

Abstract
Rationale
Compulsive behavior, which is a hallmark of psychiatric disorders such as addiction and obsessive-compulsive disorder, engages corticostriatal circuits. Previous studies indicate a role for corticostriatal N-methyl-D-aspartate receptors (NMDARs) in mediating compulsive-like responding for drugs of abuse, but the specific receptor subunits controlling reward-seeking in the face of punishment remain unclear.
Objectives
The current study assessed the involvement of corticostriatal GluN2B-containing NMDARs in measures of persistent and punished food reward-seeking.
Methods
Mice with genetic deletion of GluN2B in one of three distinct neuronal populations, cortical principal neurons, forebrain interneurons, or striatal medium spiny neurons, were tested for (1) sustained food reward-seeking when reward was absent, (2) reward-seeking under a progressive ratio schedule of reinforcement, and (3) persistent reward-seeking after a footshock punishment.
Results
Mutant mice with genetic deletion of GluN2B in cortical principal neurons demonstrated attenuated suppression of reward-seeking during punishment. These mice performed normally on other behavioral measures, including an assay for pain sensitivity. Mutants with interneuronal or striatal GluN2B deletions were normal on all behavioral assays.
Conclusions
Current findings offer novel evidence that loss of GluN2B-containing NMDARs expressed on principal neurons in the cortex results in reduced punished food rewardseeking. These data support the involvement of GluN2B subunit in cortical circuits regulating cognitive flexibility in a variety of settings, with implications for understanding the basis of inflexible behavior in neuropsychiatric disorders including obsessive-compulsive disorders (OCD) and addictions.
Keywords: Addiction, NMDA, Prefrontal cortex, Motivation, Striatum, Interneuron, Alcohol, Drug
Introduction
Compulsive behaviors and thoughts are characteristic of obsessive-compulsive disorders (OCD) and addictions. Individuals with these afflictions engage in repetitive and uncontrollable actions, often despite a strong desire to stop. The preclinical literature has demonstrated a role for prefrontal cortical and striatal circuits in mediating behavioral flexibility and habit learning, respectively (Balleine et al. 2007; Dalley et al. 2004; Everitt et al. 2008; Hamilton and Brigman 2015). Additionally, studies of drug-dependent patients and individuals with OCD have shown neural activation in the prefrontal cortex and basal ganglia, including the dorsal and ventral regions of the striatum, in response to stimuli that elicit compulsive actions (Breiter et al. 1996; Engelmann et al. 2012; Maltby et al. 2005; Volkow et al. 2005; Vollstädt-Klein et al. 2010). Changes in the functioning of these neural circuits are thus implicated in compulsive behavior.
N-methyl-D-aspartate receptors (NMDARs) are comprised of an obligatory GluN1 and multiple GluN2(A-D) subunits and have been strongly implicated in rewarded behavior (Cull-Candy et al. 2001). GluN2B-containing NMDARs have slower channel kinetics, and overexpressing or deleting this subunit alters synaptic plasticity (Brigman et al. 2010; Shipton and Paulsen 2014; von Engelhardt et al. 2008). In addition to having a major role in hippocampal memory (Brigman et al. 2010; Clayton et al. 2002; von Engelhardt et al. 2008), GluN2B receptors have been shown to contribute to working memory (Davies et al. 2013; Wang et al. 2013), memory reconsolidation (Milton et al. 2013), reversal learning (Dalton et al. 2011; Duffy et al. 2008; Thompson et al. 2015), Pavlovian fear and extinction (Dalton et al. 2008; Dalton et al. 2012; Mathur et al. 2009; Otis et al. 2014; Burgos-Robles et al. 2007), and the behavioral effects of ethanol (Badanich et al. 2011). The GluN2B subunit has also been found to mediate the effects of stress on memory (Howland and Cazakoff 2010; Wong et al. 2007), stress responsivity (Kiselycznyk et al. 2011), and depression-related behavior (Kiselycznyk et al. 2015).
The NMDAR is a binding site for ethanol (EtOH) and has been implicated in the physiological and behavioral effects of alcohol (Holmes et al. 2013; Lovinger et al. 1989; Nagy 2004; Woodward 2000). GluN2B is involved in determining the sensitivity of the NMDAR to alcohol (Woodward 2000), and administration of GluN2B antagonists can reduce alcohol drinking during withdrawal (Vengeliene et al. 2005). NMDARs in the striatum have also been linked to compulsive alcohol-seeking. For instance, systemic or intra-nucleus accumbens (NAc) core administration of the partial NMDAR-agonist D-serine blocks the effects of intermittent alcohol exposure on alcohol drinking following addition of the bitter tastant quinine (Seif et al. 2013; Seif et al. 2015; Wang et al. 2007). Collectively, these prior findings suggest a possible role for GluN2B in mediating behavioral responses for reward in the face of adverse consequences that may be at the core of compulsive-like drug-seeking and taking. However, this hypothesized role of GluN2B remains to be tested.
The goal of the current study was to assess the involvement of corticostriatal GluN2B-containing NMDARs on a series of tests for persistent and punished food reward-seeking. Based on procedures previously employed to test responding for cocaine (Deroche-Gamonet et al. 2004), we first established methods in mice that measure sustained reward-seeking when food reward is absent, continued reward-seeking under a progressive ratio (PR) schedule of reinforcement, and persistent reward-seeking after footshock-based punishment. Using these methods, we examined the behavioral consequences of genetically deleted GluN2B in one of three distinct neuronal populations, cortical principal neurons, forebrain interneurons, or striatal medium spiny neurons
Materials and methods
Subjects
Subjects were adult male mice housed in a temperature- and humidity-controlled vivarium under a 12-h light/dark cycle (lights on 0600 h). GluN2B-floxed mice were generated as previously described and backcrossed repeatedly to C57BL/ 6J to produce a C57BL/6J congenic background (Brigman et al. 2010). Three separate Cre-driver transgenic lines were used to delete GluN2B in specific brain regions and neuronal subtypes when crossed with the GluN2B-floxed mice: CaMKIIα-Cre (Tsien et al. 1996) to delete GluN2B on cortical and CA1 hippocampal principal neurons (GluN2BCxNULL) (Brigman et al. 2010), Ppp1r2-Cre (Belforte et al. 2010) to delete GluN2B on forebrain interneurons (predominantly parvalbumin-positive) (GluN2BInterNULL) (Kiselycznyk et al. 2015), and Rgs9-Cre (Dang et al. 2006) to delete GluN2B on striatal neurons (GluN2BStNULL) (Brigman et al. 2013). For each of the three mutant lines, homozygous GluN2B-floxed/Cre-negative sires were bred with homozygous GluN2B-floxed/Cre-positive dams to produce groups of male GluN2B-deficient mice and control littermates for testing. GluN2BCxNULL mice were approximately 13 weeks, GluN2BInterNULL were approximately 21 weeks, and GluN2BStNULL mice were approximately 18 weeks of age at the start of training.
Test-naïve mice were used in each experiment, and the experimenter remained blind to genotype during testing. All procedures were approved by the NIAAA Animal Care and Use Committee and followed the NIH guidelines outlined in“Using Animals in Intramural Research.” The number of mice tested is indicated in the figure legends.
Behavioral training and testing
Training and testing were conducted in a chamber measuring 21.6×17.8×12.7 cm (model ENV-307W, Med Associates, St. Albans, VT, USA) housed in a sound- and light-attenuating box. On one end of the chamber was a receptacle for delivery of food pellets, a house light, a tone generator, and two levers (left=active, right=inactive). Mice were food-restricted and maintained at 80–85% free-feeding body weight to motivate performance.
Mice were first trained to press the active lever for a 14-mg food pellet (F05684, BioServ, Frenchtown, NJ, USA) on a fixed ratio (FR) 1 (for two daily sessions) schedule of reinforcement and then progressed to a FR3 schedule (Fig. 1). Training sessions were 60 min in duration and blocked into two sequential 20-min reward-available and 10-min reward-unavailable periods (block 1: 20-min available, 10-min unavailable; block 2: 20-min available, 10-min unavailable). Reward-available periods were signaled by continuous illumination of the reward receptacle. Following each reward delivery, a 2-s, 65-dB tone was played. Responses on the inactive lever had no programmed consequences. Training continued until a criterion of reliable responding on the active lever (<20% variability on three successive sessions). The final three sessions of training were averaged as the test for responding for unavailable reward.
Fig. 1.
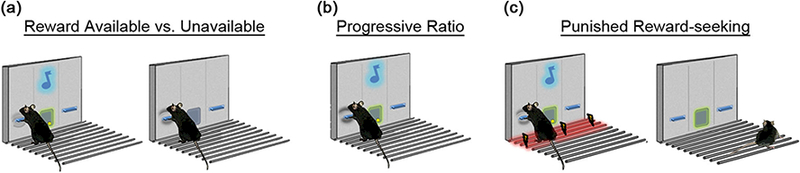
Schematic and cartoon of behavioral procedures. Mice were trained to lever-press for a 14-mg food pellet on a FR3 schedule of reinforcement then a tested for rewarded lever-pressing during “reward-available” and “reward-unavailable” periods during the final three sessions of training. b The next day, motivation for reward was assayed from lever-pressing breakpoints on a progressive ratio (PR) shedule. Responding on an FR3 schedule was reestablished on the next session. c The following day, punished suppression of reward seeking was tested with a 0.4-mA footshock
Progressive ratio responding for reward
On the next session, mice were tested on a PR schedule in which a progressively higher number of presses (range = 1–737) were required to obtain reward to measure motivation for reward. The session terminated when the mouse failed to earn a reward in a 60-min period and the breakpoint was taken as the highest number of presses made for a reward. Responding on the FR3 training schedule of reinforcement (procedure the same as training) was reestablished on the following session.
Responding for a punished reward
Punished suppression of lever pressing was assessed during one 40-min session on a FR3 schedule of reinforcement. The first active lever press in a sequence signaled a pending punishment by illuminating the house light, the second press produced a footshock (0.4 mA, 0.5 s) via a metal grid connected to a shock generator (model ENV-416S, Med Associates, St. Albans, VT, USA), and the third press resulted in the delivery of the reward and extinguished the house light. The signal-shock-reward sequence was then repeated. The reward receptacle was illuminated throughout the session to indicate reward availability. Active lever pressing during the punishment session was compared to active lever presses during the equivalent cumulative (40-min) period of reward availability during the “responding for unavailable reward” test described above. This is shown as “baseline” in the punished session data. For a cartoon depiction of the training and testing procedure, see Fig. 1.
Hot plate test for nociception
To exclude the possibility that abnormal nociception confounded the reduced sensitivity to punishment evident in GluN2BCxNULL mice (see “Results”), these mice were tested on the hot plate test, as previously described (Feyder et al. 2007). The mouse was placed on a flat plate (Columbus Instruments, Columbus, OH, USA) heated to 55 °C. The latency to show a hindpaw shake or lick was manually timed, with a maximum response latency of 30 s before the mouse was removed to avoid tissue damage.
Statistical analysis
Data were analyzed using t tests or analysis of variance (ANOVA), followed by Sidak’s or Dunnett’s post hoc tests for multiple comparisons, where appropriate, using GraphPad Prism 6.04 (GraphPad Software Inc, La Jolla, CA, USA). The threshold for statistical significance was set at P <.05.
Results
Deletion of GluN2B on cortical principal neurons reduced punished suppression of reward-seeking
Mutant mice with deletion of GluN2B in principal neurons (GluN2BCxNULL) (Fig. 2a) reached the FR3 schedule training criterion in a similar number of sessions as GluN2BFLOX controls (Fig. 2b). During training, GluN2BCxNULL mice pressed the active lever more than controls (main effect of genotype: F1,52=5.53, P <.05; main effect of lever: F1,52 = 220.80, P<.0001; genotype × lever interaction: F1,52=6.37, P<.05; post hoc tests: P<.01 for the active lever) (Table 1). Regardless of genotype, there were generally very few presses on the inactive lever during all three phases of testing (Table 2).
Fig. 2.
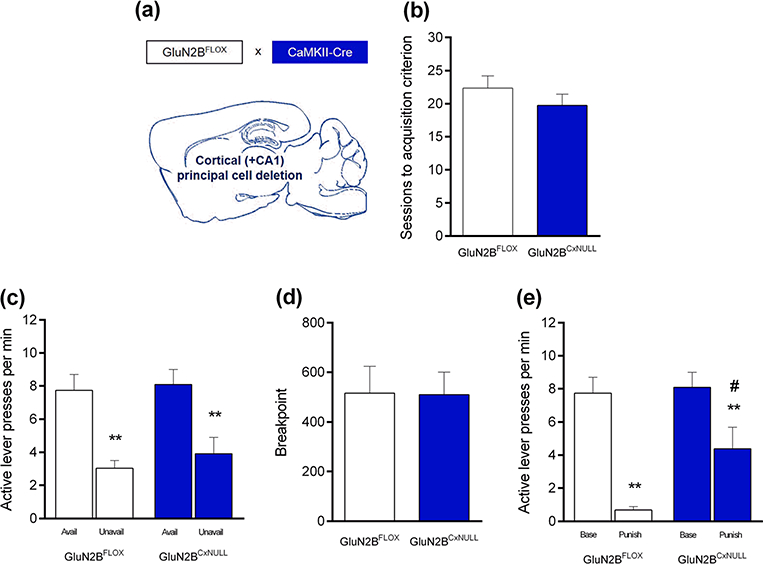
Deletion of GluN2B on cortical principal neurons attenuated punished suppression of reward-seeking. a GluN2B was deleted in cortical principal neurons by crossing GluN2B-floxed mice with CaMKIIα-Cre mice. b Genotypes reached the training criterion in a similar number of sessions. c Both genotypes pressed the active lever more often during signaled reward-available than reward-unavailable phases. d Breakpoints attained on a progressive ratio (PR) schedule were similar between genotypes. e Punishment suppressed responding in both genotypes, but did so to a lesser extent in GluN2BCxNULL than GluN2BFLOX controls. **P<.01 versus available phases or unpunished baseline, #P<.05 GluN2BCxNULL versus GluN2BFLOX. n = 13–15 per genotype. Data are means ±SEM
Table 1.
Responses during training. Responses on the active and inactive levers during FR3 training
| GluN2BFLOX | GluN2BCxNULL | GluN2BFLOX | GluN2BInterNULL | GluN2BFLOX | GluN2BStNULL | |
|---|---|---|---|---|---|---|
| Active lever | 4.59±0.57 | 6.29±0.41** | 3.70±0.38 | 5.00±0.61* | 4.76±0.61 | 5.07±0.63 |
| Inactive lever | 0.29±0.09 | 0.23±0.06 | 0.48±0.11 | 0.38±0.06 | 0.32±0.08 | 0.14±0.02 |
Data are mean presses per minute±SEM
*P<.05
**P<.01 compared to GluN2BFLOX control
Table 2.
Responding on the inactive lever during testing. Responses on the inactive lever were low across genotypes and the various phases of testing
| Test phase | GluN2BFLOX | GluN2BCxNULL | GluN2BFLOX | GluN2BInterNULL | GluN2BFLOX | GluN2BStNULL |
|---|---|---|---|---|---|---|
| Reward-unavailable (presses/min) | 0.19±0.04 | 0.14±0.03 | 0.30±0.05 | 0.39±0.05 | 0.30±0.06 | 0.13±0.04 |
| Progressive ratio (total presses) | 95.50±28.19 | 151.00±70.66 | 56.11±15.20 | 78.50±15.67 | 116.91±27.90 | 78.17±15.40 |
| Punished (presses/min) | 0.64±0.26 | 0.63±0.22 | 0.26±0.10 | 0.23±0.04 | 0.31±0.10 | 0.20±0.08 |
Data are means±SEM
Responding for unavailable reward
Both genotypes pressed the active lever significantly more often during reward-available than reward-unavailable periods (genotype × availability phase interaction: P>.05; main effect of genotype: P>.05; main effect of availability phase: F1,26=26.19, P<.01; post hoc tests: P<.01 in both genotypes) (Fig. 2c).
Progressive ratio responding for reward
The PR breakpoint attained was not different between genotypes (t test, P>.05) (Fig. 2d).
Responding for a punished reward
Punishment significantly suppressed lever pressing in both genotypes (genotype × punishment interaction: P>.05; main effect of genotype: P>.05; main effect of punishment: F1,26 = 46.79, P<.01). However, the decrease in responding during punishment was significantly lesser in GluN2BCxNULL mutants than GluN2BFLOX controls (t26=2.21, P<.05) (Fig. 2e).
Deletion of GluN2B on forebrain interneurons did not affect behavioral measures
Mutant mice with deletion of GluN2B in cortical GABAergic neurons (GluN2BInterNULL) (Fig. 3a) acquired the training criterion in a similar number of sessions as GluN2BFLOX controls (Fig. 3b). During training, GluN2BInterNULL mice pressed the active lever more than controls (main effect of genotype: P>.05; genotype × lever interaction: P>.05; main effect of lever: F1,40=105.30, P<.0001; post hoc tests: P<.05 for the active lever) (Table 1). Regardless of genotype, there were generally very few presses on the inactive lever during testing (Table 2).
Fig. 3.
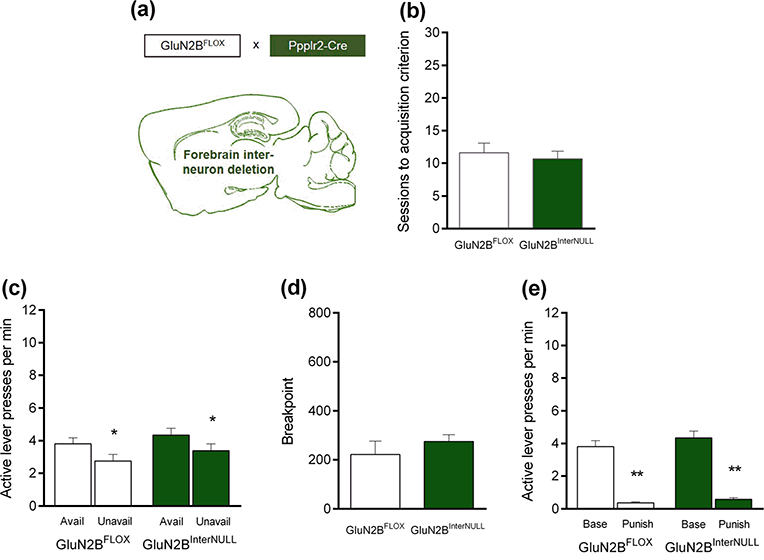
Deletion of GluN2B on forebrain interneurons did not affect measures of compulsive-like behavior a GluN2B was deleted in cortical interneurons by crossing GluN2B-floxed mice with Ppp1r2-Cre mice. b Genotypes reached the training criterion in a similar number of sessions. c Both genotypes pressed the active lever more often during signaled reward-available than reward-unavailable phases. d Breakpoints attained on a progressive ratio (PR) schedule were similar between genotypes. e Punishment suppressed responding in both genotypes. *P<.05, **P<.01 versus available phases or unpunished baseline. n=10–12 per genotype. Data are means ±SEM
Responding for unavailable reward
Both genotypes pressed the active lever significantly more often during reward-available than reward-unavailable periods (genotype × availability phase interaction: P>.05; main effect of genotype: P>.05; main effect of availability phase: F1,20=16.28, P<.01; post hoc tests: P<.05 in both genotypes) (Fig. 3c).
Progressive ratio responding for reward
The PR breakpoint attained by the two genotypes was similar (t test: P>.05) (Fig. 3d).
Responding for a punished reward
There was a significant and similar suppression of active lever responding during punishment in both genotypes (genotype × punishment interaction: P>.05; main effect of genotype: P>.05; main effect of punishment: F1j19=178.0, P<.01: post hoc tests: P<.01 in both genotypes) (Fig. 3e).
Deletion of GluN2B on striatal neurons did not affect behavioral measures
Mutant mice with deletion of GluN2B in striatal neurons (GluN2BStNULL) (Fig. 4a) attained training criterion in a similar number of sessions as GluN2BFLOX controls (Fig. 4b). There were no differences between genotypes in active or inactive lever presses during training (Table 1). Irrespective of genotype, there were generally very few presses on the inactive lever during testing (Table 2).
Fig. 4.
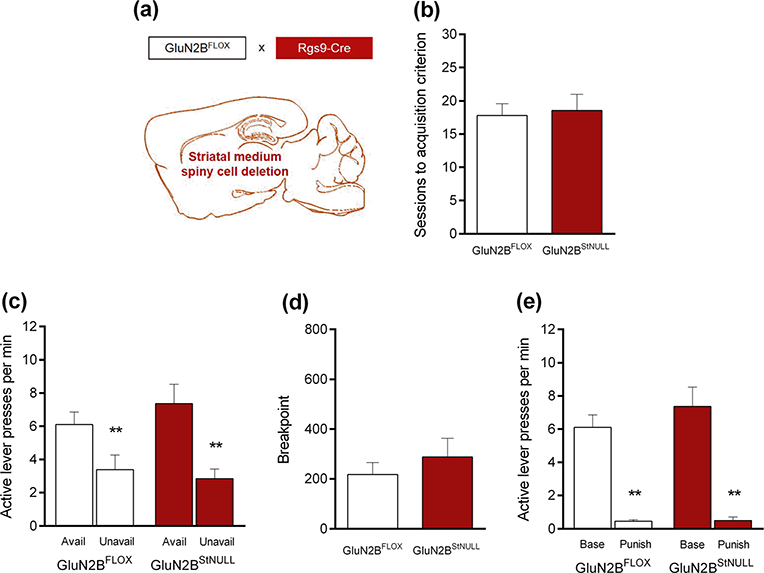
Deletion of GluN2B on striatal neurons did not affect measures of compulsive-like behavior. a GluN2B was deleted in striatal neurons by crossing GluN2B-floxed mice with Rgs9- Cre mice. b Genotypes reached the training criterion in a similar number of sessions. c Both genotypes pressed the active lever more often during signaled reward-available than reward-unavailable phases. d Breakpoints attained on a progressive ratio (PR) schedule were similar jbetween genotypes. e Punishment suppressed responding in both genotypes. **P<.01 versus available phases or unpunished baseline. n=9–11 per genotype. Data are means±SEM
Responding for unavailable reward
Both genotypes pressed the active lever significantly more times during reward-available than reward-unavailable periods (genotype × availability phase interaction: P>.05; main effect of genotype: P>.05; main effect of availability phase: F1,18 = 33.48, P<.01; post hoc tests: P<.01 in both genotypes) (Fig. 4c).
Progressive ratio responding for reward
Genotypes reached a similar breakpoint on the PR schedule (t test: P>.05) (Fig. 4d).
Responding for a punished reward
Punishment significantly suppressed responding in both genotypes (genotype × punishment interaction: P>.05; main effect of genotype: P>.05; main effect of punishment: F1,17=81.01, P<.01; post hoc tests: P<.001 in both genotypes) (Fig. 4e).
Deletion of GluN2B on cortical principal neurons did not affect hot plate nociception
There were no differences in hot plate response latencies between GluN2BCxNULL mutants and GluN2BFLOX controls (Table 3).
Table 3.
Deletion of GluN2B on cortical principal neurons did not affect pain perception on the hot plate test. Response latencies did not differ between GluN2BFLOX and GluN2BCxNULL mice
| GluN2BFLOX | GluN2BCxNULL | |
|---|---|---|
| Response latency (s) | 8.42±1.42 | 6.57±0.92 |
Data are means±SEM
Discussion
The current study used region- and cell-specific deletion of GluN2B to probe the contribution of this NMDAR subunit to the control of reward-seeking behaviors. The major finding was that mice lacking GluN2B on principal neurons throughout the cortex, as well as the dorsal CA1 hippocampal subregion, showed attenuated suppression of responding for a food reward when the response was punished. By contrast, mutation-induced loss of GluN2B on forebrain interneurons or striatal medium spiny neurons failed to produce alterations in punished responding.
The measure of punished reward-seeking employed here and in previous studies has similarities to Pavlovian fear conditioning paradigms, as well as to long-standing punishment-based assays for anxiety-like behavior, such as the Vogel conflict and Geller-Seifter tests (Cryan and Holmes 2005; Griebel and Holmes 2013). However, it is unlikely that the reduced sensitivity to punishment in GluN2BCxNULL mice reflects impaired learning or reduced anxiety-like behavior, as we have previously reported that these mutant mice associate an auditory cue with footshock at a similar level to non-mutant controls in a delay Pavlovian conditioning paradigm (Brigman et al. 2010) and exhibit normal unconditioned anxiety-like behaviors (Kiselycznyk et al. 2011). Here, we also showed that the mutants exhibit normal nociceptive responses in the hot plate test, discounting the possibility that impaired pain perception accounted for the attenuated punishment suppression of reward-seeking. Furthermore, arguing against general learning deficits, abnormal punished reward-seeking in these mice were not accompanied by deficits in initial operant learning or in persistent reward-seeking during reward absence or progressive ratio responding for reward ( Deroche-Gamonet et al. 2004; Radwanska and Kaczmarek 2012). In this context, recent studies in rats have found that chronic exposure to ethanol or cocaine causes changes in two or all three of these measures (Belin et al. 2011; Deroche-Gamonet et al. 2004; Jonkman et al. 2012; Vendruscolo et al. 2012) (but see Waters et al. 2014). The behaviorally selective (punishment-specific) phenotypic profile shown by the GluN2BCxNULL mice demonstrates that while the mechanisms regulating these various measures are likely to be overlapping, they can also be dissociated. Of note in this context, we have found that C57BL/6J mice chronically exposed to ethanol also show a selective deficit in punished suppression of ethanol-seeking and altered cortical expression of NMDARs (Radke et al., unpublished observations).
A contribution of GluN2B-containing NMDARs to punished reward-seeking adds to the growing evidence of an important role for this subunit in a variety of behavioral settings which require the inhibition of a previously learned response. For example, systemic administration of the GluN2B selective antagonist, Ro 25–6981, has been shown to impair reversal learning (Dalton et al. 2011; Duffy et al. 2008), strategy set-shifting (Dalton et al. 2011), working memory (Davies et al. 2013), and fear extinction (Dalton et al. 2008; Dalton et al. 2012) in rats. The neural locus of these effects is likely to be cortical, given reversal learning is impaired in GluN2BCxNULL mice (Brigman et al. 2013; Thompson et al. 2015) and by microinfusion of Ro 25–6981 into the lateral orbitofrontal cortex (Brigman et al. 2013), and that GluN2B antagonist microinfusion into the medial PFC impairs working memory (Davies et al. 2013) and extinction of a cocaine-conditioned place preference (Otis et al. 2014).
A cortical basis for GluN2B regulation of punished reward-seeking would also be consistent with the broader literature implicating PFC dysfunction in loss of behavioral control (Cannella et al. 2013; Chen et al. 2013; Everitt et al. 2008; Jentsch and Taylor 1999; Schoenbaum and Shaham 2008; Seif et al. 2013). In this context, we did not find an effect on this same behavioral measure in mutants with either forebrain-interneuron or striatal GluN2B deletion. Performance on the progressive ratio and reward-unavailability tests was also normal in the GluN2BInterNULL and GluN2BStNULL mice (as it was in the GluN2BCxNULL mice). These negative phenotypes were present, even though the overall vigor of responding was noticeably lower in the GluN2BInterNULL and GluN2BStNULL mutants than was evident in the GluN2BCxNULL mice, likely due to subtle differences in age, genetic background, or extraneous factors. Notwithstanding, on the basis of our current findings alone, it would be premature to fully exclude a contribution of GluN2B-containing NMDARs localized to forebrain interneurons or striatum medium spiny neurons to effects of punishment on reward-seeking. Indeed, prior studies have implicated the nucleus accumbens (Seif et al. 2013; Seif et al. 2015), basolateral amygdala (Pelloux et al. 2013), and a lateral hypothalamus-ventral tegmental area pathway (Nieh et al. 2015) in compulsive-like responding for alcohol, cocaine, or sucrose rewards. Further delineating the critical neural circuits mediating compulsive-like behaviors, and the functional involvement of GluN2B within these circuits, will be a key area for future work.
In summary, the results of the current study provide novel evidence that loss of GluN2B-containing NMDARs expressed on principal neurons in the cortex, but not deletion on forebrain interneurons or striatal medium spiny neurons, significantly attenuated the ability of punishment to inhibit responding for a food reward. This phenotype was unrelated to deficits in pain perception, operant learning, or measures of persistent reward-seeking and motivation for reward. These data add to emerging evidence that the GluN2B subunit has a critical role in cortical circuits that regulate the ability to flexibly adapt behavior in the face of changing environmental demands. They could also have implications for elucidating the pathophysiological basis of maladaptive compulsions in neuropsychiatric disorders including OCD and addictions.
Acknowledgments
We are grateful to Munisa Bachu, Shaun Flynn, and Adrina Kocharian for technical assistance and to Dr. Jonathan Brigman for the cartoons of the behavioral procedures. This research was supported by the NIAAA Intramural Research Program and NIMH grant K22MH099164.
References
- Badanich KA, Doremus-Fitzwater TL, Mulholland PJ, Randall PK, Delpire E, Becker HC (2011) NR2B-deficient mice are more sensitive to the locomotor stimulant and depressant effects of ethanol. Genes Brain Behav 10:805–816 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Balleine BW, Delgado MR, Hikosaka O (2007) The role of the dorsal striatum in reward and decision-making. J Neurosci 27:8161–8165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belforte JE, Zsiros V, Sklar ER, Jiang Z, Yu G, Li Y, Quinlan EM, Nakazawa K (2010) Postnatal NMDA receptor ablation in corticolimbic interneurons confers schizophrenia-like phenotypes. Nat Neurosci 13:76–83 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belin D, Berson N, Balado E, Piazza PV, Deroche-Gamonet V (2011) High-novelty-preference rats are predisposed to compulsive cocaine self-administration. Neuropsychopharmacology 36:569–579 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breiter HC, Rauch SL, Kwong KK, Baker JR, Weisskoff RM, Kennedy DN, Kendrick AD, Davis TL, Jiang A, Cohen MS (1996) Functional magnetic resonance imaging of symptom provocation in obsessive-compulsive disorder. Arch Gen Psychiatry 53:595–606 [DOI] [PubMed] [Google Scholar]
- Brigman JL, Daut RA, Wright T, Gunduz-Cinar O, Graybeal C, Davis MI, Jiang Z, Saksida LM, Jinde S, Pease M (2013) GluN2B in corticostriatal circuits governs choice learning and choice shifting. Nat Neurosci 16:1101–1110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brigman JL, Wright T, Talani G, Prasad-Mulcare S, Jinde S, Seabold GK, Mathur P, Davis MI, Bock R, Gustin RM (2010) Loss of GluN2B-containing NMDA receptors in CA1 hippocampus and cortex impairs long-term depression, reduces dendritic spine density, and disrupts learning. J Neurosci 30:4590–4600 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgos-Robles A, Vidal-Gonzalez I, Santini E, Quirk GJ (2007) Consolidation of fear extinction requires NMDA receptor-dependent bursting in the ventromedial prefrontal cortex. Neuron 53(6):871–880 [DOI] [PubMed] [Google Scholar]
- Cannella N, Halbout B, Uhrig S, Evrard L, Corsi M, Corti C, Deroche- Gamonet V, Hansson AC, Spanagel R (2013) The mGluR2/3 agonist LY379268 induced anti-reinstatement effects in rats exhibiting addiction-like behavior. Neuropsychopharmacology 38:2048–2056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen BT, Yau H-J, Hatch C, Kusumoto-Yoshida I, Cho SL, Hopf FW, Bonci A (2013) Rescuing cocaine-induced prefrontal cortex hypoactivity prevents compulsive cocaine seeking. Nature 496: 359–362 [DOI] [PubMed] [Google Scholar]
- Clayton DA, Mesches MH, Alvarez E, Bickford PC, Browning MD (2002) A hippocampal NR2B deficit can mimic age-related changes in long-term potentiation and spatial learning in the Fischer 344 rat. J Neurosci 22:3628–3637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cryan JF, Holmes A (2005) The ascent of mouse: advances in modelling human depression and anxiety. Nat Rev Drug Discov 4:775–790 [DOI] [PubMed] [Google Scholar]
- Cull-Candy S, Brickley S, Farrant M (2001) NMDA receptor subunits: diversity, development and disease. Curr Opin Neurobiol 11:327–335 [DOI] [PubMed] [Google Scholar]
- Dalley JW, Cardinal RN, Robbins TW (2004) Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neurosci Biobehav Rev 28:771–784 [DOI] [PubMed] [Google Scholar]
- Dalton GL, Ma LM, Phillips AG, Floresco SB (2011) Blockade of NMDA GluN2B receptors selectively impairs behavioral flexibility but not initial discrimination learning. Psychopharmacology 216: 525–535 [DOI] [PubMed] [Google Scholar]
- Dalton GL, Wang YT, Floresco SB, Phillips AG (2008) Disruption of AMPA receptor endocytosis impairs the extinction, but not acquisition of learned fear. Neuropsychopharmacology 33:2416–2426 [DOI] [PubMed] [Google Scholar]
- Dalton GL, Wu DC, Wang YT, Floresco SB, Phillips AG (2012) NMDA GluN2A and GluN2B receptors play separate roles in the induction of LTP and LTD in the amygdala and in the acquisition and extinction of conditioned fear. Neuropharmacology 62:797–806 [DOI] [PubMed] [Google Scholar]
- Dang MT, Yokoi F, Yin HH, Lovinger DM, Wang Y, Li Y (2006) Disrupted motor learning and long-term synaptic plasticity in mice lacking NMDAR1 in the striatum. Proc Natl Acad Sci 103:15254–15259 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davies DA, Greba Q, Howland JG (2013) GluN2B-containing NMDA receptors and AMPA receptors in medial prefrontal cortex are necessary for odor span in rats. Front Behav Neurosci 7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deroche-Gamonet V, Belin D, Piazza PV (2004) Evidence for addiction-like behavior in the rat. Science 305:1014–1017 [DOI] [PubMed] [Google Scholar]
- Duffy S, Labrie V, Roder JC (2008) D-serine augments NMDA-NR2B receptor-dependent hippocampal long-term depression and spatial reversal learning. Neuropsychopharmacology 33:1004–1018 [DOI] [PubMed] [Google Scholar]
- Engelmann JM, Versace F, Robinson JD, Minnix JA, Lam CY, Cui Y, Brown VL, Cinciripini PM (2012) Neural substrates of smoking cue reactivity: a meta-analysis of fMRI studies. Neuroimage 60:252–262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Everitt BJ, Belin D, Economidou D, Pelloux Y, Dalley JW, Robbins TW (2008) Neural mechanisms underlying the vulnerability to develop compulsive drug-seeking habits and addiction. Phil Trans R Soc B: Biol Sci 363:3125–3135 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feyder M, Wiedholz L, Sprengel R, Holmes A (2007) Impaired associative fear learning in mice with complete loss or haploinsufficiency of AMPA GluR1 receptors. Front Behav Neurosci 1:4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griebel G, Holmes A (2013) 50 years of hurdles and hope in anxiolytic drug discovery. Nat Rev Drug Discov 12:667–687 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamilton DA, Brigman JL (2015) Behavioral flexibility in rats and mice: contributions of distinct frontocortical regions. Genes Brain Behav [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holmes A, Spanagel R, Krystal JH (2013) Glutamatergic targets for new alcohol medications. Psychopharmacology 229:539–554 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Howland JG, Cazakoff BN (2010) Effects of acute stress and GluN2B-containing NMDA receptor antagonism on object and object-place recognition memory. Neurobiol Learn Mem 93:261–267 [DOI] [PubMed] [Google Scholar]
- Jentsch JD, Taylor JR (1999) Impulsivity resulting from frontostriatal dysfunction in drug abuse: implications for the control of behavior by reward-related stimuli. Psychopharmacology 146:373–390 [DOI] [PubMed] [Google Scholar]
- Jonkman S, Pelloux Y, Everitt BJ (2012) Differential roles of the dorsolateral and midlateral striatum in punished cocaine seeking. J Neurosci 32:4645–4650 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiselycznyk C, Jury NJ, Halladay LR, Nakazawa K, Mishina M, Sprengel R, Grant SG, Svenningsson P, Holmes A (2015) NMDA receptor subunits and associated signaling molecules mediating antidepressant-related effects of NMDA-GluN2B antagonism. Behav Brain Res 287:89–95 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiselycznyk C, Svenningsson P, Delpire E, Holmes A (2011) Genetic, pharmacological and lesion analyses reveal a selective role for corticohippocampal GLUN2B in a novel repeated swim stress paradigm. Neuroscience 193:259–268 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovinger DM, White G, Weight FF (1989) Ethanol inhibits NMDA-activated ion current in hippocampal neurons. Science 243:1721–1724 [DOI] [PubMed] [Google Scholar]
- Maltby N, Tolin DF, Worhunsky P, O’Keefe TM, Kiehl KA (2005) Dysfunctional action monitoring hyperactivates frontal-striatal circuits in obsessive-compulsive disorder: an event-related fMRI study. Neuroimage 24:495–503 [DOI] [PubMed] [Google Scholar]
- Mathur P, Graybeal C, Feyder M, Davis MI, Holmes A (2009) Fear memory impairing effects of systemic treatment with the NMDA NR2B subunit antagonist, Ro 25–6981, in mice: attenuation with ageing. Pharmacol Biochem Behav 91:453–460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Milton AL, Merlo E, Ratano P, Gregory BL, Dumbreck JK, Everitt BJ (2013) Double dissociation of the requirement for GluN2B-and GluN2A-containing NMDA receptors in the destabilization and restabilization of a reconsolidating memory. J Neurosci 33:1109–1115 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagy J (2004) The NR2B subtype of NMDA receptor: a potential target for the treatment of alcohol dependence. Curr Drug Targets CNS Neurol Disord 3:169–179 [DOI] [PubMed] [Google Scholar]
- Nieh EH, Matthews GA, Allsop SA, Presbrey KN, Leppla CA, Wichmann R, Neve R, Wildes CP, Tye KM (2015) Decoding neural circuits that control compulsive sucrose seeking. Cell 160:528–541 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otis JM, Fitzgerald MK, Mueller D (2014) Infralimbic BDNF/TrkB enhancement of GluN2B currents facilitates extinction of a cocaine-conditioned place preference. J Neurosci 34:6057–6064 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pelloux Y, Murray JE, Everitt BJ (2013) Differential roles of the prefrontal cortical subregions and basolateral amygdala in compulsive cocaine seeking and relapse after voluntary abstinence in rats. Eur J Neurosci [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radwanska K, Kaczmarek L (2012) Characterization of an alcohol addiction-prone phenotype in mice. Addict Biol 17:601–612 [DOI] [PubMed] [Google Scholar]
- Schoenbaum G, Shaham Y (2008) The role of orbitofrontal cortex in drug addiction: a review of preclinical studies. Biol Psychiatry 63:256–262 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seif T, Chang S-J, Simms JA, Gibb SL, Dadgar J, Chen BT, Harvey BK, Ron D, Messing RO, Bonci A (2013) Cortical activation of accum-bens hyperpolarization-active NMDARs mediates aversion-resistant alcohol intake. Nat Neurosci 16:1094–1100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seif T, Simms JA, Lei K, Wegner S, Bonci A, Messing RO, Hopf FW (2015) D-serine and D-cycloserine reduce compulsive alcohol intake in rats. Neuropsychopharmacology [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shipton OA, Paulsen O (2014) GluN2A and GluN2B subunit-containing NMDA receptors in hippocampal plasticity. Phil Trans R Soc London B: Biol Sci 369:20130163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thompson SM, Josey M, Holmes A, Brigman JL (2015) Conditional loss of GluN2B in cortex and hippocampus impairs attentional set fomation. Behav Neurosci 129:105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsien JZ, Chen DF, Gerber D, Tom C, Mercer EH, Anderson DJ, Mayford M, Kandel ER, Tonegawa S (1996) Subregion-and cell type-restricted gene knockout in mouse brain. Cell 87:1317–1326 [DOI] [PubMed] [Google Scholar]
- Vendruscolo LF, Barbier E, Schlosburg JE, Misra KK, Whitfield TW, Logrip ML, Rivier C, Repunte-Canonigo V, Zorrilla EP, Sanna PP (2012) Corticosteroid-dependent plasticity mediates compulsive alcohol drinking in rats. J Neurosci 32:7563–7571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vengeliene V, Bachteler D, Danysz W, Spanagel R (2005) The role of the NMDA receptor in alcohol relapse: a pharmacological mapping study using the alcohol deprivation effect. Neuropharmacology 48:822–829 [DOI] [PubMed] [Google Scholar]
- Volkow ND, Wang G-J, Ma Y, Fowler JS, Wong C, Ding Y-S, Hitzemann R, Swanson JM, Kalivas P (2005) Activation of orbital and medial prefrontal cortex by methylphenidate in cocaine-addicted subjects but not in controls: relevance to addiction. J Neurosci 25:3932–3939 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollstädt-Klein S, Wichert S, Rabinstein J, Bühler M, Klein O, Ende G, Hermann D, Mann K (2010) Initial, habitual and compulsive alcohol use is characterized by a shift of cue processing from ventral to dorsal striatum. Addiction 105:1741–1749 [DOI] [PubMed] [Google Scholar]
- von Engelhardt J, Doganci B, Jensen V, Hvalby Ø, Göngrich C, Taylor A, Barkus C, Sanderson DJ, Rawlins JNP, Seeburg PH(2008) Contribution of hippocampal and extra-hippocampal NR2B-containing NMDA receptors to performance on spatial learning tasks. Neuron 60:846–860 [DOI] [PubMed] [Google Scholar]
- Wang J, Carnicella S, Phamluong K, Jeanblanc J, Ronesi JA, Chaudhri N, Janak PH, Lovinger DM, Ron D (2007) Ethanol induces long-term facilitation of NR2B-NMDA receptor activity in the dorsal striatum: implications for alcohol drinking behavior. J Neurosci 27:3593–3602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang M, Yang Y, Wang C-J, Gamo NJ, Jin LE, Mazer JA, Morrison JH, Wang X-J, Arnsten AF (2013) NMDA receptors subserve persistent neuronal firing during working memory in dorsolateral prefrontal cortex. Neuron 77:736–749 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Waters RP, Moorman DE, Young AB, Feltenstein MW, See RE (2014) Assessment of a proposed “three-criteria” cocaine addiction model for use in reinstatement studies with rats. Psychopharmacology (Berl) 231:3197–3205 [DOI] [PubMed] [Google Scholar]
- Wong TP, Howland JG, Robillard JM, Ge Y, Yu W, Titterness AK, Brebner K, Liu L, Weinberg J, Christie BR (2007) Hippocampal long-term depression mediates acute stress-induced spatial memory retrieval impairment. Proc Natl Acad Sci 104:11471–11476 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodward JJ (2000) Ethanol and NMDA receptor signaling. Critical Reviews™ in Neurobiology 14 [DOI] [PubMed] [Google Scholar]