

Abstract
The syntaxin family of soluble N-ethyl maleimide sensitive factor adaptor protein receptors (SNAREs) is known to play an important role in the fusion of transport vesicles with specific organelles. Twenty-four syntaxins are encoded in the genome of the model plant Arabidopsis thaliana. These 24 genes are found in 10 gene families and have been reclassified as syntaxins of plants (SYPs). Some of these gene families have been previously characterized, with the SYP2-type syntaxins being found in the prevacuolar compartment (PVC) and the SYP4-type syntaxins on the trans-Golgi network (TGN). Here we report on two previously uncharacterized syntaxin groups. The SYP5 group is encoded by a two-member gene family, whereas SYP61 is a single gene. Both types of syntaxins are localized to multiple compartments of the endomembrane system, including the TGN and the PVC. These two groups of syntaxins form SNARE complexes with each other, and with other Arabidopsis SNAREs. On the TGN, SYP61 forms complexes with the SNARE VTI12 and either SYP41 or SYP42. SYP51 and SYP61 interact with each other and with VTI12, most likely also on the TGN. On the PVC, a SYP5-type syntaxin interacts specifically with a SYP2-type syntaxin, as well as the SNARE VTI11, forming a SNARE complex likely involved in TGN-to-PVC trafficking.
INTRODUCTION
Vesicle trafficking is an essential function in all eukaryotic cells (reviewed in Sanderfoot and Raikhel, 1999). The formation, targeting, and fusion of a transport vesicle with its correct target membrane are required for movement of cargo between the various endomembrane organelles, as well as for recovery of proteins that have escaped their resident organelle. A class of proteins called syntaxins is required on the target membrane for selection and fusion of vesicles. Syntaxins are a part of a larger structural group of proteins called soluble N-ethyl malemide sensitive factor adaptor protein receptors (SNAREs), all of which are membrane proteins that contain a conserved coiled-coil domain (called a SNARE helix) proximal to a C-terminal transmembrane domain or lipid anchor. The function of the SNAREs in membrane fusion has been recently made clear in a series of articles (Fukuda et al., 2000; McNew et al., 2000; Parlati et al., 2000). Syntaxins are involved in recognizing and complexing with one or two other SNAREs on the target membrane creating a three-helix bundle called the t-SNARE complex. This tertiary complex is then primed to recognize a fourth helix derived from a v-SNARE, which resides on the vesicle membrane. The creation of the four-helix bundle, called a trans-SNARE complex or “SNAREpin,” leads eventually to vesicle fusion with the target membrane and delivery of the cargo into the lumen or limiting membrane of a particular organelle.
Analysis of the genome of the model plant Arabidopsis thaliana (The Arabidopsis Genome Initiative, 2000) revealed 24 genes that encoded members of the syntaxin family, each recently being reclassified with the name “syntaxin of plants” or SYP (Sanderfoot et al., 2000). These 24 genes comprise 10 gene families that each contain one to five members (Sanderfoot et al., 2000). The presence of the gene families does not necessarily indicate genetic redundancy, because gene disruptions of single members of three gene families lead to a lethal phenotype. The knolle mutation (in the SYP111 gene) results in a seedling lethal phenotype (Lukowitz et al. 1996) despite the presence of a second gene in the SYP11 gene family. Disruption of either SYP21 or SYP22 in the three member SYP2 gene family results in lethality at the gametophytic stage; a similar phenotype occurs within the SYP4 gene family when either SYP41 or SYP42 is disrupted (Sanderfoot et al., 2001). These results indicate that each syntaxin has unique essential functions, despite the fact that a high degree of sequence homology exists within a particular gene family.
Distinct types of syntaxins are found resident on the various membranes of the secretory pathway. The Arabidopsis syntaxins that have been characterized appear to be distributed similar to their yeast or mammalian orthologs upon the endomembrane compartments of the plant cell. The Arabidopsis SYP2-type syntaxins SYP21 and SYP22 are both found on the prevacuolar compartment (PVC; Conceição et al., 1997; Sanderfoot et al., 1998, 1999), although others have reported that SYP22 may be found on the tonoplast in the shoot apical meristem (Sato et al., 1997). This localization is consistent with the PVC or endosomal localization of the SYP2-type orthologs from yeast (Pep12p; Becherer et al., 1996) and mammals (syntaxins 7 and 13; Prekeris et al., 1999). Similarly, the Arabidopsis SYP4-type syntaxins and their orthologs from yeast (Tlg2p) and mammals (syntaxin 16) have all been localized to the trans-Golgi network (TGN) (Holthuis et al., 1998; Simonsen et al., 1998; Bassham et al., 2000). Interestingly, the two most divergent members (SYP41 and SYP42) are found on distinct domains of the TGN (Bassham et al., 2000), suggesting distinct functions for each in vesicle trafficking at the plant TGN.
To further our studies on vesicle trafficking, we have begun to characterize syntaxin gene families that may have roles in the late secretory/endosomal system of Arabidopsis cells. Here, we report that both SYP51 and SYP61 are found on multiple organelles in the late secretory system, and interact with other syntaxins and SNAREs of the VTI1-type. On the TGN, SYP61 forms separate complexes with the SNARE VTI12 and either SYP41 or SYP42, as well as a distinct complex with SYP51 and a member of the VTI1-type SNAREs. SYP5-type syntaxins show an interaction with SYP2-type syntaxins on the PVC. In addition, because both SYP21 and SYP51 interact with the SNARE VTI11, we propose that these three proteins are part of a SNARE complex on the PVC involved in TGN-to-PVC trafficking.
MATERIALS AND METHODS
Cloning of New Arabidopsis SYP Genes
A recent examination of the completed genome sequence of Arabidopsis thaliana revealed 24 syntaxins representing 10 gene families and 8 sequence-homology groups (Sanderfoot et al., 2000). Three of the gene families were most homologous to the yeast syntaxin Tlg1p and were further examined. The SYP5 gene family was represented by two genes, SYP51 (identified as open reading frame [ORF] F3O9.4, At1g15930; The Arabidopsis Genome Initiative, 2000) and SYP52 (F20B17.2, At1g73260), each of which was represented by expressed sequence tags (ESTs), although none the ESTs for SYP52 were full length. A full-length clone of SYP52 was acquired from a cDNA library. The SYP6 gene family was represented by a single gene, SYP61 (F3 M18.7, At1g27550). No full-length EST was found for this gene, so the ORF was acquired by reverse transcriptase-polymerase chain reaction (RT-PCR) with the use of specific primers (61NdeF: CATATGTTCTCAGCTCAAGATCCATTC; 61SacIR: GAGCTCTGTTCTTTAGGTCAAGAAGAC). The SYP7 family was represented by three genes: SYP71 (F11F8.33, At3g09740), SYP72 (F18N11.40, At3g45280), and SYP73 (F2A19.50, At3g61450). The full-length cDNA of SYP71 was acquired from an EST, whereas the other two members of this gene family were acquired by RT-PCR with the use of gene-specific primers (72F: GAGACATTCCACAAGAAGAAGAAGAAG; 72R: CCAGAAGAATTTCGGAGTGGGACATC; 73F: CCTGAAAAAAACCTTTCGTCGGAGAAATCT; 73R: GTGTAATGTAAATGACAGGCATCACCA). The full sequence of each cDNA clone was determined at the Michigan State University Sequencing Center (East Lansing, MI). The sequences of each of these SYPs have been deposited in GenBank under the following accession numbers: SYP51, AF355755; SYP52, AF355756; SYP61, AF355754; SYP71, AF355757; SYP72, AF355758; and SYP73, AF355759.
To examine the expression pattern of the SYP genes, total RNA was extracted from various tissues of Arabidopsis (roots, stems, leaves, flowers, siliques, and total seedlings). Equal amounts of total RNA were reverse transcribed with the use of oligo-dT as a primer under a manufacturer's protocol (Invitrogen, Carlsbad, CA). These cDNAs were subsequently used in PCR with gene-specific primers for each of the Arabidopsis SYP genes. The PCR products were separated by agarose gels and the amount of the appropriate sized product visually quantified. Total genomic DNA was used as a control for the specificity and fidelity of the primers, and products were also sequenced to confirm amplification. As a control, the pattern of SYP21 and SYP22 expression seen by RT-PCR matched that seen previously by Northern analysis (Bassham et al., 1995; Sato et al., 1997), suggesting that this method was adequate for qualitatively judging the expression pattern of these genes.
Bacterial Overexpression and Antisera Production
The full-length cDNAs of the SYPs were each further modified by PCR with the use of a specific primer for each, which included an NdeI restriction site immediately before the start codon of each open reading frame (51NdeF: AGATCTCATATGGCGTCTTCATCGGATTC; 61NdeF listed above; 71NdeF: CATATGACTGTGATCGATATTCTGACT; reverse primers were same as listed above), and the amplified products of each were cloned into the pGEM-T Easy vector (Promega, Madison, WI), creating pNde-SYP51, pNde-SYP61, and pNde-SYP71. The sequence of each plasmid was verified by dideoxy sequencing.
To produce antibodies to each of these classes of syntaxins, N-terminal fragments of SYP51, 61, and 71 (encoding amino acids 1–189, 1–168, and 1–144, respectively) were fused to a C-terminal 6xHis tag, and separately expressed in Escherichia coli stain BL21 (DE3) pLysS. Protein was purified by Ni-affinity chromatography, dialyzed, and concentrated before injection into rabbits at Cocalico Biologicals (Reamstown, PA).
Glutathione-S-transferase (GST) fusions of SYP51, 61, and 71 were created similarly by fusing amino acids 1–189, 1–168, and 1–236, respectively, to the C terminus of GST, and expressed in E. coli stain BL21. Protein was bound to a glutathione-affinity chromatography column, washed extensively, and then cross-linked with dimethyl pimelimidate as previously described (Bassham et al., 2000). Antisera raised against SYP51, 61, or 71 were affinity purified over the appropriate columns with the use of a procedure described previously (Bassham et al., 2000).
To test the specificity of the antibodies, we produced full-length fusions of several Arabidopsis SYPs in bacteria. pNde-SYP21 (previously called pNde-AtPEP12) is described in Sanderfoot et al. (1999), whereas pNde-SYP31 (previously called pNde-AtSED5) and pNdeSYP41 (previously called pNde-AtTLG2a) are described in Bassham et al. (2000). Each was cloned into an appropriately digested pET-28a (Novagen, Madison, WI) creating an N-terminal 6xHis fusion to each full-length ORF. The pET-28a-derived fusions were each transformed into E. coli stain BL21 (DE3) pLysS, and protein overexpressed by induction with isopropyl β-d-thiogalactoside. All of the 6xHis-tagged SYPs were soluble and could be purified by Ni-affinity chromatography. The purified proteins were separated by SDS-PAGE, transferred to nitrocellulose, and then probed with the various antisera raised to the Arabidopsis SYPs. Each antiserum recognized its cognate antigen, but no cross-reactivity was seen with noncognate SYPs, indicating the antiserum was specific for the products of its particular gene family.
SYP21 chicken antibodies are described in Sanderfoot et al. (2001), rabbit antisera specific to either SYP21 or SYP22 are described in Sanderfoot et al. (1999), and SYP41 rabbit antibodies are described in Bassham et al. (2000).
Epitope Tagging of SYP51 and 52
The plant binary plasmids pCAMBIA1300MCS1 and 3300MCS1 are derivatives of the pCAMBIA series of plasmids (Cambia GPO, Canberra, Australia) in which the endogenous multicloning site is replaced with the cauliflower mosaic virus 35S promoter and nopaline synthase terminator from pBI121 separated by a new multicloning site. These plasmids allow constitutive expression of proteins in plants after Agrobacterium-mediated transformation with selection based on either hygromycin (1300MCS1) or glufosinate (3300MCS1).
SYP51 was epitope tagged with a T7 peptide by cloning the BglII to NotI (found in the vector downstream of the stop codon) fragment of pNde-SYP51 into a BamHI to NotI digested pET-21a. This plasmid encodes for a T7-epitope tag fused to the N terminus of the full-length SYP51 ORF. The sequence of this plasmid was confirmed before cloning of the XbaI to XhoI fragment (containing the T7-SYP51 ORF) into XbaI and SalI digested pCAMBIA-3300MCS.
SYP52 was tagged with an CRUZ22 peptide (Santa Cruz Biotechnology, Santa Cruz, CA) by introducing an EcoRI site at the start of the ORF by PCR (52EcoRIF: GAATTCATGGCCTCTTCTTCGGATC, 52SacIR: CTCGAGCCACCACATTGGCATTACAGG), and then cloning the EcoRI to PstI fragment of the amplified product into the pCRUZ22B vector. A primer specific to the 5′ region of the CRUZ22 tag preceded by an XbaI restriction site (TCTAGAATGGATATGCGCGACGCCCTG) was used with a primer within the SYP52 fragment (52R2: ACATGGTAATCTAAGTCATCAATAAGCCT) in PCR to retrieve the tagged fragment from the pCRUZ22B plasmid. This amplified product was cloned into pGEM-T Easy, and the sequence confirmed before the XbaI to PstI fragment was rejoined with the rest of the SYP52 ORF (digested with PstI and XhoI) by ligating to XbaI and SalI digested pCAMBIA-1300MCS.
Both of the binary plasmids were transformed into Agrobacterium tumefaciens, and subsequently vacuum infiltrated into Arabidopsis ecotype Columbia as described previously (Bent et al., 1994). Transformants were screened based upon the appropriate selectable marker and expression of the epitope-tagged proteins verified by T7 monoclonal antibodies (Novagen) or CRUZ22 rabbit polyclonal antibodies (Santa Cruz Biotechnology).
Immunoprecipitation of Arabidopsis SYPs
Root cultured Arabidopsis plants (wild-type, T7-SYP51, C22-SYP52, or T7-SYP42-expressing) were grown as described previously (Bar-Peled et al., 1995). Detergent extracts of microsomes and subsequent immunoprecipitations (IPs) were performed essentially as described in Bassham et al. (2000). Immobilized antibodies to the CRUZ22 tag were acquired from Santa Cruz Biotechnology, and used in IPs in a manner similar to that described for immobilized T7 monoclonal antibodies (Bassham et al., 2000). Each IP was repeated at least twice from independent plants, and representative results are shown in the figures.
Similar extracts were made from suspension cultures of Arabidopsis cell lines (cultured as described in Conceição et al., 1997) and were used in similar experiments to those listed above, as well as for the large-scale IPs used in our proteomic approach. The details of this method will be described elsewhere in detail, but briefly, after elution from the immobilized antibody column, proteins were precipitated by 5% trichloroacetic acid, washed in cold acetone, and then resuspended in SDS sample buffer. After separation by SDS-PAGE, proteins were visualized by silver staining, and specific bands excised. Bands were provided to the Protein Microsequencing and Proteomic Mass Spectroscopy Lab at the University of Massachusetts Medical School (Worchester, MA). After in-gel tryptic digestion, eluted peptides were applied to a Finnigan Electrospray LCQ Deca ion trap mass spectrometer, and the resulting peptide masses used to identify potential matches within the Arabidopsis proteome.
Electron Microscopy
Cryosections of Arabidopsis root tips were prepared as described in Sanderfoot et al. (1998) and used for all immunogold labeling experiments. Immunolabeling was performed as described in Sanderfoot et al. (1998) and Zheng et al. (1999). For double-labeling experiments, after incubation of the grids with the first antibody, a second fixation step followed by a second blocking step was used to prevent cross-reactivity of the antibodies in later stages of the protocol. Many sections from independent plants were observed for each combination of antibodies. Controls were performed with the use of the corresponding preimmune serum substituted for one or both of the antisera. In all cases, these controls demonstrated that the labeling seen was highly specific.
RESULTS
SYP5- and SYP6-Group of Arabidopsis Syntaxins
Analysis of the completed Arabidopsis genome identified three syntaxin gene families (SYP5, SYP6, and SYP7) that had not been previously characterized (Sanderfoot et al., 2000). Based on secondary structure predictions (NNPredict; Kneller et al., 1990), the members of each of these groups of syntaxins have a secondary structure very similar to that of mammalian syntaxin 1, whose crystal structure has been solved (Misura et al., 2000). With respect to the eight yeast syntaxins, the members of each of these families showed homology with the late-Golgi/endosomal syntaxin Tlg1p. The SYP5 family contained two members (SYP51 and SYP52), whose gene products share 82% sequence identity and were most similar in sequence to mammalian syntaxin 8 (37% sequence identity; Figure 1A). SYP51/52 (as well as syntaxin 8) also shared ∼20% sequence identity with the yeast vacuolar SNARE Vam7p (Sato et al., 1998). However, the SYP5/syntaxin 8-type proteins lack the essential PX-domain found at the N terminus of Vam7p, whereas Vam7p does not contain a transmembrane domain at the C terminus like the SYP5/syntaxin8-type proteins. Comparisons between SYP51 and Vam7p with the PX-sequence deleted (i.e., the syntaxin-like domain) showed a higher level (28%) of sequence identity. The SYP6-group is encoded by a single gene (SYP61), the product of which shows highest sequence identity to mammalian syntaxins 6 and 10 (16 and 20% sequence identity, respectively), as well as yeast Tlg1p (19.2% sequence identity). The SYP7-gene family encompasses three genes that show ∼15% sequence identity to Tlg1p, and lower identity with the mammalian syntaxins listed above. SYP7-type syntaxins show no obvious mammalian homologs and may be a distinct subgroup of Tlg1p-like syntaxins.
Figure 1.

SYP5- and SYP6-type syntaxins of Arabidopsis. (A) Arabidopsis SYP51 (AF355755), SYP52 (AF355756), SYP61 (AF355754) human syntaxin 6 (O43752), syntaxin 8 (AF115323), syntaxin 10 (AF035531), yeast Tlg1p (NP_010756), and Vam7p (NP_011303) were aligned with the CUSTAL W algorithm. The aligned sequences were subjected to secondary structure predictions (NNPredict; Kneller et al., 1990). A comparison of the predicted secondary structures to the solved crystal structure of mammalian syntaxin 1 (Misura et al., 2000) produced the schematic diagrams of the predicted structure of the five proteins: Ha, Hb, and Hc are three N-terminal α-helices found in all syntaxins examined thus far, SNARE indicates the conserved coiled-coil domain found in all SNAREs, TM indicates the transmembrane helix found in the syntaxins, PX indicates the NADPH oxidase p40phox domain found at the N terminus of Vam7p. The percentage of amino acid identity of SYP51 to related syntaxins is shown above the schematic of each protein (percentage of identity of SYP51 and the SNARE-like domain of Vam7p shown in parentheses), whereas the percentage of identity of SYP61 to related syntaxins is shown below the schematics. Phylogenetic trees resulting from this alignment can be found at http://www.msu.edu/∼sanderfo/atsnare.htm. (B) Microsomal extracts of Arabidopsis root were separated by SDS-PAGE and transferred to nitrocellulose. Individual lanes were probed with affinity-purified rabbit antisera raised against Arabidopsis SYP51, SYP61, and SYP71. (C) Microsomal extracts were prepared from tissue extracted from each of the major organs of an Arabidopsis plant. Approximately equal amounts of total microsomal protein were separated by SDS-PAGE, transferred to nitrocellulose, and probed with antisera specific to the indicated Arabidopsis syntaxins. SYP21 is shown as a control because its expression is very consistent across different tissues.
Based upon RT-PCR analysis, members of the SYP5- and SYP7-families and SYP61 are expressed widely in plant tissues. Like other previously characterized Arabidopsis syntaxins (Bassham et al., 1995; Sato et al., 1997), these genes typically had their highest expression levels in roots and seedlings, but detectable levels were observed in all other tissues (our unpublished results). Analysis of the EST databases from other plants revealed that orthologs of these syntaxins are found in other dicotyledonous plants such as soybean (Glycine max), barrel medic (Medicago truncatula), and tomato (Lycopersicon esculentum), as well as several monocots (Zea mays, Triticum aestivum, Sorghum bicolor), indicating that Arabidopsis is typical with respect to these gene families (see http://www.msu.edu/∼sanderfo/atsnare.htm).
To better study these syntaxins, a polyclonal antiserum was raised to an N-terminal fragment of SYP51. The affinity-purified SYP51 antibodies recognized a single band of ∼30 kDa (Figure 1B) that was found in microsomes extracted from all major tissues of Arabidopsis (Figures 1C). The molecular mass of the SYP51-cross-reacting band was of a larger apparent size than predicted, but this is typical of syntaxins, which often migrate as larger proteins in SDS-PAGE (Conceição et al., 1997; Bassham et al., 2000). The antiserum was found to cross-react with both SYP5-type syntaxins, and this cross-reactivity could not be reduced by affinity purification procedures (our unpublished results). This was not surprising considering the high degree of sequence homology between SYP51 and SYP52. Because both SYP51 and SYP52 have a similar expression pattern, the bands observed in Figure 1, B and C, are likely to be a summation of both proteins. Antisera were also raised to N-terminal fragments of SYP61 and SYP71. These antisera recognized bands of ∼35 and ∼36 kDa (respectively; Figure 1B) in microsomes from all major organs (Figure 1C). The SYP71-antiserum is likely to recognize all three of the SYP7-type syntaxins. The specificity of all the antisera was checked against bacterially expressed Arabidopsis syntaxins (see MATERIALS AND METHODS), and each was found to be specific to its cognate antigen.
In examining the localization and protein–protein complexes between these groups of syntaxins, we observed an overall similarity between the SYP5- and SYP6-type syntaxins that was not shared with the SYP7-group (see below). For this reason, we will focus here on the SYP5- and SYP6-type syntaxins, whereas SYP7-type syntaxins will be covered in more detail elsewhere.
SYP51 Is Found on PVC and SYP42-Domain of TGN
To examine the intracellular localization of SYP51, cryosections of Arabidopsis roots were probed with affinity-purified antibodies specific to SYP51 (Figure 2). Labeling with the preimmune serum to each protein showed no significant staining of any organelles (Figure 2, A and E). Use of the SYP51 immune serum showed a significant labeling of the TGN region of the Golgi (Figure 2B). No significant labeling of the nuclei, endoplasmic reticulum, plasma membrane, cell wall, or of the vacuole was seen with SYP51 antibodies (our unpublished results). In addition, gold label was seen on other organelles that were found distal to the Golgi and occasionally in perivacuolar regions (Figure 2, C and D) that resembled the PVC. This was confirmed by performing double immunolabeling with antisera specific for the PVC marker SYP21. Cryosections of Arabidopsis roots were stained with affinity-purified SYP51 antibodies, followed by 10 nm of gold; then with rabbit affinity-purified SYP21 antibodies, followed by 5 nm of gold. As a control, preimmune antiserum was used in place of one or both (Figure 2, E and F) of the antisera. As has been found previously (Conceição et al., 1997; Sanderfoot et al., 1998, 1999), the SYP21 antibodies was found only on the PVC, electron-dense organelles that lie either proximal to the Golgi stacks or are found in the perivacuolar region (Figure 2, G and H). Many of the PVC structures stained by SYP21 antibodies were also labeled by the SYP51 antibodies (Figure 2, G and H, and Table 1). Occasionally, PVC structures stained with only SYP21 were observed, and SYP51 was found alone, especially over the Golgi stacks. Identical results were found in double immunolabeling when SYP22 antiserum was used in place of SYP21 (Table 1). All of the colocalization between SYP22 and SYP51 was found over prevacuolar-like structures, which is consistent with the colocalization of SYP21 and SYP22 on the PVC in these root cells (Sanderfoot et al., 1999). SYP22 may be found on other organelles such as the tonoplast in other cell types (Sato et al., 1997; Rojo, Zouhar, and Raikhel, unpublished results), although we have no evidence for SYP51 being found on organelles other than the PVC or TGN in any cell type (our unpublished results).
Figure 2.

SYP51 is found on the TGN and PVC. Cryosections of Arabidopsis root cells were immunolabeled with either SYP51 preimmune (A), affinity-purified SYP51 antibodies (B–D) followed by detection with 10 nm of colloidal gold. Double immunolabeling was performed with the use of cryosections from wild-type (E–H), or HA-SYP41 and T7-SYP42-expressing (I–M) roots. In all cases, SYP51 (or its preimmune) immunolabeling was detected with 10 nm of gold, whereas the second antibody was detected with 5 nm of gold (as indicated in the lower right on each panel). SYP51 preimmune (E and I) or affinity-purified antibodies (G and H, J–M, and insets) were detected with 10 nm of gold. A second labeling was then preformed with SYP21 preimmune (E and F) or affinity-purified antibodies (G and H, and inset), with a nonspecific control (I), T7-monoclonal antibodies (J and K), or HA-monoclonal antibodies (L and M), each detected with 5 nm of gold. All images were captured at the same magnification. G, Golgi; V, vacuole. Bar in A, 100 nm. Arrowheads are used to highlight the 5-nm gold particles.
Table 1.
Quantification of immunogold localizations For each combination of proteins shown, the number of Golgi stacks or prevacuolar structures where colocalization was seen (Together; defined as within 80 nm), with antibodies against epitope-tagged or endogenous proteins, and where the labeling was seen over different membranes or membrane domains (Separate) is indicated. The total number of sections examined for each combination is indicated in the Total column.
| Together | Separate | Total | |
|---|---|---|---|
| SYP21 + SYP51 | 48 | 10 | 58 |
| SYP22 + SYP51 | 26 | 11 | 37 |
| HA-SYP41 + SYP51 | 15 | 30 | 45 |
| T7-SYP42 + SYP51 | 18 | 8 | 26 |
| SYP21 + SYP61 | 19 | 9 | 28 |
| HA-SYP41 + SYP61 | 31 | 20 | 51 |
| T7-SYP42 + SYP61 | 44 | 20 | 64 |
| SYP51 + SYP61 | 40 | 16 | 56 |
SYP41 and SYP42 define distinct domains of the plant TGN (Bassham et al., 2000). Because SYP51 was found to label the TGN, we attempted to determine whether this syntaxin was restricted to a particular domain of the TGN (as defined by SYP41 or SYP42). We produced cryosections of plants expressing both HA-SYP41 and T7-SYP42. The sections were incubated with SYP51 antibodies detected with 10 nm of gold, followed by incubation with either HA- or T7-specific antibodies (for HA-SYP41 or T7-SYP42, respectively) detected with 5 nm of gold. Controls where preimmune serum was used in place of one or both of the antisera during the double labeling ensured that the staining was specific (Figure 2I). We found that SYP51 significantly colocalized with T7-SYP42 on the TGN (Figure 2, J and K, and Table 1), although SYP51 was found separate from T7-SYP42 in a small percentage of sections (Table 1). On the other hand, colocalization of SYP51 with HA-SYP41 was rare, with the majority of the labeling for each marker being found on distinct regions of the TGN (Figure 2, L and M, and Table 1).
SYP61 Is Found on TGN and PVC
In a similar manner as described above, we examined the localization of SYP61 with the use of cryosections and immunolabeling with affinity-purified SYP61 antibodies. SYP61 immunolabeling was found over the Golgi and TGN (Figure 3, B and C), and occasionally over regions distal from the Golgi (Figure 3D). No specific labeling with the SYP61 antibodies was found over the nuclei, endoplasmic reticulum, plasma membrane, cell wall, or the tonoplast membrane (our unpublished results). With the use of SYP21 as a marker for the PVC in double immunolabeling, we were able to show significant colocalization of SYP61 and SYP21 (Figure 3F and Table 1), indicating that SYP61 is also found on the PVC. When double labeling was performed on cryosections from the HA-SYP41- and T7-SYP42-expressing plants, we found that SYP61 colocalized with both SYP41 and SYP42 to approximately equal extent (Figure 3, G and H, and Table 1). These results suggested that SYP61 was distributed across both domains of the TGN.
Figure 3.

SYP61 is found on the Golgi and TGN. Cryosections of Arabidopsis root cells were immunolabeled with either SYP61 preimmune (A) or affinity-purified SYP61 antibodies (B–D) followed by detection with 10 nm of colloidal gold. Double immunolabeling was performed with the use of cryosections from wild-type (E, F, and I) or from HA-SYP41- and T7-SYP42-expressing roots (G and H). SYP61 preimmune (E) or affinity-purified antibodies were detected with 10 nm of gold (F–H) or with 5 nm of gold (I). A second labeling was then performed with nonspecific (E), SYP21 affinity-purified (F), HA-monoclonal (G), or T7-monoclonal (H) antibodies followed by 5 nm of gold, or with SYP51 affinity-purified antibodies followed by 10 nm of gold (I). All images are shown at the same magnification. G, Golgi; V, vacuole. Bar in A, 100 nm. Arrowheads are used to highlight the 5-nm gold particles.
Double immunolabeling with SYP51 and SYP61 (on sections of wild-type Arabidopsis) showed that both colocalized extensively (Figure 3I and Table 1). The colocalization occurred both near the Golgi as well as distal from the stacks (likely the PVC) with approximately equal extent. Meanwhile, some of the Golgi staining for SYP61 was unique, probably representing the pool of SYP61 found on the SYP41-domain of the TGN where SYP51 staining is rare. These results suggest that SYP51 and SYP61 colocalize both on the TGN and PVC.
Interactions between Arabidopsis SYPs
Because recent results have suggested that different syntaxins may interact in vivo (Holthuis et al., 1998; Antonin et al., 2000; Wade et al., 2001), we examined the potential interactions between Arabidopsis syntaxins with the use of IP of detergent extracts of Arabidopsis microsomes. Extracts of Arabidopsis root-cultured tissue were immunoprecipitated (IPd) with antisera to various SYPs, and then the eluates of these experiments were probed with the antisera indicated in Figure 4. Similar results were found if extracts from suspension cultured cell lines of Arabidopsis were used instead for the IPs (our unpublished results).
Figure 4.

Interaction between Arabidopsis SYPs and VTI1-type SNAREs. Detergent extracts of Arabidopsis microsomes (see MATERIALS AND METHODS) from wild-type (A and C), or T7-SYP42-expressing (B) root cultures were immunoprecipitated with affinity-purified chicken anti-SYP21, affinity-purified rabbit anti-SYP41, anti-SYP51, or anti-SYP61. (A) Equal amounts of total (T) and flowthrough (FT), as well as approximately threefold more eluate (E) were separated by SDS-PAGE and then probed with rabbit antibodies specific for the indicated SYPs. Asterisks indicate the SYP used in a particular IP. (B) Extracts of plants expressing T7-SYP42 were used for IPs as in A to better differentiate between SYP41 and SYP42. (C) Extracts of wild-type plants were immunoprecipitated as in A except that 10-fold more eluate was loaded to better detect the small amounts of VTI11-type SNAREs coimmunoprecipitated in these experiments. Coimmunoprecipitation of SYP41 and VTI12 (and not VTI11) has been previously described in detail in Bassham et al. (2000).
We found many distinct complexes between the TGN and PVC SYPs that are shown in Figure 4A. Use of SYP21-specific antibodies showed that SYP51 was coimmunoprecipitated, although no interaction was seen with other SYPs (including SYP22). A similar pattern was observed by IP with SYP22 antibodies (our unpublished results). This interaction was confirmed in reverse by the use of SYP51 antibodies to coimmunoprecipitated SYP21. The fact that SYP21 and SYP22 do not coimmunoprecipitate, yet SYP51 interacts with both indicates that two distinct SYP21 + SYP51 and SYP22 + SYP51 complexes must form. SYP51 also coimmunoprecipitated SYP61, and because no interaction was found between SYP21 and SYP61, the SYP51 + SYP61 interaction is again distinct. Use of SYP41 antibodies showed a coimmunoprecipitation with SYP61, but not with other SYPs. It should be noted that the SYP41 antibodies used here cross-react with SYP42 (Bassham et al., 2000). Extracts of T7-SYP42-expressing plants were used in parallel experiments to clarify any potential distinctions between SYP41 and SYP42, although none were identified in these experiments (Figure 4B). Because SYP41 and SYP42 are found on distinct domains of the TGN, and T7-SYP42 does not coimmunoprecipitate SYP41 (Bassham et al., 2000), it is clear that distinct SYP41 + SYP61 and SYP42 + SYP61 complexes must form. None of these SYPs were found to interact with SYP71, indicating that this group is distinct from the SYP5- or SYP6-type syntaxins. As should be expected, the complexes between the SYPs were restricted to those proteins that have been shown to colocalize (i.e., SYP21 + SYP51 or SYP41 + SYP61), although, importantly, not all SYPs that are found on the same organelle were found to interact (i.e., SYP51 does not coimmunoprecipitate SYP42, and SYP21 does not coimmunoprecipitate SYP61). In summary, our results indicated PVC-localized complexes of SYP21 + SYP51 and SYP22 + SYP51, TGN-localized complexes of SYP41 + SYP61, SYP42 + SYP61, and a fifth complex containing SYP51 + SYP61 that may be either on the PVC or the TGN.
As an independent verification of the above-mentioned results, we also used a proteomic approach to identify potential partners for the various SYPs. Much of this analysis will be presented elsewhere, but the following is of relevance to this work. We performed a large-scale IP of Arabidopsis suspension cell extract with SYP41 antibodies, followed by separation of the eluate by SDS-PAGE, silver staining, and excision of specific bands. Peptides were then subjected to in-gel tryptic digestion and mass identification with the use of electron spray ion trap mass spectroscopy. Aside from identifying peptides known from previous analysis (i.e., VPS45; Bassham et al., 2000), we were able to identify a peptide corresponding to SYP61 (195IGGVGLTIHDELVAQER211), as well as peptides corresponding to SYP41, SYP42, and SYP43 (recall that the SYP41 antibodies cross-react with other members of the SYP4-group; Bassham et al., 2000). Peptides from no other SYPs could be identified, consistent with the results presented above.
Interactions with Arabidopsis Vti1p-Homologs
Arabidopsis encodes three homologs of the yeast SNARE Vti1p (Zheng et al., 1999; Sanderfoot et al., 2000). VTI11 and VTI12 (formerly VTI1a and VTI1b) show the lowest degree of sequence identity (58%) among the Arabidopsis homologs, yet have a similar distribution pattern within the endomembrane system, both being found on the TGN and Golgi-derived vesicles, as well as on the PVC (Zheng et al., 1999). However, each is believed to serve distinct functions in vesicle trafficking based upon their differential ability to complement aspects of yeast Vti1p functions (Zheng et al., 1999). Furthermore, it has been previously shown that the members of the SYP4 family interact only with the VTI12 paralog, having no detectable interaction with VTI11 (Bassham et al., 2000). Members of the SYP2 family have previously been shown to interact with VTI11 (Sanderfoot et al., 1999; Zheng et al., 1999), and we now show that the PVC syntaxins coimmunoprecipitate very little VTI12 (Figure 4C). Thus, there seems to be a clear preference for VTI11 to form complexes with the PVC syntaxins, whereas VTI12 interacts with the TGN syntaxins.
To investigate any preference between VTI11 and VTI12 with respect to the other SYPs, we examined detergent extracts of Arabidopsis microsomes. In this experiment, the eluate was substantially overloaded with respect to the total and flow-through (10-fold) so as to allow comparison between the two VTI1 paralogs. SYP51 antibodies coimmunoprecipitated a much greater amount of VTI11 compared with VTI12 (Figure 4C), indicating a potentially greater affinity for VTI11. Unlike the SYP2-type syntaxins, however, some VTI12 was coimmunoprecipitated with SYP51. On the other hand, SYP61 antibodies coimmunoprecipitated a much greater amount of VTI12 than VTI11 (Figure 4C). Unlike the SYP4-type syntaxins, a small amount of VTI11 was coimmunoprecipitated by SYP61. Consistent with the previous results, VTI11 may preferentially form PVC-localized complexes (i.e., with SYP2- and SYP5-type syntaxins), whereas VTI12 preferentially forms TGN-localized complexes (i.e., with SYP4-, SYP5-, and SYP6-type syntaxins. These results further strengthen the hypothesis that VTI11 and VTI12 serve distinct functions in vesicle trafficking.
SYP51 and SYP52 May Have Redundant Functions
Because the SYP51 antibodies cross-reacted with SYP52, to clearly distinguish each member of the SYP5 gene family, we labeled each with a distinct epitope tag. SYP51 was epitope tagged with a T7-peptide (T7-SYP51), whereas SYP52 was tagged with a CRUZ22 epitope (C22-SYP52). Arabidopsis plants stably expressing both epitope-tagged proteins were produced by Agrobacterium-mediated transformation. Arabidopsis plants were chosen that expressed only a moderate amount of each epitope-tagged protein to prevent any potential problems due to overexpression. Based upon density gradient analysis of the transgenic plants, the epitope-tagged proteins were found to fractionate identically to that of the endogenous SYP5 (our unpublished results), suggesting that the epitope tags did not affect the intracellular localization.
To determine whether there was any difference between the two SYP5-type syntaxins, we made microsomal extracts from root cultures expressing T7-SYP51 and C22-SYP52, and used immobilized antibodies specific for each tag to immunoprecipitate them individually (Figure 5). The two SYP5-type syntaxins do not interact with each other, similar to what was found for the SYP2-type syntaxins mentioned above, indicating that distinct complexes must exist for SYP51 and SYP52. No other distinctions were found between the SYP5 paralogs with respect to interaction with other SYPs, and identical interactions were observed for the epitope-tagged versions as was found above for the polyclonal antiserum. Because of the high degree of sequence identity, together with an identical expression pattern and biochemical behavior, we believe that SYP51 and SYP52 may serve redundant functions, although this will require future analysis of gene disruptions.
Figure 5.
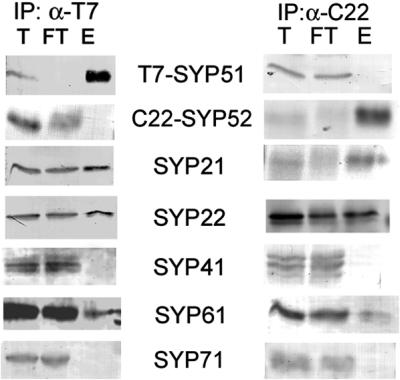
Use of epitope tags to differentiate SYP51 and SYP52. Microsomes were prepared from Arabidopsis plants expressing T7-SYP51 and C22-SYP52. Detergent extracts of the microsomes were IPd with either immobilized T7-monoclonal (left) or CRUZ22-rabbit polyclonal antibodies (right). Equal amounts of the total (T) and flowthrough (FT), as well as approximately threefold more eluate (E) were separated by SDS-PAGE and then probed with rabbit antisera specific for the indicated proteins.
DISCUSSION
In this work, we have examined the intracellular localization and protein–protein interactions of the SYP5- and SYP6-type syntaxins, previously undescribed groups of Arabidopsis syntaxins. Among the eight yeast syntaxins, both SYP5- and SYP6-type syntaxins show highest overall sequence identity with the early endosomal syntaxin Tlg1p. SYP61 shows further homology to the mammalian TGN-localized syntaxins 6 and 10 (Bock et al., 1997; Tang et al., 1998). SYP5-type syntaxins, also show similarity to the SNARE domain of the yeast vacuolar SNARE Vam7p (Sato et al., 1998) and to mammalian syntaxin 8, a syntaxin found on many organelles within the late endocytic system of the mammalian cell (Prekeris et al., 1999). Based upon these similarities, we propose the multicellular eukaryotes may have subgroups of the Tlg1p-like syntaxins: one including SYP61 and mammalian syntaxins 6 and 10, and a second of SYP51/52 and mammalian syntaxin 8.
The yeast syntaxin Tlg1p is required for the endocytosis and correct Golgi localization of many membrane proteins in yeast, and consistent with this role is generally reported to be localized to the equivalent of an early endosome in yeast cells, although it is also found on the TGN and the PVC (Holthuis et al., 1999; Gurunathan et al., 2000; Lewis et al., 2000). Yeast Vam7p is found mostly in the soluble fraction (Sato et al., 1998), although it is also found associated with the vacuolar membrane (Sato et al., 1998; Ungermann and Wickner, 1998). The Ustilago maydis ortholog of Vam7p (Yup1p) is found to localize to small vesicles and the equivalent of an early endosome (Wedlich-Söldner et al., 2000), suggesting that this SNARE may function on organelles other than the vacuole. Mammalian syntaxins 6 and 8 are each found on many organelles, including the TGN and early and late endosomes, as well as at lower levels on the plasma membrane and lysosomes (Bock et al., 1997; Klumperman et al., 1998; Prekeris et al., 1999). Here, we have shown that SYP51 and SYP61 are found on multiple organelles, including the TGN and PVC. Note that we have found no evidence of vacuolar or plasma membrane localization for either SYP51 or SYP61. The finding that, while on the TGN, SYP51 preferentially localizes with SYP42 (compared with SYP41) is further evidence that the TGN has distinct domains. Because SYP51 and SYP42 do not directly interact, these two syntaxins probably have distinct functions on this particular domain of the TGN. The vacuolar cargo receptor AtELP also preferentially localizes with SYP42 on this TGN domain (Bassham et al., 2000). At this point, it is unclear whether the colocalization of these markers to a particular domain of the TGN has any significance, but future studies may clarify the distinct roles of the domains of the TGN.
The yeast SNAREs Tlg1p and Vam7p interact with other syntaxins. For example, Tlg1p interacts with the TGN-localized syntaxin Tlg2p (Holthuis et al., 1998). Vam7p has been shown to interact with the vacuolar syntaxin Vam3p (Sato et al., 1998). With respect to the Tlg1p + Tlg2p interaction, we find one orthologous interaction between SYP41 or 42 + SYP61, but not between SYP41 or 42 + SYP51; perhaps indicating a functional distinction between the Arabidopsis SYPs. With respect to the Vam7p + Vam3p interaction, this becomes contentious. No other eukaryote aside from budding yeast has a clear ortholog of Vam3p (Pelham, 1999; Sanderfoot et al., 1999, 2000). Most commonly, the members of the plant SYP2-type syntaxins and mammalian syntaxins 7/13 are described as Pep12p-orthologs because they are found in most cells on prevacuolar or endosomal compartments (Prekeris et al., 1999; Sanderfoot et al., 1999). However, in some cells types, representatives of each of these groups (SYP22 and syntaxin 7) have been reported to be localized to the tonoplast or lysosomal membranes (Sato et al., 1997; Mullock et al., 2000), suggesting that perhaps particular members of these groups may also play similar roles as yeast Vam3p. It is therefore possible that the Vam7p + Vam3p interaction represents an analog of the SYP2 + SYP5 interaction we observe, or the orthologous syntaxin 7 + 8 interaction observed in mammalian cells by Antonin et al. (2000). We have also shown an interaction between SYP5-type syntaxins and SYP61. Thus far, no equivalent interaction between the mammalian orthologs of these syntaxins (syntaxin 8 and syntaxin 6) has been reported. On the contrary, a recent report has indicated an interaction between mammalian syntaxins 6 and 7 (Wade et al., 2001), an interaction we do not observe between the orthologous plant proteins (SYP2 and SYP6; Figure 4). Clearly, significant differences remain between plants and animals with respect to SNARE–SNARE interactions.
We have also identified other members of the SYP-complexes within the VTI1-like family of Arabidopsis SNAREs. Because both SYP2-type and SYP5-type syntaxins each interact with VTI11, and all three SNAREs can be found localized on the PVC (Conceição et al., 1997; Sanderfoot et al., 1999; Zheng et al., 1999), it seems reasonable to suggest that these three SNAREs may form a complex (SYP2 + SYP5 + VTI11). Because the amount of VTI11 IPd with either SYP21 or SYP51 is very small, it is possible that VTI11 may be acting as a v-SNARE for a SYP21 + SYP51 t-SNARE complex. It is typical that the amount of v-SNARE IPd from detergent extracts is quite small relative to other members of the t-SNARE complex (Holthuis et al., 1998; Antonin et al., 2000; Gurunathan et al., 2000; Lewis et al., 2000). Similar arguments can be made to show that there exist TGN complexes of SYP4 + SYP6 + VTI12 where VTI12 may act as a v-SNARE. In the case of the SYP5 + SYP6 complex, because both interact to some extent with both VTI11 and VTI12, it is not immediately apparent which VTI1-type SNARE may be a part of this complex. It is most likely that this complex contains VTI12, considering the very small amount of VTI11 coimmunoprecipitated by SYP61, but we cannot be sure at this time. With preliminary experiments attempting IPs with either VTI11 or VTI12 antibodies, we have not observed any interactions between VTI11 and VTI12 (our unpublished results), suggesting that the SYP5 + SYP6 complex must contain one or the other, not both. Regardless of the order of the tertiary complex between these SNAREs, a fourth SNARE is required to form a functional SNARE complex. Preliminary evidence indicates that none of the members of the VAMP72-group nor of the novel NPSN-group of SNAREs (Sanderfoot et al., 2000) are IPd by any of the above-mentioned syntaxins (our unpublished results), suggesting that these SNAREs are unlikely to be part of this complex. Many other SNAREs are found in Arabidopsis (Sanderfoot et al., 2000), and we are currently investigating the possibility that any of these may be a part of these complexes. At this point, little can be gained from examining the other members of the mammalian syntaxin complexes. For example, the SNAREs mVti1b and VAMP8 constitute the other members of the syntaxin 7 + 8 complex (Antonin et al., 2000). Although Arabidopsis encodes for three Vti1p-type SNAREs (including VTI11), all are more homologous to mVti1a as opposed to the much more divergent mVti1b (Zheng et al., 1999; Sanderfoot et al., 2000). Similarly, none of the large number VAMP-homologs found in plants show homology to VAMP8 (Sanderfoot et al., 2000). It is therefore clear that the SYP2/SYP5 complex will be distinct from the analogous mammalian complex, and research into the fourth member of the SYP2 + SYP5 + VTI11 complex continues.
In conclusion, we have identified at least five distinct SNARE complexes on the TGN or PVC of Arabidopsis cells that contain more than one member of the syntaxin-group of SNAREs (Figure 6). We have tentatively placed the SYP51/SYP61 complex on the TGN due to the likelihood of it containing VTI12 (which seems to prefer TGN-localized SYPs), but we cannot rule out the possibility of this complex forming on the PVC or other endosomal organelles. At this point, we are not able to clearly assign a function to any of these complexes. Gene disruptions of many Arabidopsis syntaxins have shown that each are essential genes; however, the lethality associated with loss of gene function occurs too early in development to allow analysis of the defects (Sanderfoot et al., 2001). Based on several lines of evidence, including intracellular localization and the interactions listed above, we believe we can give a preliminary model for at least one of these complexes. VTI11 is found on the TGN, and on TGN-derived vesicles where it is in proximity to the vacuolar cargo receptor AtELP (Zheng et al., 1999). Because VTI11 interacts with the PVC SYP21 + SYP51 t-SNARE complex (which requires a third, as yet unidentified SNARE), we propose that VTI11 is mediating the trafficking of the AtELP-bound vacuolar cargo to the PVC. Much more work is required to prove this hypothesis, as well as to identify functions for the other SNARE complexes we have characterized in this work.
Figure 6.

Schematic model of the Arabidopsis secretory system. A diagram of the late secretory pathway of Arabidopsis is shown. On two different domains of the TGN are found complexes containing SYP41 or 42, each in a complex containing VTI12 and SYP61. SYP51 is found on the SYP42-labeled domain of the TGN, as well as on the PVC. On the PVC, SYP51 is found in a complexes with VTI11 and either SYP21 or SYP22. SYP51 and SYP61 also form a complex, likely with VTI12 (although it is possible that this may be VTI11 instead, indicated by asterisk). The exact location of the SYP51/61 complex in unclear, but is drawn on the TGN because of the likelihood of containing VTI12 (see text).
ACKNOWLEDGMENTS
We thank Emily Avila-Teeguarden for donating RNA from various Arabidopsis tissues, the Arabidopsis Biological Resource Center for providing ESTs, and John Leszyk for performing the mass spectroscopy analysis. A.A.S. was partially supported by a National Institute of Health postdoctoral fellowship (GM-18861). N.V.R. was supported by funds from the Department of Energy (DE-FG02-91ER-20021) and the National Science Foundation (MCB-9507030).
Abbreviations used:
- IP
immunoprecipitate
- IPd
immunoprecipitated
- PVC
prevacuolar compartment
- SNARE
soluble N-ethyl maleimide sensitive factor adaptor protein receptors
- SYP
syntaxin of plants
- TGN
trans-Golgi network
REFERENCES
- Antonin W, Holroyd C, Fasshauer D, Pabst S, Fischer Von Mollard G, Jahn R. A SNARE complex mediating fusion of late endosomes defines conserved properties of SNARE structure and function. EMBO J. 2000;19:6453–6464. doi: 10.1093/emboj/19.23.6453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bar-Peled M, Conceição AS, Frigerio L, Raikhel NV. Expression and regulation of aERD2, a gene encoding the KDEL receptor homolog in plants, and other genes encoding proteins involved in ER-Golgi vesicular trafficking. Plant Cell. 1995;7:667–676. doi: 10.1105/tpc.7.6.667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassham DC, Gal S, Conceição AS, Raikhel NV. An Arabidopsis syntaxin homologue isolated by functional complementation of a yeast pep12 mutant. Proc Natl Acad Sci USA. 1995;92:7262–7266. doi: 10.1073/pnas.92.16.7262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bassham DC, Sanderfoot AA, Kovaleva V, Zheng H, Raikhel NV. AtVPS45 complex formation at the trans-Golgi network. Mol Biol Cell. 2000;11:2251–2265. doi: 10.1091/mbc.11.7.2251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becherer KA, Reide SE, Emr SD, Jones EW. Novel syntaxin homologue, Pep12p, required for the sorting of lumenal hydrolases to the lysosome-like vacuole of yeast. Mol Biol Cell. 1996;7:579–594. doi: 10.1091/mbc.7.4.579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bent AF, Kunkel BN, Dahlbeck D, Brown KL, Schmidt R, Giraudat J, Leung J, Staskawicz BJ. RPS2 of Arabidopsis thaliana: a leucine-rich repeat class of plant disease resistance genes. Science. 1994;265:1856–1860. doi: 10.1126/science.8091210. [DOI] [PubMed] [Google Scholar]
- Bock JB, Klumperman J, Davanger S, Scheller RH. Syntaxin 6 functions in trans-Golgi network vesicle trafficking. Mol Biol Cell. 1997;8:1261–1271. doi: 10.1091/mbc.8.7.1261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conceição AS, Marty-Mazars D, Bassham DC, Sanderfoot AA, Marty F, Raikhel NV. The syntaxin homologue AtPEP12p resides on a late post-Golgi compartment in plants. Plant Cell. 1997;9:571–582. [PMC free article] [PubMed] [Google Scholar]
- Fukuda R, McNew JA, Weber T, Parlati F, Engel T, Nickel W, Rothman JE, Söllner TH. Functional architecture of an intracellular membrane t-SNARE. Nature. 2000;407:198–202. doi: 10.1038/35025084. [DOI] [PubMed] [Google Scholar]
- Gurunathan S, Chapman-Shimshoni D, Trajkovic S, Gerst JE. Yeast exocytic v-SNAREs confer endocytosis. Mol Biol Cell. 2000;11:3629–3643. doi: 10.1091/mbc.11.10.3629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holthuis JCM, Nichols BJ, Dhruvakumar S, Pelham HRB. Two syntaxin homologues in the TGN/endosomal system of yeast. EMBO J. 1998;17:113–126. doi: 10.1093/emboj/17.1.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holthuis JCM, Nichols BJ, Pelham HRB. The syntaxin Tlg1p mediates trafficking of chitin synthase III to polarized growth sites in yeast. Mol Biol Cell. 1999;9:3383–3397. doi: 10.1091/mbc.9.12.3383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kneller DG, Cohen FE, Langridge R. Improvements in protein secondary structure prediction by an enhanced neural network. J Mol Biol. 1990;214:171–182. doi: 10.1016/0022-2836(90)90154-E. [DOI] [PubMed] [Google Scholar]
- Klumperman J, Kuliawat R, Griffith JM, Geuze HJ, Arvan P. Mannose-6-phosphate receptors are sorted from immature secretory granule via adaptor protein AP-1, clathrin, and syntaxin 6-positive vesicles. J Cell Biol. 1998;141:359–371. doi: 10.1083/jcb.141.2.359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lewis MJ, Nichols BJ, Prescianotto-Baschong C, Riezman H, Pelham HR. Specific retrieval of the exocytic SNARE Snc1p from early yeast endosomes. Mol Biol Cell. 2000;11:23–38. doi: 10.1091/mbc.11.1.23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lukowitz W, Mayer U, Jürgens G. Cytokinesis in the Arabidopsis embryo involves the syntaxin-related KNOLLE gene product. Cell. 1996;84:61–71. doi: 10.1016/s0092-8674(00)80993-9. [DOI] [PubMed] [Google Scholar]
- McNew JA, Parlati F, Fukuda R, Johnston J, Paz K, Paumet F, Söllner TH, Rothman JH. Compartmental specificity of cellular membrane fusion encoded in SNARE proteins. Nature. 2000;407:153–159. doi: 10.1038/35025000. [DOI] [PubMed] [Google Scholar]
- Misura KM, Scheller RH, Weis WI. Three-dimensional structure of the neuronal-Sec1-syntaxin 1a complex. Nature. 2000;404:355–62. doi: 10.1038/35006120. [DOI] [PubMed] [Google Scholar]
- Mullock BM, et al. Syntaxin 7 is localized to late endosome compartments, associates with Vamp 8, and Is required for late endosome-lysosome fusion. Mol Biol Cell. 2000;11:3137–3153. doi: 10.1091/mbc.11.9.3137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parlati F, McNew JA, Fukuda R, Miller R, Söllner TH, Rothman JH. Topological restriction of SNARE-dependent membrane fusion. Nature. 2000;407:194–198. doi: 10.1038/35025076. [DOI] [PubMed] [Google Scholar]
- Pelham HR. SNAREs and the secretory pathway—Lessons from yeast. Exp Cell Res. 1999;247:1–8. doi: 10.1006/excr.1998.4356. [DOI] [PubMed] [Google Scholar]
- Prekeris R, Yang B, Oorschot V, Klumperman J, Scheller RH. Differential roles of syntaxin 7 and syntaxin 8 in endosomal trafficking. Mol Biol Cell. 1999;10:3891–3908. doi: 10.1091/mbc.10.11.3891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderfoot AA, Ahmed SU, Marty-Mazars D, Rapoport I, Kirchhausen T, Marty F, Raikhel NV. A putative vacuolar cargo receptor partially colocalizes with AtPEP12p on a prevacuolar compartment in Arabidopsis roots. Proc Natl Acad Sci USA. 1998;95:9920–9925. doi: 10.1073/pnas.95.17.9920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderfoot AA, Assaad FF, Raikhel NV. The Arabidopsis Genome: an abundance of soluble N-ethylmaleimide sensitive factor receptors. Plant Physiol. 2000;121:929–938. doi: 10.1104/pp.124.4.1558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderfoot AA, Kovaleva V, Zheng H, Raikhel NV. The t-SNARE AtVAM3p resides on the prevacuolar compartment in Arabidopsis root cells. Plant Physiol. 1999;121:929–938. doi: 10.1104/pp.121.3.929. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderfoot AA, Pilgrim M, Adam L, Raikhel NV. Disruption of individual members of Arabidopsis syntaxin gene families indicates each have essential functions. Plant Cell. 2001;13:659–666. doi: 10.1105/tpc.13.3.659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanderfoot AA, Raikhel NV. The specificity of vesicle trafficking: coat proteins and SNAREs. Plant Cell. 1999;11:629–641. doi: 10.1105/tpc.11.4.629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato TK, Darsow T, Emr SD. Vam7p, a SNAP-25-like molecule, and Vam3p, a syntaxin homolog, function together in yeast vacuolar protein trafficking. Mol Cell Biol. 1998;18:5309–5319. doi: 10.1128/mcb.18.9.5308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato MH, Nakamura N, Ohsumi Y, Kouchi H, Kondo M, Hara-Nishimura I, Nishimura M, Wada Y. The AtVAM3 encodes a syntaxin-related molecule implicated in the vacuolar assembly in Arabidopsis thaliana. J Biol Chem. 1997;272:24530–24535. doi: 10.1074/jbc.272.39.24530. [DOI] [PubMed] [Google Scholar]
- Simonsen A, Bremnes B, Rønning E, Aasland R, Stenmark H. Syntaxin-16, a putative Golgi t-SNARE. Eur J Cell Biol. 1998;75:223–231. doi: 10.1016/S0171-9335(98)80116-7. [DOI] [PubMed] [Google Scholar]
- Tang BL, Low DY, Tan AE, Hong W. Syntaxin 10: a member of the syntaxin family localized to the trans-Golgi network. Biochem Biophys Res Commun. 1998;242:345–350. doi: 10.1006/bbrc.1997.7966. [DOI] [PubMed] [Google Scholar]
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature. 2000;408:796–815. doi: 10.1038/35048692. [DOI] [PubMed] [Google Scholar]
- Ungermann C, Wickner W. Vam7p, a vacuolar SNAP-25 homolog, is required for SNARE complex integrity and vacuole docking and fusion. EMBO J. 1998;17:3269–3276. doi: 10.1093/emboj/17.12.3269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wade N, Bryant NJ, Connolly LM, Simpson RJ, Luzio JP, Piper RC, James DE. Syntaxin 7 complexes with mouse Vps10p tail interactor 1b, syntaxin 6, vesicle associated membrane protein (VAMP)8 and VAMP7 in B16 melanoma cells. J Biol Chem. 2001;276:19820–19827. doi: 10.1074/jbc.M010838200. [DOI] [PubMed] [Google Scholar]
- Wedlich-Söldner R, Bölker M, Kahlmann R, Steinberg G. A putative endosomal t-SNARE links exo- and endocytosis in the phytopathogenic fungus Ustilago maydis. EMBO J. 2000;19:1974–1986. doi: 10.1093/emboj/19.9.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng H, Fischer von Mollard G, Kovaleva V, Stevens TH, Raikhel NV. The plant vesicle-associated SNARE AtVTI1a likely mediates vesicle transport from the trans-Golgi network to the prevacuolar compartment. Mol Biol Cell. 1999;10:2251–2264. doi: 10.1091/mbc.10.7.2251. [DOI] [PMC free article] [PubMed] [Google Scholar]