

Abstract
An intronless endoglucanase from thermotolerant Aspergillus fumigatus DBINU-1 was cloned, characterized and expressed in the yeast Kluyveromyces lactis. The full-length open reading frame of the endoglucanase gene from A. fumigatus DBiNU-1, designated Cel7, was 1383 nucleotides in length and encoded a protein of 460 amino acid residues. The predicted molecular weight and the isoelectric point of the A. fumigatus Cel7 gene product were 48.19 kDa and 5.03, respectively. A catalytic domain in the N-terminal region and a fungal type cellulose-binding domain/module in the C-terminal region were detected in the predicted polypeptide sequences. Furthermore, a signal peptide with 20 amino acid residues at the N-terminus was also detected in the deduced amino acid sequences of the endoglucanase from A. fumigatus DBiNU-1. The endoglucanase from A. fumigatus DBiNU-1 was successfully expressed in K. lactis, and the purified recombinant enzyme exhibited its maximum activity at pH 5.0 and 60 °C. The enzyme was very stable in a pH range from 4.0 to 8.0 and a temperature range from 30 to 60 °C. These features make it suitable for application in the paper, biofuel, and other chemical production industries that use cellulosic materials.
Keywords: Aspergillus fumigatus, Cellulosic material, Gene cloning, Kluyveromyces lactis, Thermotolerant fungus
Introduction
Cellulose, the most abundant organic and renewable biomass, has been considered a good candidate for the production of alternative biofuels and chemicals.1 Cellulose consists of a linear chain of several glucose units that are linked by β (1→4) glycosidic bonds.2, 3, 4 It is one of the major structural constituents of plant cell walls and is commonly found in the stalks, stems, trunks and all the woody portions of the plant body.5 Conversion of cellulose into fermentable sugar, i.e., glucose, requires the synergistic action of many cellulolytic enzymes known as cellulases. Cellulases refer to a multienzyme complex composed of endoglucanase (endo-1,4-β-glucanase, EC 3.2.1.4), exoglucanase (exo-1,4-β-glucanase, EC 3.2.1.91), and cellobiase (β-glucosidase, EC 3.2.1.21).6, 7 The enzymes are widely found in many organisms, such as actinomycetes, bacteria, fungi and plants. Among the three major groups of cellulases, endoglucanase is considered to be the most important enzyme because its action results in a non-reducing end that is necessary for the action of exoglucanase and cellobiase.8
There are several sources of endoglucanase such as bacteria, fungi and plant,7, 8, 9, 10 but fungal endoglucanase is the major source of enzyme for industrial applications. Although several endoglucanases have been cloned, identified and characterized, most efforts have focused on the enzymes from the filamentous fungi, such as Aspergillus usamii,7 A. kawachii,12 Trichoderma reesei,13 A. niger,14 Volvariella volvacea,15, 16 Penicillium canescens,11 P. funiculosum17 and Verticillium dahlia.18 Nierman et al.19 reported that there are approximately eighteen different genes that encode endoglucanases in the entire sequence of the A. fumigatus Af293 genome. Most recently, the endoglucanase gene belonging to the family AA9 (formerly known as GH61) from the alkali-tolerant A. fumigatus MKU1, which is isolated from paper and pulp industry wastes, has also been cloned and sequenced.3 Das et al.20 reported the kinetic properties of the purified halostable endoglucanase from a culture extract of A. fumigatus ABK9, and the authors demonstrated that the calculated molecular weight of the enzyme was approximately 56.3 kDa. The enzyme displayed its maximum activity at a temperature of 50 °C and a pH value of 5.0. This enzyme is highly stable in a pH range from 4.0 to 7.0 and at NaCl concentrations up to 3.0 M. Xu et al.21 reported the expression and secretion of fungal endoglucanase II and chimeric cellobiohydrolase I in the oleaginous yeast Lipomyces starkeyi and the authors demonstrated that the engineered L. starkeyi was capable of expressing and secreting both enzymes with high activity toward cellulose.
Currently, Kluyveromyces lactis, an ascomycetous budding yeast belonging to the endoascomycetales, is considered as one of the most favored systems for the expression of several heterologous proteins because it has several advantages including protein folding, protein processing and posttranslational modification. It has been reported that K. lactis produces and secretes heterologous proteins at a higher level than Saccharomyces cerevisiae. It also secretes few native proteins and a secreted recombinant product is therefore easily recovered and purified from the culture medium.22, 23 Most importantly, K. lactis can be grown in inexpensive media. Furthermore, it has been asserted as a “Generally Recognized As Safe” microorganism by the US FDA and can thus be applied in several industries such as food, beverage, flavor, vitamin and feed production.24
In the current study, the cloning and molecular characterization of the gene encoding endoglucanase from the thermotolerant fungus A. fumigatus DBiNU-1 were investigated. In addition, the heterologous expression of the A. fumigatus endoglucanase gene in yeast K. lactis GG799 and the enzymatic properties of the gene product were also described. We demonstrated in the current study that the endoglucanase from A. fumigatus DBiNU-1 is a thermostable enzyme.
Materials and methods
Strains, plasmid vectors and culture conditions
The thermotolerant fungus A. fumigatus DBiNU-1 used in this work was obtained from Department of Biology, Faculty of Science, Naresuan University. It was isolated from a soil sample collected in the northern part of Thailand, and was identified by morphological analysis and DNA sequencing of the internal transcribed spacer (ITS) and large subunit (LSU) of rDNA region. It was used as a source for genomic DNA isolation. The fungus was maintained on potato dextrose agar (PDA) and stored at 4 °C. K. lactis GG799 (New England Biolabs, US) was used as a host system for protein expression, while Escherichia coli DH5α was used as a host system for gene cloning. The pGEM-T Easy (Promega, Madison, WI, USA) and pKLAC2 vectors were used for gene cloning in E. coli and for gene expression in K. lactis, respectively.
Genomic DNA extraction
A. fumigatus DBiNU-1 was cultured in YPD medium on a rotary incubator shaker (220 rpm) at 30 °C for 24 h. The fungal mycelia were harvested by filtration and washed twice with sterile distilled water. Genomic DNA was isolated from the fungal mycelia using the GF-1 nucleic acid extraction kit (Vivantis, USA), and the purified genomic DNA was used as a DNA template for gene amplification using the polymerase chain reaction (PCR) technique.
PCR amplification, cloning and nucleotide sequencing of the endoglucanase gene from A. fumigatus DBiNU-1
Based on the alignment of the multiple amino acid sequences of the endoglucanases from A. fumigatus MKU1 (AFJ54162), A. niger ATCC10574 (ADC84000) and A. oryzae KBN616 (BAA22588), the conserved regions at the N-terminus (GALYLSEM) and C-terminus (IRWGDIG) were found. The design of the degenerate primers Cel7asper_1F (5′-GGNGCNYTNWSNGARATG-3′) and Cel7asper_1R (5′-NCCDATRTCNCCCCANCKDAT-3′) were based on these two regions, and the primers were used to amplify a partial sequence of the endoglucanase gene from A. fumigatus DBiNU-1. Since the partial sequence of the endoglucanase gene from A. fumigatus DBiNU-1 was highly homologous to that from A. fumigatus MKU1 (JQ796069), the amplification of the full length open reading frame (ORF) of the endoglucanase gene from A. fumigatus DBiNU-1 was performed using specific primers Cel7full-F (5′-ATGGACTCCAAAAGAGGCGTCGT-3′) and Cel7full-R (5′-CTACAGACACTGAGAGTACCACGGA-3′), which were synthesized based on the nucleotide sequences of the endoglucanase gene from A. fumigatus MKU1. The PCR reaction mixture (25 μl) consisted of 2× KOD Xtreme™ PCR buffer, 2 mM dNTPs, 10 pmol/μl each forward and reverse primer, 1.0 U/μl KOD Xtreme™ Hot Start DNA polymerase, and 100 ng/μl template DNA, as per the manufacturer's instructions (Merck, Germany). The template DNA was first denatured at 95 °C for 3 min. Thirty cycles of the amplification reaction were then performed using of the following conditions: denaturation for 1 min at 94 °C, annealing for 1 min at 50 °C and extension for 2 min at 72 °C. A final extension was performed at 72 °C for 7 min. The PCR product was visualized on 0.7% agarose gel, extracted and purified from the gel using the NucleoSpin Extract II Kit (Macherey-Nagel, Germany). The Rapid DNA Ligation and Transformation Kit (Fermentas, USA) was used to clone the purified PCR product into the pGEM T-easy vector and to transform the resulting plasmid into E. coli DH5α. Transformants were screened at 37 °C on LB medium containing 100 μg/ml ampicillin. The recombinant plasmid DNA was isolated from the selected transformants and subjected to DNA sequencing by the Sanger method using the MegaBACE 1000 automated DNA sequencer (Pharmacia Biotech, Sweden). The nucleotide and the deduced amino acid sequences of the endoglucanase gene product from A. fumigatus DBiNU-1 were analyzed using GENETYX (Software Development, Tokyo, Japan), while a homology analysis was carried out using the FASTA and BLAST program in the GenBank database.
Construction of the expression vector and transformation of K. lactis
The ORF of the endoglucanase gene from A. fumigatus DBiNU-1 was amplified by PCR using the following primer pairs: CEL7PKLAC_F: 5′ CGCTCGAGAAAAGAATGG ACTCCAAAAGAGGC-3′ (the underline bases represent the XhoI restriction site) and CEL7PKLAC_R: 5′-CCGGAATTCCTACAGACACTGAGAGTACCA-3′ (the underline bases represent the EcoRI restriction site). All PCR primers were synthesized by Integrated DNA technology (IDT, Singapore). The PCR reaction mixture (25 μl) consisted of 2× KOD Xtreme™ PCR buffer, 2 mM dNTPs, 10 pmol/μl each forward and reverse primer, 1.0 U/μl KOD Xtreme™ Hot Start DNA polymerase and 20 ng/μl template DNA, as per the manufacturer's instruction (Merck, Germany). The thermocycling conditions consisted of a 94 °C step for 2 min, followed by 35 cycles of successive incubation at 94 °C for 30 s, 60 °C for 1 min and 72 °C for 2 min. After the thermocycling, a final extension was performed at 72 °C for 7 min. To construct the expression cassette pKLAC2-Cel7, the purified amplified fragment was digested with XhoI and EcoRI and then ligated into the XhoI/EcoRI site of the K. lactis expression vector pKLAC2. The accuracy of the nucleotide sequence of the cloned endoglucanase gene was confirmed by DNA sequencing as described above.
The transformation of the expression cassette into K. lactis GG799 was performed using lithium acetate with the heat shock method.25 To integrate the cloned endoglucanase gene into the LAC4 locus of the K. lactis chromosome, the expression cassette pKLAC2-Cel7 was linearized with BstXI before transformation. The transformants harboring the pKLAC2-Cel7 plasmid were screened and selected at 30 °C using yeast carbon base (YCB) agar medium supplemented with 5 mM acetamide. The selected transformants were further analyzed on YPD agar medium containing 1% carboxymethyl cellulose (CMC) (Fluka, Germany).
Detection of the recombinant endoglucanase using 1% Congo red
The selected transformants were grown in YPGal medium at 30 °C in an incubator shaker (200 rpm) for 24 h. Thereafter, 50 μl of the yeast culture was transferred onto the YPD agar medium that contained 1% CMC and incubated at 30 °C. Sterile distilled water was used as a control in this study. After 7 days of incubation, the agar medium was flooded with 1% Congo red and subsequently incubated at room temperature (RT) for 15 min. The agar medium was rinsed twice with sterile distilled water and then washed with a 1 M NaCl solution for 15 min. The clear zone that appeared on the agar medium was monitored and photographed.
SDS-polyacrylamide gel electrophoresis and zymogram analysis of the recombinant endoglucanase
The selected transformant was grown in YPD liquid medium supplemented with 1% CMC at 30 °C in an incubator shaker (200 rpm) for 7 days. The culture supernatant was collected by centrifugation at 13,000 rpm and 4 °C for 15 min, and the endoglucanase activity in the clear supernatant (crude enzyme) was assayed. The protein concentration in the culture supernatant was measured by the Bradford method26 using bovine serum albumin (BSA) as the standard. For SDS-polyacrylamide gel electrophoresis (SDS-PAGE), the crude enzyme was prepared based on the method described by Eriksson and Pettersson27 and Béguin,28 using a sample buffer containing 2% SDS, 0.5 M Tris–HCl (pH 6.8), 10% glycerol and 0.01% bromophenol blue. The SDS-PAGE was carried out based on the standard method described by Li,29 using 30 μg of protein sample and a 12% polyacrylamide gel, and the separation of the protein bands in the gel was visualized by Coomassie brilliant blue R-250 (Sigma) staining.
The zymogram analysis was based on the method described by Grigorevski-Lima et al.,30 and was carried out to determine the enzyme activity. The crude enzyme was subjected to SDS-PAGE using a 12% polyacrylamide gel that contained 1% CMC as substrate. After electrophoresis, the gel was soaked in 2.5% (v/v) Triton X-100 for 2 h and then incubated in 50 mM sodium acetate buffer (pH 5.0) for 1 h at RT in order to remove the SDS and renature the enzyme. The resulting gel was washed with 50 mM sodium acetate buffer (pH 5.0) at 45 °C for 30 min, stained with 1% (w/v) Congo red dye solution at RT for 30 min, and subsequently destained with 1 N NaCl until the excess dye was removed. The clear zone, which corresponded to enzyme activity, was monitored and documented with a gel documentation system (Bio-Rad, Singapore).
Purification of protein using an aqueous two-phase system
The recombinant endoglucanase was purified using an aqueous two-phase system.31 Various concentrations of polyethylene glycol 6000 (PEG 6000) and dextran T70 were used in this system. To prepare the two-phase system, a stock solution composed of 20% (w/w) dextran and 40% (w/w) PEG and the enzyme solution were mixed and left to settle for at least 1.5 h. Protein samples in the top and bottom phases were taken, and the enzyme activity in both phases was further determined.
The activity of the purified recombinant endoglucanase was assayed using the method described by Li et al.,16 using CMC as substrate. One unit (U) of enzyme activity was defined as the amount of enzyme capable of releasing 1 μmol of reducing sugar from CMC per minute under the assay conditions. d-Glucose was used as the standard.
Effect of pH on enzyme activity and stability
The effect of pH on the activity of the purified recombinant endoglucanase was evaluated using the standard method described by Li et al.,16 with different buffers including sodium acetate buffer (pH 3.0–6.0) and phosphate buffer (pH 7.0–11.0). The stability of the purified endoglucanase at different pH values was evaluated by incubating the purified enzyme at 50 °C for 1 h in the same buffers in the pH range from 3.0 to 11.0 and measuring the remaining enzyme activity under standard conditions.
Effect of temperature on enzyme activity and stability
The effect of temperature on the activity of the purified recombinant endoglucanase was evaluated at the optimum pH at temperatures ranging from 30 to 80 °C. The thermal stability of the purified enzyme was evaluated by preincubating the enzyme solution in 50 mM sodium acetate buffer (pH 5.0) at various temperatures (30–80 °C) for 1 h. The remaining enzyme activity was assayed under standard conditions.
Results
Cloning and nucleotide sequencing analysis of the endoglucanase gene from A. fumigatus DBiNU-1
The ORF of the gene encoding the endoglucanase in A. fumigatus DBiNU-1 was amplified by PCR using a genomic DNA template that was isolated from A. fumigatus DBiNU-1 and specific primers whose design was based on the nucleotide sequences of the endoglucanase gene from A. fumigatus MKU1, as described in the Materials and methods section. A PCR product of approximately 1.3 kb was detected after agarose gel electrophoresis analysis, and it was purified and cloned, and the resulting vector was transformed into E. coli DH5α. The recombinant plasmid DNA that was isolated from the selected transformants was then subjected to DNA sequencing. As found in the present study, the complete nucleotide sequence of the endoglucanase gene from A. fumigatus DBiNU-1, designated Cel7, was 1,383 nucleotides in length and encoded a polypeptide of 460 amino acid residues (Fig. 1). The predicted molecular weight and the isoelectric point of the A. fumigatus Cel7 gene product were found to be 48.19 kDa and 5.03, respectively. Based on the sequence similarities in the carbohydrate-active enzyme (CAZy) database (http://www.cazy.org/), the deduced amino acid sequence of the endoglucanase gene product from A. fumigatus DBiNU-1 was classified into glycoside hydrolase family 7 (GH7).32, 33, 34 These comprise two domains, i.e., a catalytic domain at the N-terminus, which contains the glycosyl hydrolase family 7 active-site residues (amino acids at positions 22–398), and a fungal type cellulose-binding domain/module (CBD/CBM) at the C-terminus (amino acids at positions 428–456), which is involved in substrate binding, as observed in most fungal cellulases.35 The prediction of the amino acid sequences by the SignalP 3.0 server (http://www.cbs.dtu.dk/services/SignalP/) revealed the signal peptide (MDSKRGVVAAVLALLPLVSA) at the N-terminal region and a signal peptide cleavage site between Ala20 and Gln21.
Fig. 1.

Nucleotide and deduced amino acid sequences of an intronless endoglucanase from thermotolerant A. fumigatus DBiNU-1. The signal peptide from Met1 to Ala20 is underlined. The conserved amino acid regions, GALYLSEM and IRWGDIG, are shown in bold letters. The putative catalytic amino acid residues Asp191 and Glu369 are shown in the boxes.
The homology analysis of the deduced amino acid sequence showed that the endoglucanase from A. fumigatus DBiNU-1 shared 98%, 74%, 59%, 51%, 46% and 34% identity with the GH7 endoglucanases from A. fumigatus KMU1 (AFJ54162), A. terreus MS-31 (ADR78837), T. reesei M5 (ADM08177), A. oryzae KBN616 (BAA22589), A. nidulans FGSCA4 (EAA63386), and Fusarium oxysporum (AAA65586), respectively (Fig. 2). The putative catalytic amino acid residues Asp191 and Glu369 were found to be highly conserved among the GH7 endoglucanases and are similar to those reported in the GH12 and AA9 family.3, 7 A three-dimensional structure of the predicted protein sequence of the endoglucanase from A. fumigatus DBiNU-1 was predicted using the Swiss-PdbViewer (http://www/expasy.org/spdbv/), and the result is shown in Additional information section, Fig. 1. The protein structure was observed at the tertiary level, and two completely distinct structures corresponding to the catalytic domain and CBM1 domain were detected.
Fig. 2.

Alignments between the A. fumigatus DBiNU-1 endoglucanase sequence and the sequences of other fungal endoglucanases from GH7. The putative catalytic amino acid residues Asp191 and Glu369 are shown in the boxes.
Expression of the endoglucanase gene from A. fumigatus DBiNU-1 in K. lactis GG799
The expression vector pKLAC2-Cel7 was constructed and transformed into K. lactis GG799, and the transformants were selected using YCB agar medium, as described in the Materials and methods section. The selected transformants were further analyzed on YPD agar medium containing 1% CMC. Transformants harboring the pKLAC2-Cel7 exhibited a clear zone on agar plates after Congo red staining, suggesting that the cloned endoglucanase gene from A. fumigatus DBiNU-1 was expressed in the K. lactis (Fig. 3).
Fig. 3.

The activity of the endoglucanase from A. fumigatus DBiNU-1 that was expressed in the yeast K. lactis, as determined using YPD agar medium containing 1% CMC.
The crude enzyme in the cell-free supernatant was analyzed by SDS-PAGE using a 12% polyacrylamide gel, and the resolved proteins were visualized using Coomassie brilliant blue R-250 staining. As shown in Fig. 4A, a protein band of approximately 48 kDa that corresponded to the predicted molecular weight of the endoglucanase from A. fumigatus DBiNU-1 was detected. The endoglucanase activity was also determined based on the degradation of CMC in the polyacrylamide gel using a zymogram analysis. A protein band of approximately 48 kDa that coincided with the band that was observed in the SDS-PAGE analysis was detected on a polyacrylamide gel after Congo red staining (Fig. 4A), suggesting that this protein band was the recombinant endoglucanase from A. fumigatus DBiNU-1. The endoglucanase activity in the cell-free supernatant from K. lactis harboring the pKLAC2-Cel7 plasmid was measured, and the highest enzyme activity was found to be 0.80 U/ml with a specific activity of 3.08 U/mg protein. There was no endoglucanase activity in the cell-free supernatant from the negative control (K. lactis transformed with pKLAC2) under the same cultural conditions.
Fig. 4.

SDS-PAGE and zymogram analysis of the endoglucanase (A) and the purified recombinant endoglucanase (B) from A. fumigatus DBiNU-1. The protein samples (30 μg) were loaded on the gel and subjected to electrophoresis. (A) M, protein marker; 1, the cell-free supernatant from K. lactis GG799; 2, the cell-free supernatant from K. lactis harboring pKLAC2-Cel7; 3, zymogram analysis of the endoglucanase after Congo red staining. (B) M, protein marker; 1, crude recombinant endoglucanase; 2, purified recombinant endoglucanase.
Purification and characterization of the purified recombinant endoglucanase
The recombinant endoglucanase from the K. lactis strain that harbored pKLAC2-Cel7 was purified using the aqueous two-phase system, as described in the Materials and methods section. A maximum enzyme activity and specific activity of 1.57 U/ml and 15.39 U/mg protein, respectively, were achieved using the two-phase system that contained 7% PEG and 5% dextran. Fig. 4B shows the SDS-PAGE analysis of the purified recombinant endoglucanase that was obtained from aqueous two-phase system. Most of the protein band at approximately 48 kDa, which corresponded to the monomer of the Cel7 gene product from A. fumigatus DBiNU-1, was observed, while those low-molecular weight proteins that were contained in the cell-free supernatant were mostly removed.
The effect of pH on the activity and stability of the purified recombinant endoglucanase was evaluated, and the results are summarized in Fig. 5A. The purified recombinant endoglucanase displayed its highest activity at pH 5.0, and more than 80% of its original activity remained in the pH range from 4.0 to 8.0.
Fig. 5.

The effect of pH (A) and temperature (B) on the activity and stability of the purified recombinant endoglucanase from A. fumigatus DBiNU-1. The maximum enzyme activity (1.57 U/ml) assayed under the standard condition (60 °C, pH 5.0) was taken as a control and recorded as 100%.
The effect of temperature on the recombinant endoglucanase activity and stability is illustrated in Fig. 5B. The maximal activity of the purified recombinant endoglucanase at pH 5.0 was attained at a temperature of 60 °C. The activity of the purified enzyme remarkably decreased at temperatures above 60 °C. More than 80% of the original activity of the purified enzyme remained in a temperature range from 30 to 60 °C, whereas only half of the original activity was detected at 70 °C.
Discussion
Based on the whole genome sequence of A. fumigatus Af293, 18 different genes encoding endoglucanases have been reported.19 They belong to various families, such as GH5, GH7, GH12, GH45, AA9 (formerly known as GH61), and GH81, based on their amino acid sequence similarities. According to a nucleotide sequence analysis, three endoglucanase genes from A. fumigatus are intronless (one from GH7 and two from AA9), and two out of these three known intronless endoglucanase genes are predicted to have a cellulose-binding domain/module in the C-terminal region. In the current study, a gene that encodes an intronless endoglucanase in A. fumigatus DBiNU-1 was cloned, and its nucleotide and putative amino acid sequences were analyzed using GENETYX (Software Development, Tokyo, Japan). The results showed that the complete sequence of the gene that encodes the endoglucanase in A. fumigatus DBiNU-1, designated Cel7, was 1383 bp in length and encoded a polypeptide of 460 amino acid residues (Fig. 1). Based on the amino acid sequence similarity in the CAZy database (http://www.cazy.org), the endoglucanase from A. fumigatus DBiNU-1 was classified into the GH7 family,32, 33, 34 similar to endoglucanases from Aspergillus spp. such as A. terreus MS-31, A. oryzae KBN616, and A. nidulans FGSCA4, and endoglucanases from T. reesei M5 and Penicillium sp. C7. According to the Pfam tool (http://pfam.sanger.ac.uk/), two domains, i.e., a catalytic domain in the N-terminal region (amino acids from position 22 to 398) and a fungal type CBD/CBM in the C-terminal region (amino acids from position 428 to 456), were found in the deduced amino acid sequences of the A. fumigatus Cel7 gene product. As demonstrated by Sugimoto,36 the CBD/CBM is important for the binding of cellulose. Cellulases that lack the CBD/CBM exhibited a reduction in cellulose degradation. It should be noted from the current study that a signal peptide containing 20 amino acid residues was detected in the N-terminal region, suggesting that the endoglucanase from A. fumigatus DBiNU-1 is an extracellular enzyme because the same feature has been found in an intronless endoglucanase from A. fumigatus MKU1.3 In addition to a signal peptide, two regions, GALYLSEM and IRWGDIG, that are highly conserved in all fungal endoglucanases in the GH7 family were detected in the endoglucanase from A. fumigatus DBiNU-1. This finding also verified that the A. fumigatus Cel7 gene product was a member of the fungal GH7 family. No putative N-glycosylation site (N-W-T) was found in the predicted protein sequences, suggesting that the endoglucanase from A. fumigatus DBiNU-1 is not an N-glycosylated protein, which is distinct from the reported case of the endoglucanase from A. usamii E001.7
The predicted molecular weight of the endoglucanase from A. fumigatus DBiNU-1, as confirmed by SDS-PAGE analysis, was approximately 48 kDa, which was distinct from other endoglucanases from other Aspergillus strains, e.g., A. terreus M11 (25 kDa),37 A. awamori VTCC-F099 (32 kDa),38 A. fumigatus KMU1 (35 kDa),3 A. oryzae AG1 (45 kDa),39 A. usamii E001 (24 kDa),7 and A. fumigatus ABK9 (56 kDa).20 Many glycoside hydrolases such as GH7, GH12, GH16 and AA9 have a modular structure that consists of a catalytic domain and a substrate-binding domain.40 The Swiss-PdbViewer (http://www/expasy.org/spdbv) was used to predict the three-dimensional structure of the putative amino acid sequence of the endoglucanase from A. fumigatus DBiNU-1 (Additional information section, Fig. 1). The GH7 endoglucanase from this thermotolerant fungus exhibited an arrangement of catalytic residues and a fold that are similar to those of the β-1,3-glucanases and β-1,3-1,4-glucanases of family 16.41
The aqueous two-phase system has been widely used to purify several enzymes. Compared with conventional purification methods, this technique provides a high yield of recovery with high level of enzyme activity and stability.31 In the current study, the recombinant endoglucanase from A. fumigatus DBiNU-1 that was expressed in K. lactis with the pKLAC2-Cel7 vector was purified using the aqueous two-phase system. A maximum enzyme activity and specific activity of 1.57 U/ml and 15.39 U/mg protein, respectively, were achieved; these values were approximately 2- and 5-fold greater than the corresponding original activities. The optimum pH of the activity of the purified recombinant endoglucanase from A. fumigatus DBiNU-1 was 5.0, and the enzyme was highly stable in a pH range from 4.0 to 8.0 (Fig. 5A). The purified recombinant endoglucanase also exhibited its highest activity at 60 °C, and it over 80% of its original activity in a temperature range from 30 to 60 °C (Fig. 5B). The results of the current study were in good agreement with those reported for most of the endoglucanase from Aspergillus strains, such as A. terreus DSM 826,42 A. awamori VTCC-F099,38 A. usamii E0017 and A. fumigatus ABK9.20 However, a high thermal stability (70 °C) was also reported for endoglucanases from T. aurantiacus43 and A. fumigatus MKU1.3 The differences in thermal stability indicated the different molecular properties of the enzymes, including the bonding that stabilizes in the structures and the conformations among the various species. It has been reported that endoglucanases with optimal activities between 55 and 80 °C are considered thermostable enzymes.44 The high thermal stability of the endoglucanase from A. fumigatus DBiNU-1 (60 °C) make this enzyme a good candidate for application in the paper industry and in saccharification processes for the production of biofuel and other chemicals using cellulosic materials.
Conflicts of interest
The authors declare that they have no conflict of interest.
Acknowledgments
The authors thank Khon Kaen University for financial support and the Fermentation Research Center for Value Added Agricultural Products (FerVAAPs) at Khon Kaen University for technical support.
Associate Editor: Rodrigo Galhardo
Footnotes
Supplementary data associated with this article can be found, in the online version, at doi:10.1016/j.bjm.2017.10.001.
Appendix A. Supplementary data
The following are the supplementary data to this article.
Supplementary Fig. 1.
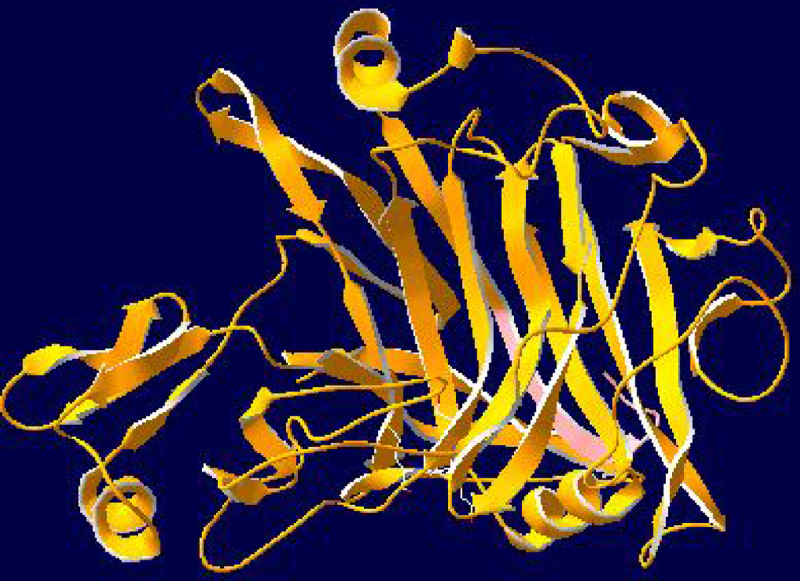
The three-dimensional structure of the predicted protein sequences of the endoglucanase from A. fumigatus DBiNU-1 using Swiss-PdbViewer V4.10 (http://www.expasy.org/spdbv/).
References
- 1.Hong J., Wang Y., Kumagai H., Tamaki H. Construction of thermotolerant yeast expressing thermostable cellulase genes. J Biotechnol. 2007;130:114–123. doi: 10.1016/j.jbiotec.2007.03.008. [DOI] [PubMed] [Google Scholar]
- 2.Makut M., Godiya E. A survey of cellulolytic mesophilic fungi in the soil environment of Keffi Metropolis, Nasarawa State, Nigeria. Afr J Microbiol Res. 2010;4:2191–2195. [Google Scholar]
- 3.Meera B., Vanitha M.C., Ramani G., Manjula A., Gunasekaran P. Cloning and expression of an intronless gene encoding endoglucanase from Aspergillus fumigatus MKU1. Indian J Biotechnol. 2011;10:480–486. [Google Scholar]
- 4.Rowell R.M. CRC press; New York, USA: 2012. Handbook of Wood Chemistry and Wood Composites. [Google Scholar]
- 5.Chaudhary N., Qazi J.I. Lignocellulose for ethanol production: a review of issues relating to bagasse as a source material. Afr J Biotechnol. 2013;10:1270–1274. [Google Scholar]
- 6.Adsul M.G., Bastawde M.G., Varma A.J., Gokhale D.V. Strain improvement of Penicillium janthinellum NCIM 1171 for increased cellulase production. Bioresour Technol. 2007;98:1467–1473. doi: 10.1016/j.biortech.2006.02.036. [DOI] [PubMed] [Google Scholar]
- 7.Shi H., Yin X., Wu M., Tang C., Zhang H., Li J. Cloning and bioinformatics analysis of an endoglucanase gene (Aucel12A) from Aspergillus usamii and its functional expression in Pichia pastoris. J Ind Microbiol Biotechnol. 2012;39:347–357. doi: 10.1007/s10295-011-1039-z. [DOI] [PubMed] [Google Scholar]
- 8.Wang J., Gao G., Li Y. Cloning, expression, and characterization of a thermophilic endoglucanase, AcCel12B from Acidothermus cellulolyticus 11B. Int J Mol Sci. 2015;16:25080–25095. doi: 10.3390/ijms161025080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wei K.S.C., Teoh T.C., Koshy P., Salmah I., Zainudin A. Cloning, expression and characterization of the endoglucanase gene from Bacillus subtilis UMC7 isolated from the gut of the indigenous termite Macrotermes malaccensis in Escherichia coli. Electron J Biotechnol. 2015;18:103–109. [Google Scholar]
- 10.del Campillo E. Multiple endo-1,4-beta-D-glucanase (cellulase) genes in Arabidopsis. Curr Top Dev Biol. 1999;46:39–61. doi: 10.1016/s0070-2153(08)60325-7. [DOI] [PubMed] [Google Scholar]
- 11.Chulkin A.M., Loginov D.S., Vavilova E.A. Cloning of the Penicillium canescens endo-1,4-β-glucanase gene egl3 and the characterization of the recombinant enzyme. Appl Biochem Microbiol. 2009;45:143–149. [PubMed] [Google Scholar]
- 12.Sakamoto S., Tamura G., Tto K., Ishikawa T., Iwano K., Nishiya N. Cloning and sequencing of cellulase cDNA from Asperillus kawachii and its expression in Saccharomyces cerevisiae. Curr Genet. 1995;27:435–439. doi: 10.1007/BF00311212. [DOI] [PubMed] [Google Scholar]
- 13.Sandgren M., Shaw A., Ropp T.H. The X-ray crystal structure of the Trichoderma reesei family 12 endoglucanase 3 Cel12A, at 1.9 Å resolution. J Mol Biol. 2001;308:295–310. doi: 10.1006/jmbi.2001.4583. [DOI] [PubMed] [Google Scholar]
- 14.Hong J., Tamaki H., Akiba S., Yamamoto K., Kumagai H. Cloning of a gene encoding a highly stable endo-β-1,4-glucanase from Aspergillus niger and its expression in yeast. J Biosci Bioeng. 2001;92:434–441. doi: 10.1263/jbb.92.434. [DOI] [PubMed] [Google Scholar]
- 15.Ding S.J., Ge W., Buswell J.A. Secretion purification and characterization of a recombinant Volvariella volvacea endoglucanase expressed in the yeast Pichia pastoris. Enzyme Microb Technol. 2002;31:621–626. [Google Scholar]
- 16.Li J.F., Tang C.D., Shi H.L., Wu M.C. Cloning and optimized expression of a neutral endoglucanase gene (ncel5A) from Volvariella volvacea WX32 in Pichia pastoris. J Biosci Bioeng. 2011;111:537–540. doi: 10.1016/j.jbiosc.2011.01.002. [DOI] [PubMed] [Google Scholar]
- 17.Castro A.M., Caryalho M.L.A., Leite S.G.F., Pereira N., Jr. Cellulases from Penicillium funiculosum: production, properties and application to cellulose hydrolysis. J Ind Microbiol Biotechnol. 2010;37:151–158. doi: 10.1007/s10295-009-0656-2. [DOI] [PubMed] [Google Scholar]
- 18.Eboigbe L., Tzima A.K., Paplomatas E.J., Typas M.A. The role of the β-1,6-endoglucanase gene vegB in physiology and virulence of Verticillium dahliae. Phytopathol Mediterr. 2014;53:94–107. [Google Scholar]
- 19.Nierman W.C., Pain A., Anderson M.J. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature. 2005;438:1151–1156. doi: 10.1038/nature04332. [DOI] [PubMed] [Google Scholar]
- 20.Das A., Jana A., Paul T. Thermodynamics and kinetic properties of halostable endoglucanase from Aspergillus fumigatus ABK9. J Basic Microbiol. 2014;54:S142–S151. doi: 10.1002/jobm.201300350. [DOI] [PubMed] [Google Scholar]
- 21.Xu Q., Knoshaug E.P., Wang W. Expression and secretion of fungal endoglucanase II and chimeric cellobiohydrolase I in the oleaginous yeast Lipomyces starkeyi. Microb Cell Fact. 2017;16:126. doi: 10.1186/s12934-017-0742-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Van der Vlugt-Bergmans C., Van Ooyen A. Expression cloning in Kluyveromyces lactis. Biotechnol Tech. 1999;13:87–92. [Google Scholar]
- 23.Schaffrath R., Breunig K.D. Genetics and molecular physiology of the yeast Kluyveromyces lactis. Fungal Genet Biol. 2000;30:173–190. doi: 10.1006/fgbi.2000.1221. [DOI] [PubMed] [Google Scholar]
- 24.Bonekamp J.F., Oosterom J. On the safety of Kluyveromyces lactis – a review. Appl Microbiol Biotechnol. 1994;41:1–3. [Google Scholar]
- 25.Ito H., Fukuda Y., Murata K., Kimura A. Transformation of intact yeast cells treated with alkali cations. J Bacteriol. 1983;153:163–168. doi: 10.1128/jb.153.1.163-168.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Bradford M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1006/abio.1976.9999. [DOI] [PubMed] [Google Scholar]
- 27.Eriksson K.E., Pettersson B. A zymogram technique for the detection of carbohydrases. Anal Biochem. 1973;56:618–620. doi: 10.1016/0003-2697(73)90233-9. [DOI] [PubMed] [Google Scholar]
- 28.Béguin P. Detection of cellulase activity in polyacrylamide gels using Congo red-stained agar replicas. Anal Biochem. 1983;131:333–336. doi: 10.1016/0003-2697(83)90178-1. [DOI] [PubMed] [Google Scholar]
- 29.Li S.F.Y. Elsevier; Singapore: 1992. Capillary Electrophoresis: Principles Practice and Applications. [Google Scholar]
- 30.Grigorevski-Lima A., Da Vinha F., Souza D. Aspergillus fumigatus thermophilic and acidophilic endoglucanases. Appl Biochem Biotechnol. 2009;155:18–26. doi: 10.1007/s12010-008-8482-y. [DOI] [PubMed] [Google Scholar]
- 31.Asenjo J.A., Andrews B.A. Aqueous two-phase systems for protein separation: a perspective. J Chromatogr A. 2011;1218:8826–8835. doi: 10.1016/j.chroma.2011.06.051. [DOI] [PubMed] [Google Scholar]
- 32.Naumoff D.G. Hierarchical classification of glycoside hydrolases. Biochemistry. 2011;76:622–635. doi: 10.1134/S0006297911060022. [DOI] [PubMed] [Google Scholar]
- 33.Lombard V., Golaconda Ramulu H., Drula E., Coutinho P.M., Henrissat B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014;42:490–495. doi: 10.1093/nar/gkt1178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Terrapon N., Lombard V., Gilbert H.J., Henrissat B. Automatic prediction of polysaccharide utilization loci in Bacteroidetes species. Bioinformatics. 2015;31:647–655. doi: 10.1093/bioinformatics/btu716. [DOI] [PubMed] [Google Scholar]
- 35.Marchler-Bauer A., Derbyshire M.K., Gonzales N.R. CDD: NCBI's conserved domain database. Nucleic Acids Res. 2014;34:222–226. doi: 10.1093/nar/gku1221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sugimoto N. Shinshu University; Japan: 2012. Functional Analysis and Application of Fungal Cellulose-Binding Domains. (Ph.D. thesis) [Google Scholar]
- 37.Gao J., Weng H., Xi Y., Zhu D., Han S. Purification and characterization of a novel endo-β-1, 4-glucanase from the thermoacidophilic Aspergillus terreus. Biotechnol Lett. 2008;30:323–327. doi: 10.1007/s10529-007-9536-x. [DOI] [PubMed] [Google Scholar]
- 38.Nguyen V.T., Quyen D.T. Purification and properties of a novel thermoactive endoglucanase from Aspergillus awamori VTCC-F099. Aust J Basic Appl Sci. 2010;4:6211–6216. [Google Scholar]
- 39.Youssef G.A. Physiological studies of cellulase complex enzymes of Aspergillus oryzae and characterization of carboxymethyl cellulase. Afr J Microbiol Res. 2011;5:1311–1321. [Google Scholar]
- 40.Davies G., Henrissat B. Structures and mechanisms of glycosyl hydrolases. Structure. 1995;3:853–859. doi: 10.1016/S0969-2126(01)00220-9. [DOI] [PubMed] [Google Scholar]
- 41.Divne C., Ståhlberg J., Reinikainen T. The three-dimensional crystal structure of the catalytic core of cellobiohydrolase I from Trichoderma reesei. Science. 1994;265:524–528. doi: 10.1126/science.8036495. [DOI] [PubMed] [Google Scholar]
- 42.Elshafei A.M., Hassan M.M., Haroun B.M., Abdel-Fatah O.M., Atta H.M., Othman A.M. Purification and properties of an endoglucanase of Aspergillus terreus DSM 826. J Basic Microbiol. 2009;49:426–432. doi: 10.1002/jobm.200800227. [DOI] [PubMed] [Google Scholar]
- 43.Voutilainen S.P., Puranen T., Siika-aho M. Cloning, expression, and characterization of novel thermostable family 7 cellobiohydrolases. Biotechnol Bioeng. 2008;101:515–528. doi: 10.1002/bit.21940. [DOI] [PubMed] [Google Scholar]
- 44.Maheshwari R., Bharadwaj G., Bhat M.K. Thermophilic fungi: their physiology and enzymes. Microbiol Mol Biol R. 2000;64:461–488. doi: 10.1128/mmbr.64.3.461-488.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]