

Abstract
Interest in connectivity has increased in the aquatic sciences, partly because of its relevance to the Clean Water Act. This paper has two objectives: (1) provide a framework to understand hydrological, chemical, and biological connectivity, focusing on how headwater streams and wetlands connect to and contribute to rivers; and (2) review methods to quantify hydrological and chemical connectivity. Streams and wetlands affect river structure and function by altering material and biological fluxes to the river; this depends on two factors: (1) functions within streams and wetlands that affect material fluxes; and (2) connectivity (or isolation) from streams and wetlands to rivers that allows (or prevents) material transport between systems. Connectivity can be described in terms of frequency, magnitude, duration, timing, and rate of change. It results from physical characteristics of a system, e.g., climate, soils, geology, topography, and the spatial distribution of aquatic components. Biological connectivity is also affected by traits and behavior of the biota. Connectivity can be altered by human impacts, often in complex ways. Because of variability in these factors, connectivity is not constant but varies over time and space. Connectivity can be quantified with field-based methods, modeling, and remote sensing. Further studies using these methods are needed to classify and quantify connectivity of aquatic ecosystems and to understand how impacts affect connectivity.
Keywords: connectivity, isolation, streams, tributaries, riparian areas, wetlands, geographically isolated wetlands, watersheds, Clean Water Act
INTRODUCTION
In recent decades, the aquatic sciences have expanded from studying the function and dynamics of individual ecosystems (e.g., Valiela and Teal, 1979) to developing an understanding of interactions between multiple ecosystems and the ways in which these interactions result in larger-scale landscape function (e.g., Cohen et al., 2016). For freshwater aquatic ecosystems, a typical scale and focus for such research is the watershed and resulting watershed functions. Aquatic ecosystems (e.g., streams, rivers, lakes, and wetlands) can interact because of their ability to import and export material and energy, and through their ability to alter the fluxes of these materials. A key determinant of these interactions is connectivity, which describes the degree to which components of a river system are joined, or connected, by various transport mechanisms (U.S. EPA, 2015). Here, we adopt the U.S. EPA (2015) and Naiman and Bilby (1998) terminology and define a stream and river as, respectively, either a relatively small or relatively large volume of flowing water within a visible channel, including subsurface water moving in the same direction as the surface water, and lateral flows exchanged with associated floodplain and riparian areas. We define a river system as a river and its entire drainage basin, including its river network, associated riparian areas, floodplains, alluvial aquifers, regional aquifers, connected water bodies, geographically isolated waters, and terrestrial ecosystems. A river network is defined as a hierarchical, interconnected population of channels or swales that drain water to a river. Connectivity of river systems—hydrological, chemical, and biological—is determined by characteristics of the physical landscape, climate, and the biota, as well as human impacts.
In just the last few years, numerous studies have focused on aquatic connectivity, including: (1) connectivity between wetlands (McIntyre et al., 2014; Uden et al., 2014; Hayashi et al., 2016; Leibowitz et al., 2016; Vanderhoof et al., 2016); (2) connectivity between hillslopes and streams (Jencso et al., 2010; Jencso and McGlynn, 2011; Bracken et al., 2013; Reaney et al., 2013; Janzen and McDonnell, 2015); (3) connectivity between rivers, floodplains, and floodplain wetlands (Rooney et al., 2013; Vilizzi et al., 2013; Wolf et al., 2013; Zilli and Paggi, 2013; Scott et al., 2014; Jones et al., 2015; Reid et al., 2015); (4) connectivity between wetlands occurring in non-floodplain areas and rivers (McLaughlin et al., 2014; McDonough et al., 2015; Cohen et al., 2016; Evenson et al., 2016; Fossey et al., 2016; Golden et al., 2016; Rains et al., 2016); and (5) connectivity between other river system components (Giblin et al., 2014; Harvey and Gooseff, 2015; Moore, 2015; Hauer et al., 2016). Recent research also has investigated how connectivity contributes to ecosystem services (Mitchell et al., 2013; Jordan and Benson, 2015) and its importance to watershed and aquatic ecosystem management and protection of vulnerable waters (Uden et al., 2014; Crook et al., 2015; Moore, 2015; Creed et al., 2017).
Much of the scientific interest in connectivity has been generated because of its relevance to protection of aquatic ecosystems under the U.S. Clean Water Act (CWA; Alexander et al., submitted), the goal of which is to restore and maintain the chemical, physical, and biological integrity of the nation’s waters. Thus, there has been particular interest in connectivity with respect to the effects of wetlands and non-perennial streams on the integrity of downstream waters (Leibowitz and Nadeau, 2003; Caruso, 2011; McLaughlin et al., 2014; Alexander, 2015; Marton et al., 2015; Cohen et al., 2016; Rains et al., 2016). The absence of a connection—i.e., isolation—has also been recognized as contributing to these systems (U.S. EPA, 2015). As a result, recent work has begun to focus on the full connectivity-isolation gradient of aquatic ecosystems (Cohen et al., 2016; Rains et al., 2016), including ephemeral and intermittent streams and many so-called geographically isolated wetlands (wetlands completely surrounded by upland; Leibowitz, 2015; Mushet et al., 2015; Calhoun et al., 2017).
Given the growing scientific interest in aquatic system connectivity and its relevance to the goals of the CWA, a conceptual overview of the subject and approaches towards its study is timely. This paper has two objectives. First, we provide a framework for understanding physical, chemical, and biological connectivity, specifically focusing on how aquatic ecosystems such as headwater streams and wetlands contribute to rivers and other downstream waters and the connections between those ecosystems and downstream waters. This framework integrates studies from multiple disciplines to synthesize how researchers across diverse fields approach connectivity studies. Because the study of connectivity between different types of aquatic systems is a relatively new research endeavor, the framework also draws conclusions based on logical inferences and first principles. Second, we review some of the approaches that have been used to quantify hydrological, chemical, and biological connectivity.
This paper is adapted from a chapter on this topic from an EPA report on connectivity of aquatic systems (U.S. EPA, 2015) that reviewed and synthesized more than 1,300 peer-reviewed publications on this subject. The current work is updated with recent additions to the literature. In the sections below, we address three topics: (1) the influence that streams and wetlands have on downstream waters; (2) factors that influence connectivity, including human activities; and (3) approaches for quantifying connectivity.
INFLUENCE OF STREAMS AND WETLANDS ON DOWNSTREAM WATERS
The structure and function of rivers are highly dependent on the constituent materials stored in and transported through them. Much of these materials, broadly defined here as any physical, chemical, or biological entities (including water, heat energy, sediment, wood, organic matter, nutrients, chemical contaminants, and organisms), originate outside of the river. The materials come from either the upstream river network or other components of the river system, and then are transported to the river by water movement or other mechanisms (e.g., wind, movement of biota). For example, most of the water in higher order rivers (Strahler, 1957) comes from tributaries, and not from precipitation or groundwater directly entering the river (Winter, 2007; Bukaveckas, 2009). Also, riparian areas are a critical source for allochthonous inputs of organic matter to streams, especially in headwater catchments (Tank et al., 2010). This includes large woody debris, which comes from riparian areas or hillslopes and acts to dissipate energy, trap material, and provide habitat in the stream (Sobota et al., 2006; Fritz et al., submitted). Thus, our framework for understanding how streams and wetlands contribute to downstream waters identifies alterations of material fluxes to the river as the fundamental way in which streams and wetlands affect river structure and function. This framework considers two key factors that cause this alteration of material fluxes: (1) stream and wetland functions that increase or decrease material fluxes, and (2) connectivity (or isolation) from streams and wetlands to rivers that allows (or prevents) transport of materials between the systems. Below, we discuss these two factors, as well as the spatial and temporal variability of connectivity.
Effects of Streams and Wetlands on Material Fluxes
Streams and wetlands affect the amounts and types of materials that are or are not delivered to downstream waters, ultimately contributing to the structure and function of those waters. Leibowitz et al. (2008) identified three functions, or general mechanisms of action, by which streams and wetlands influence material fluxes to downstream waters: source, sink, and refuge. U.S. EPA (2015) expanded this framework to include two additional functions: lag and transformation. These five functions (Table 1) describe the general mechanisms by which physical, chemical, and biological connections between streams and wetlands and downstream waters influence river systems. Each of these general functions are driven by specific mechanisms; e.g., adsorption, chelation, and soil biogeochemical reactions are the mechanisms that drive the sink and transformation functions for many water quality constituents. While here, we focus on functions that benefit downstream waters; these functions also can have negative effects. For example, wetlands can be a source of methylmercury, a contaminant that can travel to downstream systems, bioaccumulate in fish, and pose a health risk to humans and aquatic life (Galloway and Branfireun, 2004).
TABLE 1.
Functions by which streams and wetlands affect material and energy fluxes to downstream waters. Arrows indicate material and energy imports to and exports from a stream or wetland, in terms of mass or energy; arrow widths represent relative material mass or energy and differences in arrow shades represent timing (lag) or composition (transformation) changes. Imports to streams and wetlands can come from upland terrestrial areas, other streams and wetlands, or from the river itself. Arrows are meant to be illustrative, and do not necessarily represent upstream/downstream relationships. For example, materials and energy can move downstream, upstream, or laterally into streams and wetlands. Examples of commonly exchanged materials and energy include water, heat energy, nutrients, contaminants, sediment, particulate organic matter, organisms, and reproductive propagules; note that exchange of materials and energy between streams and wetlands and downstream systems can result in positive or negative effects on downstream waters. For the refuge function, arrows on the left side represent a river that lacks a refuge and so experiences a reduction in material or energy.
| Function | Definition | Examples |
|---|---|---|
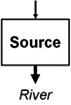
|
Net increase in a material or energy flux (exports > imports) | Streams: invertebrate production (Wipfli and Gregovich, 2002) |
| Wetlands: phytoplankton production from floodplain (Schemel et al., 2004; Lehman et al., 2008) | ||
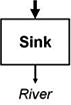
|
Net decrease in a material or energy flux (exports < imports) | Streams: upstream fish populations that are not sustainable without net immigration from downstream areas (Woodford and McIntosh, 2010) |
| Wetlands: sediment deposition, denitrification (Johnston, 1991) | ||
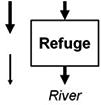
|
Avoidance of a nearby sink function, thereby preventing a net decrease in material or energy flux (exports = imports) | Streams: headwaters as summer coldwater refuges (Curry et al., 1997; Ebersole et al., 2015) |
| Wetlands: riparian wetlands as aquatic refuges in dryland rivers (Leigh et al., 2010) | ||
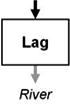
|
Temporary storage and subsequent release of materials or energy without affecting cumulative flux (exports = imports); delivery is delayed and can be prolonged | Streams: delay of downstream peak flows due to bank storage (Burt, 1997); temporary heat storage within the alluvial aquifer (Arrigoni et al., 2008) |
| Wetlands: flood attenuation (Bullock and Acreman, 2003) | ||
|
Conversion of a material or energy into a different form; the amount of the base material or energy is unchanged (base exports = base imports), but its composition (e.g., mass of the different forms) can vary | Streams: conversion of coarse to fine particulate organic matter (Wallace et al., 1995) |
| Wetlands: mercury methylation (Galloway and Branfireun, 2004; Selvendiran et al., 2008) |
Source: U.S. EPA (2015).
The five functions are neither static nor mutually exclusive, and often the distinctions between them are not sharp. A stream or wetland can provide different functions at the same time. These functions can vary with the material considered (e.g., acting as a source of organic matter and a sink for nitrogen; Jones et al., 2015) and can change over time (e.g., acting as a water sink during wet periods and a water source during dry periods; McLaughlin et al., 2014). The magnitude of a given function also is likely to vary temporally: for example, streams can be greater sources of organic matter and contaminants during high flows (Rabiet et al., 2010; Tank et al., 2010; but see Kolpin et al., 2004). The magnitude and timing of these functions is dependent on conditions within the stream or wetland. For example, uptake and transformation of stream nutrients through spiraling is affected by water velocity, residence time, and characteristics of the benthos (Ensign and Doyle, 2006). If a function depends in part on material or energy imports (e.g., denitrification requires a nitrogen source), the function will not be realized in the absence of those imports. However, this function could be provided under appropriate conditions (e.g., changing land use leading to non-point source nitrogen imports). The capacity for such potential functions can be instrumental in protecting waters from future impacts (Leibowitz et al., 2008).
Three factors can influence the effect that material and energy fluxes from streams and wetlands have on downstream waters: (1) proportion of the material originating from (or reduced by) streams and wetlands relative to the importance of other system components, such as the river itself; (2) residence time of the material in the downstream water; and (3) relative importance of the material to river function or ecosystem services. In many cases, the effects on downstream waters need to be considered in aggregate. For example, although macroinvertebrate production per unit length of stream is relatively low in first-order streams of the Little Tennessee River basin, smaller, lower order streams contribute at least 10% of all macroinvertebrate production because they represent a large percentage of the basin’s total stream length (Freeman et al., 2007). Integrating contributions over time also might be necessary, depending on the purpose of the analysis.
In addition to the three factors just described, the more frequently a material is delivered to a downstream water, the greater the opportunity it will have to affect function. The effect of an infrequently supplied material, however, can be large if the material has a long residence time in the river (Leibowitz et al., 2008). For example, woody debris might be exported to downstream waters infrequently but it can persist in stream channels for hundreds of years (Hyatt and Naiman, 2001). In addition, some materials are more important in determining the structure and function of a river. For example, salmon can serve as a keystone species that regulates other populations and serves as a source of marine-derived nutrients (Schindler et al., 2005).
Connectivity and Transport of Materials to and from Streams and Wetlands
For the functions discussed above to affect a river, transport mechanisms that deliver (or could deliver) these materials to the river are necessary. In this section, we define connectivity, discuss different types of connectivity as well as the directionality of connectivity.
We define connectivity as the degree to which components of a system are connected and interact through various transport mechanisms. This structural definition is related to, but distinct from, functional definitions of connectivity based on the actual flow of materials between system components (e.g., Pringle, 2001). Connectivity among river system components is not a new concept. In fact, much of the theory developed to explain how these systems work focuses on connectivity and the importance of linkages between system components (Vannote et al., 1980; Newbold et al., 1982a; Newbold et al., 1982b; Junk et al., 1989; Ward, 1989; Benda et al., 2004; Thorp et al., 2006). The River Continuum Concept (Vannote et al., 1980) viewed the entire length of rivers, from source to mouth, as a complex hydrologic gradient with predictable longitudinal patterns of ecological structure and function. The key pattern is that downstream communities are organized, in large part, by upstream communities and processes (Vannote et al., 1980; Battin et al., 2008). The Serial Discontinuity Concept (Ward and Stanford, 1983) built on the River Continuum Concept to improve our understanding of how dams and impoundments disrupt the longitudinal patterns of flowing waters with predictable downstream effects. The Spiraling Concept (Webster and Patten, 1979; Newbold et al., 1981; Elwood et al., 1983) described how river network connectivity can be evaluated and quantified as materials cycle from dissolved forms to transiently stored forms taken up by living organisms, then back to dissolved forms, as they are transported downstream. These three conceptual frameworks focused on the longitudinal connections of river ecosystems, whereas the subsequent flood pulse concept (Junk et al., 1989) examined the importance of lateral connectivity of river channels to floodplains, including wetlands and open waters, through seasonal expansion and contraction of river networks. Ward (1989) summarized the importance of connectivity to lotic ecosystems along four dimensions: longitudinal, lateral, vertical (surface-subsurface), and temporal connections; he concluded that running water ecosystems are open systems that are highly interactive with both contiguous habitats and other ecosystems in the surrounding landscape. Euliss et al. (2004) added to these ideas by proposing the wetland continuum concept, which considered wetland change over time in response to dynamic changes in climate. As these conceptual frameworks illustrate, scientists have long recognized the hydrologic connectivity that the physical structure of river networks represents.
Water movement through the river system (Figure 1) is the primary mechanism providing physical connectivity both within river networks and between those networks and the surrounding landscape (Fullerton et al., 2010). Hydrologic connectivity results from the flow of water, which provides a “hydraulic highway” (Fausch et al., 2002) along which physical, chemical, and biological materials associated with the water (e.g., sediment, woody debris, nutrients, contaminants, organisms) are transported. However, the effect of this water movement can depend on the specific type of connectivity. Leibowitz et al. (2016) found that the type of connectivity—fill and spill vs. fill and merge—affected the hydrological, chemical, and biological response between connected wetlands. Fill and spill occurs when net input of water exceeds wetland storage capacity (fill), causing the wetland to overflow (spill) onto the land surface and into other surface-water bodies (Leibowitz and Vining, 2003; Tromp-van Meerveld and McDonnell, 2006; Shaw et al., 2013; McCauley and Anteau, 2014). Fill and merge occurs when the water depth of one basin exceeds the internal spill point of another basin, but the surface storage of the combined basin is greater than the total net input of water. In this latter case, the basins merge without spilling externally. Fill and spill promotes external hydrologic exports while fill and merge favors internal storage. However, wetlands can also be subsumed by or merge with other aquatic systems through expansion, resulting in continuous surface-water connections to streams over long (>35 km) spatial distances (Vanderhoof and Alexander, 2016). Hydrologic connectivity can also occur through groundwater, e.g., between the river, hyporheic zone, and alluvial aquifer (Boulton et al., 1998; Gooseff, 2010; Bencala et al., 2011; Goodrich et al., submitted).
FIGURE 1.

Water movement in river systems. (A) Common hydrologic flowpaths by which water flows between watersheds and river networks. The three-dimensional process of hyporheic flow, or the movement of water from a river or stream to nearby alluvium and then back to the river or stream, laterally (B) and longitudinally (C). Source: U.S. EPA (2015), as modified from Winter (1998).
River system structure and function also depend on biological connectivity among the system’s populations of aquatic and semiaquatic organisms (Bornette et al., 1998; Steiger et al., 2005; Meyer et al., 2007). Biological connectivity is established by the active and passive movements of living organisms or their reproductive materials (e.g., seeds, eggs, genes) through space (e.g., dispersal, migration) or time (e.g., dormancy). These movements can occur via multiple pathways (e.g., water, wind, other organisms) and multiple mechanisms (e.g., swimming, flying, walking) within aquatic ecosystems and across ecosystem or watershed boundaries (see Schofield et al., submitted for further discussion of biological connectivity in aquatic systems).
Streams and rivers are not pipes (Bencala, 1993; Bencala et al., 2011); the water and associated materials they carry interact with internal components (e.g., alluvium, organisms) through the five functions by which streams and wetlands alter material fluxes (Table 1). Connectivity between streams and wetlands provides opportunities for material and energy fluxes to be altered sequentially by multiple streams and wetlands as the materials are transported downstream (Newbold et al., 1981; Ensign and Doyle, 2006). The aggregate effect of these sequential fluxes determines the proportion, form, and type of material that ultimately reaches downstream waters (Figure 2). These sequential transformations can also create a cumulative time lag for material transported downstream (e.g., through sediment retention or biological uptake).
FIGURE 2.

Illustration of the sequential transformation of materials as they move through the river network, via either downstream transport with water flow (solid black arrows) or via aerial or terrestrial movements (dashed black arrows). Here, an ephemeral headwater stream exports organic matter (at left) and an intermittent headwater stream exports ammonium, which is incorporated into algal biomass (at right). Macroinvertebrates consume these basal food resources and transform them into biomass, which in turn is eaten and transformed into fish biomass in both local and downstream reaches. The blue and green hashed polygons represent non-floodplain wetlands with and without surface outlets, respectively. Source: U.S. EPA (2015).
The opposite of connectivity is isolation, or the degree to which transport mechanisms (i.e., pathways between system components) are lacking. Isolation reduces material fluxes between system components. Although here, we primarily focus on the benefits that connectivity can have on downstream systems, isolation also can have important positive effects on the condition and function of downstream waters. For example, waterborne contaminants that enter a wetland cannot be transported to a river, except by non-hydrologic pathways, if the wetland is hydrologically isolated from the river. Increased isolation can decrease the spread of pathogens (Hess, 1996) and invasive species (e.g., Bodamer and Bossenbroek, 2008), and increase the rate of local adaptation (e.g., Fraser et al., 2011). Isolation can also reduce downstream flooding, as in the case of surface-water storage by depressional wetlands (Vining, 2002; Yang et al., 2010). Thus, both connectivity and isolation should be considered when examining material fluxes from streams and wetlands, and these fluxes—including biological interactions—should be viewed in terms of a balance between these two factors.
Directionality is an important consideration for the effects of streams and wetlands on downstream waters, especially for connectivity driven by hydrologic flows. Wetlands that occur in riparian or floodplain settings (referred to as riparian/floodplain wetlands) can have bidirectional, lateral hydrologic flows in addition to some of the same hydrologic connections that occur in non-floodplain settings (Górski et al., 2014; Yurek et al., 2016). For example, wetlands within a riparian area are connected to the river network through lateral movement of water between the channel and riparian areas (e.g., through overbank flooding [Figure 1A] and hyporheic flow [Figures 1B and 1C]). In contrast, non-floodplain landscape settings include upgradient areas such as hillslopes or upland areas outside of the floodplain. U.S. EPA (2015) stated that wetlands occurring in such settings, referred to as non-floodplain wetlands, can hydrologically connect to the stream network only through unidirectional, lateral hydrologic flows of surface water or groundwater. However, other studies have shown that this is not always true; rather, directionality of hydrologic flows for non-floodplain wetlands is influenced by the type of connection. Fill and spill connectivity is unidirectional with respect to hydrology, since spillage must follow gravity (Tromp-van Meerveld and McDonnell, 2006; Leibowitz et al., 2016). In contrast, lateral expansion of river networks may occur via multiple mechanisms, including surface flow that is generated when the soil system adjacent to the river is fully saturated from a rise in the water table (Hewlett and Hibbert, 1967; McDonnell, 2013). This can result in bidirectional fill and merge connectivity between a non-floodplain wetland and a downstream water (Devitio et al., 1996; Chu, 2015; Vanderhoof and Alexander, 2016). Because of the existence of a high water table during these events, bidirectional subsurface connectivity might also be expected. Which type of directionality dominates in non-floodplain wetlands and under what circumstances are questions requiring further research. However, Leibowitz et al. (2016) suggest that slope should affect the type of connectivity—with fill and merge occurring in flatter areas and fill and spill occurring in steeper areas—while relief and climate should influence the frequency of hydrologic connectivity. More detailed discussion concerning riparian/floodplain and non-floodplain wetlands is given in (Fritz et al., submitted; Lane et al., submitted).
A major consequence of the directionality of hydrologic connections is that, for a non-floodplain wetland connected by fill and spill, waterborne materials can be transported only from the wetland to the river network, whereas waterborne materials can be transported from the wetland to the river network and vice versa for a riparian/floodplain wetland or a non-floodplain wetland connected by fill and merge. With bidirectional flow, there is a mutual, interacting effect on the structure and function of both the wetland and river network. In contrast, wetlands with unidirectional connections to a river can affect the river through the transport of waterborne materials, but the opposite is not true. Note that hydrological directionality does not necessarily control the directionality of geochemical or biological flows. For example, mobile organisms such as fish can move against the flow of water to a headwater stream or non-floodplain wetland (Leibowitz et al., 2016; Schofield et al., submitted), and carcasses of salmon returning to watersheds from the ocean can serve as a source of marine-derived nutrients (Schindler et al., 2005).
Both riparian/floodplain and non-floodplain wetlands can include so-called geographically isolated wetlands (Leibowitz, 2015; Mushet et al., 2015; Calhoun et al., 2017), or wetlands completely surrounded by uplands (Tiner, 2003). These wetlands have no apparent surface-water outlets, but can hydrologically connect to downstream waters through fill and spill, fill and merge, and shallow or deep subsurface flow. Note that the term “geographically isolated” should not be misconstrued as implying functional isolation (Leibowitz, 2003; Tiner, 2003). Determining the functional connectivity of geographically isolated wetlands is essential because of their important hydrological, chemical, and biological contributions to downstream watersheds and other watershed components (Marton et al., 2015; Cohen et al., 2016; Rains et al., 2016; Schofield et al., submitted). However, additional research is required since the hydrologic connectivity of these wetlands is generally difficult to characterize.
Spatial and Temporal Variability of Connectivity
Connectivity is not a fixed characteristic of a system, but varies over space and time (Ward, 1989; Leibowitz, 2003; Leibowitz et al., 2016). In fact, Cohen et al. (2016) have argued that the hydrologic function of watersheds depends on the connectivity-isolation continuum, which results from this variation. Variability in hydrologic connectivity results from the longitudinal, lateral, and vertical expansion and contraction of the river network (Figures 1, 3, and 4) and transient connections with other watershed components (e.g., through ephemeral and episodic connections). Poff et al. (1997) described this variability in terms of frequency, duration, magnitude, timing, and rate of change of connections among system components.
FIGURE 3.

Examples of longitudinal expansion of river networks. Extent and connectivity of streams with flowing water, wetlands, and other water bodies in Spring Valley Creek, Oregon during dry summer (a) and wet winter (b) conditions, and Spoon Creek, Oregon during summer (c) and winter (d) conditions. Source: (a) and (b) U.S. EPA (2015); (c) and (d) modified from Wigington et al. (2005).
FIGURE 4.

Example of lateral expansion of a river network. Landsat 5 satellite images of the Mississippi River along the borders of Tennessee, Kentucky, Missouri, and Arkansas on (A) May 12, 2006 and (B) May 10, 2011. Green and white shades represent land while blue shades represent water. Discharge on May 12, 2006 and May 10, 2011 was 14,977 and 36,529 m3 s−1, respectively, based on gage data at Baton Rouge, Louisiana (USGS gage 07374000, data downloaded from waterdata.usgs.gov/nwis on Oct. 19, 2017; discharge for May 12, 2006 was estimated from discharge-gage height relationship). Images courtesy of U.S. Geological Survey/National Aeronautics Space Administration.
The expansion and contraction of river networks affect the spatial extent, magnitude, timing, and type of hydrologic connectivity. For example, intermittent and ephemeral streams, which are estimated to account for 59% of the total length of streams in the contiguous United States (Nadeau and Rains, 2007), flow continuously only at certain times of year or briefly in direct response to precipitation, respectively. Thus, the spatial extent of connectivity between streams and wetlands and downstream waters increases greatly during these seasonal or episodic high-flow events (Wigington et al., 2005; Figure 3). Changes in the spatial extent of connectivity due to expansion and contraction are particularly pronounced in the arid and semiarid Southwest (Goodrich et al., submitted), where more than 80% of all streams are intermittent or ephemeral (Levick et al., 2008). Expansion and contraction also affect the magnitude of connectivity, because larger flows provide greater potential for material transport (Fritz et al., submitted).
Besides affecting the spatial extent and magnitude of hydrologic connectivity, expansion and contraction of the stream network also affect the duration and timing of flow in different portions of the network. For example, perennial streams have year-round hydrologic connectivity with a downstream river via surface-water flows, whereas intermittent streams have only seasonal surface-water connectivity with downstream segments. Goodrich et al. (submitted) include examples of the importance of intermittent and ephemeral streams. The temporal characteristics of connectivity for ephemeral streams depend on the duration, intensity, and timing of storm events. Similarly, connectivity between wetlands and downstream waters can range from permanent to seasonal to episodic.
The expansion and contraction of river systems also affect the type of hydrologic connectivity. For example, during wet periods, when input from precipitation can exceed evapotranspiration and available storage, non-floodplain wetlands could have connectivity with other wetlands or streams through surface spillage (Rains et al., 2008; Leibowitz et al., 2016). When spillage ceases due to drier conditions, hydrologic connectivity could only occur through groundwater (Rains et al., 2006; Rains et al., 2008).
When the flow of water mediates dispersal, migration, and other forms of biotic movement, biological and hydrologic connectivity can be tightly coupled. For example, seasonal flooding of riparian/floodplain wetlands creates temporary habitat that fish, aquatic insects, and other organisms use (Junk et al., 1989; Smock, 1994; Tockner et al., 2000; Robinson et al., 2002; Tronstad et al., 2007). Factors other than hydrologic dynamics also can affect the temporal and spatial dynamics of biological connectivity. Such factors include movement associated with seasonal habitat use (Moll, 1990; Lamoureux and Madison, 1999) and shifts in habitat use due to life-history changes (Huryn and Gibbs, 1999; Gibbons et al., 2006; Subalusky et al., 2009), quality or quantity of food resources (Smock, 1994), presence or absence of favorable dispersal conditions (Schalk and Luhring, 2010), physical differences in aquatic habitat structure (Grant et al., 2007), or the number and sizes of nearby populations (Gamble et al., 2007). For a specific river system with a given spatial configuration, variability in biological connectivity also occurs due to variation in the dispersal distance of organisms and reproductive propagules (Semlitsch and Bodie, 2003).
Finally, just as connectivity from temporary or seasonal wetting of channels can affect downstream waters, temporary or seasonal drying (i.e., isolation) also can affect river networks. Riverbeds or streambeds that temporarily dry up are used by aquatic organisms that are specially adapted to wet and dry conditions, and can serve as egg and seed banks for organisms including aquatic invertebrates and plants (Steward et al., 2012). These temporary dry areas also can affect nutrient dynamics due to reduced microbial activity, increased oxygen availability, and inputs of terrestrial sources of organic matter and nutrients (Steward et al., 2012). These materials can then be washed downstream during rewetting.
FACTORS INFLUENCING CONNECTIVITY
Numerous factors affect physical, chemical, and biological connectivity within river systems. These factors operate at multiple spatial and temporal scales, and interact with each other in complex ways to determine where components of a system fall on the connectivity-isolation gradient at a given time. Here, we focus on three key factors: climate and watershed characteristics, spatial distribution patterns of waterbodies, and human activities and alterations (see Schofield et al., submitted for a discussion of biota, which is another key determinant of biological connectivity). These are by no means the only factors influencing connectivity, but they illustrate how numerous variables can ultimately shape physical, chemical, and biological connectivity. While many of these factors are derived from basic hydrological and biological concepts, it is important to describe them here since they are fundamental to our framework.
Climate and Watershed Characteristics
The movement and storage of water in watersheds varies with climatic, geologic, physiographic, and edaphic characteristics of river systems (Winter, 2001; Wigington et al., 2013). At broad spatial scales, climate determines the amount, timing, and duration of water available to watersheds and river basins. Key characteristics of water availability that influence connectivity over various temporal scales include annual precipitation surplus (precipitation minus evapotranspiration); timing (seasonality) of water surplus during the year (Stieglitz et al., 2003), which is heavily influenced by precipitation timing and form (e.g., rain, snow); and precipitation intensity (Gomi et al., 2008).
Annual runoff generally reflects water surplus and varies widely across the United States. Seasonality of water surplus during the year determines when and for how long runoff and groundwater recharge occur. Precipitation and water surplus in the eastern United States is well-distributed throughout the year and, therefore, less seasonal than in the West (Finkelstein and Truppi, 1991). The Southwest experiences summer monsoonal rains (Goodrich et al., submitted), whereas the West Coast and Pacific Northwest receive most precipitation during the winter season (Wigington et al., 2013). Throughout the West, winter precipitation in the mountains occurs as snowfall, where it accumulates in seasonal snowpack and is released during the spring and summer melt seasons to sustain streamflow during late spring and summer months (Brooks et al., 2012). The flowing portions of river networks tend to have their maximum extent during seasons with the highest water surplus (Goodrich et al., submitted; Figure 3), when conditions for flooding are most likely. Typically, the occurrence of ephemeral and intermittent streams is greatest in watersheds with low annual runoff and high water surplus seasonality, but this also is influenced by watershed geologic and edaphic features (Gleeson et al., 2011).
Rainfall intensity can affect hydrologic connectivity in localities where watershed surfaces have low infiltration capacities relative to rainfall intensities (Gomi et al., 2008). Infiltration-excess overland flow occurs when rainfall intensity exceeds watershed surface infiltration, and it can be an important mechanism providing water to wetlands and river networks (Goodrich et al., 1997; Levick et al., 2008). Overland flow is common at low elevations in the Southwest, due to the presence of desert soils with low infiltration capacities combined with relatively high rainfall intensities (Goodrich et al., submitted). The Pacific Northwest has low rainfall intensities, whereas many locations in the Mid-Atlantic, Southeast, and Great Plains have higher rainfall intensities. The prevalence of impermeable surfaces in urban areas can generate overland flow in virtually any setting (Booth et al., 2002).
River system topography and landscape form can profoundly influence river network drainage patterns, distribution of wetlands, and groundwater and surface-water flowpaths. Winter (2001) described six generalized hydrologic landscape forms common throughout the United States (Figure 5). These landscape forms can affect the spatial pattern and timing of water delivery. For example, Mountain Valleys (Figure 5A) have constrained valleys with proportionately long, steep sides with narrow to nonexistent floodplains. This results in the rapid movement of water downslope. In contrast, Riverine Valleys (Figure 5D) have extensive floodplains that promote strong surface water, hyporheic water, and alluvial groundwater connections between wetlands and rivers. Small changes in water table elevations can influence the water levels and hydrologic connectivity of wetlands over extensive areas in this landscape form (Vanderhoof and Alexander, 2016; Figure 5D).
FIGURE 5.

Generalized hydrologic landscape forms. A fundamental hydrologic landscape unit is defined by land-surface form, geology, and climate. (A) Mountain Valley: narrow uplands and lowlands separated by large steep valley sides; (B) Playa: large broad lowland separated from narrow uplands by steeper valley sides (playas and basins of interior drainage); (C) Plateau and High Plains: small narrow lowlands separated from broad uplands by steeper valley sides; (D) Riverine Valley: small fundamental landscape units nested inside broader fundamental landscape unit; (E) Coastal Terrain: small fundamental landscape units nested inside broader fundamental landscape unit (coastal plain with terraces and scarps); and (F) Hummocky Terrain: small fundamental landscape units superimposed randomly on larger fundamental landscape unit. Source: U.S. EPA (2015), as modified from Winter (2001).
Hydrologic connectivity between floodplains and rivers and streams occurs primarily through overbank flooding, shallow groundwater flow, and hyporheic flows and exchanges. Water-table depth can influence connectivity across a range of hydrologic landscape forms, especially within floodplains. Wet, high water table conditions influence both groundwater and surface-water connectivity. For example, when water tables are near the watershed surface, they create conditions in which swales and small stream channels fill with water and flow to nearby water bodies (Wigington et al., 2003; Wigington et al., 2005).
Within hydrologic landscape forms, the permeability of soil and geologic formations is an important determinant of hydrologic flowpaths (Wolock et al., 2004; Figure 6). Permeable soils promote infiltration that results in groundwater hydrologic flowpaths (Figures 6A and 6B), whereas the presence of impermeable soils with low infiltration capacities is conducive to overland flow (Figures 6C and 6D). In situations in which groundwater outflows from watersheds or landscapes dominate, the fate of water depends in part on the permeability of deeper geologic strata. The presence of an aquiclude near the land surface increases the potential for shallow subsurface flows through soil or geologic materials (Figure 6A).
FIGURE 6.

Major hydrologic flowpaths for hillslopes with combinations of permeable and impermeable soils and geologic formations. (A) Permeable soil and impermeable underlying geologic formation; (B) permeable soil and permeable underlying geologic formation; (C) impermeable soil and impermeable underlying geologic formation; and (D) impermeable soil and permeable underlying geologic formation. Width of arrow indicates relative magnitude of flow. Note that pavement can be another source of impermeable surfaces and subsequent overland flow in anthropogenically influenced settings. Source: U.S. EPA (2015).
These local groundwater flowpaths connect portions of watersheds to nearby wetlands or streams. Alternatively, if a deep permeable geologic material (an aquifer) is present, water is likely to move deeper into the subsurface, recharge deeper aquifers, and potentially form regional groundwater flowpaths (Figure 6B). Also, the higher the hydraulic conductivity of an alluvial aquifer, the greater the exchange rate between the alluvial aquifer and river waters (Whiting and Pomeranets, 1997). The permeability of soils and geologic formations both can influence the range of hydrologic connectivity between non-floodplain wetlands and river networks. For example, a wetland that is the origin of a stream can have a permanent or temporary surface-water connection with downstream waters through a channelized outlet (Figure 7A); a wetland can be connected to downstream waters by transient surface-water flows through swales (Figure 7B) or by shallow groundwater flows (Figure 7C); or a wetland can be hydrologically isolated from downstream waters (Figure 7D) because it recharges a deep groundwater aquifer that does not feed surface waters, or it is located in a basin where evapotranspiration is the dominant form of water loss.
FIGURE 7.

Types of hydrologic connections between non-floodplain wetlands and streams or rivers. (A) A wetland connected to a river by surface flow through a headwater stream channel. (B) A wetland connected to a river by surface flow through a nonchannelized swale. Such a wetland would be considered geographically isolated if the swale did not meet the Cowardin et al. (1979) three attribute wetland criteria. (C) A geographically isolated wetland connected to a river by groundwater flow (flowpath can be local, intermediate, or regional). (D) A geographically isolated wetland that is hydrologically isolated from a river. Note that in A–C, flows connecting the wetland and river may be perennial, intermittent, or ephemeral. Source: U.S. EPA (2015).
Spatial Distribution Patterns
Climate and watershed characteristics directly affect spatial and temporal patterns of connectivity between streams and wetlands and downstream waters by influencing the timing and extent of river network expansion and contraction (Wigington et al., 2005; Figure 3). They also influence the spatial distribution of water bodies within a watershed (e.g., Tihansky, 1999).
Hydrologic connectivity between streams and rivers can be a function of the distance between the two water bodies (Bracken and Croke, 2007; Peterson et al., 2007). Water in streams and rivers can be lost from the channel through evapotranspiration and bank storage or added through downstream inputs. Thus, material from a headwater stream that flowed directly into a river along a shorter channel length would be subject to less transformation or dilution. On the other hand, the greater the distance a material travels between a particular stream reach and the river, the greater the opportunity for that material to be altered (e.g., taken up, transformed, or assimilated; Figure 2) in intervening stream reaches. This illustrates how relative isolation—in this case through increased distance—can affect a downstream water; this effect will be beneficial if the material being isolated would have a negative impact on the downstream water (e.g., as would be the case for a contaminant). Higher order streams generally are located closer to rivers and, therefore, tend to be more directly connected to downstream waters than upstream reaches of lower order. Although an individual low-order stream can have less connectivity than a high-order stream, a river network has many more low-order streams; thus, the cumulative effect of these low-order streams is significant.
Distance also affects connectivity between riparian/floodplain and non-floodplain wetlands and downstream waters. Riverine wetlands that serve as origins for headwater streams that connect directly to a mainstem river (e.g., first-order streams that directly connect to fifth-order streams) have a more direct connection to that river than wetlands that serve as origins for headwater streams high in a drainage network (e.g., first-order streams that connect to second-order streams). This also applies to riparian/floodplain wetlands that have direct surface-water connections to streams or rivers. For example, Reese and Batzer (2007) examined invertebrate communities in the Altamaha River catchment in Georgia and found evidence of weak biological connectivity between upper reach floodplains and the stream, and strong connectivity in mid-reach floodplains. However, biological connectivity of lower reach floodplains was lower. Further, wetlands that border first-order streams may process a larger percentage of the water entering a stream than wetlands bordering higher order streams (Whigham et al., 1988). If geographically isolated non-floodplain wetlands have surface-water outputs (e.g., depressions that experience surface-water spillage or groundwater seeps; Figure 7B), the probability that surface water will infiltrate or be lost through evapotranspiration increases with distance, and can vary substantially within and among regions (Cohen et al., 2016; Vanderhoof et al., 2017a).
For non-floodplain wetlands connected through groundwater flows, less distant areas are generally connected through shallower flowpaths, assuming similar soil and geologic properties. These shallower groundwater flows have the greatest interchange with surface waters and travel between points in the shortest amount of time. Although elevation is the primary factor determining areas that are inundated through overbank flooding, connectivity with the river generally will be higher for riparian/floodplain wetlands located near the river’s edge compared with riparian/floodplain wetlands occurring near the floodplain edge.
Distance from the river network also influences biological connectivity among streams and wetlands. For example, mortality of an organism due to predators and other natural hazards generally increases with the distance it has to travel to reach the river network. The likelihood that organisms or propagules traveling randomly or by diffusive mechanisms such as wind will arrive at the river network generally decreases as distance increases.
The distribution of distances between wetlands and river networks depends on both the drainage density of the river network (the total length of stream channels per unit area) and the density of wetlands. Climate, geological history, and watershed characteristics influence these spatial patterns, as do human activities, all of which can vary widely. Figure 8 shows wetland landscape settings ranging from a few nearby streams with high wetland density (Figures 8A–C), to less spatially uniform wetlands (Figure 8D), to areas with higher drainage densities coupled with either riparian (Figure 8E) or larger, more extensive (Figure 8F) wetlands. These maps illustrate the degree to which landscape setting can affect the interspersion―and thus average distance―between wetlands and the river network, and the large variability in distances, and therefore connectivity, that can result.
FIGURE 8.

Examples of different landscapes showing interspersion of wetlands and streams or rivers, based on National Wetlands Inventory maps (http://www.fws.gov/wetlands) and high resolution National Hydrography Dataset streams (ftp://nhdftp.usgs.gov/DataSets/Staged/States/FileGDB/HighResolution). (A) Prairie potholes within the Missouri Coteau in North Dakota; (B) prairie potholes within the Drift Prairie in North Dakota; (C) playas in Texas; (D) vernal pools in California; (E) bottomland hardwood wetlands in Illinois; and (F) Carolina bays in North Carolina. Note all maps are at the same scale. Wetlands smaller than the minimum mapping unit (currently 0.4 ha) might not appear on maps.
All factors being equal, wetlands closer to the stream network will have greater hydrologic and biological connectivity to downstream waters than wetlands located farther from the same network. However, all factors often are not equal, and so more distant wetlands can have higher connectivity than wetlands that are closer to downstream waters due to variability in factors such as topography, slope, and soil permeability. For example, Vanderhoof et al. (2017a) found that the average distance over which wetlands showed a surface-water connection to streams varied substantially between ecoregions across the prairie pothole region. In areas dominated by flat, open basins and lakes, small changes in surface-water levels can consolidate wetlands that were previously disconnected by distances greater than 1 km (Vanderhoof and Alexander, 2016). In this case, wetlands often connect to one another first, forming wetland complexes, prior to connecting to a stream channel (Vanderhoof et al., 2017a).
An analysis by Cohen et al. (2016) found that the distribution of wetland distances to the nearest stream with respect to different wetland landscapes was complex, although in most cases followed exponential scaling. They concluded that wetlands occur in a continuum of sizes and separation distances from streams, and hypothesized that geographically isolated wetlands could enable functions that larger wetlands nearer to streams did not support. In particular, they suggested that travel times between geographically isolated wetlands and a stream would vary depending on whether the geographically isolated wetland had frequent, intermittent, or infrequent surface-water connections with the stream, and that the hydrographic response of a watershed was the convolution of all these different travel times. Cohen et al. (2016) suggested that the hydrologic function of watersheds was dependent on the entire continuum of wetland connectivity and isolation.
Human Activities and Alterations
Human activities frequently alter connectivity—and therefore the transfer and movement of materials and energy—between headwater streams, riparian/floodplain wetlands, non-floodplain wetlands, and downgradient river networks (Fullerton et al., 2010; Jackson and Pringle, 2010; Jaeger et al., 2014; Fritz et al., submitted; Lane et al., submitted). This includes alterations that reduce connectivity, such as dams, levees, water abstraction, piping, and burial, and those that increase connectivity, including wetland drainage, irrigation, impervious surfaces, and interbasin transfers. Each of these can alter the transfer and movement of materials and energy between river system components. Furthermore, the individual or cumulative effects of headwater streams and wetlands on river networks are often more discernible—through impairments caused by their loss—following human-mediated changes in degree of connectivity.
Landscape alterations can increase or decrease hydrologic and biological connectivity (or, alternatively, decrease or increase hydrologic and biological isolation). For example, dams and impoundments might impede biotic movement, whereas nonnative species introductions artificially increase biotic movement. Further complicating the issue is that a given activity or alteration might simultaneously increase and decrease connectivity, depending on which part of the river system is considered. For example, channelization can increase hydrologic connectivity by increasing water flow between portions of a river, but decrease connectivity by reducing flow to floodplain wetlands. Similarly, agricultural ditches and tile drainage can increase hydrologic connectivity by facilitating rapid water transport into channels or wetlands, but can also decrease hydrologic connectivity by filling wetlands with soil and lowering the water table (De Laney, 1995; Blann et al., 2009; McCauley et al., 2015).
The presence of dams illustrates the complex effects that human-mediated landscape modifications can have on connectivity. Numerous studies have shown that dams impede biotic movements, reduce biological connectivity between upstream and downstream locations (e.g., Greathouse et al., 2006; Hall et al., 2011), and form a discontinuity in the normal stream-order-related progression in stream ecosystem structure and function (Stanford and Ward, 1984). However, dams can have the opposite effect with respect to natural lakes. For example, a dam may increase a lake’s biological connectivity with respect to invasive species by adding impoundments that decrease average distances between lakes and serving as stepping stone habitat (Johnson et al., 2008). Large dams can also permanently inundate upstream riparian areas, increasing lateral hydrologic connectivity in these locations. In contrast, dams attenuate downstream flow variability by reducing peak stream volumes during normal high-runoff seasons and increasing minimum flows during normal low-flow seasons (Poff et al., 2007). Because many riverine organisms are adapted to the seasonality of natural flow regimes, attenuating flow variability can have deleterious effects on species persistence where dams have been built (Lytle and Poff, 2004). This reduction in high flows also decreases the connectivity of riparian wetlands with the stream by reducing the potential for overbank lateral flow, which can affect downstream water quality (Hupp et al., 2009).
Wetland drainage, primarily for agricultural purposes, has had a great impact on wetland loss and connectivity. Drainage causes a direct loss of function and connectivity in cases where wetland characteristics are completely lost. In the Des Moines lobe of the prairie pothole region, more than 90% of the original wetlands have been drained, disproportionately impacting both small and large wetlands. Accompanying this loss has been significant decreases in wetland perimeter-area ratios. High perimeter-area ratios are associated with greater biogeochemical processing and groundwater recharge rates, so these processes would be negatively impacted by decreased ratios. Further, loss is also associated with increased mean distances between wetlands, which reduce biological connectivity (Van Meter and Basu, 2014). Wetland drainage also enhances hydrologic connectivity between the landscape and downstream waters. Effects of this enhanced hydrologic connectivity include (1) reduced water storage and more rapid conveyance of water to the network, with subsequent increases in total runoff, baseflow, stormflows, and flooding risk (Wiskow and van der Ploeg, 2003; Blann et al., 2009); (2) increased delivery of sediment and pollutants to downstream waters; and (3) increased transport of water-dispersing organisms (Babbitt and Tanner, 2000; Baber et al., 2002; Mulhouse and Galatowitsch, 2003). Biological connectivity also can decrease with drainage and ditching, as average distances between wetlands increase and limit the ability of organisms to disperse aerially or terrestrially between systems (Leibowitz, 2003).
QUANTIFYING CONNECTIVITY
Quantifying connectivity is important for a number of reasons, e.g., to establish a baseline against which changes can be measured, to allow the degree of connectivity to be related to various watershed functions, and to develop restoration and protection criteria. Connectivity can be quantified using structural metrics, measurements, or models of physical landscape features (e.g., watershed topography, the spatial arrangement of habitat patches), or functional metrics, measurements, or models of system dynamics, which integrate information about processes and interactions that influence hydrologic flows or biological dispersal. Selection of specific tools for quantifying connectivity depends on purpose of the assessment, environmental context, type of connection, spatial and temporal scale of interest, and available data (Calabrese and Fagan, 2004; Lexartza-Artza and Wainwright, 2009). An overview of the diverse methods that can be used to quantify hydrologic connectivity and the scale, environment, and applications relevant to each method is provided in Table 2. See Schofield et al. (submitted) for discussion of metrics and measures used to quantify biological connectivity.
TABLE 2.
An overview of methodological approaches to quantify hydrologic connectivity and an assessment of the appropriate scale and environments in which to apply the methods, as well as connectivity type and metrics easily extractable with each method. This table is not comprehensive but represents interpretation of significant efforts within the existing literature.
| Method | Scale | Environments | Connectivity Metrics | Connectivity Type |
Examples |
|---|---|---|---|---|---|
| Water quality indicators | Site | Wetlands, streams | Magnitude | Functional | Johnston et al. (1990); Leibowitz and Vining (2003); Leibowitz et al. (2016) |
| Tracer experiments | Site, watershed | Wetlands, streams | Temporal frequency, magnitude, timing | Functional | Mulholland et al. (2004); Bencala et al. (2011); O'Brien et al. (2012) |
| Flow meters | Site | Streams | Temporal frequency, duration, timing | Functional | McDonough et al. (2015) |
| Modeling stream type | Watershed | Streams | Duration | Structural | Fritz et al. (2009); Nadeau et al. (2015) |
| Topographic indices | Watershed | Wetlands, streams | Spatial frequency and extent | Structural/ Functional | Quinn et al. (1995); Creed and Beall (2009); Chu et al. (2013); Ali et al. (2014) |
| Sediment Delivery Ratio | Watershed | Streams | Magnitude | Structural | Atkinson (1995); Hooke (2003); Bracken and Croke (2007) |
| Catchment scale metrics (e.g., connected to total stream reach ratio | Watershed | Streams | Spatial and temporal frequency, magnitude | Structural/Functional | Phillips et al. (2011); Spence and Phillips (2015) |
| Directional Connectivity Index | Watershed | Wetlands, streams | Spatial frequency | Structural | Larsen et al. (2012) |
| Volume-to-breakthrough | Watershed | Wetlands, streams | Spatial frequency, magnitude | Structural/Functional | Bracken and Croke (2007) |
| Hydrological models (surface, groundwater or coupled) | Watershed | Wetlands, streams | Magnitude, spatial extent, temporal frequency, duration, timing, rate of change | Functional | McLaughlin et al. (2014); Evenson et al. (2015); Evenson et al. (2016); Golden et al. (2016) |
| Geostatistical modeling approaches | Watershed | Streams | Magnitude, spatial extent | Structural | Fagan (2002); Ganio et al. (2005); Peterson et al. (2007) |
| Dynamic contributing area model | Watershed | Wetlands, streams | Spatial frequency and extent | Structural | Shaw et al. (2013) |
| Remotely sensed imagery | Site, watershed | Wetlands, lakes, streams | Spatial frequency and extent, timing, rate of change | Functional | Sass and Creed (2008); Huang et al. (2014); Vanderhoof et al. (2016) |
Movement of water is the primary mechanism by which chemical substances are transported downstream. Therefore, quantifying chemical connectivity is closely related to quantifying hydrologic connectivity (Michalzik et al., 2001; Borselli et al., 2008). Metrics for quantifying hydrologic connections between upstream and downstream waters vary both in the scale at which connections are observed or predicted (e.g., individual waterbody, watershed, region) and the methodological approach taken (e.g., field-based measurements, topographical approaches, hydrological modeling, and remote sensing; Golden et al., 2017).
Field-based approaches can contribute to a general understanding of connectivity between individual water features. They can also be used to evaluate the magnitude of water and associated materials transported, as well as the temporal frequency, timing, and duration of connections. For example, water quality indicators have been used to identify functional connectivity, or the magnitude of actual movement of water between wetlands and streams (Johnston et al., 1990; Leibowitz and Vining, 2003; Leibowitz et al., 2016). Tracer experiments using 15N, bromide, salt solutions, fluorescing particles, or other conservative compounds have also been conducted to assess the frequency, magnitude, and timing of flowpaths in aquatic systems (Mulholland et al., 2004; Bencala et al., 2011; O'Brien et al., 2012). Furthermore, sensors that detect the presence of water in streams can be used to determine the duration of functional connections (McDonough et al., 2015) and can assist in characterizing the hydrologic permanence of streams, which can inform estimates of the duration of connections (Leibowitz et al., 2008; Fritz et al., 2009; Nadeau et al., 2015).
Structural indices derived from topography, alone or in combination with other watershed characteristics (e.g., infiltration and storage capacity, presence of barriers), can be used to predict structural patterns in the spatial frequency, magnitude, and spatial extent of potential connections. For example, the Topographic Wetness Index (Quinn et al., 1995) and other landscape wetness and community indices (Ali et al., 2014), as well as quasi-dynamic indices that calculate the effective contributing area (variable source area) in a watershed (e.g., Barling et al., 1994; Tarboton, 1997; Creed and Beall, 2009), can be used to predict the spatial frequency, total magnitude, and spatial extent of hydrologic connectivity at the watershed scale. These indices can be used to predict the location of hydrologic flowpaths and areas of a watershed that might be efficient material exporters (Creed and Beall, 2009; Lane et al., 2009). The mass transfer efficiency of a watershed can be measured and modeled using a parameter such as the sediment delivery ratio, which describes and predicts the relationship between erosion and sediment yield in a watershed. This information can indicate the degree of structural connectivity in a watershed, thereby allowing for inferences concerning water movement based on the spatial relationships between wetlands and streams within a watershed (Atkinson, 1995; Hooke, 2003; Bracken and Croke, 2007). These kinds of models can be used to analyze functional connectivity if they simulate actual fluxes of water or materials between landscape elements (e.g., a wetland and stream) in response to precipitation events. But they can also be used to examine structural connectivity, i.e., by predicting potential water movement as precipitation increases, based on variations in topography (Chu et al., 2013). Other indices and models can be used to quantify basin-scale hydrologic connectivity using the ratio of stream reaches with flow connected to the outlet to the total potential number of reaches that could connect to the outlet (Phillips et al., 2011; Spence and Phillips, 2015), or as transport potential in a given direction (e.g., the directional connectivity index; Larsen et al., 2012). Similar to this, the volume-to-breakthrough concept quantifies the frequency and magnitude of hydrologic connectivity as actual runoff relative to water inputs, where connectivity decreases with increased infiltration, depression storage, slope length, barriers, or other factors (Bracken and Croke, 2007).
Numerous modeling and simulation tools can be modified and applied to investigate watershed scale hydrologic connectivity dynamics from geographically isolated wetland systems (Golden et al., 2014) and headwaters (e.g., TOPMODEL; Beven and Kirkby, 1979) to downstream surface-water systems. For example, at the watershed-scale, hydrological models have begun to be modified and applied to quantify the effect of geographically isolated wetlands on stream flow (McLaughlin et al., 2014; Evenson et al., 2015; Evenson et al., 2016; Fossey and Rousseau, 2016). Furthermore, groundwater (Harbaugh, 2005) and coupled surface-water-groundwater models (Ameli and Creed, 2017) are beginning to be adapted and applied to answer questions about wetland connectivity. Such approaches could be structural or functional, depending on how estimates are made about water movement (fluxes) across the landscape in response to precipitation inputs. These approaches have the potential to provide a number of connectivity metrics, including the magnitude, spatial extent, temporal frequency, duration, timing, and rate of change (Golden et al., 2016). Geostatistical modeling approaches are also being advanced to quantify connectivity within stream networks, but tend to only address structural connectivity (Fagan, 2002; Ganio et al., 2005; Peterson et al., 2007). Consideration should be given to watershed characteristics (e.g., climate, geology, soils) in order to select the best set of modeling tools for a given application. For example, in flatter landscapes a more dynamic contributing area model is typically required (Shaw et al., 2013). In these systems, the model should have the capacity to quantify wetland fill and spill dynamics (Evenson et al., 2016), which can cause watershed contributing area to expand.
Remotely sensed imagery can also be used to directly map and evaluate functional surface-water connections, including spatial extent and frequency, timing, and changes in hydrologic connectivity over time (Sass and Creed, 2008; Lang et al., 2012; Huang et al., 2014; Vanderhoof et al., 2016). While moderate resolution sources of imagery, such as Landsat, are effective to identify connections that occur through the expansion of larger features, such as lakes (Vanderhoof and Alexander, 2016), finer resolution sources of imagery (e.g., lidar, Worldview-2) can be used to identify changes in spatial extent of surface water for smaller wetlands and facilitate the identification of narrower connections (Huang et al., 2011; Simon et al., 2015; Vanderhoof et al., 2017b).
While all of the preceding approaches are scientifically sound and have their own advantages and disadvantages, there is currently limited consensus regarding the best methods or metrics to quantify or predict hydrologic or chemical connectivity (Lexartza-Artza and Wainwright, 2009; Ali and Roy, 2010; Bracken et al., 2013). Future efforts to quantify connectivity based on frequency, magnitude, duration, timing, and rate of change or other connectivity metrics are necessary. The best path forward will likely require the integration of various types of measurements and models to provide information on connectivity based on convergent evidence (Golden et al., 2017).
SUMMARY AND CONCLUSIONS
Streams and wetlands can influence downstream waters because of (1) their ability to alter material fluxes through source, sink, refuge, lag, and transformation functions, and (2) the presence of various transport mechanisms that connect these systems. Connectivity between these systems is not constant, but varies over space and time. This variability can be described in terms of frequency, duration, magnitude, timing, and rate of change. These characteristics are influenced by climate and the physical features of the watershed—including geology, topography, and the spatial patterns of the component systems—as well as human alterations. Biological connectivity is also affected by species’ traits and behavioral responses to climate and the landscape. Human activities, such as dam construction and wetland drainage, affect connectivity, often in complex ways. There are many scientifically sound approaches for quantifying connectivity, including tracer studies, use of remote sensing, hydrologic models, and graph theoretic approaches, and new applications of these methods continue to appear in the literature. The literature suggests that different analytical approaches and methods are necessary to quantify different types of connectivity across diverse environments and scales.
As aquatic scientists have increasingly focused on interactions between aquatic ecosystem components, there has been an upsurge in research on how connectivity supports these interactions and the factors affecting connectivity. Besides its importance in determining watershed function, research into connectivity has been spurred on by science needs related to the CWA. The current work adds to the literature by providing an integrated systems framework for understanding how streams and wetlands contribute to downstream waters. Although we focus on these specific systems and downstream waters because of their relevance to the CWA, many of the concepts discussed herein are applicable to other systems, such as connectivity between rivers and estuaries, between aquatic and terrestrial systems, and connectivity between similar ecosystem types (e.g., between wetlands).
The existing scientific literature provides substantial information about the connectivity of streams and wetlands to downstream waters. However, further research is necessary in many areas to improve our ability to understand and maintain the long-term sustainability and resiliency of valued aquatic resources. These research areas include:
Although we now have a good general understanding of connectivity between non-floodplain wetlands and rivers, this has not translated into a specific understanding of how these wetlands affect downstream systems. For example, we know that non-floodplain wetlands can serve as source areas that export organic matter and other materials to rivers, but their importance relative to exports from other sources is difficult to assess and not well understood.
Although most research has been focused on the effects of greater connectivity, hydrologic isolation of aquatic ecosystems can also reduce material and energy fluxes that would otherwise arrive at downstream waters. Further study is needed on the benefits that isolated ecosystems provide to downstream waters.
Research is needed to classify and quantify the connectivity of ecosystems in specific watersheds or regional landscapes, e.g., watersheds containing prairie potholes, Carolina and Delmarva bays, pocosins, western vernal pools in California, and Texas coastal prairie wetlands, as well as watersheds dominated by ephemeral streams. This need includes the level of connectivity or isolation prior to and following human impacts. Such information could allow us to better understand the aggregate function of these waters within different kinds of landscapes.
Additional research is needed to understand and predict how different impacts may affect connectivity, and how this might alter watershed function and the integrity of downstream waters. In particular, research is needed on how climate change will affect connectivity. Climate change is expected to cause more extreme weather (Hirabayashi et al., 2008; Diffenbaugh et al., 2012). Studies are needed to consider how these changes might affect downstream waters by impacting the frequency, duration, magnitude, timing, and rate of change of connectivity.
Methods need to be developed based on the best available scientific information that would allow regulators and aquatic managers to incorporate advances in the scientific understanding of connectivity into the process of identifying waters of national, state, or local importance.
Acknowledgments
We thank Rose Kwok of EPA’s Wetlands Division and Karyn Wendelowski of EPA’s Office of General Counsel for useful suggestions relating to the Clean Water Act. Thanks also to Daniel McLaughlin and three anonymous reviewers for their helpful comments. We acknowledge Randy Comeleo for contributions on two of the figures. The information in this document has been funded entirely by the U.S. Environmental Protection Agency, in part through an appointment to the Internship/Research Participation Program at the Office of Research and Development, U.S. Environmental Protection Agency, administered by the Oak Ridge Institute for Science and Education through an interagency agreement between the U.S. Department of Energy and EPA. This manuscript has been subjected to Agency review and has been approved for publication. The views expressed in this paper are those of the authors and do not necessarily reflect the views or policies of the U.S. Environmental Protection Agency. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
LITERATURE CITED
- Alexander LC. Science at the Boundaries: Scientific Support for the Clean Water Rule. Freshwater Science. 2015;34:1588–1594. [Google Scholar]
- Alexander LC, Fritz KM, Schofield KA, Autrey BC, DeMeester JE, Golden HE, Goodrich DC, Kepner WG, Lane CR, LeDuc SD, Leibowitz SG, McManus MG, Pollard AI, Raanan Kiperwas H, Ridley CE, Vanderhoof MK, Wigington PJ., Jr Featured Collection Introduction: Connectivity of Streams and Wetlands to Downstream Waters. Journal of the American Water Resources Association. doi: 10.1111/1752-1688.12632. submitted. (this issue) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ali G, Birkel C, Tetzlaff D, Soulsby C, McDonnell JJ, Tarolli P. A Comparison of Wetness Indices for the Prediction of Observed Connected Saturated Areas under Contrasting Conditions. Earth Surface Processes and Landforms. 2014;39:399–413. [Google Scholar]
- Ali GA, Roy AG. Shopping for Hydrologically Representative Connectivity Metrics in a Humid Temperate Forested Catchment. Water Resources Research. 2010;46:W12544. [Google Scholar]
- Ameli AA, Creed IF. Quantifying Hydrologic Connectivity of Wetlands to Surface Water Systems. Hydrology and Earth System Sciences. 2017;21:1791–1808. [Google Scholar]
- Arrigoni AS, Poole GC, Mertes LAK, O'Daniel SJ, Woessner WW, Thomas SA. Buffered, Lagged, or Cooled? Disentangling Hyporheic Influences on Temperature Cycles in Stream Channels. Water Resources Research. 2008;44:W09418. [Google Scholar]
- Atkinson E. Methods for Assessing Sediment Delivery in River Systems. Hydrological Sciences Journal. 1995;40:273–280. [Google Scholar]
- Babbitt KJ, Tanner GW. Use of Temporary Wetlands by Anurans in a Hydrologically Modified Landscape. Wetlands. 2000;20:313–322. [Google Scholar]
- Baber MJ, Childers DL, Babbitt KJ, Anderson DH. Controls on Fish Distribution and Abundance in Temporary Wetlands. Canadian Journal of Fisheries and Aquatic Sciences. 2002;59:1441–1450. [Google Scholar]
- Barling RD, Moore ID, Grayson RB. A Quasi-Dynamic Wetness Index for Characterizing the Spatial Distribution of Zones of Surface Saturation and Soil Water Content. Water Resources Research. 1994;30:1029–1044. [Google Scholar]
- Battin TJ, Kaplan LA, Findlay S, Hopkinson CS, Marti E, Packman AI, Newbold JD, Sabater F. Biophysical Controls on Organic Carbon Fluxes in Fluvial Networks. Nature Geoscience. 2008;1:95–100. [Google Scholar]
- Bencala KE. A Perspective on Stream-Catchment Connections. Journal of the North American Benthological Society. 1993;12:44–47. [Google Scholar]
- Bencala KE, Gooseff MN, Kimball BA. Rethinking Hyporheic Flow and Transient Storage to Advance Understanding of Stream-Catchment Connections. Water Resources Research. 2011;47 [Google Scholar]
- Benda L, Poff NL, Miller D, Dunne T, Reeves G, Pess G, Pollock M. The Network Dynamics Hypothesis: How Channel Networks Structure Riverine Habitats. Bioscience. 2004;54:413–427. [Google Scholar]
- Beven KJ, Kirkby MJ. A Physically Based, Variable Contributing Area Model of Basin Hydrology. Hydrological Sciences Bulletin. 1979;24:43–69. [Google Scholar]
- Blann KL, Anderson JL, Sands GR, Vondracek B. Effects of Agricultural Drainage on Aquatic Ecosystems: A Review. Critical Reviews in Environmental Science and Technology. 2009;39:909–1001. [Google Scholar]
- Bodamer BL, Bossenbroek JM. Wetlands as Barriers: Effects of Vegetated Waterways on Downstream Dispersal of Zebra Mussels. Freshwater Biology. 2008;53:2051–2060. [Google Scholar]
- Booth DB, Hartley D, Jackson R. Forest Cover, Impervious-Surface Area, and the Mitigation of Stormwater Impacts. Journal of the American Water Resources Association. 2002;38:835–845. [Google Scholar]
- Bornette G, Amoros C, Lamouroux N. Aquatic Plant Diversity in Riverine Wetlands: The Role of Connectivity. Freshwater Biology. 1998;39:267–283. [Google Scholar]
- Borselli L, Cassi P, Torri D. Prolegomena to Sediment and Flow Connectivity in the Landscape: A GIS and Field Numerical Assessment. Catena. 2008;75:268–277. [Google Scholar]
- Boulton AJ, Findlay S, Marmonier P, Stanley EH, Valett HM. The Functional Significance of the Hyporheic Zone in Streams and Rivers. Annual Review of Ecology and Systematics. 1998;29:59–81. [Google Scholar]
- Bracken LJ, Croke J. The Concept of Hydrological Connectivity and Its Contribution to Understanding Runoff-Dominated Geomorphic Systems. Hydrological Processes. 2007;21:1749–1763. [Google Scholar]
- Bracken LJ, Wainwright J, Ali GA, Tetzlaff D, Smith MW, Reaney SM, Roy AG. Concepts of Hydrological Connectivity: Research Approaches, Pathways and Future Agendas. Earth-Science Reviews. 2013;119:17–34. [Google Scholar]
- Brooks JR, Wigington PJ, Jr, Phillips DL, Comeleo R, Coulombe R. Willamette River Basin Surface Water Isoscape (δ18O and δ2H): Temporal Changes of Source Water within the River. Ecosphere. 2012;3:39. [Google Scholar]
- Bukaveckas PA. Rivers. In: Likens GE, editor. Encyclopedia of Inland Waters. Elsevier; Oxford, UK: 2009. pp. 721–732. [Google Scholar]
- Bullock A, Acreman M. The Role of Wetlands in the Hydrological Cycle. Hydrology and Earth System Sciences. 2003;7:358–389. [Google Scholar]
- Burt TP. The Hydrological Role of Buffer Zones within the Drainage Basin System. In: Haycock NE, Burt TP, Goulding KWT, Pinay G, editors. Buffer Zones: Their Processes and Potential in Water Protection. Proceedings of the International Conference on Buffer Zones, September 1996. Quest Environmental; Hertfordshire, UK: 1997. pp. 21–32. [Google Scholar]
- Calabrese JM, Fagan WF. A Comparison-Shopper's Guide to Connectivity Metrics. Frontiers in Ecology and the Environment. 2004;2:529–536. [Google Scholar]
- Calhoun AJK, Mushet DM, Alexander LC, DeKeyser ES, Fowler L, Lane CR, Lang MW, Rains MC, Richter SC, Walls SC. The Significant Surface-Water Connectivity of “Geographically Isolated Wetlands”. Wetlands. 2017;37:801–806. doi: 10.1007/s13157-017-0887-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caruso BS. Science and Policy Integration Issues for Stream and Wetland Jurisdictional Determinations in a Semi-Arid Region of the Western U.S. Wetlands Ecology and Management. 2011;19:351–371. [Google Scholar]
- Chu X. Delineation of Pothole-Dominated Wetlands and Modeling of Their Threshold Behaviors. Journal of Hydrologic Engineering. 2015 doi: 10.1061/(ASCE)HE.1943-5584.0001224. [DOI] [Google Scholar]
- Chu X, Yang J, Chi Y, Zhang J. Dynamic Puddle Delineation and Modeling of Puddle-to-Puddle Filling-Spilling-Merging-Splitting Overland Flow Processes. Water Resources Research. 2013;49:3825–3829. [Google Scholar]
- Cohen MJ, Creed IF, Alexander L, Basu NB, Calhoun AJK, Craft C, D’Amico E, DeKeyser E, Fowler L, Golden HE, Jawitz JW, Kalla P, Kirkman LK, Lane CR, Lang M, Leibowitz SG, Lewis DB, Marton J, McLaughlin DL, Mushet DM, Raanan-Kiperwas H, Rains MC, Smith L, Walls SC. Do Geographically Isolated Wetlands Influence Landscape Functions? Proceedings of the National Academy of Sciences. 2016;113:1978–1986. doi: 10.1073/pnas.1512650113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cowardin LM, Carter V, Golet FC, LaRoe ET. Classification of Wetlands and Deepwater Habitats of the United States. Fish and Wildlife Service, Office of Biological Services; Washington, D.C.: 1979. [Google Scholar]
- Creed IF, Beall FD. Distributed Topographic Indicators for Predicting Nitrogen Export from Headwater Catchments. Water Resources Research. 2009;45:W10407. [Google Scholar]
- Creed IF, Lane CR, Serran JN, Alexander LC, Basu NB, Calhoun AJK, Christensen JR, Cohen MJ, Craft C, D'Amico E, DeKeyser E, Fowler L, Golden HE, Jawitz JW, Kalla P, Kirkman LK, Lang M, Leibowitz SG, Lewis DB, Marton J, McLaughlin DL, Raanan-Kiperwas H, Rains MC, Rains KC, Smith L. Enhancing Protection for Vulnerable Waters. Nature Geoscience advance online publication. 2017 doi: 10.1038/ngeo3041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crook DA, Lowe WH, Allendorf FW, Erős T, Finn DS, Gillanders BM, Hadwen WL, Harrod C, Hermoso V, Jennings S, Kilada RW, Nagelkerken I, Hansen MM, Page TJ, Riginos C, Fry B, Hughes JM. Human Effects on Ecological Connectivity in Aquatic Ecosystems: Integrating Scientific Approaches to Support Management and Mitigation. Science of the Total Environment. 2015;534:52–64. doi: 10.1016/j.scitotenv.2015.04.034. [DOI] [PubMed] [Google Scholar]
- Curry RA, Brady C, Noakes DLG, Danzmann RG. Use of Small Streams by Young Brook Trout Spawned in a Lake. Transactions of the American Fisheries Society. 1997;126:77–83. [Google Scholar]
- De Laney TA. Benefits to Downstream Flood Attenuation and Water Quality as a Result of Constructed Wetlands in Agricultural Landscapes. Journal of Soil and Water Conservation. 1995;50:620–626. [Google Scholar]
- Devitio KJ, Hill AR, Roulet N. Groundwater-Surface Water Interactions in Headwater Forested Wetlands of the Canadian Shield. Journal of Hydrology. 1996;181:127–147. [Google Scholar]
- Diffenbaugh NS, Scherer M, Ashfaq M. Response of Snow-Dependent Hydrologic Extremes to Continued Global Warming. Nature Climate Change. 2012 doi: 10.1038/NCLIMATE1732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ebersole JL, Wigington PJ, Jr, Leibowitz SG, Comeleo RL, Van Sickle J. Predicting the Occurrence of Cold-Water Patches at Intermittent and Ephemeral Tributary Confluences with Warm Rivers. Freshwater Science. 2015;34:111–124. [Google Scholar]
- Elwood JW, Newbold JD, O'Neill RV, Van Winkle W. Resource Spiralling: An Operational Paradigm for Analyzing Lotic Ecosystems. In: Fontaine TD, Bartell SM, editors. Dynamics of Lotic Ecosystems. Ann Arbor Science Publishers; Ann Arbor, Michigan: 1983. pp. 3–23. [Google Scholar]
- Ensign SH, Doyle MW. Nutrient Spiraling in Streams and River Networks. Journal of Geophysical Research-Biogeosciences. 2006;111 [Google Scholar]
- Euliss NH, LaBaugh JW, Fredrickson LH, Mushet DM, Laubhan MK, Swanson GA, Winter TC, Rosenberry DO, Nelson RD. The Wetland Continuum: A Conceptual Framework for Interpreting Biological Studies. Wetlands. 2004;24:448–458. [Google Scholar]
- Evenson GR, Golden HE, Lane CR, D'Amico E. Geographically Isolated Wetlands and Watershed Hydrology: A Modified Model Analysis. Journal of Hydrology. 2015;529(Part 1):240–256. [Google Scholar]
- Evenson GR, Golden HE, Lane CR, D'Amico E. An Improved Representation of Geographically Isolated Wetlands in a Watershed-Scale Hydrologic Model. Hydrological Processes. 2016;30:4168–4184. [Google Scholar]
- Fagan WF. Connectivity, Fragmentation, and Extinction Risk in Dendritic Metapopulations. Ecology. 2002;83:3243–3249. [Google Scholar]
- Fausch KD, Torgersen CE, Baxter CV, Li HW. Landscapes to Riverscapes: Bridging the Gap between Research and Conservation of Stream Fishes. BioScience. 2002;52:483–498. [Google Scholar]
- Finkelstein PL, Truppi LE. Spatial Distribution of Precipitation Seasonality in the United States. Journal of Climate. 1991;4:373–385. [Google Scholar]
- Fossey M, Rousseau AN. Can Isolated and Riparian Wetlands Mitigate the Impact of Climate Change on Watershed Hydrology? A Case Study Approach. Journal of Environmental Management. 2016;184(Part 2):327–339. doi: 10.1016/j.jenvman.2016.09.043. [DOI] [PubMed] [Google Scholar]
- Fossey M, Rousseau AN, Savary S. Assessment of the Impact of Spatio-Temporal Attributes of Wetlands on Stream Flows Using a Hydrological Modelling Framework: A Theoretical Case Study of a Watershed under Temperate Climatic Conditions. Hydrological Processes. 2016;30:1768–1781. [Google Scholar]
- Fraser DJ, Weir LK, Bernatchez L, Hansen MM, Taylor EB. Extent and Scale of Local Adaptation in Salmonid Fishes: Review and Meta-Analysis. Heredity. 2011;106:404–420. doi: 10.1038/hdy.2010.167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Freeman MC, Pringle CM, Jackson CR. Hydrologic Connectivity and the Contribution of Stream Headwaters to Ecological Integrity at Regional Scales. Journal of the American Water Resources Association. 2007;43:5–14. [Google Scholar]
- Fritz KM, Glime JM, Hribljan J, Greenwood JL. Can Bryophytes Be Used to Characterize Hydrologic Permanence in Forested Headwater Streams? Ecological Indicators. 2009;9:681–692. [Google Scholar]
- Fritz KM, Schofield KA, Alexander LC, McManus MG, Golden HE, Lane CR, Kepner WG, LeDuc SD, DeMeester JE, Pollard AI. Physical and Chemical Connectivity of Streams and Riparian Wetlands to Downstream Waters: A Synthesis. Journal of the American Water Resources Association. doi: 10.1111/1752-1688.12632. submitted. (this issue) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fullerton AH, Burnett KM, Steel EA, Flitcroft RL, Pess GR, Feist BE, Torgersen CE, Miller DJ, Sanderson BL. Hydrological Connectivity for Riverine Fish: Measurement Challenges and Research Opportunities. Freshwater Biology. 2010;55:2215–2237. [Google Scholar]
- Galloway ME, Branfireun BA. Mercury Dynamics of a Temperate Forested Wetland. Science of the Total Environment. 2004;325:239–254. doi: 10.1016/j.scitotenv.2003.11.010. [DOI] [PubMed] [Google Scholar]
- Gamble DE, Grody J, Mack JJ, Micacchion M. An Ecological and Functional Assessment of Urban Wetlands in Central Ohio. Columbus, Ohio. Ohio Environmental Protection Agency, Wetland Ecology Group, Division of Surface Water; Columbus, OH: 2007. [Google Scholar]
- Ganio LM, Torgersen CE, Gresswell RE. A Geostatistical Approach for Describing Spatial Pattern in Stream Networks. Frontiers in Ecology and the Environment. 2005;3:138–144. [Google Scholar]
- Gibbons JW, Winne CT, Scott DE, Willson JD, Glaudas X, Andrews KM, Todd BD, Fedewa LA, Wilkinson L, Tsaliagos RN, Harper SJ, Greene JL, Tuberville TD, Metts BS, Dorcast ME, Nestor JP, Young CA, Akre T, Reed RN, Buhlmann KA, Norman J, Croshaw DA, Hagen C, Rothermel BB. Remarkable Amphibian Biomass and Abundance in an Isolated Wetland: Implications for Wetland Conservation. Conservation Biology. 2006;20:1457–1465. doi: 10.1111/j.1523-1739.2006.00443.x. [DOI] [PubMed] [Google Scholar]
- Giblin SM, Houser JN, Sullivan JF, Langrehr HA, Rogala JT, Campbell BD. Thresholds in the Response of Free-Floating Plant Abundance to Variation in Hydraulic Connectivity, Nutrients, and Macrophyte Abundance in a Large Floodplain River. Wetlands. 2014;34:413–425. [Google Scholar]
- Gleeson T, Marklund L, Smith L, Manning AH. Classifying the Water Table at Regional to Continental Scales. Geophysical Research Letters. 2011;38:L05401. [Google Scholar]
- Golden HE, Creed IF, Ali G, Basu NB, Neff B, Rains MC, McLaughlin DL, Alexander LC, Ameli AA, Christensen JR, Evenson GR, Jones CN, Lane CR, Lang M. Scientific Tools for Integrating Geographically Isolated Wetlands into Land Management Decisions. Frontiers in Ecology and the Environment. 2017 doi: 10.1002/fee.1504. (accepted) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Golden HE, Lane CR, Amatya DM, Bandilla KW, Raanan Kiperwas H, Knightes CD, Ssegane H. Hydrologic Connectivity between Geographically Isolated Wetlands and Surface Water Systems: A Review of Select Modeling Methods. Environmental Modelling & Software. 2014;53:190–206. [Google Scholar]
- Golden HE, Sander HA, Lane CR, Zhao C, Price K, D’Amico E, Christensen JR. Relative Effects of Geographically Isolated Wetlands on Streamflow: A Watershed-Scale Analysis. Ecohydrology. 2016;9:21–38. [Google Scholar]
- Gomi T, Sidle RC, Miyata S, Kosugi K, Onda Y. Dynamic Runoff Connectivity of Overland Flow on Steep Forested Hillslopes: Scale Effects and Runoff Transfer. Water Resources Research. 2008;44 [Google Scholar]
- Goodrich DC, Kepner WG, Levick LR, Wigington PJ. Southwestern Intermittent and Ephemeral Stream Connectivity. Journal of the American Water Resources Association. submitted. (this issue) [Google Scholar]
- Goodrich DC, Lane LJ, Shillito RM, Miller SN, Syed KH, Woolhiser DA. Linearity of Basin Response as a Function of Scale in a Semiarid Watershed. Water Resources Research. 1997;33:2951–2965. [Google Scholar]
- Gooseff MN. Defining Hyporheic Zones – Advancing Our Conceptual and Operational Definitions of Where Stream Water and Groundwater Meet. Geography Compass. 2010;4/8:945–955. [Google Scholar]
- Górski K, Collier KJ, Hamilton DP, Hicks BJ. Effects of Flow on Lateral Interactions of Fish and Shrimps with Off-Channel Habitats in a Large River-Floodplain System. Hydrobiologia. 2014;729:161–174. [Google Scholar]
- Grant EHC, Lowe WH, Fagan WF. Living in the Branches: Population Dynamics and Ecological Processes in Dendritic Networks. Ecology Letters. 2007;10:165–175. doi: 10.1111/j.1461-0248.2006.01007.x. [DOI] [PubMed] [Google Scholar]
- Greathouse EA, Pringle CM, McDowell WH, Holmquist JG. Indirect Upstream Effects of Dams: Consequences of Migratory Consumer Extirpation in Puerto Rico. Ecological Applications. 2006;16:339–352. doi: 10.1890/05-0243. [DOI] [PubMed] [Google Scholar]
- Hall CJ, Jordaan A, Frisk MG. The Historic Influence of Dams on Diadromous Fish Habitat with a Focus on River Herring and Hydrologic Longitudinal Connectivity. Landscape Ecology. 2011;26:95–107. [Google Scholar]
- Harbaugh AW. U.S. Geological Survey Techniques and Methods 6-A16. U.S. Geological Survey; Reston, Virginia: 2005. MODFLOW-2005, the U.S. Geological Survey Modular Ground-Water Model: The Ground-Water Flow Process. [Google Scholar]
- Harvey J, Gooseff M. River Corridor Science: Hydrologic Exchange and Ecological Consequences from Bedforms to Basins. Water Resources Research. 2015;51:6893–6922. [Google Scholar]
- Hauer FR, Locke H, Dreitz VJ, Hebblewhite M, Lowe WH, Muhlfeld CC, Nelson CR, Proctor MF, Rood SB. Gravel-Bed River Floodplains Are the Ecological Nexus of Glaciated Mountain Landscapes. Science Advances. 2016;2 doi: 10.1126/sciadv.1600026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayashi M, van der Kamp G, Rosenberry DO. Hydrology of Prairie Wetlands: Understanding the Integrated Surface-Water and Groundwater Processes. Wetlands. 2016;36(Suppl 2):S237–S254. [Google Scholar]
- Hess GR. Linking Extinction to Connectivity and Habitat Destruction in Metapopulation Models. Am Nat. 1996;148:226–236. [Google Scholar]
- Hewlett JD, Hibbert AR. Proceedings of the International Symposium on Forest Hydrology. Pergamon Press; Oxford: 1967. Factors Affecting the Response of Small Watersheds to Precipitation in Humid Areas; pp. 275–290. W.E. Sopper and H.W. Lull (W.E. Sopper and H.W. Lull(W.E. Sopper and H.W. Lulls) [Google Scholar]
- Hirabayashi Y, Kanae S, Emori S, Oki T, Kimoto M. Global Projections of Changing Risks of Floods and Droughts in a Changing Climate. Hydrological Sciences Journal. 2008;53:754–772. [Google Scholar]
- Hooke J. Coarse Sediment Connectivity in River Channel Systems: A Conceptual Framework and Methodology. Geomorphology. 2003;56:79–94. [Google Scholar]
- Huang C, Peng Y, Lang M, Yeo I-Y, McCarty G. Wetland Inundation Mapping and Change Monitoring Using Landsat and Airborne Lidar Data. Remote Sensing of Environment. 2014;141:231–242. [Google Scholar]
- Huang S, Young C, Feng M, Heidemann K, Cushing M, Mushet DM, Liu S. Demonstration of a Conceptual Model for Using Lidar to Improve the Estimation of Floodwater Mitigation Potential of Prairie Pothole Region Wetlands. Journal of Hydrology. 2011;405:417–426. [Google Scholar]
- Hupp CR, Pierce AR, Noe GB. Floodplain Geomorphic Processes and Environmental Impacts of Human Alteration Along Coastal Plain Rivers, USA. Wetlands. 2009;29:413–429. [Google Scholar]
- Huryn AD, Gibbs KE. Riparian Sedge Meadows in Maine. A Macroinvertebrate Community Structured by River-Floodplain Interaction. In: Batzer DP, Rader RB, Wissinger SA, editors. Invertebrates in Freshwater Wetlands of North America: Ecology and Management. John Wiley & Sons; New York, NY: 1999. pp. 363–382. [Google Scholar]
- Hyatt TL, Naiman RJ. The Residence Time of Large Woody Debris in the Queets River, Washington, USA. Ecological Applications. 2001;11:191–202. [Google Scholar]
- Jackson CR, Pringle CM. Ecological Benefits of Reduced Hydrologic Connectivity in Intensively Developed Landscapes. Bioscience. 2010;60:37–46. [Google Scholar]
- Jaeger KL, Olden JD, Pelland NA. Climate Change Poised to Threaten Hydrologic Connectivity and Endemic Fishes in Dryland Streams. Proceedings of the National Academy of Sciences. 2014;111:13894–13899. doi: 10.1073/pnas.1320890111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Janzen D, McDonnell JJ. A Stochastic Approach to Modelling and Understanding Hillslope Runoff Connectivity Dynamics. Ecological Modelling. 2015;298:64–74. [Google Scholar]
- Jencso KG, McGlynn BL. Hierarchical Controls on Runoff Generation: Topographically Driven Hydrologic Connectivity, Geology, and Vegetation. Water Resources Research. 2011;47 [Google Scholar]
- Jencso KG, McGlynn BL, Gooseff MN, Bencala KE, Wondzell SM. Hillslope Hydrologic Connectivity Controls Riparian Groundwater Turnover: Implications of Catchment Structure for Riparian Buffering and Stream Water Sources. Water Resources Research. 2010;46 [Google Scholar]
- Johnson PTJ, Olden JD, Vander Zanden MJ. Dam Invaders: Impoundments Facilitate Biological Invasions into Freshwaters. Frontiers in Ecology and the Environment. 2008;6:357–363. [Google Scholar]
- Johnston CA. Sediment and Nutrient Retention by Freshwater Wetlands: Effects on Surface Water Quality. Critical Reviews in Environmental Control. 1991;21:491–565. [Google Scholar]
- Johnston CA, Detenbeck NE, Niemi GJ. The Cumulative Effect of Wetlands on Stream Water Quality and Quantity. A Landscape Approach. Biogeochemistry. 1990;10:105–141. [Google Scholar]
- Jones CN, Scott DT, Guth C, Hester ET, Hession WC. Seasonal Variation in Floodplain Biogeochemical Processing in a Restored Headwater Stream. Environmental Science & Technology. 2015;49:13190–13198. doi: 10.1021/acs.est.5b02426. [DOI] [PubMed] [Google Scholar]
- Jordan SJ, Benson WH. Sustainable Watersheds: Integrating Ecosystem Services and Public Health. Environmental Health Insight. 2015;9(S2):1–7. doi: 10.4137/EHI.S19586. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Junk WJ, Bayley PB, Sparks RE. The Flood Pulse Concept in River-Floodplain Systems. In: Dodge DP, editor. Proceedings of the International Large River Symposium. Ottawa: 1989. pp. 110–127. [Google Scholar]
- Kolpin DW, Skopec M, Meyer MT, Furlong ET, Zaugg SD. Urban Contribution of Pharmaceuticals and Other Organic Wastewater Contaminants to Streams During Differing Flow Conditions. Science of the Total Environment. 2004;328:119–130. doi: 10.1016/j.scitotenv.2004.01.015. [DOI] [PubMed] [Google Scholar]
- Lamoureux VS, Madison DM. Overwintering Habitats of Radio-Implanted Green Frogs, Rana Clamitans. Journal of Herpetology. 1999;33:430–435. [Google Scholar]
- Lane CR, Leibowitz SG, Autrey BC, LeDuc SD, Alexander LC. Hydrological, Physical, and Chemical Functions and Connectivity of Non-Floodplain Wetlands to Downstream Waters: A Review. Journal of the American Water Resources Association. doi: 10.1111/1752-1688.12633. submitted. (this issue) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane SN, Reaney SM, Heathwaite AL. Representation of Landscape Hydrological Connectivity Using a Topographically Driven Surface Flow Index. Water Resources Research. 2009;45:W08423. [Google Scholar]
- Lang M, McDonough O, McCarty G, Oesterling R, Wilen B. Enhanced Detection of Wetland-Stream Connectivity Using Lidar. Wetlands. 2012;32:461–473. [Google Scholar]
- Larsen LG, Choi J, Nungesser MK, Harvey JW. Directional Connectivity in Hydrology and Ecology. Ecological Applications. 2012;22:2204–2220. doi: 10.1890/11-1948.1. [DOI] [PubMed] [Google Scholar]
- Lehman PW, Sommer T, Rivard L. The Influence of Floodplain Habitat on the Quantity and Quality of Riverine Phytoplankton Carbon Produced During the Flood Season in San Francisco Estuary. Aquatic Ecology. 2008;42:363–378. [Google Scholar]
- Leibowitz SG. Isolated Wetlands and Their Functions: An Ecological Perspective. Wetlands. 2003;23:517–531. [Google Scholar]
- Leibowitz SG. Geographically Isolated Wetlands: Why We Should Keep the Term. Wetlands. 2015;35:997–1003. [Google Scholar]
- Leibowitz SG, Mushet DM, Newton WE. Intermittent Surface Water Connectivity: Fill and Spill Vs. Fill and Merge Dynamics. Wetlands. 2016;36(Suppl 2):S323–S342. [Google Scholar]
- Leibowitz SG, Nadeau T-L. Isolated Wetlands: State-of-the-Science and Future Directions. Wetlands. 2003;23:663–684. [Google Scholar]
- Leibowitz SG, Vining KC. Temporal Connectivity in a Prairie Pothole Complex. Wetlands. 2003;23:13–25. [Google Scholar]
- Leibowitz SG, Wigington PJ, Jr, Rains MC, Downing DM. Non-Navigable Streams and Adjacent Wetlands: Addressing Science Needs Following the Supreme Court's Rapanos Decision. Frontiers in Ecology and the Environment. 2008;6:364–371. [Google Scholar]
- Leigh C, Sheldon F, Kingsford RT, Arthington AH. Sequential Floods Drive 'Booms' and Wetland Persistence in Dryland Rivers: A Synthesis. Marine and Freshwater Research. 2010;61:896–908. [Google Scholar]
- Levick L, Fonseca J, Goodrich D, Hernandez M, Semmens D, Leidy R, Scianni M, Guertin P, Tluczek M, Kepner W. The Ecological and Hydrological Significance of Ephemeral and Intermittent Streams in the Arid and Semi-Arid American Southwest. U.S. Environmental Protection Agency, Office of Research and Development and U.S. Department of Agriculture/Agricultural Research Service. Southwest Watershed Research Center; Washington, DC: 2008. [Google Scholar]
- Lexartza-Artza I, Wainwright J. Hydrological Connectivity: Linking Concepts with Practical Implications. Catena. 2009;79:146–152. [Google Scholar]
- Lytle DA, Poff NL. Adaptation to Natural Flow Regimes. Trends in Ecology and Evolution. 2004;19:94–100. doi: 10.1016/j.tree.2003.10.002. [DOI] [PubMed] [Google Scholar]
- Marton JM, Creed IF, Lewis DB, Lane CR, Basu NB, Cohen MJ, Craft CB. Geographically Isolated Wetlands Are Important Biogeochemical Reactors on the Landscape. Bioscience. 2015;65:408–418. [Google Scholar]
- McCauley LA, Anteau MJ. Generating Nested Wetland Catchments with Readily-Available Digital Elevation Data May Improve Evaluations of Land-Use Change on Wetlands. Wetlands. 2014;34:1123–1132. [Google Scholar]
- McCauley LA, Anteau MJ, van der Burg MP, Wiltermuth MT. Land Use and Wetland Drainage Affect Water Levels and Dynamics of Remaining Wetlands. Ecosphere. 2015;6:1–22. [Google Scholar]
- McDonnell JJ. Are All Runoff Processes the Same? Hydrological Processes. 2013;27:4103–4111. [Google Scholar]
- McDonough OT, Lang MW, Hosen JD, Palmer MA. Surface Hydrologic Connectivity between Delmarva Bay Wetlands and Nearby Streams Along a Gradient of Agricultural Alteration. Wetlands. 2015;35:41–53. [Google Scholar]
- McIntyre NE, Wright CK, Swain S, Hayhoe K, Liu G, Schwartz FW, Henebry GM. Climate Forcing of Wetland Landscape Connectivity in the Great Plains. Frontiers in Ecology and the Environment. 2014;12:59–64. [Google Scholar]
- McLaughlin DL, Kaplan DA, Cohen MJ. A Significant Nexus: Geographically Isolated Wetlands Influence Landscape Hydrology. Water Resources Research. 2014;50:7153–7166. [Google Scholar]
- Meyer JL, Strayer DL, Wallace JB, Eggert SL, Helfman GS, Leonard NE. The Contribution of Headwater Streams to Biodiversity in River Networks. Journal of the American Water Resources Association. 2007;43:86–103. [Google Scholar]
- Michalzik B, Kalbitz K, Park JH, Solinger S, Matzner E. Fluxes and Concentrations of Dissolved Organic Carbon and Nitrogen – a Synthesis for Temperate Forests. Biogeochemistry. 2001;52:173–205. [Google Scholar]
- Mitchell MGE, Bennett EM, Gonzalez A. Linking Landscape Connectivity and Ecosystem Service Provision: Current Knowledge and Research Gaps. Ecosystems. 2013;16:894–908. [Google Scholar]
- Moll D. Population Sizes and Foraging Ecology in a Tropical Freshwater Stream Turtle Community. Journal of Herpetology. 1990;24:48–53. [Google Scholar]
- Moore JW. Bidirectional Connectivity in Rivers and Implications for Watershed Stability and Management. Canadian Journal of Fisheries and Aquatic Sciences. 2015;72:785–795. [Google Scholar]
- Mulholland PJ, Valett HM, Webster JR, Thomas SA, Cooper LW, Hamilton SK, Peterson BJ. Stream Denitrification and Total Nitrate Uptake Rates Measured Using a Field 15N Tracer Addition Approach. Limnology and Oceanography. 2004;49:809–820. [Google Scholar]
- Mulhouse JM, Galatowitsch SM. Revegetation of Prairie Pothole Wetlands in the Mid-Continental Us: Twelve Years Post-Reflooding. Plant Ecology. 2003;169:143–159. [Google Scholar]
- Mushet DM, Calhoun AJK, Alexander LC, Cohen MJ, DeKeyser ES, Fowler L, Lane CR, Lang MW, Rains MC, Walls SC. Geographically Isolated Wetlands: Rethinking a Misnomer. Wetlands. 2015;35:423–431. [Google Scholar]
- Nadeau T-L, Leibowitz SG, Wigington PJ, Jr, Ebersole JL, Fritz KM, Coulombe RA, Comeleo RL, Blocksom KA. Validation of Rapid Assessment Methods to Determine Streamflow Duration Classes in the Pacific Northwest, USA. Environmental Management. 2015;56:34–53. doi: 10.1007/s00267-015-0466-4. [DOI] [PubMed] [Google Scholar]
- Nadeau TL, Rains MC. Hydrological Connectivity between Headwater Streams and Downstream Waters: How Science Can Inform Policy. Journal of the American Water Resources Association. 2007;43:118–133. [Google Scholar]
- Naiman RJ, Bilby RE. River Ecology and Management: Lessons from the Pacific Coastal Ecoregion. New York, NY: Springer-Verlag; 1998. [Google Scholar]
- Newbold JD, Elwood JW, O'Neill RV, Van Winkle W. Measuring Nutrient Spiralling in Streams. Canadian Journal of Fisheries and Aquatic Sciences. 1981;38:860–863. [Google Scholar]
- Newbold JD, Mulholland PJ, Elwood JW, Oneill RV. Organic Carbon Spiralling in Stream Ecosystems. Oikos. 1982a;38:266–272. [Google Scholar]
- Newbold JD, O’Neill RV, Elwood JW, Van Winkle W. Nutrient Spiraling in Streams: Implications for Nutrient Limitation and Invertebrate Activity. Am Nat. 1982b;120:628–665. [Google Scholar]
- O'Brien JM, Hamilton SK, Kinsman-Costello LE, Lennon JT, Ostrom NE. Nitrogen Transformations in a through-Flow Wetland Revealed Using Whole-Ecosystem Pulsed 15N Additions. Limnology and Oceanography. 2012;57:221–234. [Google Scholar]
- Peterson EE, Theobald DM, Ver Hoef JM. Geostatistical Modelling on Stream Networks: Developing Valid Covariance Matrices Based on Hydrologic Distance and Stream Flow. Freshwater Biology. 2007;52:267–279. [Google Scholar]
- Phillips RW, Spence C, Pomeroy JW. Connectivity and Runoff Dynamics in Heterogeneous Basins. Hydrological Processes. 2011;25:3061–3075. [Google Scholar]
- Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC. The Natural Flow Regime. Bioscience. 1997;47:769–784. [Google Scholar]
- Poff NL, Olden JD, Merritt DM, Pepin DM. Homogenization of Regional River Dynamics by Dams and Global Biodiversity Implications. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:5732–5737. doi: 10.1073/pnas.0609812104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pringle CM. Hydrologic Connectivity and the Management of Biological Reserves: A Global Perspective. Ecological Applications. 2001;11:981–998. [Google Scholar]
- Quinn PF, Beven KJ, Lamb R. The ln(a/tanβ) Index: How to Calculate It and How to Use It within the Topmodel Framework. Hydrological Processes. 1995;9:161–182. [Google Scholar]
- Rabiet M, Margoum C, Gouy V, Carluer N, Coquery M. Assessing Pesticide Concentrations and Fluxes in the Stream of a Small Vineyard Catchment – Effect of Sampling Frequency. Environmental Pollution. 2010;158:737–748. doi: 10.1016/j.envpol.2009.10.014. [DOI] [PubMed] [Google Scholar]
- Rains MC, Dahlgren RA, Fogg GE, Harter T, Williamson RJ. Geological Control of Physical and Chemical Hydrology in California Vernal Pools. Wetlands. 2008;28:347–362. [Google Scholar]
- Rains MC, Fogg GE, Harter T, Dahlgren RA, Williamson RJ. The Role of Perched Aquifers in Hydrological Connectivity and Biogeochemical Processes in Vernal Pool Landscapes, Central Valley, California. Hydrological Processes. 2006;20:1157–1175. [Google Scholar]
- Rains MC, Leibowitz SG, Cohen MJ, Creed IF, Golden HE, Jawitz JW, Kalla P, Lane CR, Lang MW, McLaughlin DL. Geographically Isolated Wetlands Are Part of the Hydrological Landscape. Hydrological Processes. 2016;30:153–160. [Google Scholar]
- Reaney SM, Bracken LJ, Kirkby MJ. The Importance of Surface Controls on Overland Flow Connectivity in Semi-Arid Environments: Results from a Numerical Experimental Approach. Hydrological Processes. 2013;28:2116–2128. [Google Scholar]
- Reese EG, Batzer DP. Do Invertebrate Communities in Floodplains Change Predictably Along a River's Length? Freshwater Biology. 2007;52:226–239. [Google Scholar]
- Reid MA, Reid MC, Thoms MC. Ecological Significance of Hydrological Connectivity for Wetland Plant Communities on a Dryland Floodplain River, Macintyre River, Australia. Aquatic Sciences. 2015;78:139–158. [Google Scholar]
- Robinson CT, Tockner K, Ward JV. The Fauna of Dynamic Riverine Landscapes. Freshwater Biology. 2002;47:661–677. [Google Scholar]
- Rooney RC, Carli C, Bayley SE. River Connectivity Affects Submerged and Floating Aquatic Vegetation in Floodplain Wetlands. Wetlands. 2013;33:1165–1177. [Google Scholar]
- Sass GZ, Creed IF. Characterizing Hydrodynamics on Boreal Landscapes Using Archived Synthetic Aperture Radar Imagery. Hydrological Processes. 2008;22:1687–1699. [Google Scholar]
- Schalk CM, Luhring TM. Vagility of Aquatic Salamanders: Implications for Wetland Connectivity. Journal of Herpetology. 2010;44:104–109. [Google Scholar]
- Schemel LE, Sommer TR, Muller-Solger AB, Harrell WC. Hydrologic Variability, Water Chemistry, and Phytoplankton Biomass in a Large Floodplain of the Sacramento River, Ca, USA. Hydrobiologia. 2004;513:129–139. [Google Scholar]
- Schindler DE, Leavitt PR, Brock CS, Johnson SP, Quay PD. Marine-Derived Nutrients, Commercial Fisheries, and Production of Salmon and Lake Algae in Alaska. Ecology. 2005;86:3225–3231. [Google Scholar]
- Schofield KA, Alexander LC, Ridley CE, Vanderhoof MK, Fritz KM, Autry BC, DeMeester JE, Kepner WG, Lane CR, Leibowitz SG, Pollard AI. Biota Connect Aquatic Habitats Throughout Freshwater Ecosystem Mosaics. Journal of the American Water Resources Association. doi: 10.1111/1752-1688.12634. submitted. (this issue) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott DT, Keim RF, Edwards BL, Jones CN, Kroes DE. Floodplain Biogeochemical Processing of Floodwaters in the Atchafalaya River Basin During the Mississippi River Flood of 2011. Journal of Geophysical Research: Biogeosciences. 2014;119:537–546. [Google Scholar]
- Selvendiran P, Driscoll CT, Bushey JT, Montesdeoca MR. Wetland Influence on Mercury Fate and Transport in a Temperate Forested Watershed. Environmental Pollution. 2008;154:46–55. doi: 10.1016/j.envpol.2007.12.005. [DOI] [PubMed] [Google Scholar]
- Semlitsch RD, Bodie JR. Biological Criteria for Buffer Zones around Wetlands and Riparian Habitats for Amphibians and Reptiles. Conservation Biology. 2003;17:1219–1228. [Google Scholar]
- Shaw DA, Pietroniro A, Martz LW. Topographic Analysis for the Prairie Pothole Region of Western Canada. Hydrological Processes. 2013;27:3105–3114. [Google Scholar]
- Simon RN, Tormos T, Danis PA. Very High Spatial Resolution Optical and Radar Imagery in Tracking Water Level Fluctuations of a Small Inland Reservoir. International Journal of Applied Earth Observation and Geoinformation. 2015;38:36–39. [Google Scholar]
- Smock LA. Movements of Invertebrates between Stream Channels and Forested Floodplains. Journal of the North American Benthological Society. 1994;13:524–531. [Google Scholar]
- Sobota DJ, Gregory SV, Van Sickle J. Riparian Tree Fall Directionality and Modeling Large Wood Recruitment to Streams. Canadian Journal of Forest Research. 2006;36:1243–1254. [Google Scholar]
- Spence C, Phillips RW. Refining Understanding of Hydrological Connectivity in a Boreal Catchment. Hydrological Processes. 2015;29:3491–3503. [Google Scholar]
- Stanford JA, Ward JV. The Effects of Regulation on the Limnology of the Gunnison River: A North American Case History. In: Lillehammer A, Salveit SJ, editors. Regulated Rivers. Proceedings of the Second International Symposium on Regulated Streams, Oslo, August 1982. Universitetsforlaget AS; Oslo, Norway: 1984. pp. 467–480. [Google Scholar]
- Steiger J, Tabacchi E, Dufour S, Corenblit D, Peiry JL. Hydrogeomorphic Processes Affecting Riparian Habitat within Alluvial Channel–Floodplain River Systems: A Review for the Temperate Zone. River Research and Applications. 2005;21:719–737. [Google Scholar]
- Steward AL, von Schiller D, Tockner K, Marshall JC, Bunn SE. When the River Runs Dry: Human and Ecological Values of Dry Riverbeds. Frontiers in Ecology and the Environment. 2012;10:202–209. [Google Scholar]
- Stieglitz M, Shaman J, McNamara J, Engel V, Shanley J, Kling GW. An Approach to Understanding Hydrologic Connectivity on the Hillslope and the Implications for Nutrient Transport. Global Biogeochemical Cycles. 2003;17:16-l–16-15. [Google Scholar]
- Strahler AN. Quantitative Analysis of Watershed Geomorphology. Transactions of the American Geophysical Union. 1957;38:913–920. [Google Scholar]
- Subalusky AL, Fitzgerald LA, Smith LL. Ontogenetic Niche Shifts in the American Alligator Establish Functional Connectivity between Aquatic Systems. Biological Conservation. 2009;142:1507–1514. [Google Scholar]
- Tank JL, Rosi-Marshall EJ, Griffiths NA, Entrekin SA, Stephen ML. A Review of Allochthonous Organic Matter Dynamics and Metabolism in Streams. Journal of the North American Benthological Society. 2010;29:118–146. [Google Scholar]
- Tarboton DG. A New Method for the Determination of Flow Directions and Upslope Areas in Grid Digital Elevation Models. Water Resources Research. 1997;33:309–319. [Google Scholar]
- Thorp JH, Thoms MC, Delong MD. The Riverine Ecosystem Synthesis: Biocomplexity in River Networks across Space and Time. River Research and Applications. 2006;22:123–147. [Google Scholar]
- Tihansky AB. Sinkholes, West-Central Florida. In: Galloway D, Jones DR, Ingebritsen SE, editors. Land Subsidence in the United States. USGS Circular 1182. U.S. Department of the Interior, U.S. Geological Survey; Reston, VA: 1999. pp. 121–140. [Google Scholar]
- Tiner RW. Estimated Extent of Geographically Isolated Wetlands in Selected Areas of the United States. Wetlands. 2003;23:636–652. [Google Scholar]
- Tockner K, Malard F, Ward JV. An Extension of the Flood Pulse Concept. Hydrological Processes. 2000;14:2861–2883. [Google Scholar]
- Tromp-van Meerveld HJ, McDonnell JJ. Threshold Relations in Subsurface Stormflow 2. The Fill and Spill Hypothesis. Water Resources Research. 2006;42:W02411. [Google Scholar]
- Tronstad LM, Tronstad BP, Benke AC. Aerial Colonization and Growth: Rapid Invertebrate Responses to Temporary Aquatic Habitats in a River Floodplain. Journal of the North American Benthological Society. 2007;26:460–471. [Google Scholar]
- U.S. EPA. EPA/600/R-14/475F. Office of Research and Development, U.S. Environmental Protection Agency; Washington, D.C.: 2015. Connectivity of Streams and Wetlands to Downstream Waters: A Review and Synthesis of the Scientific Evidence. [Google Scholar]
- Uden DR, Hellman ML, Angeler DG, Allen CR. The Role of Reserves and Anthropogenic Habitats for Functional Connectivity and Resilience of Ephemeral Wetlands. Ecological Applications. 2014;24:1569–1582. doi: 10.1890/13-1755.1. [DOI] [PubMed] [Google Scholar]
- Valiela I, Teal JM. The Nitrogen Budget of a Salt Marsh Ecosystem. Nature. 1979;280:652–656. [Google Scholar]
- Van Meter KJ, Basu NB. Signatures of Human Impact: Size Distributions and Spatial Organization of Wetlands in the Prairie Pothole Landscape. Ecological Applications. 2014;25:451–465. doi: 10.1890/14-0662.1. [DOI] [PubMed] [Google Scholar]
- Vanderhoof MK, Alexander LC. The Role of Lake Expansion in Altering the Wetland Landscape of the Prairie Pothole Region, United States. Wetlands. 2016;36(Suppl 2):S309–S321. doi: 10.1007/s13157-015-0728-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderhoof MK, Alexander LC, Todd MJ. Temporal and Spatial Patterns of Wetland Extent Influence Variability of Surface Water Connectivity in the Prairie Pothole Region, United States. Landscape Ecology. 2016;31:805–824. [Google Scholar]
- Vanderhoof MK, Christensen JR, Alexander LC. Patterns and Drivers for Wetland Connections in the Prairie Pothole Region, United States. Wetlands Ecology and Management. 2017a;25:275–297. doi: 10.1007/s11273-016-9516-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vanderhoof MK, Distler HE, Lang MW, Alexander LC. The Influence of Data Characteristics on Detecting Wetland/Stream Surface-Water Connections in the Delmarva Peninsula, Maryland and Delaware. Wetlands Ecology and Management. 2017b doi: 10.1007/s11273-017-9554-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE. The River Continuum Concept. Canadian Journal of Fisheries and Aquatic Sciences. 1980;37:130–137. [Google Scholar]
- Vilizzi L, McCarthy BJ, Scholz O, Sharpe CP, Wood DB. Managed and Natural Inundation: Benefits for Conservation of Native Fish in a Semi-Arid Wetland System. Aquatic Conservation: Marine and Freshwater Ecosystems. 2013;23:37–50. [Google Scholar]
- Vining KC. Water-Resources Investigations Report 02-4113. U.S. Geological Survey; Bismarck, ND: 2002. Simulation of Streamflow and Wetland Storage, Starkweather Coulee Subbasin, North Dakota, Water Year 1981–98. [Google Scholar]
- Wallace JB, Whiles MR, Eggert S, Cuffney TF, Lugthart GH, Chung K. Long-Term Dynamics of Coarse Particulate Organic Matter in Three Appalachian Mountain Streams. Journal of the North American Benthological Society. 1995;14:217–232. [Google Scholar]
- Ward JV. The Four-Dimensional Nature of Lotic Ecosystems. Journal of the North American Benthological Society. 1989;8:2–8. [Google Scholar]
- Ward JV, Stanford JA. The Serial Discontinuity Concept of Lotic Ecosystems. In: Fontaine TD III, Bartell SM, editors. Dynamics of Lotic Ecosystems. Ann Arbor Science Publishers; Ann Arbor, MI: 1983. pp. 29–42. [Google Scholar]
- Webster JR, Patten BC. Effects of Watershed Perturbation on Stream Potassium and Calcium Dynamics. Ecological Monographs. 1979;49:51–72. [Google Scholar]
- Whigham DF, Chitterling C, Palmer B. Impacts of Freshwater Wetlands on Water Quality: A Landscape Perspective. Environmental Management. 1988;12:663–671. [Google Scholar]
- Whiting PJ, Pomeranets M. A Numerical Study of Bank Storage and Its Contribution to Streamflow. Journal of Hydrology. 1997;202:121–136. [Google Scholar]
- Wigington PJ, Griffith SM, Field JA, Baham JE, Horwath WR, Owen J, Davis JH, Rain SC, Steiner JJ. Nitrate Removal Effectiveness of a Riparian Buffer Along a Small Agricultural Stream in Western Oregon. Journal of Environmental Quality. 2003;32:162–170. doi: 10.2134/jeq2003.1620. [DOI] [PubMed] [Google Scholar]
- Wigington PJ, Leibowitz SG, Comeleo RL, Ebersole JL. Oregon Hydrologic Landscapes: A Classification Framework. Journal of the American Water Resources Association. 2013;49:163–182. [Google Scholar]
- Wigington PJ, Moser TJ, Lindeman DR. Stream Network Expansion: A Riparian Water Quality Factor. Hydrological Processes. 2005;19:1715–1721. [Google Scholar]
- Winter TC. The Concept of Hydrologic Landscapes. Journal of the American Water Resources Association. 2001;37:335–349. [Google Scholar]
- Winter TC. The Role of Ground Water in Generating Streamflow in Headwater Areas and in Maintaining Base Flow. Journal of the American Water Resources Association. 2007;43:15–25. [Google Scholar]
- Winter TC, Harvey JW, Franke OL, Alley WM. USGS Circular 1139. U.S. Government Printing Office; Washington, DC: 1998. Ground Water and Surface Water: A Single Resource. [Google Scholar]
- Wipfli MS, Gregovich DP. Export of Invertebrates and Detritus from Fishless Headwater Streams in Southeastern Alaska: Implications for Downstream Salmonid Production. Freshwater Biology. 2002;47:957–969. [Google Scholar]
- Wiskow E, van der Ploeg RR. Calculation of Drain Spacings for Optimal Rainstorm Flood Control. Journal of Hydrology. 2003;272:163–174. [Google Scholar]
- Wolf KL, Noe GB, Ahn C. Hydrologic Connectivity to Streams Increases Nitrogen and Phosphorus Inputs and Cycling in Soils of Created and Natural Floodplain Wetlands. Journal of Environmental Quality. 2013;42:1245–1255. doi: 10.2134/jeq2012.0466. [DOI] [PubMed] [Google Scholar]
- Wolock DM, Winter TC, McMahon G. Delineation and Evaluation of Hydrologic-Landscape Regions in the United States Using Geographic Information System Tools and Multivariate Statistical Analyses. Environmental Management. 2004;34:S71–S88. doi: 10.1007/s00267-003-5077-9. [DOI] [PubMed] [Google Scholar]
- Woodford DJ, McIntosh AR. Evidence of Source-Sink Metapopulations in a Vulnerable Native Galaxiid Fish Driven by Introduced Trout. Ecological Applications. 2010;20:967–977. doi: 10.1890/08-1909.1. [DOI] [PubMed] [Google Scholar]
- Yang W, Wang X, Liu Y, Gabor S, Boychuk L, Badiou P. Simulated Environmental Effects of Wetland Restoration Scenarios in a Typical Canadian Prairie Watershed. Wetlands Ecology and Management. 2010;18:269–279. [Google Scholar]
- Yurek S, DeAngelis DL, Trexler JC, Klassen JA, Larsen LG. Persistence and Diversity of Directional Landscape Connectivity Improves Biomass Pulsing in Simulations of Expanding and Contracting Wetlands. Ecological Complexity. 2016;28:1–11. [Google Scholar]
- Zilli FL, Paggi AC. Ecological Responses to Different Degrees of Hydrologic Connectivity: Assessing Patterns in the Bionomy of Benthic Chironomids in a Large River-Floodplain System. Wetlands. 2013;33:837–845. [Google Scholar]