

Abstract
Introduction: The prebiotic potential of herbal medicines has been scarcely studied.
Methods: The authors therefore used anaerobic human fecal cultivation to investigate whether three herbal medicines commonly used in gastrointestinal health and disease in Ayurveda alter the growth and abundance of specific bacterial species.
Results: Profiling of cultures supplemented with Glycyrrhiza glabra, Ulmus rubra, or triphala formulation by 16S rDNA sequencing revealed profound changes in diverse taxa in human gut microbiota. Principal coordinate analysis highlights that each herbal medicine drives the formation of unique microbial communities. The relative abundance of approximately one-third of the 299 species profiled was altered by all 3 medicines, whereas additional species displayed herb-specific alterations. Herb supplementation increased the abundance of many bacteria known to promote human health, including Bifidobacterium spp., Lactobacillus spp., and Bacteroides spp. Herb supplementation resulted in the reduced relative abundance of many species, including potential pathogens such as Citrobacter freundii and Klebsiella pneumoniae. Herbal medicines induced blooms of butyrate- and propionate-producing species. U. rubra and triphala significantly increased the relative abundance of butyrate-producing bacteria, whereas G. glabra induced the largest increase in propionate-producing species. To achieve greater insight into the mechanisms through which herbal medicines alter microbial communities, the authors assessed the shifts in abundance of glycosyl hydrolase families induced by each herbal medicine. Herb supplementation, particularly G. glabra, significantly increased the representation and potential expression of several glycosyl hydrolase families.
Discussion: These studies are novel in highlighting the significant prebiotic potential of medicinal herbs and suggest that the health benefits of these herbs are due, at least in part, to their ability to modulate the gut microbiota in a manner predicted to improve colonic epithelium function, reduce inflammation, and protect from opportunistic infection. Forthcoming studies in human clinical trials will test the concordance of the results generated in vitro and the predictions made by genome analyses.
Keywords: : prebiotic, ayurveda, microbiota, gastrointestinal, anti-inflammatory, herb
Introduction
In the United States alone, 60–70 million people are affected by some type of digestive disease.1 Ayurveda is the traditional system of medicine originating in India that emphasizes gastrointestinal health and disease prevention with herbal and lifestyle medicine. The herbal medicines commonly used for gastrointestinal health and disease in Ayurvedic Medicine, as well as other traditional systems of medicine, include Ulmus rubra (common name: slippery elm) and Glycyrrhiza glabra (common name: licorice), as well as the widely known polyherbal medicinal formulation (common name: triphala) of Emblica officinalis, Terminalia bellerica, and Terminalia chebula, which were three herbal medicines of greatest interest for the current study.
Triphala is an extensively clinically studied formulation containing three dried fruits, most commonly in equal proportions. While this herbal medicine has many clinical applications in Ayurveda, it is considered of special importance for gastrointestinal disorders ranging from constipation to colitis. Triphala exerts enteroprotective effects and promotes health of the gut epithelium and villi through improved barrier function and nutrient absorption.2 Licorice root, which is referred to as Yashtimadhu in Ayurveda, is often prescribed as a demulcent, mild laxative, and anti-inflammatory.3 Slippery elm (common names: red or Indian elm) is frequently used as a demulcent and emollient for gastrointestinal and other mucous membranes.4 Both licorice and slippery elm are often used to treat hyperacidity, ulcerative conditions, and compromised mucous membranes. However, slippery elm is classified as endangered by multiple plant preservation-focused organizations such as the United Plant Savers, and thus, substitutes should be considered in clinical treatment.
While the health benefits of herbal medicines on gastrointestinal health have been well documented, the prebiotic potential of medicinal herbs has only been scarcely investigated. The most commonly accepted, but debated, definition of prebiotics refers to dietary carbohydrates selectively fermented by gut microbiota that modulate the microbiota composition to result in health benefits to the host.5,6 The impact of diet on gut microbiota has been reported in several studies.7,8 Likewise, various plants and botanical extracts alter the gut microbiota composition as do compounds derived or isolated from those plants, for example, various polyphenolic compounds.9–11 Prebiotics are defined as undigested dietary carbohydrates that alter the gut microbiota and promote human health.6,12 This restrictive language is distinct because the effects of prebiotics are specific to gut microbiota without direct effects on the host, which lacks the glycosyl hydrolases (GHs) required to catabolize or digest polysaccharides.
Prebiotics reach the site of action in the colon mostly unmetabolized and are broken down by extracellular GH; for example, Bacteroides spp., Parabacteroides spp., and Bifidobacterium spp. liberate oligo-, di-, and monosaccharides that are subsequently fermented by saccharolytic microbes. The end products of microbial carbohydrate metabolism in anaerobic environments such as the colon often include short chain fatty acids (SCFAs) such as butyrate which can act as a histone deacetylase (HDAC) inhibitor, inhibit nuclear factor kappa B (NF-κB) and the subsequent expression of pro-inflammatory cytokines, induce regulatory T cells (Tregs), promote improved gut barrier function, and participate in gut–brain axis signaling.13,14
Prebiotics have been evaluated in several diverse disease contexts with varying outcomes.15–17 These diseases, which are largely inflammatory diseases of the gut and associated with reduced barrier function, share a common sharp increase in incidence over recent decades that strongly implicate environmental factors, namely diet and gut microbiota, as important drivers of disease. In the United States, the average adult consumes only 12–18 g/day of dietary fiber, whereas people in rural Africa subsisting on a plant-based diet consume over 100 g/day.18 The origin, chemical composition, and physicochemical properties, including chain length, cross-linking, and branching, dictate the specificity of bacterial taxa capable of efficiently metabolizing glycans.19,20 Thus, the authors hypothesized that the complex carbohydrates present in herbs may be potent drivers to alter gut microbiota composition.2
Methods
Study participants and sample collection
Healthy English-speaking women and men aged 30–60 years who had previously adhered to a vegetarian or vegan diet for >1 year were recruited to donate a single stool sample. The study was approved by the Sanford Burnham Prebys Medical Discovery Institute Institutional Review Board (IRB-2014-020), and all study participants provided written informed consent before participation. Participants ate their normal diets and donated a morning fecal sample in stool hats. The fecal samples were transferred to conical tubes then frozen on ice and stored at −80°C until further processing.
Anaerobic fecal cultures
Equal volumes of stool collected from 12 participants were pooled and used to inoculate a chemically defined medium (CDM). CDM is buffered with HEPES and NaHCO3 and contains all 20 amino acids, phosphate (KH2PO4, Na2HPO4), and a mineral mix supplying sulfate in the form of Mg and Mn salts, Fe, Zn, Cu, Al salts and Co, B, Ca, Mo, Se, W and Ni salts. Nucleoside bases, including Inosine, Xanthine, and additional compounds including: Choline, Ascorbic acid, Lipoic acid, Hemin, and Inositol were added to the CDM. Resazurin (1 mg/L) was added to visually monitor dissolved oxygen. 2 × CDM and herbs (powder) in sterile water (2%) were reduced in an anaerobic chamber (Coy Labs) for 3 days. The herb sources are as follows: triphala powder (Banyan Botanicals) (Emblica officinalis, Terminalia bellerica, and Terminalia chebula) (fruits), slippery elm powder (Now foods) (Ulmus rubra) (bark), and licorice (Solaray Nutraceutical) (Glycyrrhiza glabra) (root). Fecal pools were used to inoculate reduced 2 × CDM that was distributed in equal volumes to replicate Hungate tubes containing 2 × herbs. Anaerobic cultures (20% CO2, 5% H2, 75% N2) were grown statically for 3–4 days at 37°C. Technical replicates of cultures (n = 6) with each herb and control cultures without herb (n = 2) were grown to approximate saturation.
Microbial DNA isolation
Genomic DNA was isolated from cultures, as well as the fecal inoculums (FIs), using the procedures of the QIAamp DNA Stool Kit (Qiagen) with a modification that included an additional step of bead beating using the FastPrep instrument (Bio 101) to ensure uniform lysis of bacterial cells.
Microbiota analysis by 16S rDNA sequencing
Multiplexed 16S rDNA libraries were prepared using standard 16S metagenomic sequencing library protocols from Illumina, which uses the V3–V4 region of 16S rDNA for target amplification. The authors performed paired end reads (250 bp) sequencing to generate ∼200,000 sequences/sample using the Illumina MiSeq. Subsequent analysis was done in CLC Microbial Genomics Module 2.5 (Qiagen) and R.21 Paired end reads were merged (mismatch cost–2, minimum score–8, gap cost–3, maximum unaligned end mismatches–0) and trimmed to the same length. Additional quality filter steps were applied to exclude short reads, sequences with poor quality scores, and chimeras. To ensure comparable high coverage in all samples, the authors excluded samples producing <35,000 reads. They did not use OTU-based enumeration of taxa due to the over-merging that occurs. Instead each unique 16S rDNA sequence was subjected to BLAST using the NCBI 16S rRNA database (Bacteria and Archaea) to identify best matches to taxa at the genus and species levels based on % identity.
Results
Triphala, slippery elm, and licorice have potent microbiota modulatory effects in vitro
To study the direct effects of these herbal medicines in the absence of dietary and host-driven processes, the authors used in vitro anaerobic cultivation of human FIs representing a pool of 12 healthy vegetarian donors. Gut bacterial taxa capable of catabolizing herbal medicines as a source of energy will have a growth advantage revealed by increased relative abundance in the community. The authors generated an average of 337,186 reads from each anaerobic culture (average triphala: 185,681; slippery elm: 364,942; Licorice: 460,937). The authors compared the relative abundance of gut species after cultivation in CDM lacking carbohydrate energy sources to cultures grown in CDM supplemented with licorice, slippery elm, or triphala. Collectively, 299 unique bacterial species belonging to 131 genera were observed in the cultures (Supplementary Table S1; Supplementary Data are available online at www.liebertpub.com/acm). By comparing the taxa present in the fecal pool inoculum (fecal pool without cultivation), to those present in one or more cultures, the authors conclude that their optimized anaerobic cultivation conditions represent virtually all (>99%) taxa present in human gut microbiota. The nearly complete representation of fecal microbiota is unique and not observed in screens involving a wide variety of 30 prebiotic isolates (unpublished observation).
Supplementation of CDM with each of the three herbal medicines resulted in significant shifts in the relative abundance of specific taxa compared to control cultures. The magnitude of these shifts was largely equivalent for each herbal medicine. As expected, each herbal medicine generated a unique microbial profile that is well separated from one another, as depicted in PCoA plots using weighted UniFrac22 (Fig. 1). These plots indicate that control cultures lacking carbohydrate energy sources were the most significantly different compared to the FI. The bacterial communities generated in response to herbal medicines underwent prominent shifts resulting in communities that were more similar to the FI. In addition, cultures supplemented with triphala and slippery elm were more similar to each other compared to licorice. The bacterial response to triphala and slippery elm was consistent across replicate cultures, whereas profiles generated from licorice-supplemented cultures displayed more variability.
FIG. 1.
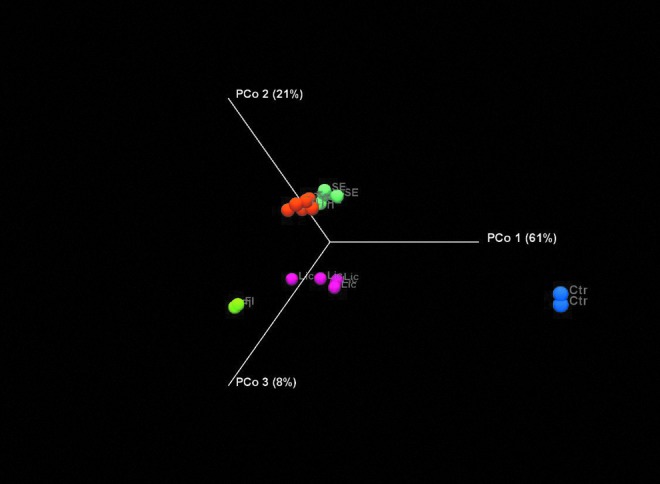
PCoA weighted UniFrac. Weighted UniFrac analysis of β−diversity of each culture measures abundance of bacterial communities present in each culture. Fecal inoculum (FI) (bright green), control (Ctr) (blue), triphala (Tri) (orange), slippery elm (SE) (light green), licorice (Lic) (pink).
Genus level impact of herbal medicines
Among the 131 genera observed, the relative abundance of 99 (75%) was significantly altered by these three herbal medicines (Supplementary Table S1). The number of genera displaying increased relative abundance for each herb was comparable for triphala (47), slippery elm (56), and licorice (44); however, only 17 genera were induced by all three herbal medicines. The magnitude of these changes ranged from 3.4 to over 2000-fold increases (Table 1). Conversely, the relative abundance of 10 genera was reduced by all 3 herbal supplements. The largest increases in relative abundance induced by these herbal medicines were associated with genera present in very low abundance in control cultures. In some instances, these changes induced certain genera to become dominant in the community, such as Acidaminococcus.
Table 1.
Genera Altered by Herbs
| NCBI genus | Ctrl (avg) | Triph (avg) | SE (avg) | Lic (avg) |
|---|---|---|---|---|
| Genera increased by herbs, n (%) | ||||
| Acidaminococcus | 0.002 | 4.884 | 0.415 | 3.421 |
| Bacteroides | 1.954 | 24.472 | 24.042 | 45.932 |
| Bariatricus | 0.000 | 0.013 | 0.040 | 0.349 |
| Clostridium | 0.009 | 0.094 | 2.844 | 0.030 |
| Collinsella | 0.008 | 0.077 | 0.117 | 0.129 |
| Eisenbergiella | 0.000 | 0.020 | 0.186 | 0.013 |
| Emergencia | 0.001 | 0.025 | 0.034 | 0.249 |
| Erysipelatoclostridium | 0.000 | 0.807 | 0.825 | 0.036 |
| Flavonifractor | 0.000 | 0.121 | 0.379 | 0.180 |
| Frisingicoccus | 0.001 | 0.523 | 0.108 | 0.044 |
| Longibaculum | 0.000 | 0.102 | 0.042 | 0.026 |
| Lutispora | 0.005 | 0.091 | 0.430 | 0.044 |
| Parabacteroides | 0.304 | 2.232 | 1.409 | 1.025 |
| Phascolarctobacterium | 0.028 | 4.344 | 4.742 | 5.235 |
| Pseudoflavonifractor | 0.002 | 0.147 | 0.463 | 0.154 |
| Clostridium (Ruminococcaceae) | 0.079 | 0.252 | 0.499 | 0.544 |
| Sutterella | 0.018 | 7.407 | 5.482 | 2.786 |
| Genera inhibited by herbs, n (%) | ||||
| Akkermansia | 0.040 | 0.000 | 0.000 | 0.000 |
| Blautia | 2.011 | 0.154 | 0.614 | 0.326 |
| Citrobacter | 39.793 | 0.422 | 0.366 | 0.241 |
| Enterobacter | 2.928 | 0.014 | 0.434 | 0.744 |
| Enterococcus | 13.878 | 2.359 | 4.897 | 1.264 |
| Kluyvera | 0.096 | 0.000 | 0.008 | 0.010 |
| Muribaculum | 0.048 | 0.000 | 0.000 | 0.000 |
| Prevotella | 0.018 | 0.006 | 0.000 | 0.000 |
| Pseudocitrobacter | 0.032 | 0.000 | 0.003 | 0.000 |
| Shimwellia | 0.011 | 0.000 | 0.000 | 0.000 |
avg, average; Lic, licorice; SE, slippery elm; Triph, triphala.
The authors focused on genera that were present in relative abundance of >0.01% and further increased or decreased by the herbal medicines. Triphala induced large increases in the relative abundance of Dorea, Sutterella, Phascolarctobacterium, Lactobacillus, and Butyrivibrio (range = 116–580-fold). Smaller increases were observed for Lachnoclostridium, Oscillibacter, Eubacterium, Roseburia, and Bacteroides (range = 12–70-fold). While Bacteroides were only increased by 12-fold, this shift is highly significant since its final average relative abundance represented 24.5% of the community. Slippery elm yielded similar outcomes with large increases for Dorea, Sutterella, and Phascolarctobacterium (range = 169–298-fold), whereas Lactobacillus, Lachnoclostridium, Oscillibacter, Flintibacter, Anaerostipes, Bifidobacterium, Desulfovibrio, Bacteroides, Eubacterium, and Butyrivibrio were induced to a smaller extent (range = 11–92-fold) with final average relative abundance of 0.2%–24%. The taxa induced by licorice were distinct, although shared features in common with triphala and slippery elm. Licorice induced very large increases in genera that were not detected or present in very low abundance in control cultures that achieved high final average relative abundance values, including Pseudomonas, Acinetobacter, Victivallis, and Acidaminococcus (range = 1.2%–3.0%), whereas Achromobacter, Anaerofilum, Anseongella, Bariatricus, Marvinbryantia, Rhizobium, and Sphingomonas achieved lower but significant final average relative abundance (range = 0.1%–0.3%). Finally, Phascolarctobacterium and Sutterella displayed large increases (range = 152–186-fold), whereas Parasutterella, Cloacibacillus, Desulfovibrio, Roseburia, Bacteroides, Bifidobacterium, and Gemmiger were induced to a lesser extent (range = 10–85-fold). These results highlight both common and distinct features of each herbal medicine and its potential prebiotic effects on gut microbiota.
Species responsiveness to herbal medicine supplementation
Among the 299 species profiled, 224 were observed at an average relative abundance of >0.01% of the total community in at least 1 of the culture conditions analyzed. Subsequent analyses were restricted to these moderate-to-high abundance species (Fig. 2). A total of 75 species (33%) responded uniformly in direction to all 3 herbal medicines with 42 (∼19%) displaying increased and 33 (∼15%) displaying decreased relative abundance compared to control cultures. Prominent among the positive response to all three herbal medicines tested were six Clostridium spp., 14 Bacteroides spp., three Parabacteroides spp., and two Bifidobacterium, Phascolarctobacterium, and Sutterella spp. Among the species negatively impacted by all herbal supplements were five Citrobacter spp., including C. freundii. In addition, the relative abundance of three Enterobacter spp. and Enterococcus spp. was reduced by all three herbal medicines. Two Cronobacter and Klebsiella spp., including K. pneumonia, were reduced in abundance in cultures containing each herbal medicine tested.
FIG. 2.

Hierarchical cluster plots. Heat maps were prepared using R and show relative abundance of species in each herbal culture. Accompanying box shows the color scale with dark green being least abundant and red being most abundant. C, control; Lic, licorice; SE, slippery elm; Tri, triphala.
Overall, 16S rDNA profiles highlighted the stronger relationship between the taxa responsive to triphala and slippery elm, consistent with PCoA plots (Fig. 1). The authors observed concordant behavior of 159 species (71%) in response to these herbal medicines, whereas licorice supplementation displayed more differences than similarities compared to the other herbal medicines tested. Triphala and slippery elm both induced the expanded representation of Clostridium spp. (11), Ruminococcus spp. (2), Bacteroides spp. (18), Eubacterium spp. (4), and Oscillibacter spp. (2). Licorice uniquely induced the expansion of many poorly described taxa; however, licorice induced an increased relative abundance of five sulfate-reducing bacteria belonging to Desulfovibrio spp., three of which were uniquely observed in cultures supplemented with licorice.
The complexity of the microbial responses to these herbal medicines is too high to easily describe; however, some noteworthy observations can be summarized. Licorice generally had the greatest magnitude increase of Bacteroides spp. compared to slippery elm and triphala. Licorice supplementation generally resulted in communities with reduced relative abundance of Eubacterium spp. It is of potential importance that licorice strongly reduced the relative abundance of opportunistic pathogens Enterococcus faecalis and Klebsiella pneumoniae. At the species level, the response of fecal microbiota to slippery elm and triphala is highly similar, although notable differences are also evident. Slippery elm compared to other herbs generally induced the greatest increases in Ruminococcus spp., R. obeum, R. gnavus, and R. torques. Triphala induced the strongest increase in the relative abundance of Butyrivibrio crossotus and Clostridium hathewayi, renamed Hungatella hathewayi and H. effluvii.
Impact of herbal medicines on butyrate and propionate producers
The authors compiled the species known to encode gene functions required for butyrate and propionate generation. Most of butyrate producers described in the literature were observed in the cultures, particularly those supplemented with slippery elm and triphala (Fig. 3a). As expected, control cultures with no exogenous sugar source featured reduced relative abundance of butyrate producing bacterial species. Licorice supplementation elevated the relative abundance of butyrate producers only modestly as the result of increased relative abundance of specific Clostridium spp., whereas both triphala and slippery elm displayed a significant increase in the relative abundance of butyrate producers. Slippery elm promoted the expansion of Flintibacter spp., predominantly F. plautii and Clostridium spp. belonging to cluster XIVa, whereas triphala induced a similar expansion of Clostridium spp.; additional butyrate producing potential may be driven by Butyrivibrio spp. but not Flintibacter. A smaller number of propionate producers were observed in fecal culture. All cultures supplemented with herbs displayed increased relative abundance of propionate producers compared to control cultures (Fig. 3b). Slippery elm displayed modest but significant increases, whereas triphala and Licorice induced a greater expansion of propionate producers, driven primarily by two Bacteroides spp., namely B. thetaiotaomicron and B. vulgatus.
FIG. 3.

(a) Species with butyrate biosynthetic potential. Abundance of butyrate producing species was summed and depicted as genera. (b) Species with propionate biosynthetic potential. Abundance of propionate producing species was summed.
Herbal medicine supplementation influences glycosyl hydrolase representation
In vitro prebiotic effects are driven by media components utilized by members of the community as energy sources (e.g., carbohydrates and amino acids). The taxa impacted by licorice, slippery elm, and triphala suggested that the catabolism of polysaccharides present in each herbal medicine may be a prominent fitness determinant in fecal communities. The authors therefore analyzed the 16S rDNA profiles in the context of the suite of GH encoded by genomes characterized in the CAZy database.23 The authors identified 66 reference genomes profiled in fecal communities for which GH families have been annotated. They tabulated the GH families and the number of gene copies present in each genome together with the relative abundance measure of each species to visualize differential representation of the most abundant GH families (Fig. 4). Even though GH representation has not been documented in all gut species profiled, the results are representative of the community as a whole since the reference genomes used provided excellent coverage in terms of total community abundance: control (62%), licorice (42%), slippery elm (50%), and triphala (37%). All herbal medicine supplemented cultures increased the potential representation of GH3 (β-glucosidase), GH4 (maltose-6-phosphate glucosidase), GH21, GH29 (α-L-fucosidase), GH44 (endoglucanase), and GH93 (exo-α-L-1,5-arabinanase). These activities are likely to be the key to driving the alterations in species abundance and catabolism of herbal medicines and reflect the possible major sugar components of licorice, slippery elm, and triphala.
FIG. 4.

GH representation. The y-axis depicts the weighted functional potential of GHs encoded by communities with and without herb supplementation. GH families and copy number derived from reference genomes observed in culture were multiplied by percent abundance of each species encoding them. The sum of these values is shown. GH, glycosyl hydrolase.
Discussion
The health benefits of triphala, slippery elm, and licorice are well documented; however, the impact of these herbal medicines on the human gut microbiota has been only scarcely studied. Triphala, licorice, and slippery elm are cornerstone treatments for gastrointestinal health and disease in Ayurveda and other traditional systems of medicine. Triphala is considered a rasayana medicine, which classically categorizes it as a tonic that provides immunity and rejuvenation. In the context of gut health for which is it most renowned, the formula is often used to treat ulcers, constipation, abdominal pain, and colitis.2 Similarly, licorice is a rasayana and used to treat mucosal inflammation, dryness, and ulcerations.3 Slippery elm is similarly used to protect mucosal barrier function in gastrointestinal illness and is more mucilaginous compared to licorice.4,24 The effects of the mucilaginous features of each herbal medicine based on their respective carbohydrate content in the context of the human gut microbiota metabolism was of particular interest.
The study of in vitro anaerobic fecal cultures offers several advantages, including the ease and low-cost cultivation highly representative of human microbiota. In vitro cultures simplify interpretation of microbiologic processes that occur in the absence of complex and poorly defined host-dependent effects. The percentage of gut microbes considered cultivable is widely debated; however, it is noteworthy that the three herbal medicines tested in this study allowed virtually all the observed species present in feces to be cultivated, suggesting that herbal medicine supplementation represents unique media that permit a highly representative microbiota community architecture. It remains largely untested to what extent in vitro cultures stimulated with prebiotics are reproduced in vivo.
Fecal cultivation permits the study of gut microbial communities in CDM that facilitates single variable comparisons to be examined. While useful, such cultivation techniques fail to replicate the acid and enzymatic hydrolysis of dietary components before entering the site of action in the colon, which is where the majority of prebiotic catabolism occurs. Prebiotics reach the colon relatively intact suggesting that their catabolism and metabolism are microbiologic processes largely occurring independent of host-derived influence.25 It is anticipated that the effects of medicinal herbs in vivo would be more constrained than those observed in vitro due to the complexity of individual diets, immune regulation, or bile acids that further influence microbiota composition.26 The authors elected to study the effect of herbal medicines in vegetarian subjects reasoning that their gut microbiota would be well adapted to catabolism of plant-based diets without the potentially confounding effects accompanying an omnivorous diet. It might be anticipated that similar studies examining these herbal medicines in human omnivores would result in potentially overlapping but distinct outcomes.27–30 While these limitations should be noted, anaerobic cultivation is advantageous in delineating microbiologic phenomenon that occurs independent of host factors as detailed in this study.
Herbal medicine supplementation of human fecal cultures resulted in profound changes in the relative abundance of many phylogenetically diverse gut species. Each herbal medicine evaluated in this study uniquely altered gut bacterial communities; however, the microbial composition of triphala and slippery elm supplemented cultures more closely resembled one another, whereas licorice induced substantially unique profiles.
Alterations in fecal cultures are driven by media components that bacteria can readily metabolize as energy sources that provide a fitness advantage by promoting increased cell division. In anaerobic environments, saccharolytic bacteria derive energy predominantly from sugar fermentation, whereas other bacteria derive energy through amino acid metabolism. By comparing control cultures lacking carbohydrate energy sources, but containing amino acids, the authors observed that species specializing in amino acid fermentation are uniformly, but not exclusively, diminished in cultures supplemented with herbal medicines, thus highlighting the preference for sugar fermentation when complex carbohydrates are available. Featured prominently in control cultures are members of the Firmicutes such as Clostridium oroticum and Enterococcus spp. and Proteobacteria such as Escherichia spp., Enterobacter spp., Citrobacter spp., and Klebsiella spp. These observations are consistent with known amino acid fermentation pathways and activities of these taxonomic groups.31
While difficult to quantify, herbal medicine supplementation induced a carbohydrate catabolism dominant response based on the species undergoing the greatest increase in relative abundance. These species were predominantly those encoding large repertoires of GH (i.e., Bacteroides, Parabacteroides, and Bifidobacteria). GHs are expressed and localized to the extracellular space to permit energy extraction in the form of mono-, di-, and oligosaccharides that also promote cross-feeding of bacterial species lacking relevant GH specificities.32 The strong enrichment of GH families induced by each herbal medicine substantiates their conclusion that sugar metabolism is a driver of alterations in gut microbiota composition. Cross-feeding on liberated sugars was also evident in herb-supplemented cultures featuring known sugar-fermenting species (i.e., Clostridium, Ruminococcus, Flintibacter, and many others).
These findings are consistent with the sources of glycans and sugar composition of the herbal medicines evaluated. The sugar composition of triphala, slippery elm, and licorice is incompletely defined; however, some information for each has been documented in the literature. In terms of relevant biochemical constituents, slippery elm is known to contain a high content of polysaccharides, including linear chains of alternating D-galacturonic acid and L-rhamnose residues with α-linkages containing galactose and 3-O-methyl galactose.33,34 Also present in high quantities are starch (cellulose) and lignin with lower quantities of mucilage, gums, and pectin. Licorice has a wide range of sugars, including fucose, rhamnose, arabinose, galactose, glucose, mannose, ribose, xylose, fructose, galacturonic acid, and glucuronic acid.35 The sugars present in triphala include arabinose, galactose, xylose, glucose, mannose, rhamnose, galacturonic acid, and arabinogalactan.36,37 The specific sugars and sugar linkages present in each herbal medicine ultimately dictate which species are able to catabolize and, therefore, metabolize herb glycan components as an energy source.
Interestingly, herbal medicine supplementation also resulted in increased relative abundance of a group of asaccharolytic species that presumably derive energy through protein catabolism of herbal medicines; these species include: Cloacibacillus evryensis, Phascolarctobacterium faecium, P. succinatutans, Sutterella wadsworthensis, S. massillensis, Parasutterella excrementihominis, Collinsella aerofaciens, Eggerthella lenta, and Slackia isoflavoniconvertens.38,39 These results indicate that both the sugar and protein content of these herbal medicines drive alterations in gut microbiota profiles.
The authors speculate that human consumption of triphala, licorice, or slippery elm will induce similar alterations in gut microbiota since prebiotic effects are largely microbiologic processes driven by glycan microbial metabolism. The magnitude of alterations observed in vitro may be significantly dampened in vivo owing to inherent resilience of gut microbiota. It is predicted that in humans these herbal medicines have a strong potential to elevate butyrate and propionate levels. Butyrate is known to promote anti-inflammatory responses through activation of IL-10 producing Tregs, an improved gut barrier, and the accompanying reduction of circulating lipopolysaccharide and microbial and food antigens.40 In addition, butyrate is a HDAC inhibitor that increases expression of Muc2 and other mucins, which further promotes proper gut barrier function and global alterations of gene expression.41,42 Propionate is transported to the liver where it serves as substrate for gluconeogenesis, lipogenesis, and protein synthesis.43,44 Moreover, both butyrate and propionate are GPR41 ligands45 expressed by enteroendocrine L cells inducing GLP-1 production that drives pancreatic β cells to secrete insulin and control blood glucose levels.46 Thus, the induction of gut microbiota SCFA production confers diverse health benefits to the host.
The taxa most positively impacted by herbal medicine supplementation predominantly represent species with documented health promoting qualities. A growing number of studies demonstrate health benefits of Bifidobacterium spp. and Lactobacillus spp. in probiotic formulations.47,48 Some Bacteroides species have demonstrated immune modulating activities, including B. thetaiotaomicron and B. fragilis which secrete a polysaccharide (PSA) in outer membrane vesicles to stimulate IL-12/Stat4 dependent differentiation of CD4+ cells to TH1 lineage.49 Furthermore, B. fragilis promotes improved gut barrier function.50 These speculations require testing and prebiotic validation in healthy human subjects, as well as those suffering from diverse gastrointestinal disorders and disease such as allergy, autoimmunity, asthma, neurodegeneration, and cancer, and represent areas in need of further study. It will be of interest to determine whether the suppression of opportunistic pathogens observed in vitro will occur in humans. Citrobacter freundii and Klebsiella pneumoniae have been implicated as drivers of “leaky gut” or increased intestinal permeability, and Enterococcus faecalis is a well-documented enteric pathogen capable of causing significant disease.51,52 While these species are often detected in healthy individuals, additional studies focused on subjects with gastrointestinal disorders, or overgrowth of such species will be needed to assess the repressive potential of these herbal medicines.
These results, for the first time, provide evidence that the health benefits of Ayurvedic herbal medicines may be mediated by the glycan catabolic activities of the human gut microbiome. These studies are novel in highlighting the significant prebiotic potential of herbal medicines and suggest that the health benefits of these herbs are due, at least in part, to their ability to modulate the gut microbiota in a manner predicted to improve colonic epithelium function, reduce inflammation, and promote protection from bacterial opportunistic pathogenic infection. Forthcoming studies in human clinical trials will test the concordance of the results generated in vitro and the predictions made by genome analyses.
Supplementary Material
Acknowledgments
This work was supported by a research grant from the Samuel Lawrence Foundation and a fellowship grant from the Chopra Foundation.
Author Disclosure Statement
D.C. is a founder of the Chopra Foundation and Chopra Center and a co-owner of the Chopra Center. P.J.M. is Director of Research for the Chopra Foundation.
References
- 1.DHHS. Opportunities and Challenges in Digestive Diseases Research: Recommendations of the National Commission on Digestive Diseases. Bethesda, MD: National Institutes of Health 08-6514, 2009 [Google Scholar]
- 2.Peterson CT, Denniston K, Chopra D. Therapeutic uses of triphala in ayurvedic medicine. J Altern Complement Med 2017;23:607–614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hosseinzadeh H, Nassiri-Asl M. Pharmacological effects of glycyrrhiza spp. and its bioactive constituents: Update and review. Phytother Res 2015;29:1868–1886 [DOI] [PubMed] [Google Scholar]
- 4.Ke F, Yadav PK, Ju LZ. Herbal medicine in the treatment of ulcerative colitis. Saudi J Gastroenterol 2012;18:3–10 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gibson GR, Roberfroid MB. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J Nutr 1995;125:1401–1412 [DOI] [PubMed] [Google Scholar]
- 6.Hutkins RW, Krumbeck JA, Bindels LB, et al. Prebiotics: Why definitions matter. Curr Opin Biotechnol 2016;37:1–7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.David LA, Maurice CF, Carmody RN, et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014;505:559–563 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Llewellyn SR, Britton GJ, Contijoch EJ, et al. Interactions between diet and the intestinal microbiota alter intestinal permeability and colitis severity in mice. Gastroenterology 2017;154:1037–1046 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Liu D, Zhang Y, Liu Y, et al. Berberine modulates gut microbiota and reduces insulin resistance via the TLR4 signaling pathway. Exp Clin Endocrinol Diabetes 2018. DOI: 10.1055/s-0043-125066 [DOI] [PubMed]
- 10.Cheng M, Zhang X, Zhu J, et al. A metagenomics approach to the intestinal microbiome structure and function in high fat diet-induced obesity mice fed with oolong tea polyphenols. Food Funct 2018;9:1079–1087 [DOI] [PubMed] [Google Scholar]
- 11.Wang S, Yao J, Zhou B, et al. Bacteriostatic effect of Quercetin as an antibiotic alternative in vivo and its antibacterial mechanism in vitro. J Food Prot 2018;81:68–78 [DOI] [PubMed] [Google Scholar]
- 12.Bird AR, Conlon MA, Christophersen CT, Topping DL. Resistant starch, large bowel fermentation and a broader perspective of prebiotics and probiotics. Benef Microbes 2010;1:423–431 [DOI] [PubMed] [Google Scholar]
- 13.Kim CH, Park J, Kim M. Gut microbiota-derived short-chain fatty acids, T cells, and inflammation. Immune Netw 2014;14:277–288 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Furusawa Y, Obata Y, Fukuda S, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013;504:446–450 [DOI] [PubMed] [Google Scholar]
- 15.Manning TS, Gibson GR. Microbial-gut interactions in health and disease. Prebiotics. Best Pract Res Clin Gastroenterol 2004;18:287–298 [DOI] [PubMed] [Google Scholar]
- 16.Toward R, Montandon S, Walton G, Gibson GR. Effect of prebiotics on the human gut microbiota of elderly persons. Gut Microbes 2012;3:57–60 [DOI] [PubMed] [Google Scholar]
- 17.Kolida S, Gibson GR. Prebiotic capacity of inulin-type fructans. J Nutr 2007;137:2503S–2506S [DOI] [PubMed] [Google Scholar]
- 18.Sonnenburg ED, Sonnenburg JL. Starving our microbial self: The deleterious consequences of a diet deficient in microbiota-accessible carbohydrates. Cell Metab 2014;20:779–786 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Elleuch M, Bedigian D, Maazoun B, et al. Improving halva quality with dietary fibres of sesame seed coats and date pulp, enriched with emulsifier. Food Chem 2014;145:765–771 [DOI] [PubMed] [Google Scholar]
- 20.Bourquin LD, Titgemeyer EC, Fahey GC. Jr. Vegetable fiber fermentation by human fecal bacteria: Cell wall polysaccharide disappearance and short-chain fatty acid production during in vitro fermentation and water-holding capacity of unfermented residues. J Nutr 1993;123:860–869 [DOI] [PubMed] [Google Scholar]
- 21.Eglen SJ. A quick guide to teaching R programming to computational biology students. PLoS Comput Biol 2009;5:e1000482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lozupone CA, Hamady M, Kelley ST, Knight R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl Environ Microbiol 2007;73:1576–1585 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Lombard V, Golaconda Ramulu H, Drula E, et al. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res 2014;42:D490–D495 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hawrelak JA, Myers SP. Effects of two natural medicine formulations on irritable bowel syndrome symptoms: A pilot study. J Altern Complement Med 2010;16:1065–1071 [DOI] [PubMed] [Google Scholar]
- 25.Kolida S, Tuohy K, Gibson GR. Prebiotic effects of inulin and oligofructose. Br J Nutr 2002;87 Suppl 2:S193–S197 [DOI] [PubMed] [Google Scholar]
- 26.Zheng X, Huang F, Zhao A, et al. Bile acid is a significant host factor shaping the gut microbiome of diet-induced obese mice. BMC Biol 2017;15:120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Healey GR, Murphy R, Brough L, et al. Interindividual variability in gut microbiota and host response to dietary interventions. Nutr Rev 2017;75:1059–1080 [DOI] [PubMed] [Google Scholar]
- 28.Ferrocino I, Di Cagno R, De Angelis M, et al. Fecal microbiota in healthy subjects following omnivore, vegetarian and vegan diets: Culturable populations and rRNA DGGE profiling. PLoS One 2015;10:e0128669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Wu GD, Compher C, Chen EZ, et al. Comparative metabolomics in vegans and omnivores reveal constraints on diet-dependent gut microbiota metabolite production. Gut 2016;65:63–72 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Zimmer J, Lange B, Frick JS, et al. A vegan or vegetarian diet substantially alters the human colonic faecal microbiota. Eur J Clin Nutr 2012;66:53–60 [DOI] [PubMed] [Google Scholar]
- 31.Dai ZL, Wu G, Zhu WY. Amino acid metabolism in intestinal bacteria: Links between gut ecology and host health. Front Biosci (Landmark Ed) 2011;16:1768–1786 [DOI] [PubMed] [Google Scholar]
- 32.Sathya TA, Khan M. Diversity of glycosyl hydrolase enzymes from metagenome and their application in food industry. J Food Sci 2014;79:R2149–R2156 [DOI] [PubMed] [Google Scholar]
- 33.Hough L, Jones JK, Hirst EL. Chemical constitution of slippery elm mucilage; isolation of 3-methyl d-galactose from the hydrolysis products. Nature 1950;165:34. [DOI] [PubMed] [Google Scholar]
- 34.Bacon JS, Cheshire MV. Apiose and mono-O-methyl sugars as minor constituents of the leaves of deciduous trees and various other species. Biochem J 1971;124:555–562 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Wittschier N, Faller G, Hensel A. Aqueous extracts and polysaccharides from liquorice roots (Glycyrrhiza glabra L.) inhibit adhesion of Helicobacter pylori to human gastric mucosa. J Ethnopharmacol 2009;125:218–223 [DOI] [PubMed] [Google Scholar]
- 36.Chatterjee UR, Bandyopadhyay SS, Ghosh D, et al. In vitro anti-oxidant activity, fluorescence quenching study and structural features of carbohydrate polymers from Phyllanthus emblica. Int J Biol Macromol 2011;49:637–642 [DOI] [PubMed] [Google Scholar]
- 37.Nosalova G, Jurecek L, Chatterjee UR, et al. Antitussive activity of the water-extracted carbohydrate polymer from Terminalia chebula on citric acid-induced cough. Evid Based Complement Alternat Med 2013;2013:650134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kiwerski J. [Spinal injuries caused by the flexion mechanism]. Chir Narzadow Ruchu Ortop Pol 1989;54:304–311 [PubMed] [Google Scholar]
- 39.Watanabe Y, Nagai F, Morotomi M. Characterization of phascolarctobacterium succinatutens sp. nov., an asaccharolytic, succinate-utilizing bacterium isolated from human feces. Appl Environ Microbiol 2012;78:511–518 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Arpaia N, Campbell C, Fan X, et al. Metabolites produced by commensal bacteria promote peripheral regulatory T-cell generation. Nature 2013;504:451–455 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Burger-van Paassen N, Vincent A, Puiman PJ, et al. The regulation of intestinal mucin MUC2 expression by short-chain fatty acids: Implications for epithelial protection. Biochem J 2009;420:211–219 [DOI] [PubMed] [Google Scholar]
- 42.Willemsen LE, Koetsier MA, van Deventer SJ, van Tol EA. Short chain fatty acids stimulate epithelial mucin 2 expression through differential effects on prostaglandin E(1) and E(2) production by intestinal myofibroblasts. Gut 2003;52:1442–1447 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Wong JM, de Souza R, Kendall CW, et al. Colonic health: Fermentation and short chain fatty acids. J Clin Gastroenterol 2006;40:235–243 [DOI] [PubMed] [Google Scholar]
- 44.Boets E, Gomand SV, Deroover L, et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: A stable isotope study. J Physiol 2017;595:541–555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hudson BD, Tikhonova IG, Pandey SK, et al. Extracellular ionic locks determine variation in constitutive activity and ligand potency between species orthologs of the free fatty acid receptors FFA2 and FFA3. J Biol Chem 2012;287:41195–41209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tolhurst G, Heffron H, Lam YS, et al. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein-coupled receptor FFAR2. Diabetes 2012;61:364–371 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Parker EA, Roy T, D'Adamo CR, Wieland LS. Probiotics and gastrointestinal conditions: An overview of evidence from the Cochrane Collaboration. Nutrition 2018;45:125–134 e111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hidalgo-Cantabrana C, Delgado S, Ruiz L, et al. Bifidobacteria and Their Health-Promoting Effects. Microbiol Spectr 2017;5 DOI: 10.1128/microbiolspec.BAD-0010-2016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Mazmanian SK, Liu CH, Tzianabos AO, Kasper DL. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005;122:107–118 [DOI] [PubMed] [Google Scholar]
- 50.Hsiao EY, McBride SW, Hsien S, et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013;155:1451–1463 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Maes M, Mihaylova I, Leunis JC. Increased serum IgA and IgM against LPS of enterobacteria in chronic fatigue syndrome (CFS): Indication for the involvement of gram-negative enterobacteria in the etiology of CFS and for the presence of an increased gut-intestinal permeability. J Affect Disord 2007;99:237–240 [DOI] [PubMed] [Google Scholar]
- 52.Saffari F, Dalfardi MS, Mansouri S, Ahmadrajabi R. Survey for correlation between Biofilm formation and virulence determinants in a collection of pathogenic and fecal enterococcus faecalis isolates. Infect Chemother 2017;49:176–183 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.