

Vascular endothelial growth factor (VEGF), an angiogenic factor with neuroprotective effects: The VEGF was initially characterized by its vasculogenic and angiogenic activities and its capacity to promote vascular permeability (Yancopoulos et al., 2000). VEGF is also known as VEGF-A and is the prototype member of a related group of five trophic factors, VEGF-B, VEGF-C, VEGF-D and placental growth factor (PlGF; Lange et al., 2016). Interestingly, VEGF-B shares a high degree of homology with VEGF but, in contrast to VEGF, has low angiogenic activity and is not pro-inflammatory. VEGF-A binds to two tyrosine quinase receptors, named VEGFR-1 and VEGFR-2, and also to the coreceptor neuropilin-1. By contrast, VEGF-B only binds to the tyrosine kinase receptor VEGFR-1 and also to the coreceptor neuropilin-1 (Lange et al., 2016).
Nowadays, it is emerging that, in addition to its vascular actions, VEGF also has a significant impact on neurons by protecting them from trauma or disease. Thus, the administration of VEGF, both in vitro and in vivo through different delivery procedures, prevents or delays the death of neurons exposed to different types of insult (Lange et al., 2016). Since the knockout mice for VEGF is not viable, leading to embryonic lethality due to insufficient development of the vascular system, cardiovascular researchers designed a mutant mice with low levels of VEGF by deleting the hypoxia response element in the promoter region of the VEGF gene. Unexpectedly, these mutant mice developed an adult-onset muscle weakness accompanied by motoneuron degeneration that resembled the human disease amyotrophic lateral sclerosis (Oosthuyse et al., 2001). This work provided a novel insight to explain motoneuron degeneration in amyotrophic lateral sclerosis, since it established, for the first time, a link between low levels of VEGF and motoneuron atrophy or death. Studies of cerebrospinal fluid samples obtained from patients suffering amyotrophic lateral sclerosis, or post-mortem investigation of human spinal cord have also confirmed a correlation between low levels of VEGF and/or its receptors and amyotrophic lateral sclerosis (Sathasivam, 2008).
In this line, it was expected that the administration of VEGF in animal models of motoneuron degeneration might lead to neuroprotective effects. Several experiments indeed confirmed this possibility. When mice that overexpress VEGF (VEGF+/+) were crossed with the mutant mice G93A superoxide dismutase 1 (SOD1G93A), a classical model of amyotrophic lateral sclerosis, the double-transgenic mice showed delayed motoneuron loss and prolonged survival as compared to the SOD1G93A single-transgenic mice. Similar results were obtained when VEGF was injected intrathecally using an adeno-associated virus vector (Wang et al., 2016). Moreover, VEGF delivery with retrogradely transported lentivector to SOD1G93A mice also delayed the onset of the disease, slowed the progression of motoneuronal degeneration and increased life expectancy (Azzouz et al., 2004). Using a different model of motoneuron degeneration, it has also been shown that the chronic administration of VEGF in the spinal cord through a miniosmotic pump prevents the paralysis and motoneuronal death in rats exposed to excitotoxic motoneuron damage (Tovar-y-Romo et al., 2007). It is important to emphasize that VEGF can also exert neuroprotective roles in other neuronal types following different types of insult, such as ischemia, seizures, or neurological diseases (Lange et al., 2016).
VEGF neurophysiological effects on motoneurons: All these investigations have led to the statement that VEGF can be considered as a neurotrophic factor, especially for motoneurons, and have promising implications for the use of VEGF in the treatment of motoneuron disease. However, although it seems clear that VEGF can rescue vulnerable motoneurons from cell death, nothing is known about the physiological state in which these “rescued” motoneurons remain, for example, do they receive the same synaptic inputs or, on the contrary, do they lose afferent boutons as happens in severed motoneurons? Do these VEGF-treated motoneurons retain normal discharge characteristics or, rather, do they exhibit an altered firing? Damaged motoneurons not only need to be protected from cell death or atrophy, but should also maintain a normal synaptic complement and firing properties if VEGF treatment is expected to lead to functional recovery. We have pursued these questions recently (Calvo et al., 2018) in the chronic alert animal preparation, using the abducens motoneurons as the experimental model. It should be mentioned that although extraocular motoneurons, located in the abducens, trochlear and oculomotor nuclei, are more resistant to amyotrophic lateral sclerosis, they also exhibit some signs of degeneration and cell death at later stages of the disease.
Abducens motoneurons innervate the extraocular lateral rectus muscle. Their discharge correlates with eye movements in the horizontal plane. These motoneurons show a typical tonic-phasic firing pattern, thus having eye position and eye velocity sensitivities. Following axotomy, the discharge of abducens motoneurons shows several alterations such as an overall reduction in firing rate and a significant reduction in eye position and velocity sensitivities. Moreover, axotomy is followed by the withdrawal of synaptic boutons and a reduction in the efficacy of synaptically-driven signals (Davis-López de Carrizosa et al., 2009, 2010). Our recent study (Calvo et al., 2018) indicates that VEGF administration at the time of axotomy completely prevents the lesion-induced alterations in the discharge pattern as well as the synaptic stripping. If VEGF is administered after a delayed period of axotomy, this factor is able to recover the physiological activity of abducens motoneurons (both the tonic and the phasic components) and their synaptic inputs. To the best of our knowledge, this is the first study demonstrating that a single neurotrophic factor is able to recover lesion-induced alterations in the discharge pattern and synaptic complement of motoneurons from a multidisciplinary approach that includes physiological and morphological experiments.
Previous studies in our research group have also supplied axotomized abducens motoneurons with different neurotrophic factors, i.e., nerve growth factor (NGF), brain-derived neurotrophic factor (BDNF) and neurotrophic-3 (NT-3). Interestingly, neither of these factors yielded a complete recovery in firing and synaptic inputs after lesion. Thus, BDNF recovered only the tonic component of the discharge, whereas NT-3 recovered only the phasic firing (Davis-López de Carrizosa et al., 2009). These findings indicate that these two factors produced only a partial recovery. NGF administration yielded very peculiar results in axotomized abducens motoneurons, yielding to high eye position and velocity sensitivities and an irregular firing rate (Davis-López de Carrizosa et al., 2010) (Figure 1). Although the administration of neurotrophins (NGF, BDNF or NT-3) or other neurotrophic factors stimulates motoneuron survival in amyotrophic lateral sclerosis mouse models, the loss of these factors does not cause adult-onset motoneuron degeneration resembling the disease, so that muscle paralysis and motoneuron degeneration are not induced in mice lacking these factors (Lange et al., 2016). In contrast, VEGF deficit leads to the development of amyotrophic lateral sclerosis-like motoneuron degeneration and paralysis. Altogether, these findings strongly suggest that VEGF might be the crucial neurotrophic factor for the maintenance of a normal physiology in motoneurons.
Figure 1.
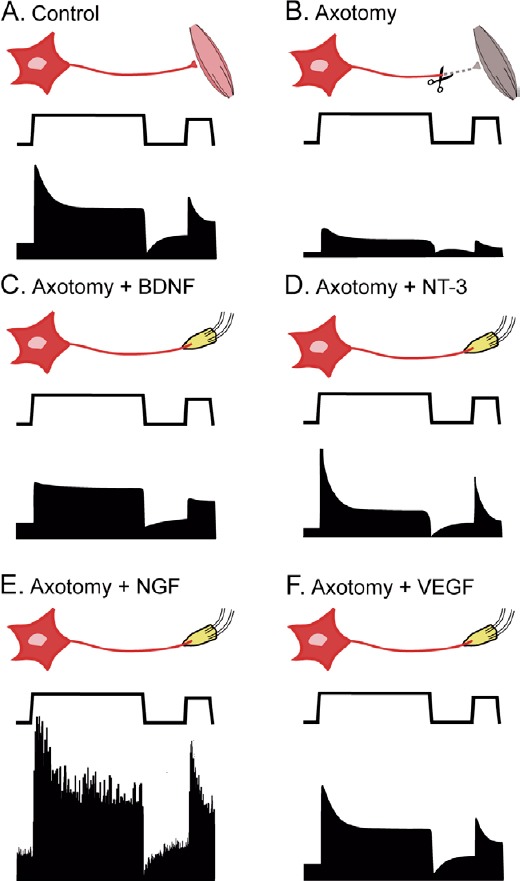
Schematic diagram comparing the results obtained after different neurotrophic factor administration on axotomized abducens motoneurons.
In each panel, from top to bottom: experimental situation, eye position, and associated firing rate. (A) Control motoneurons show a typical tonic-phasic discharge proportional to eye position and velocity. (B) Axotomy decreases the discharge, affecting both the tonic and the phasic components. (C) Brain-derived neurotrophic factor (BDNF) administration recovers essentially the tonic firing. (D) Neurotrophic-3 (NT-3) treatment restores basically the phasic activity. (E) Nerve growth factor (NGF) increases both the tonic and the phasic components as well as discharge variability over control values. (F) Vascular endothelial growth factor (VEGF) treatment completely recovers the firing pattern of axotomized motoneurons to control values.
It has been hypothesized that VEGF can mediate its trophic actions by two possible mechanisms: i) acting as an angiogenic factor, VEGF can increase the supply of oxygen and nutrients to lesioned motoneurons, and ii) as a direct neurotrophic factor for motoneurons (Oosthuyse et al., 2001; Azzouz et al., 2004). It has been demonstrated that VEGF only induces angiogenic response in the central nervous system after the administration of extremely large doses. Thus, a dose-dependent study in ischemic rat brain has shown that low (2 µg/7 days) and intermediate (8 µg/7 days) doses of VEGF are not followed by angiogenesis, whereas a high dose (60 µg/7 days) does induce a powerful angiogenic response (Manoonkitiwongsa et al., 2004). Most of the authors who have applied VEGF exogenously to damaged neurons have used doses close to the low one (2 µg/7 days; (Tovar-y-Romo et al., 2007; Lange et al., 2016; Calvo et al., 2018) and have found neuroprotective effects. Altogether, it can be concluded that the neurotrophic and synaptotrophic effects of VEGF are due to a direct action on neurons.
VEGF-B is neuroprotective and non-angiogenic: There is less information regarding the effects of VEGF-B on damaged neurons as compared to VEGF-A. However, recent evidence points to a direct neuroprotective role of VEGF-B in degenerating motoneurons, with the advantage, in relation to VEGF, of not producing vascular side effects, such as inflammation and tissue edema (Poesen et al., 2008). We have also checked the effects of VEGF-B administration to axotomized abducens motoneurons using the same experimental approach as described above. We found that VEGF-B treatment also prevented axotomy-induced alterations (Calvo et al., 2018). This finding is of great clinical relevance because the angiogenic and permeability-promoting activity of VEGF precludes unrestricted delivery at high doses. In contrast, VEGF-B does not induce any vascular side effects, and exerts similar neurotrophic effects with a greater safety, therefore, providing an attractive alternative to VEGF for the treatment of motoneuron diseases.
VEGF neurophysiological actions mediated through its receptors: VEGF binds to two tyrosine kinase receptors, VEGFR-1 and VEGFR-2. Both receptors are present in abducens motoneurons. We found that the selective blockage of VEGFR-2 with the inhibitor SU 1498 prevented the full recovery of abducens motoneuron discharge after axotomy during spontaneous, but not during vestibularly-induced, eye movements (Calvo et al., 2018). These data are in congruence with previous findings indicating that both the vascular and neural effects induced by VEGF are mediated mainly through VEGFR-2 (Lange et al., 2016). On the other hand, VEGF-B only binds to VEGFR-1 and we found that VEGFR-1 prevented abducens motoneurons to enter into an axotomy-like mode of discharge maintaining firing characteristics similar to control during both spontaneous and vestibularly-induced eye movements (Calvo et al., 2018). Since spontaneous and vestibular driven-signals arise from different sources of afferent to abducens motoneurons, our data suggest that signaling through VEGFR-1 or VEGFR-2 could mediate synaptotrophic actions preferentially on different synaptic inputs. VEGF and VEGF-B can also bind to another membrane protein named neuropilin-1. A widely accepted mechanism of action of neuropilin-1 is the formation of VEGF-dependent complexes of VEGFR-1 or VEGFR-2 with neuropilin-1. Through these complexes, neuropilin-1 would act as a co-receptor of VEGFR-1 or VEGFR-2, enhancing the signaling activity and biological function of VEGF. However, other studies suggest the possibility that neuropilin-1 can signal independently of VEGFR-1 or VEGFR-2 acting as a receptor by itself (Lange et al., 2016). Abducens motoneurons express neuropilin-1 so it would be interesting to analyze the possible physiological role of neuropilin-1 in our model.
Conclusions and future perspectives: The trophic theory of neural connections postulates that neurons are strictly dependent on neurotrophic factors, mainly arising from target tissue, for both survival and the maintenance of a normal functional state. After the discovery of the first neurotrophic factor, NGF, it was soon established that this molecule was crucial for postganglionic sympathetic neurons to survive and to exhibit a proper structure/function. In contrast, for decades, neuroscientists have been looking for a specific neurotrophic factor that could be crucial for motoneurons. However, the list of trophic molecules with beneficial effects on motoneurons is extensive, and there is still no consensus about any particular factor as being the main neurotrophic molecule for this neuronal type. We hypothesize that VEGF might be that molecule acting as an essential neurotrophic factor for motoneurons. According to our data, it is the only neurotrophic factor among all those tested in our experimental model able to prevent and recover completely the firing alterations and synaptic loss induced by axotomy in motoneurons (Davis-López de Carrizosa et al., 2009, 2010; Calvo et al., 2018). Moreover, data obtained by other authors (Oosthuyse et al., 2001; Azzouz et al., 2004; Tovar-y-Romo et al., 2007; Lange et al., 2016; Wang et al., 2016) indicate that VEGF is the only neurotrophic factor studied to date whose deficit is followed by motoneuron degeneration while its administration is able to rescue degenerating motoneurons from cell death. If our hypothesis is correct, then it would be expected that the blockage of VEGF in control intact animals should lead to motoneuron alterations in synaptic inputs and firing pattern similar to those found after axotomy. VEGF blockage could be performed either by using virus vectors carrying the sequence of the RNA interference for inhibiting VEGF gene expression, or pharmacologically following the administration of VEGF neutralizing antibodies. This prospect would reveal the likely significant role played by VEGF in motoneuron physiology and provide valuable information regarding the potential use of VEGF as a therapeutic agent for the treatment of damaged or diseased motoneurons.
This work was supported by Ministerio de Economía y Competitividad-FEDER (Grant reference: BFU2015-64515-P) in Spain. PMC was a scholar of MEC (BES-2016-077912) in Spain.
Footnotes
Copyright transfer agreement: The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
References
- 1.Azzouz M, Ralph GS, Storkebaum E, Walmsley LE, Mitrophanous KA, Kingsman SM, Carmeliet P, Mazarakis ND. VEGF delivery with retrogradely transported lentivector prolongs survival in a mouse ALS model. Nature. 2004;429:413–417. doi: 10.1038/nature02544. [DOI] [PubMed] [Google Scholar]
- 2.Calvo PM, de la Cruz RR, Pastor AM. Synaptic loss and firing alterations in Axotomized Motoneurons are restored by vascular endothelial growth factor (VEGF) and VEGF-B. Exp Neurol. 2018;304:67–81. doi: 10.1016/j.expneurol.2018.03.004. [DOI] [PubMed] [Google Scholar]
- 3.Davis-López de Carrizosa MA, Morado-Díaz CJ, Morcuende S, de la Cruz RR, Pastor ÁM. Nerve growth factor regulates the firing patterns and synaptic composition of motoneurons. J Neurosci. 2010;30:8308–8319. doi: 10.1523/JNEUROSCI.0719-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Davis-López de Carrizosa MA, Morado-Díaz CJ, Tena JJ, Benítez-Temiño B, Pecero ML, Morcuende SR, de la Cruz RR, Pastor AM. Complementary actions of BDNF and neurotrophin-3 on the firing patterns and synaptic composition of motoneurons. J Neurosci. 2009;29:575–587. doi: 10.1523/JNEUROSCI.5312-08.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lange C, Storkebaum E, de Almodovar CR, Dewerchin M, Carmeliet P. Vascular endothelial growth factor: a neurovascular target in neurological diseases. Nat Rev Neurol. 2016;12:439–454. doi: 10.1038/nrneurol.2016.88. [DOI] [PubMed] [Google Scholar]
- 6.Manoonkitiwongsa PS, Schultz RL, McCreery DB, Whitter EF, Lyden PD. Neuroprotection of ischemic brain by vascular endothelial growth factor is critically dependent on proper dosage and may be compromised by angiogenesis. J Cereb Blood Flow Metab. 2004;24:693–702. doi: 10.1097/01.WCB.0000126236.54306.21. [DOI] [PubMed] [Google Scholar]
- 7.Oosthuyse B, Moons L, Storkebaum E, Beck H, Nuyens D, Brusselmans K, Van Dorpe J, Hellings P, Gorselink M, Heymans S, Theilmeier G, Dewerchin M, Laudenbach V, Vermylen P, Raat H, Acker T, Vleminckx V, Van Den Bosch L, Cashman N, Fujisawa H, et al. Deletion of the hypoxia-response element in the vascular endothelial growth factor promoter causes motor neuron degeneration. Nat Genet. 2001;28:131–138. doi: 10.1038/88842. [DOI] [PubMed] [Google Scholar]
- 8.Poesen K, Lambrechts D, Van Damme P, Dhondt J, Bender F, Frank N, Bogaert E, Claes B, Heylen L, Verheyen A, Raes K, Tjwa M, Eriksson U, Shibuya M, Nuydens R, Van Den Bosch L, Meert T, D’Hooge R, Sendtner M, Robberecht W, et al. Novel role for vascular endothelial growth factor (VEGF) receptor-1 and its ligand VEGF-B in motor neuron degeneration. J Neurosci. 2008;28:10451–10459. doi: 10.1523/JNEUROSCI.1092-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sathasivam S. VEGF and ALS. Neurosci Res. 2008;62:71–77. doi: 10.1016/j.neures.2008.06.008. [DOI] [PubMed] [Google Scholar]
- 10.Tovar-y-Romo LB, Zepeda A, Tapia R. Vascular endothelial growth factor prevents paralysis and motoneuron death in a rat model of excitotoxic spinal cord neurodegeneration. J Neuropathol Exp Neurol. 2007;66:913–922. doi: 10.1097/nen.0b013e3181567c16. [DOI] [PubMed] [Google Scholar]
- 11.Wang Y, Duan W, Wang W, Di W, Liu Y, Liu Y, Li Z, Hu H, Lin H, Cui C, Li D, Dong H, Li C. scAAV9-VEGF prolongs the survival of transgenic ALS mice by promoting activation of M2 microglia and the PI3K/Akt pathway. Brain Res. 2016;1648:1–10. doi: 10.1016/j.brainres.2016.06.043. [DOI] [PubMed] [Google Scholar]
- 12.Yancopoulos GD, Davis S, Gale NW, Rudge JS, Wiegand SJ, Holash J. Vascular-specific growth factors and blood vessel formation. Nature. 2000;407:242–248. doi: 10.1038/35025215. [DOI] [PubMed] [Google Scholar]