

Abstract
After ovulation, metaphase II oocytes undergo a time-dependent deterioration in vivo or in vitro, which is referred to as postovulatory oocyte aging, a process during which a series of deleterious molecular and cellular changes occur. In this study, we found that short-term injection of resveratrol (3,5,4'-trihydroxystilbene) effectively ameliorated oxidative stress-induced damage in postovulatory oocyte aging of middle-aged mice in vivo. Resveratrol induced changes that delayed the aging-induced oocyte deterioration including the elevated expression of the anti-aging molecule Sirtuin 1 (SIRT1); it reduced intracellular reactive oxygen species (ROS) level, and improved mitochondria function. In addition, these beneficial changes may also help to prevent apoptosis. Taken together, our data suggest that resveratrol can effectively protect against postovulatory oocyte aging in vivo primarily by preventing ROS production.
Keywords: resveratrol, postovulatory aging, oocyte, oxidative stress
Introduction
Successful pregnancy requires high-quality embryos with embryonic developmental potential that is largely dependent on oocyte quality, considering that the oocyte provides all the necessary factors for orchestrating key events during early embryo development [1]. Oocyte deterioration as a result of postovulatory aging negatively impacts embryonic development [2–4]. Postovulatory aging refers to a degenerative process in MII oocytes which is strongly associated with fertilization failures within a period of time after ovulation. Postovulatory aging occurs both in vivo and in vitro. In humans and certain primates, owing to no visible signs of ovulation, fertilization may occur between an in vivo aged oocyte and freshly ejaculated spermatozoa. Oocytes employed in assisted reproductive technologies (ART) are often inevitably subjected to postovulatory aging. In light of these considerations, it is highly important to understand the mechanisms underlying the oocyte degeneration process, which may lead to solutions to preventing or delaying postovulatory aging and allow successful ART and embryo engineering procedures.
Postovulatory aging is highly associated with reduced fertilization rates [5], poor embryo quality [2–4] and fetus development [2,6,7]; however, the molecular mechanisms that control this process still remain poorly understood. Accumulating evidence indicates that antioxidant enzymes including Mn-superoxide dismutase (Mn-SOD), catalase, and glutathione peroxidase reduce ROS, whereas the production of ROS rises gradually with increasing maternal age [8]. Oxidative stress is strongly linked to the declined oocyte quality [9,10]. In addition, because the mitochondrial DNA (mtDNA) is not protected by histones, aging oocytes are very vulnerable to oxidative stress damage and mtDNA mutations. Dysfunctional mitochondria affect the oocyte’s metabolic capacity, and thus impair adenosine triphosphate (ATP) production. The ATP content in oocytes is correlated with embryonic development and implantation [11]. Elevation of intracellular ROS also inactivates nicotinamide adenine dinucleotide (NAD+)-dependent histone deacetylase SIRT1 [12], and decreased expression of SIRT1 accelerates postovulatory aging of oocytes in vitro [13]. In addition, high level of oxidative stress is considered an inducer of apoptosis of aging oocytes [14,15].
Resveratrol is a type of natural phenolic compound and a phytoalexin produced by several plants in response to injures or attacks by pathogens. It exhibits therapeutic effects against various diseases including cancer [16–18], diabetes [19–21], obesity [22,23], cardiovascular diseases [24,25], neurodegenerative disorders [26,27] and aging [28–31], and the effects are thought to be correlated to its anti-oxidative activity of resveratrol. In addition, it has been reported that resveratrol improved in vitro maturation of oocytes and enhanced the oocytes’ resistance to chemical reagents [32,33], heat stress [34] and cryopreservation-induced damage [35] in various species. Furthermore, resveratrol improves fertilization outcome of pig oocytes [36] and subsequent embryonic developmental potential [37,38]. Extensive studies about the effects of resveratrol on oocyte maturation are carried out in vitro, while in vivo research is rarely reported. In this study, we investigated the effects of resveratrol on delaying postovulatory aging of mouse oocytes in vivo.
RESULTS
Resveratrol maintains the survival of aging MII oocytes in vivo
To investigate the potential role of resveratrol in postovulatory oocyte aging in vivo, female ICR mice at 7-8 months of age (from the time at which fertility begins to decline) were injected with resveratrol (50 mg/kg BW/day) for 15 consecutive days. Forty-eight hours after PMSG injection, hCG was administrated, and 24 h later postovulatory in vivo aging oocytes and ovaries were collected. Firstly, the weights of control and resveratrol-treated mice were monitored on D1, D6, D11 and D15, and the body weight of resveratrol-treated mice was slightly lighter than controls from D11 to D15, but without significant differences (41.78 ± 3.34 vs. 42.19 ± 3.67 g; 40.5 ± 2.93 vs. 39.95 ± 3.26 g; 40.37 ± 3.01 vs. 41.91 ± 3.76 g; 40.97 ± 2.78 vs. 42.68 ± 4.21 g, n= 30, P > 0.05) (Figure 1A). No difference was observed in ovarian weight ratio between the resveratrol-treated group and the control group (1.06 ± 0.24% vs. 1.03 ± 0.29%, n= 30, P > 0.05) (Figure 1B). The histological assessment of ovarian sections showed that the corpus luteum formed normally in both resveratrol-treated and control groups (Figure 1C), which revealed the normal occurrence of ovulation. A portion of the aging oocytes showed apoptosis or death in both groups (Figure 2A). We counted the total number of super-ovulated oocytes, and no obvious variation was found in the resveratrol-treated group and the control group (23.39 ± 5.82 vs. 25.44 ± 8.48, n= 30, P > 0.05) (Figure 2B). We calculated the rate of living oocytes, and found that it was significantly higher in the treated group compared to the control group (85.11 ± 7.15% vs. 69.79 ± 13.75%, n= 30, P < 0.05) (Figure 2C).
Figure 1.
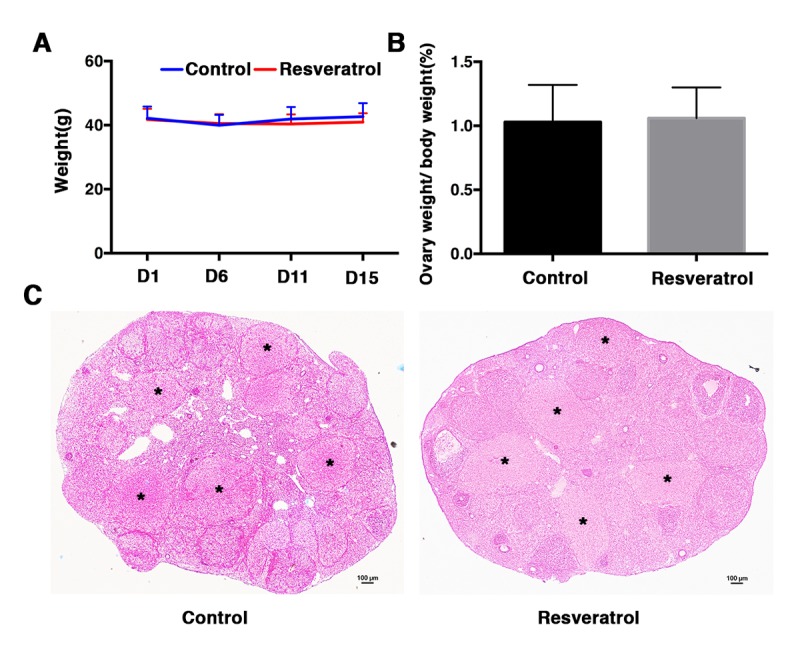
(A) Ovary weight of control and resveratrol-treated group at day 1, 6, 11 and 15 after treatment. For each time point, at least 30 mice of each group were used for analysis. Data are expressed as mean ± SEM of at least 6 independent experiments. (B) Ovary weight to body weight ratio of control and resveratrol-treated group at day 15 after treatment. At least 30 mice of each group were used for analysis. Data are expressed as mean ± SEM of at least 6 independent experiments. (C) Representative ovarian histology of control and resveratrol-treated group: 48 h after PMSG injection, hCG was administrated and ovaries were collected 24 h later for histological analysis. Black asterisks indicate corpus luteum. Scale bar: 100 μm.
Figure 2.
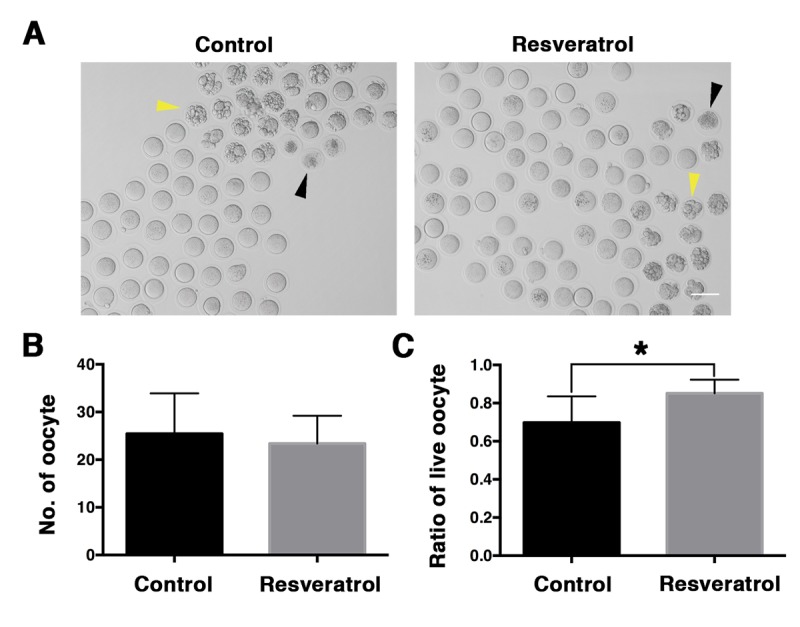
Morphological evaluation of aging MII oocytes derived from control and resveratrol-treated mice in vivo. (A) Microscopy images of aging MII oocytes from control and resveratrol-treated mice in vivo. Yellow and black arrowheads indicate apoptotic and death oocytes, respectively. (B) The number of aging MII oocytes from control and resveratrol-treated groups. Data are expressed as mean ± SEM of at least 5 independent experiments. (C) The ratio of live oocytes in control and resveratrol-treated groups. Data are expressed as mean ± SEM of at least 5 independent experiments. *Significantly different (P < 0.05).
Resveratrol elevates the expression of SIRT1
Previous studies reported that SIRT1 could protect mouse oocytes from postovulatory aging and resveratrol was an activator of SIRT1. Accordingly, we analyzed the mRNA and protein level of SIRT1 in the oocytes of control and resveratrol-treated mice. The results of real-time PCR revealed that at mRNA level SIRT1 was significantly elevated in aging oocytes from resveratrol-treated mice, when compared to the control group (Figure 3A). Besides, immunoblotting analysis showed that the level of SIRT1 protein was also higher in aging oocytes of resveratrol-treated mice compared to the control group (Figure 3B and 3C).
Figure 3.
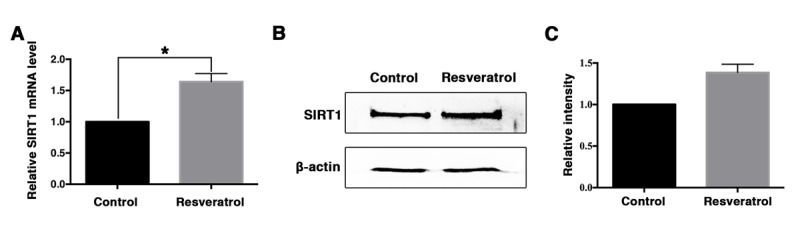
The expression of SIRT1 in aging MII oocytes derived from control and resveratrol-treated mice in vivo. (A) The expression of SIRT1 mRNA detected by quantitative RT-PCR. Error bar denotes SEM of three experiments. (B) Western blot detection of SIRT protein levels in aging MII oocytes from control and resveratrol-treated mice. (C) Quantitative analysis of gray intensity in control and resveratrol-treated groups.
Resveratrol reduces ROS levels in aging MII oocytes in vivo
Towards determining whether resveratrol might protect against oxidative stress, aging MII oocytes derived from control and resveratrol-treated mice were collected to measure the level of intracellular ROS. The results showed that ROS in aging MII oocytes from control groups displayed clustered distributions; by comparison, ROS distribution in aging MII oocytes from resveratrol-treated groups was relatively uniform (Figure 4A). Moreover, the ROS fluorescence intensity of oocytes from resveratrol-treated mice was significantly lower compared to control aging oocytes (163.23 ± 32.56 vs. 359.88 ± 91.71, n= 70, P < 0.05) (Figure 4B), indicating a reduced production of ROS.
Figure 4.
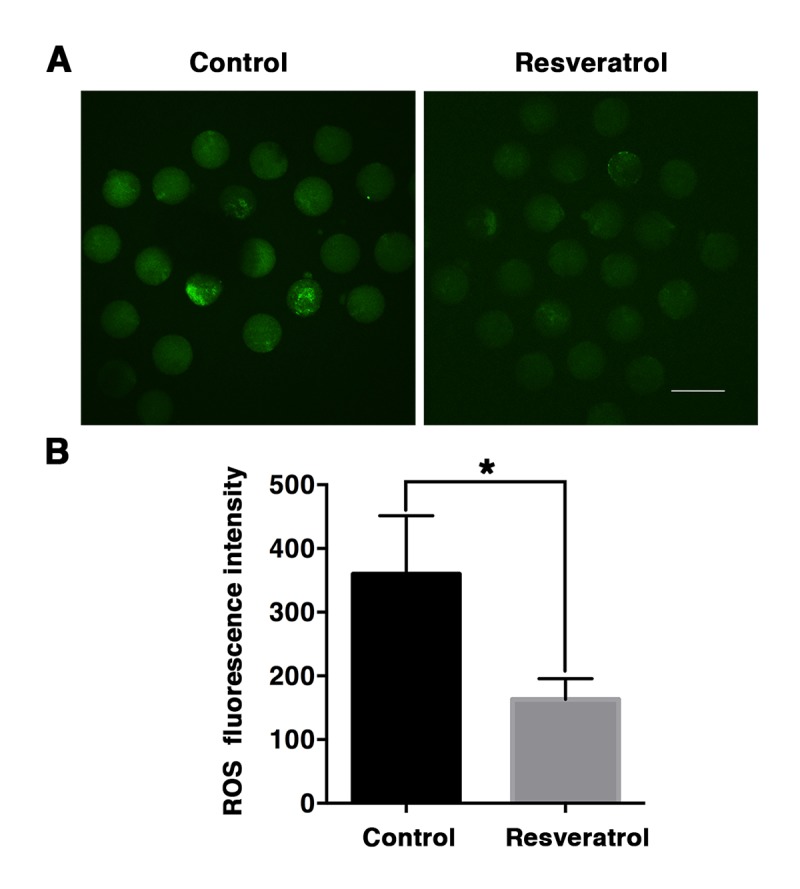
Detection of ROS production in MII oocytes during aging in vivo. (A) Representative confocal images of DCF fluorescence in control and resveratrol-treated oocytes. ROS production was measured by fluorescent probe DCFA-DA (green). Scale bar: 100 μm. (B) The relative levels of intracellular ROS determined by quantitative fluorescence intensity. Data are expressed as mean ± SEM of at least 5 independent experiments. *Significantly different (P < 0.05).
The mtDNA copy number is lower in resveratrol-treated oocytes, but mitochondrial distribution is not affected
Because damage to mitochondria is a known cause of increased production of ROS and mitochondrial dysfunction is presumably linked to postovulatory oocyte aging, we next examined mitochondrial distribution and copy number in aging MII oocytes from control and resveratrol-treated mice. The results indicated that mitochondria in MII oocytes from both groups displayed two distribution patterns: homogeneous distribution pattern and clustered distribution pattern (Figure 5A). The proportion of the two patterns was similar in the resveratrol-treated group and the control group (50, 50%, n= 40 vs. 49, 51%, n= 50, P > 0.05) (Figure 5B). In addition, real-time PCR was performed to count mtDNA copy numbers in the two groups. Unexpectedly, we found that the mtDNA copy number in the resveratrol-treated group was significantly lower compared with the control non-treated aging group (322,504 ± 27,979, n=27, vs. 396,841 ± 15,478, n=40, P < 0.05) (Figure 5C).
Figure 5.
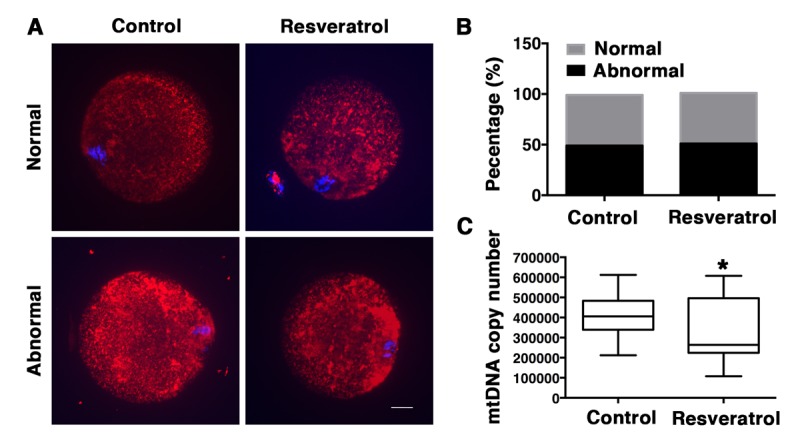
Analysis of mitochondrial distribution and mtDNA copy number in MII oocytes during aging in vivo. (A) Confocal microscopy images of normal and abnormal mitochondrial distribution patterns in oocytes recovered from control and resveratrol-treated mice. Mitochondrial distribution patterns were detected using Mito Tracker Red. DNA was counterstained with Hoechst (blue). Scale bar: 10 μm. (B) Percentages of the normal and abnormal distribution patterns in control or resveratrol-treated oocytes, respectively. (C) The average copy number of mitochondrial DNA in control and resveratrol-treated oocytes. Data are expressed as mean ± SEM of at least 3 independent experiments. *Significantly different (P < 0.05).
Resveratrol does not improve spindle assembly and chromosome alignment in aging oocytes
The spindle abnormalities have been widely reported in aging MII oocytes. Owing to oxidative stress’s negative effects on meiotic spindle integrity, we determined whether reduced oxidative stress in aging MII oocyte induced by resveratrol treatment could facilitate maintenance of spindle morphology. Immunofluorescent results revealed that more than half of the oocytes in control and resveratrol-treated group exhibited normal spindle with a typical barrel-shape positioned beneath the cortex (Figure 6A). Meanwhile, abnormal spindles were also observed in both groups. Morphological abnormalities of spindles in this experiment mainly included elongated spindles, spindles without poles, mono-polar spindles, multi-poles and unorganized spindles with astral microtubules as well as many cytoplasmic asters (Figure 6A). The proportions of defective spindles were similar in the resveratrol-treated group and the control group (60.53, 39.47% n= 120, vs. 61.54, 38.46%, n= 170, P > 0.05) (Figure 6B).
Figure 6.
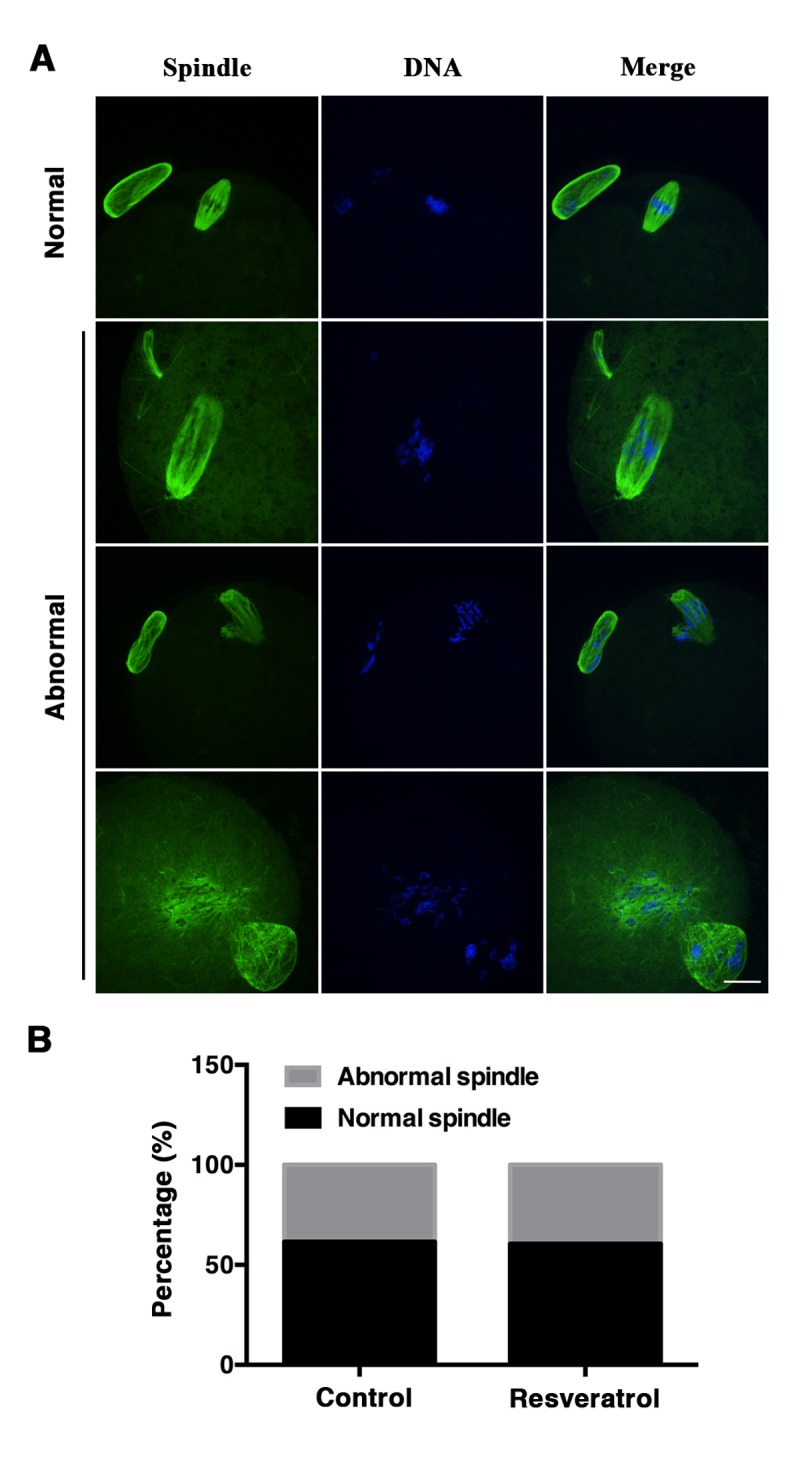
(A) Immunofluorescent images of normal and abnormal spindle morphology of control and resveratrol-treated oocytes. Oocytes were stained with α-tubulin antibody (green) and Hoechst (blue) to show spindle morphology and chromosome alignment, respectively. Scale bar: 20 μm. (B) Proportions of the normal and abnormal spindle morphology of aging MII oocytes from control and resveratrol-treated groups. Data are expressed as mean ± SEM of at least 5 independent experiments.
DISCUSSION
When a woman reaches her mid-thirties, fertility becomes decreased, and the oocytes undergo a decline in developmental potential with increasing maternal age. Although the molecular mechanisms are complex, a convincing link between oxidative stress and age-related decline in oocyte quality is well established [39]. In addition, oxidative stress also affects the quality of postovulatory oocytes both in vivo and in vitro processes [10,40]. Accordingly, it has become particularly pressing to find an effective treatment to prevent or delay the damage caused by oxidative stress. As a promising anti-oxidative small molecule, the effects of resveratrol on preventing chemical-induced oxidative damage or apoptosis have been extensively studied [32,33,41,42]. Furthermore, the possible functions of resveratrol in improving oocyte maturation, fertilization and preimplantation embryonic development have also been reported [36–38]. In addition, previous studies demonstrated that long-term treatment (12 months) of resveratrol was able to protect against age-associated infertility, as evidenced by an elevated follicle pool, decreased spindle aberrations and chromosome misalignments [28]. However, most of these studies were carried out in vitro and did not demonstrate the effects on delaying postovulatory oocyte aging. The present study fills this gap and provides important evidence for in vivo roles of resveratrol in delaying postovulatory oocyte aging.
In this study, we investigated the potential roles of resveratrol in delaying postovulatory oocyte aging in vivo. The 7-8 month-old female mice were selected as experimental subjects because the fertility begins to decrease from that age on. Our data showed that short-term injection of resveratrol delayed aging and apoptosis of postovulatory oocytes, which were possibly achieved by attenuating oxidative stress and preserving mitochondrial function. Our results showed that resveratrol treatment significantly decreased ROS levels and elevated the expression of SIRT1 in postovulatory oocytes, which was consistent with previous in vitro studies [32,43]. Oxidative stress is related to age-related mitochondrial dysfunction. Further studies indicated that resveratrol-treatment did not impact mitochondrial distribution but significantly decreased mitochondrial copy number in aging oocytes. One possible reason was that resveratrol treatment protected mitochondrial DNA from impairment by oxidative stress in aging oocytes and improved mitochondrial function, which was consistent with previous studies in patient-derived fibroblasts [44] and porcine oocytes [36]; accordingly, it was not necessary to increase mitochondria copy number to compensate mitochondrial dysfunction in resveratrol treatment group compared to control group.
Previous studies showed that chronic administration of resveratrol (6 months) prevented age-associated spindle abnormalities [28]. However, our experiment results found that short-term resveratrol treatment did not prevent spindle aberrations in aging oocytes. It is probably that low dosage and short-term treatment of resveratrol may not be sufficient to rescu the spindle aberrations in aging oocytes. Besides, similar results were observed in study of another antioxidant N-Acetyl-L-cysteine (NAC), in which NAC-treatment for 6 months showed no obvious improvement on spindle aberrations [15]. Another in vitro study found that the short-term treatment of Nicotinamide (NAM), a SIRT1, 2, 3 inhibitor, significantly elevated ROS levels but did not aggravate spindle abnormalities in aging oocytes [13].
In summary, in vivo administration of resveratrol decreases fragmentation and death of oocyte aging in the oviduct, probably by improving mitochondrial function, increasing SIRT1 expression and preventing ROS production. The results of this research may provide reproductive health care providers with new clues to improve their clinical practice. Administration of resveratrol may be an effective treatment to improve the oocyte developmental potential and increase the probability of getting pregnancy either naturally or through assisted reproductive technique. However, the effectiveness and safety of resveratrol require verification by further studies involving human subject.
MATERIALS AND METHODS
Mice
Female ICR mice were housed in a temperature-controlled room with 12D:12L (dark vs. light). Water and food were available ad libitum. All animal operations conformed to the guidelines by the Animal Research Committee principles of the Institute of Zoology, Chinese Academy of Sciences. A stock solution of resveratrol was diluted in absolute ethanol to 50g/L and stored at -20°C. Prior to administration, the stock solution was diluted with normal saline (0.9% NaCl) to a final concentration of 2.5g/L. Female ICR mice (aged 7-8 months) were randomly divided into experiment groups and control groups. For the experiment groups, resveratrol (Puhuashi Technology Development (Beijing) Co., Ltd, China; HPLC ≥ 99%) was administrated by intraperitoneal injection (50 mg/kg BW/day) for 15 consecutive days (Table 1). Control groups received injection of normal saline with 5% of ethanol (50 mg/kg BW/day) for the same period (Table 1). Body weights were assessed every 3 days during the experiment and injection doses were adjusted accordingly.
Table 1. A time table for the treatment of animals and sample collection.
| Day 1-12 | Day 13 | Day 14 | Day 15 | Day 16 | |
| 9:00 | PMSG | hCG | Egg Retrieval | ||
| 19:00 | Res/5% Ethanol | Res/5% Ethanol | Res/5% Ethanol | Res/5% Ethanol |
Tissue and oocyte collection
To collect MII eggs, all mice were injected with PMSG (10 IU) in the thirteenth day of injection, 48 h later with hCG (10 IU) (Table 1). Prior to oocyte collection, mice were weighted and sacrificed by cervical dislocation. Eggs were retrieved from oviductal ampullae 24 h after hCG (Table 1), and cumulus cells were removed by gentle pipetting in M2 (Sigma, USA) medium containing 0.3% hyaluronidase. Then denuded oocytes were examined, or collected and stored in -80°C. Ovaries were weighted and placed in 4% paraformaldehyde before subsequent processing.
Hematoxylin and eosin staining
Ovaries were fixed in 4% paraformaldehyde overnight at 4°C, dehydrated with a graded ethanol series and embedded in paraffin. Paraffin-embedded ovaries were cut into sections of 8-μm thickness and mounted on glass slides. After adequately drying for 45 °C overnight, sections were deparaffinized in xylene, hydrated by a graded alcohol series and stained with hematoxylin and eosin for histological analyses.
Western blotting
For western blotting, 120 oocytes were lysed in 2×SDS sample buffer, boiled for 5 min at 100°C and then subjected to 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). The separated proteins were transferred to a polyvinylidene difluoride (PVDF) membrane. Membrane was blocked in Tris-buffered saline (TBS) containing 5% BSA and 0.1% Tween-20 for 2 h and then incubated with rabbit anti-SIRT1 antibody (1:1000) (9475; Cell Signaling Technology, Inc.) and mouse anti-β-actin antibody (1:1000) (BE0021; Easybio Technology, Beijing). After multiple washes in TBS containing 0.1% Tween-20 and incubation with horseradish peroxidase (HRP)-conjugated anti-rabbit IgG (1:1000) (ZB-2301; Zhongshan Golden Bridge Biotechnology, Beijing) and horseradish peroxidase conjugated anti-mouse IgG (1:1000) (ZB-2305; Zhongshan Golden Bridge Biotechnology, Beijing), respectively, finally, the membranes were washed 3 times in TBS containing 0.1% Tween-20 and visualized using Bio-Rad ChemiDoc XRS+.
Quantitative RT-PCR analysis
Total RNA was extracted from 50 oocytes using RNeasy Micro Kit (Qiagen). The first strand cDNA was generated with cDNA synthesis kit (Invitrogen). Sirt1 genes were amplified. Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as control gene to correct cDNA levels of samples. Real-time PCR was conducted by using UltraSYBR Mixiure (CoWin Biosciences (Beijing) Co., Ltd., China) in Roche LightCycler480 II detection system. The relative gene expression was calculated by the 2-ΔΔCt method. The primers used were as follows. Sirt1 forward: 5′-TATCTATGCTCGCCTTGCGG-3′; Sirt1 reverse 5′-CGGGATATATTTCCTTTGCAAACTT-3. Gapdh forward: 5′-CCCCAATGTGTCCGTCGTG-3′; Gapdh reverse 5′-TGCCTGCTTCACCACCTTCT-3.
Immunofluorescence
Immunofluorescence was performed by using previously published methods [45]. Briefly, oocytes were fixed for 30 min at room temperature in 2% formaldehyde supplemented with 100mM HEPES, 50mM EGTA, 10mM MgSO4, 0.2% Triton X-100 (pH=7, titrated with KOH). Then they were treated with PBS, 0.1% Triton X-100 overnight at 4oC and incubated with anti-α-tubulin-FITC antibody (F2168, Sigma, USA) (1:1000 in PBS with 0.1% Triton X-100 and 3% BSA) overnight at 4oC. Chromosomes were stained with DAPI for 15 min. The oocytes were mounted on glass slides and examined with a laser scanning confocal microscope (Zeiss 780 META).
For staining active mitochondria, eggs were incubated for 30 min at 37°C in M2 medium containing 200nM MitoTracker ® Red CMXRos (M7512, Invitrogen, USA). After washing 3 times, eggs were stained with Hoechst 33342 (10mg/ml) for 15 min. Finally, eggs were mounted on glass slides and examined with a Perkin Elmer UltraVIEW VOX confocal Imaging System.
Detection of intracellular ROS levels
ROS levels detection was performed as previously reported [13]. Briefly, oocytes were incubated for 30 min at 37°C in M2 medium supplemented with 2',7'-dichlorodihydrofluorescein diacetate (DCFA-DA) (2 μM) (Beyotime Institute of Biotechnology (Shanghai) Co., Ltd., China). After washing three times in M2 medium, oocytes were stained with Hoechst 33342 (10mg/ml) for 15 min. Finally, oocytes were mounted on glass slides and examined in identical settings with Perkin Elmer UltraVIEW VOX confocal Imaging System.
Detection of mtDNA copy number
MtDNA copy number in oocytes was assessed according to the method previously described [46]. mtDNA-specific primers: B6 forward, 5′-AACCTGGCACTGAGTCACCA-3′, and B6 reverse, 5′-GGGTCTGAGTGTATATATCATGAAGAGAAT-3′ were used to prepare a standard amplification curve for absolute quantification of mtDNA. PCR products were ligated to the T-vector and amplified in E. coli. Then, plasmids were extracted and purified, followed by 10-fold serial dilutions for seven times. Standard curve was established afterwards. One oocyte was placed into a PCR tube with 5 µl lysis buffer and incubated at 55°C for 2 h. After heat inactivation of proteinase K at 95°C for 10 min, lysates were used for real-time PCR using Roche LightCycler480 II detection system and the above-mentioned B6 primers. Reaction conditions of PCR were as follows: initiation at 95°C for 10 min, 40 cycles of 95°C treatment for 10 s and 60°C reaction for 1 min. The melting temperature was set at 76.5°C. All standard curves of these samples were processed by linear regression, which showed a correlation coefficient of 0.98. All measurements were repeated three times.
Statistical analysis
All experiments were performed at least three times. Data were presented as mean ± SEM. Data were evaluated by Student’s t-test. P<0.05 was considered statistically significant.
ACKNOWLEDGEMENTS
We appreciated and acknowledge Lijuan Wang, Shiwen Li and Xili Zhu for their technical assistance. We thank all members of the Sun lab for their help and discussions.
Footnotes
CONFLICTS OF INTEREST: The authors have no conflict of interests to declare.
FUNDING: This work was supported by the Research Team of Female Reproductive Health and Fertility Preservation (SZSM201612065) and Project for Medical Discipline Advancement of Health and Family Planning Commission of Shenzhen Municipality (SZXJ2017003). .
REFERENCES
- 1.Schultz GA, Heyner S. Gene expression in pre-implantation mammalian embryos. Mutat Res. 1992; 296:17–31. 10.1016/0165-1110(92)90029-9 [DOI] [PubMed] [Google Scholar]
- 2.Lanman JT. Delays during reproduction and their effects on the embryo and fetus. 2. Aging of eggs. N Engl J Med. 1968; 278:1047–54. 10.1056/NEJM196805092781906 [DOI] [PubMed] [Google Scholar]
- 3.Wilcox AJ, Weinberg CR, Baird DD. Post-ovulatory ageing of the human oocyte and embryo failure. Hum Reprod. 1998; 13:394–97. 10.1093/humrep/13.2.394 [DOI] [PubMed] [Google Scholar]
- 4.Miao YL, Kikuchi K, Sun QY, Schatten H. Oocyte aging: cellular and molecular changes, developmental potential and reversal possibility. Hum Reprod Update. 2009; 15:573–85. 10.1093/humupd/dmp014 [DOI] [PubMed] [Google Scholar]
- 5.Ben-Rafael Z, Kopf GS, Blasco L, Tureck RW, Mastroianni L Jr. Fertilization and cleavage after reinsemination of human oocytes in vitro. Fertil Steril. 1986; 45:58–62. 10.1016/S0015-0282(16)49097-7 [DOI] [PubMed] [Google Scholar]
- 6.Tarín JJ, Pérez-Albalá S, Pérez-Hoyos S, Cano A. Postovulatory aging of oocytes decreases reproductive fitness and longevity of offspring. Biol Reprod. 2002; 66:495–99. 10.1095/biolreprod66.2.495 [DOI] [PubMed] [Google Scholar]
- 7.Tarín JJ, Pérez-Albalá S, Aguilar A, Miñarro J, Hermenegildo C, Cano A. Long-term effects of postovulatory aging of mouse oocytes on offspring: a two-generational study. Biol Reprod. 1999; 61:1347–55. 10.1095/biolreprod61.5.1347 [DOI] [PubMed] [Google Scholar]
- 8.Lim J, Luderer U. Oxidative damage increases and antioxidant gene expression decreases with aging in the mouse ovary. Biol Reprod. 2011; 84:775–82. 10.1095/biolreprod.110.088583 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Tamura H, Takasaki A, Miwa I, Taniguchi K, Maekawa R, Asada H, Taketani T, Matsuoka A, Yamagata Y, Shimamura K, Morioka H, Ishikawa H, Reiter RJ, Sugino N. Oxidative stress impairs oocyte quality and melatonin protects oocytes from free radical damage and improves fertilization rate. J Pineal Res. 2008; 44:280–87. 10.1111/j.1600-079X.2007.00524.x [DOI] [PubMed] [Google Scholar]
- 10.Lord T, Aitken RJ. Oxidative stress and ageing of the post-ovulatory oocyte. Reproduction. 2013; 146:R217–27. 10.1530/REP-13-0111 [DOI] [PubMed] [Google Scholar]
- 11.Van Blerkom J, Davis PW, Lee J. ATP content of human oocytes and developmental potential and outcome after in-vitro fertilization and embryo transfer. Hum Reprod. 1995; 10:415–24. 10.1093/oxfordjournals.humrep.a135954 [DOI] [PubMed] [Google Scholar]
- 12.Yang Y, Fu W, Chen J, Olashaw N, Zhang X, Nicosia SV, Bhalla K, Bai W. SIRT1 sumoylation regulates its deacetylase activity and cellular response to genotoxic stress. Nat Cell Biol. 2007; 9:1253–62. 10.1038/ncb1645 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zhang T, Zhou Y, Li L, Wang HH, Ma XS, Qian WP, Shen W, Schatten H, Sun QY. SIRT1, 2, 3 protect mouse oocytes from postovulatory aging. Aging (Albany NY). 2016; 8:685–96. 10.18632/aging.100911 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liu L, Trimarchi JR, Navarro P, Blasco MA, Keefe DL. Oxidative stress contributes to arsenic-induced telomere attrition, chromosome instability, and apoptosis. J Biol Chem. 2003; 278:31998–2004. 10.1074/jbc.M303553200 [DOI] [PubMed] [Google Scholar]
- 15.Liu J, Liu M, Ye X, Liu K, Huang J, Wang L, Ji G, Liu N, Tang X, Baltz JM, Keefe DL, Liu L. Delay in oocyte aging in mice by the antioxidant N-acetyl-L-cysteine (NAC). Hum Reprod. 2012; 27:1411–20. 10.1093/humrep/des019 [DOI] [PubMed] [Google Scholar]
- 16.Vendrely V, Peuchant E, Buscail E, Moranvillier I, Rousseau B, Bedel A, Brillac A, de Verneuil H, Moreau-Gaudry F, Dabernat S. Resveratrol and capsaicin used together as food complements reduce tumor growth and rescue full efficiency of low dose gemcitabine in a pancreatic cancer model. Cancer Lett. 2017; 390:91–102. 10.1016/j.canlet.2017.01.002 [DOI] [PubMed] [Google Scholar]
- 17.Bai Y, Yang H, Zhang G, Hu L, Lei Y, Qin Y, Yang Y, Wang Q, Li R, Mao Q. Inhibitory effects of resveratrol on the adhesion, migration and invasion of human bladder cancer cells. Mol Med Rep. 2017; 15:885–89. 10.3892/mmr.2016.6051 [DOI] [PubMed] [Google Scholar]
- 18.Sun L, Chen B, Jiang R, Li J, Wang B. Resveratrol inhibits lung cancer growth by suppressing M2-like polarization of tumor associated macrophages. Cell Immunol. 2017; 311:86–93. 10.1016/j.cellimm.2016.11.002 [DOI] [PubMed] [Google Scholar]
- 19.Tian Z, Wang J, Xu M, Wang Y, Zhang M, Zhou Y. Resveratrol Improves Cognitive Impairment by regulating apoptosis and synaptic plasticity in streptozotocin-induced diabetic rats. Cell Physiol Biochem. 2016; 40:1670–77. 10.1159/000453216 [DOI] [PubMed] [Google Scholar]
- 20.Timmers S, de Ligt M, Phielix E, van de Weijer T, Hansen J, Moonen-Kornips E, Schaart G, Kunz I, Hesselink MK, Schrauwen-Hinderling VB, Schrauwen P. Resveratrol as add-on therapy in subjects with well-controlled type 2 diabetes: a randomized controlled trial. Diabetes Care. 2016; 39:2211–17. 10.2337/dc16-0499 [DOI] [PubMed] [Google Scholar]
- 21.Wu H, Sheng ZQ, Xie J, Li R, Chen L, Li GN, Wang L, Xu B. Reduced HMGB 1-mediated pathway and oxidative stress in resveratrol-treated diabetic mice: a possible mechanism of cardioprotection of resveratrol in Diabetes Mellitus. Oxid Med Cell Longev. 2016; 2016:9836860. 10.1155/2016/9836860 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Wang S, Liang X, Yang Q, Fu X, Zhu M, Rodgers BD, Jiang Q, Dodson MV, Du M. Resveratrol enhances brown adipocyte formation and function by activating AMP-activated protein kinase (AMPK) α1 in mice fed high-fat diet. Mol Nutr Food Res. 2017; 61:16007467. 10.1002/mnfr.201600746 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zou T, Chen D, Yang Q, Wang B, Zhu MJ, Nathanielsz PW, Du M. Resveratrol supplementation of high-fat diet-fed pregnant mice promotes brown and beige adipocyte development and prevents obesity in male offspring. J Physiol. 2017; 595:1547–62. 10.1113/JP273478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Shah A, Quon A, Morton JS, Davidge ST. Postnatal resveratrol supplementation improves cardiovascular function in male and female intrauterine growth restricted offspring. Physiol Rep. 2017; 5:13109. 10.14814/phy2.13109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bai T, Hu X, Zheng Y, Wang S, Kong J, Cai L. Resveratrol protects against lipopolysaccharide-induced cardiac dysfunction by enhancing SERCA2a activity through promoting the phospholamban oligomerization. Am J Physiol Heart Circ Physiol. 2016; 311:H1051–62. 10.1152/ajpheart.00296.2016 [DOI] [PubMed] [Google Scholar]
- 26.Gocmez SS, Gacar N, Utkan T, Gacar G, Scarpace PJ, Tumer N. Protective effects of resveratrol on aging-induced cognitive impairment in rats. Neurobiol Learn Mem. 2016; 131:131–36. 10.1016/j.nlm.2016.03.022 [DOI] [PubMed] [Google Scholar]
- 27.Bernier M, Wahl D, Ali A, Allard J, Faulkner S, Wnorowski A, Sanghvi M, Moaddel R, Alfaras I, Mattison JA, Tarantini S, Tucsek Z, Ungvari Z, et al. Resveratrol supplementation confers neuroprotection in cortical brain tissue of nonhuman primates fed a high-fat/sucrose diet. Aging (Albany NY). 2016; 8:899–916. 10.18632/aging.100942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Liu M, Yin Y, Ye X, Zeng M, Zhao Q, Keefe DL, Liu L. Resveratrol protects against age-associated infertility in mice. Hum Reprod. 2013; 28:707–17. 10.1093/humrep/des437 [DOI] [PubMed] [Google Scholar]
- 29.Pearson KJ, Baur JA, Lewis KN, Peshkin L, Price NL, Labinskyy N, Swindell WR, Kamara D, Minor RK, Perez E, Jamieson HA, Zhang Y, Dunn SR, et al. Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab. 2008; 8:157–68. 10.1016/j.cmet.2008.06.011 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Park SJ, Ahmad F, Philp A, Baar K, Williams T, Luo H, Ke H, Rehmann H, Taussig R, Brown AL, Kim MK, Beaven MA, Burgin AB, et al. Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell. 2012; 148:421–33. 10.1016/j.cell.2012.01.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Monserrat Hernández-Hernández E, Serrano-García C, Antonio Vázquez-Roque R, Díaz A, Monroy E, Rodríguez-Moreno A, Florán B, Flores G. Chronic administration of resveratrol prevents morphological changes in prefrontal cortex and hippocampus of aged rats. Synapse. 2016; 70:206–17. 10.1002/syn.21888 [DOI] [PubMed] [Google Scholar]
- 32.Liu Y, He XQ, Huang X, Ding L, Xu L, Shen YT, Zhang F, Zhu MB, Xu BH, Qi ZQ, Wang HL. Resveratrol protects mouse oocytes from methylglyoxal-induced oxidative damage. PLoS One. 2013; 8:e77960. 10.1371/journal.pone.0077960 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu Y, Wang YL, He SW, Chen MH, Zhang Z, Fu XP, Fu BB, Liao BQ, Lin YH, Qi ZQ, Wang HL. Protective effects of resveratrol against mancozeb induced apoptosis damage in mouse oocytes. Oncotarget. 2017; 8:6233–45. 10.18632/oncotarget.14056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Li Y, Wang J, Zhang Z, Yi J, He C, Wang F, Tian X, Yang M, Song Y, He P, Liu G. Resveratrol compares with melatonin in improving in vitro porcine oocyte maturation under heat stress. J Anim Sci Biotechnol. 2016; 7:33. 10.1186/s40104-016-0093-9 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Giaretta E, Spinaci M, Bucci D, Tamanini C, Galeati G. Effects of resveratrol on vitrified porcine oocytes. Oxid Med Cell Longev. 2013; 2013:920257. 10.1155/2013/920257 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Takeo S, Sato D, Kimura K, Monji Y, Kuwayama T, Kawahara-Miki R, Iwata H. Resveratrol improves the mitochondrial function and fertilization outcome of bovine oocytes. J Reprod Dev. 2014; 60:92–99. 10.1262/jrd.2013-102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee S, Jin JX, Taweechaipaisankul A, Kim GA, Ahn C, Lee BC. Sonic hedgehog signaling mediates resveratrol to improve maturation of pig oocytes in vitro and subsequent preimplantation embryo development. J Cell Physiol. 2018; 233:5023–33. 10.1002/jcp.26367 [DOI] [PubMed] [Google Scholar]
- 38.Kwak SS, Cheong SA, Jeon Y, Lee E, Choi KC, Jeung EB, Hyun SH. The effects of resveratrol on porcine oocyte in vitro maturation and subsequent embryonic development after parthenogenetic activation and in vitro fertilization. Theriogenology. 2012; 78:86–101. 10.1016/j.theriogenology.2012.01.024 [DOI] [PubMed] [Google Scholar]
- 39.Miyamoto K, Sato EF, Kasahara E, Jikumaru M, Hiramoto K, Tabata H, Katsuragi M, Odo S, Utsumi K, Inoue M. Effect of oxidative stress during repeated ovulation on the structure and functions of the ovary, oocytes, and their mitochondria. Free Radic Biol Med. 2010; 49:674–81. 10.1016/j.freeradbiomed.2010.05.025 [DOI] [PubMed] [Google Scholar]
- 40.Goud AP, Goud PT, Diamond MP, Gonik B, Abu-Soud HM. Reactive oxygen species and oocyte aging: role of superoxide, hydrogen peroxide, and hypochlorous acid. Free Radic Biol Med. 2008; 44:1295–304. 10.1016/j.freeradbiomed.2007.11.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lu T, Zhou D, Gao P, Si L, Xu Q. Resveratrol attenuates high glucose-induced endothelial cell apoptosis via mediation of store-operated calcium entry. Mol Cell Biochem. 2018; 442:73–80. 10.1007/s11010-017-3194-7 [DOI] [PubMed] [Google Scholar]
- 42.Kong XX, Fu YC, Xu JJ, Zhuang XL, Chen ZG, Luo LL. Resveratrol, an effective regulator of ovarian development and oocyte apoptosis. J Endocrinol Invest. 2011; 34:e374–81. [DOI] [PubMed] [Google Scholar]
- 43.Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, Messadeq N, Milne J, Lambert P, Elliott P, Geny B, Laakso M, Puigserver P, Auwerx J. Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell. 2006; 127:1109–22. 10.1016/j.cell.2006.11.013 [DOI] [PubMed] [Google Scholar]
- 44.Mizuguchi Y, Hatakeyama H, Sueoka K, Tanaka M, Goto YI. Low dose resveratrol ameliorates mitochondrial respiratory dysfunction and enhances cellular reprogramming. Mitochondrion. 2017; 34:43–48. 10.1016/j.mito.2016.12.006 [DOI] [PubMed] [Google Scholar]
- 45.Strickland L, von Dassow G, Ellenberg J, Foe V, Lenart P, Burgess D. Light microscopy of echinoderm embryos. Methods Cell Biol. 2004; 74:371–409. 10.1016/S0091-679X(04)74016-9 [DOI] [PubMed] [Google Scholar]
- 46.Ou XH, Li S, Wang ZB, Li M, Quan S, Xing F, Guo L, Chao SB, Chen Z, Liang XW, Hou Y, Schatten H, Sun QY. Maternal insulin resistance causes oxidative stress and mitochondrial dysfunction in mouse oocytes. Hum Reprod. 2012; 27:2130–45. 10.1093/humrep/des137 [DOI] [PubMed] [Google Scholar]