

Abstract
Polypyrimidine tract binding protein, PTB/hnRNP I, is involved in pre-mRNA processing in the nucleus and RNA localization and translation in the cytoplasm. In this report, we demonstrate that PTB shuttles between the nucleus and cytoplasm in an energy-dependent manner. Deletion mutagenesis demonstrated that a minimum of the N terminus and RNA recognition motifs (RRMs) 1 and 2 are necessary for nucleocytoplasmic shuttling. Deletion of RRM3 and 4, domains that are primarily responsible for RNA binding, accelerated the nucleocytoplasmic shuttling of PTB. Inhibition of transcription directed by either RNA polymerase II alone or all RNA polymerases yielded similar results. In contrast, selective inhibition of RNA polymerase I did not influence the shuttling kinetics of PTB. Furthermore, the intranuclear mobility of GFP-PTB, as measured by fluorescence recovery after photobleaching analyses, increased significantly in transcriptionally inactive cells compared with transcriptionally active cells. These observations demonstrate that nuclear RNA transcription and export are not necessary for the shuttling of PTB. In addition, binding to nascent RNAs transcribed by RNA polymerase II and/or III retards both the nuclear export and nucleoplasmic movement of PTB. The uncoupling of PTB shuttling and RNA export suggests that the nucleocytoplasmic shuttling of PTB may also play a regulatory role for its functions in the nucleus and cytoplasm.
INTRODUCTION
Heterogeneous nuclear ribonucleoproteins (hnRNPs) are associated with RNA polymerase transcription in the nucleus and have been shown to be involved in the biogenesis and nucleocytoplasmic transport of mRNA (reviewed by Dreyfuss et al., 1993). Immunofluorescence studies of interphase HeLa nuclei revealed that although most hnRNPs are localized to the nucleus, a small subset shuttles between the nucleus and cytoplasm (Pinol-Roma and Dreyfuss, 1992; Michael et al., 1995b). Shuttling hnRNPs are involved not only in nuclear functions but also play other cellular roles, such as the transport of mature RNAs to the cytoplasm, mRNA translation, and regulation of mRNA stability (reviewed by Krecic and Swanson, 1999).
Polypyrimidine tract binding protein (PTB/hnRNP I), a member of the hnRNP family, is a 57-kDa protein that preferentially binds to the pyrimidine-rich sequences of RNA (Gil et al., 1991; Patton et al., 1991; Ghetti et al., 1992). Immunolabeling and green fluorescent protein (GFP)-tagged expression demonstrated that PTB is diffusely distributed throughout the nucleoplasm and is also concentrated in the perinucleolar compartment (PNC) (Ghetti et al., 1992; Matera et al., 1995; Huang et al., 1997). PTB consists of an amino-terminal nuclear localization sequence (NLS) (Romanelli et al., 1997) and four RNA recognition motifs (RRMs) (Ghetti et al., 1992). Structural and functional studies of the four RRMs revealed that the central part of PTB (RRM2) plays a major role in protein–protein interactions, whereas the C-terminal part of PTB (RRM3 and 4) is necessary for specific and efficient RNA binding (Perez et al., 1997; Oh et al., 1998; Conte et al., 2000; Kim et al., 2000b).
PTB has been shown to be involved in multiple steps of pre-mRNA processing, including regulation of pre-mRNA splice site selection for a number of genes (reviewed in Wagner and Garcia-Blanco, 2001), regulation of pre-mRNA polyadenylation (Lou et al., 1996, 1998; Moreira et al., 1998), and site-specific RNA localization in Xenopus embryos (Cote et al., 1999). In addition, PTB interacts with internal ribosomal entry sites of mRNA in the cytoplasm to regulate translation (Jang and Wimmer, 1990; Hellen et al., 1994; Witherell and Wimmer, 1994; Kaminski et al., 1995; Ito et al., 1998; Hunt and Jackson, 1999; Ito and Lai, 1999; Kim and Jang, 1999; Lou et al., 1999; Gosert et al., 2000). Recently, Kim et al. (2000a) and Giraud et al. (2001) showed that PTB has differential effects on the translation of internal ribosomal entry site-containing cellular mRNAs. Thus, it appears that PTB serves as a bridge between RNAs and a variety of cellular factors to fulfill cellular functions in both the nucleus and cytoplasm.
Shuttling proteins, including hnRNP A1 and SR proteins, have been shown to bind mRNA in both the nucleus and cytoplasm (Pinol-Roma and Dreyfuss, 1992; Caceres et al., 1997; Izaurralde et al., 1997). Immunoelectron microscopy analyses of Balbani ring transcripts in Chironomus tentans demonstrated that mRNA is exported as an RNP particle containing hrp36, an hnRNP related to mammalian hnRNP A1 (Visa et al., 1996). These observations led to the model that hnRNP proteins are either actively involved in nuclear export of mRNA or are passengers on the exported RNP complexes. However, it remains unclear whether the shuttling of these proteins is closely coupled with RNA export and what processes regulate nucleocytoplasmic shuttling. To begin addressing these problems, we characterized the shuttling kinetics of PTB/hnRNP I, its minimal domains required for shuttling, and the relationship between RNA binding and its shuttling. We also compared PTB shuttling kinetics with other shuttling hnRNP proteins, including A1 and K, to better understand the relationship between the shuttling kinetics and different functional roles of these hnRNPs.
MATERIALS AND METHODS
Construction of GFP-GST Fusion Proteins
The fusion protein and subsequent deletion mutants containing GFP at the amino terminus of PTB were constructed by cloning polymerase chain reaction (PCR)-generated fragments, amplified with the use of Vent polymerase with specific primers, into the HindIII and BamHI restriction sites of the pEGFP-C1 vector (CLONTECH, Palo Alto, CA) (Huang et al., 1997). A PCR-generated glutathione S-transferase (GST) fragment was cloned in-frame into the BamHI site of the GFP-PTB mutants. Thus, all PTB fusion proteins had GFP at the amino terminus and GST at the carboxy terminus. The GFP fusion protein of hnRNP A1 was constructed by cloning PCR-generated fragments with specific primers into the HindIII and BamHI restriction sites of the pEGFP-C1 vector. The GFP fusion protein of hnRNP K was constructed by cloning PCR-generated fragments with specific primers into the EcoRI restriction site of pEGFP-C1 vector.
Cell Culture and Transfection
Human HeLa cells and mouse NIH 3T3 were grown in DMEM supplemented with 10% fetal calf serum (FCS). For transient transfection, expression constructs were transfected into HeLa cells by electroporation (Sambrook et al., 1989). Briefly, subconfluent cells in 100-mm culture dishes were collected by trypsinization and mixed with 4 μg of target DNA and 16 μg of sheared salmon sperm DNA. A 280-μl mixture of cells in DMEM with 10% FCS and DNA was electroporated in a Bio-Rad (Richmond, CA) electroporator at 250 V and 950 μF. Subsequently cells were seeded onto glass coverslips in 35-mm Petri dishes and incubated for 24 h before experimentation. For transcriptional inhibition studies, HeLa cells were incubated with either 4 μg/ml actinomycin D (act D) (Sigma, St. Louis, MO), 25 μg/ml 5,6-dichloro-1-β-d-ribofuranosylbenzimidazole (DRB) (Sigma), or 50 μg/ml α-amanitin (Roche Molecular Biochemicals, Indianapolis, IN) in DMEM containing 10% FCS.
Indirect Immunofluorescence
Cells were fixed for immunofluorescence assays 24 h post-transfection. The cells were washed with phosphate-buffered saline (PBS), fixed in freshly prepared 4% paraformaldehyde in PBS for 15 min, and then washed 3 × 10 min in PBS. The cells were then permeabilized with 0.5% Triton-X (in PBS) for 5 min and washed 3 × 10 min in PBS. The fixed, permeabilized cells were then incubated for 1 h at room temperature with either anti-PTB antibody SH54 (1:200), anti-hnRNP A1 antibody 4D10 (1:2000), or anti-hnRNP K antibody (generous gift from Dr. Serafin Pinol-Roma, Mount Sinai Hospital, Manhattan, NY) (1:5). Cells were rinsed 3 × 10 min in PBS and then incubated with fluorescein isothiocyanate-conjugated goat anti-mouse antibody (1:200) (Vector Laboratories, Burlingame, CA) for 1 h at room temperature followed by 3 × 10 min washes in PBS. The coverslips were mounted onto glass slides with mounting medium containing 90% glycerol in PBS adjusted to pH 8.0 with 0.2 M bicarbonate buffer and 1 mg/ml paraphenylenediamine (Sigma) as an antifade agent. Cells were observed on Zeiss Axiovert 135 microscope and images acquired with a SenSys cooled charge-coupled device camera (Photometrics, Tucson, AZ) with the use of Metamorph, version 4.5, software (Universal Imaging, West Chester, PA). Each experiment result in this report was reproduced with the use of multiple independent transfections and experiments and the cells shown are representative of the overall effects observed under each set of conditions.
Heterokaryon Assays
HeLa cells (transfected or nontransfected) were seeded on glass coverslips for 24 h, followed by coincubation with an equal number of untransfected mouse NIH 3T3 cells for 2 h. The cells were then further incubated for an additional 2 h in presence of 100 μg/ml cycloheximide (Sigma). Cell fusions were carried out by washing the cells with PBS, incubating them in 50% (wt/vol) polyethylene glycol 1500 (Roche Molecular Biochemicals) for 2 min, and rinsing with PBS (Pinol-Roma and Dreyfuss, 1992). Heterokaryons were incubated further for the indicated times in media containing 100 μg/ml cycloheximide before fixation. For transcriptional inhibition studies heterokaryons were incubated with both 100 μg/ml cycloheximide and either 4 μg/ml act D, 0.04 μg/ml act D, 25 μg/ml DRB, or 50 μg/ml α-amanitin in DMEM containing 10% FCS for 2 h before fusion and 1 or 2 h after fusion. Immunofluorescence was carried out as described above except that cells were stained with 4,6-diamidino-2-phenylindole (DAPI) (200 ng/ml) (Sigma) for 1 min and with rhodamine-phalloidin (660 ng/ml) (Molecular Probes, Eugene, OR) for 10 min to identify heterokaryons.
Photobleaching and Live Cell Imaging
Forty-eight hours after transfection, cells were maintained in DMEM supplemented with 30 mM HEPES, pH 7.1, to stabilize the pH of the medium during imaging. Experiments done in the presence of RNA inhibitors also contained either act D (4 μg/ml for 3 h), act D (0.04 μg/ml for 3 h), DRB (25 μg/ml for 2 h), or α-amanitin (50 μg/ml for 5 h) in the media before and during imaging. The 35-mm dishes with coverslip bottoms were directly mounted onto a Zeiss 510 confocal laser-scanning microscope equipped with an argon-krypton laser (Zeiss, Thornwood, NY). The medium was kept at 37°C with the use of an ASI 400 Air Stream incubator (Nevtek, Burnsville, VA). The 488-nm laser and a 63× plan Apo lens with a 1.4 numerical aperture were used in bleaching and imaging experiments. A laser power of 1.1% of 3.75 mW was used in image acquisitions, and 100% of 3.75 mW was used in photobleaching. The time for each image acquisition was 800 ms, which did not significantly influence the fluorescent intensity through multiple acquisitions. An area of 2 μm2 was bleached with an iteration of 80. Images were collected before, immediately following, and at 1-s intervals after bleaching. Generally, photobleach analyses of GFP-tagged molecules in living cells raise concerns because the results could be influenced by phototoxicity. Several studies investigating this problem have shown that photobleaching with a low laser power results in no significant damage to the examined cells (White and Stelzer, 1999; Kruhlak et al., 2000; Phair and Misteli, 2000). In addition, we have monitored cells >24 h after receiving similar and higher doses of laser irradiation that we used in these studies. The results showed that cells survived well and that some underwent mitosis during this period of time (Chen and Huang, 2001).
Quantitation of Fluorescence Intensity
Fluorescence intensity was measured with the use of Metamorph (Universal Imaging) imaging software.
Heterokaryon Assays.
The average intensities of the same sized region of the human and mouse nuclei of fused cells were measured in images taken under the same conditions for each data set. Ratio of fluorescence intensity (Fr) in the heterokaryon analyses was calculated as Fr = Fm − Fb/Fh − Fb. Fm is the average intensity of the mouse nucleoplasmic region, Fh is the average size intensity in the human nucleoplasmic region within same fused cell, and Fb is the background fluorescence intensity for each heterokaryon. When Fm equals Fh, Fr is 1, and the two nuclei are considered to be at equilibrium for the given protein.
Fluorescent Recovery after Photobleaching (FRAP) Assays.
The average intensities of the areas of interest in images, including before, immediately after, and a series of time points after bleaching, were measured under the same conditions for each data set. The relative fluorescence intensity (RFI) in the FRAP analyses was calculated as RFI = (NB/NBEN)/(NPB/NPEN). NB is the average intensity of the bleached nucleoplasmic area at various time points after bleaching. NBEN is the average intensity of the entire nucleus at the corresponding time points. NPB is the average intensity of the bleached nucleoplasmic area before bleaching. NPEN is the average intensity of the entire nucleus before bleaching. When NB/NBEN = NPB/NPEN, namely, when the RFI is 1, fluorescence recovery of the bleached nucleoplasm reaches 100% (Phair and Misteli, 2000). The NB of the images acquired immediately after bleaching were set at zero and were used to standardize the raw data in this report. With the use of this equation, we have taken into consideration the overall fluorescence change, if any, during subsequent image acquisitions.
RESULTS
PTB Protein Shuttles between Nucleus and Cytoplasm in an Energy-dependent Manner with Slower Kinetics Compared with hnRNP A1 or K
PTB was shown to shuttle between the nucleus and cytoplasm with the use of mammalian (human HeLa cells) and amphibian (Xenopus A6 kidney cells) heterokaryon assays (Michael et al., 1995b). To evaluate PTB shuttling in the mammalian cells, we used heterokaryon assays in which human HeLa cells were fused with mouse NIH 3T3 cells. The mouse nuclei act as newly introduced compartments for human nuclear proteins. The presence of human PTB in the mouse nuclei after fusion would indicate that PTB has moved between the nuclei and through the cytoplasm. Mouse cells were selected because mouse PTB protein shows 85–100% identity with its human counterpart within the individual RRM domains. It is therefore probable that the function and overall behavior of PTB are well conserved between human and mouse, and that human PTB should act similarly in both cell types. SH54, a monoclonal antibody (mAb) that recognizes only human (Huang et al., 1997) but not mouse PTB, makes it possible to specifically detect human PTB that has shuttled into mouse nuclei. In addition, human-mouse heterokaryons were incubated at 37°C in contrast to the 30°C incubation temperature used for human-amphibian heterokaryons (Michael et al., 1995b). Therefore, we believe that the human-mouse heterokaryon system more closely represents the normal physiological conditions for nuclear import and export in mammalian cells.
With the use of the human-mouse heterokaryon assays, we evaluated the nucleocytoplasmic shuttling of the endogenous PTB and GFP-tagged PTB (GFP-PTB). GFP-PTB transfected or untransfected HeLa cells were fused with NIH 3T3 cells to form heterokaryons and were incubated in the presence of cycloheximide to prevent new protein synthesis. Cells were fixed 2 h after heterokaryon formation and the localization of endogenous human PTB was detected with the use of immunolabeling with a specific anti-human PTB antibody, SH54, whereas GFP-PTB was visualized directly with the use of fluorescence microscopy. Cells were counterstained with the DNA dye DAPI, which reveals characteristic dense DNA clusters in the mouse nuclei but not in the human nuclei, allowing identification of fused mouse nuclei. The actin filaments within the cells were also stained with rhodamine-phalloidin to identify heterokaryons based on the fusion of their cellular cytoskeletons (our unpublished data).
When HeLa cells were fused with NIH 3T3 cells in the absence of protein synthesis, human PTB was detected in the mouse nuclei of the heterokaryons, demonstrating that hPTB was exported from the human nuclei and was reimported into the mouse nuclei (Figure 1, top). GFP-PTB transfected HeLa cells showed nucleocytoplasmic shuttling similar to endogenous PTB (Figure 1, second panel from top), suggesting that GFP tagging did not significantly change the shuttling properties of PTB. To ensure the reliability of the assay, known shuttling hnRNP proteins, including GFP-tagged hnRNP A1 and hnRNP K, were also examined and were shown to shuttle as observed previously (Michael et al., 1995a) (Figure 1, bottom). Interestingly, GFP-hnRNP A1 and GFP-hnRNP K showed comparable fluorescence intensities in both the mouse and human nuclei within 2 h after fusion, whereas PTB or GFP-PTB remains predominantly in the human nuclei at this time point. This finding demonstrates that the nucleocytoplasmic shuttling kinetics of PTB and GFP-PTB is slower than that of GFP-hnRNP A1 and -hnRNP K.
Figure 1.
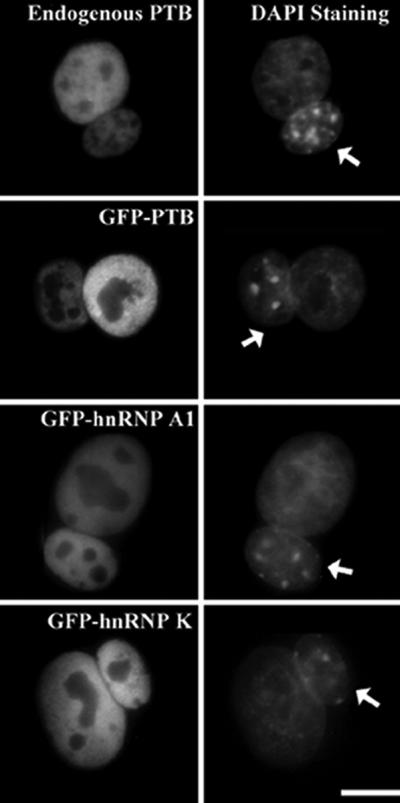
PTB and GFP-PTB shuttle between the nucleus and cytoplasm as assayed by human-mouse heterokaryons. Cells were analyzed 2 h after fusion. DNA was labeled with DAPI for differential staining patterns of human and mouse nuclei (arrows) within the heterokaryons. Bar, 10 μm.
To further characterize the shuttling kinetics of PTB protein as a function of time, heterokaryons were incubated at 37°C for 1, 2, 3, 4, or 5 h after fusion and the PTB labeling in the nuclei of fused cells were examined. The relative fluorescence intensities of the same sized nucleoplasmic regions within the mouse and human nucleus of a heterokaryon was measured with the use of Metamorph software (Universal Imaging). The ratio of relative Fr of the mouse nucleoplasm over the human nucleoplasm was calculated for each time point (see MATERIALS AND METHODS). An Fr value of 1 indicates that the shuttling of PTB between the two nuclei has reached equilibrium in terms of fluorescence intensity. PTB and GFP-PTB reach the equilibrium at 4 h postfusion, in contrast to hnRNP A1 and hnRNP K, both of which reach the equilibrium within 1 h (Figure 2, A and B). These results further demonstrate that the shuttling of PTB is slower than that of hnRNP A1 and K.
Figure 2.

Nucleocytoplasmic shuttling of PTB is slower than that of hnRNP K and A1 and is energy dependent as assayed by human-mouse heterokaryon analyses. DNA of the cells was stained with DAPI for differential staining of human and mouse nuclei (arrows) within the heterokaryons (A, bottom, and C, bottom). (A) Time-course dependent heterokaryon assays determined the time required for PTB to attain equilibrium in both nuclei (A, top). (B) Quantitative analysis of the time-course experiments represented in A and similar experiments with the use of HeLa transfected with GFP-tagged hnRNP A1 or K showed that PTB attains equilibrium in both nuclei 4 h postfusion compared with 2 h for hnRNPs A1 and K. The Fr (y-axis) was measured as fluorescence intensity in the mouse nuclei (Fm)/fluorescence intensity in the human nuclei (Fh) within a same sized area over time (x-axis). (C) Nuclear export of PTB is an energy-dependent process. Heterokaryons were incubated at either 37 or 4°C for 3 h after fusion (C, top).
To determine whether the nucleocytoplasmic shuttling of the PTB protein is an energy-dependent process, heterokaryons were incubated in a 4°C chamber for 3 h immediately after fusion and were then subjected to immunofluorescence analyses. In contrast to heterokaryons incubated at 37°C where both nuclei contain PTB at 3 h postfusion, no labeling was detected either in the mouse nuclei or in the cytoplasm of the heterokaryons at 4°C (Figure 2C). These results suggest that the nuclear export of PTB is an energy-dependent process.
Steady-State Cellular Distribution of PTB Protein Is Insensitive to Transcription Inhibition
Nuclear shuttling hnRNPs have been classified into two groups, transcription sensitive and transcription insensitive, based on their steady-state cellular distribution upon inhibition of RNA polymerase (pol) II transcription (reviewed by Dreyfuss et al., 1993). Proteins that become cytoplasmically localized during transcriptional inhibition belong to the transcription-sensitive group (e.g., hnRNP A1), whereas proteins that remain predominantly nuclear localized belong in the transcription-insensitive group (e.g., hnRNP K).
To determine to which group PTB belongs, HeLa cells were treated with various transcription inhibitors, including α-amanitin, actinomycin D, and DRB before examining the distribution of PTB. α-Amanitin binds specifically to RNA pol II large subunit and, at a higher concentration, to RNA pol III large subunit resulting in proteolytic degradation of the polymerases (Nguyen et al., 1996). Actinomycin D intercalates DNA and inhibits all three classes of transcription at a high concentration, but selectively inhibits RNA pol I transcription at a low concentration (Perry, 1963). DRB is a kinase inhibitor that blocks phosphorylation of the carboxy terminal domain of the large subunit of RNA pol II and inhibits pol II transcription (Sehgal et al., 1976; Dubois et al., 1994).
When HeLa cells were treated with any of these inhibitors, the predominantly nuclear distribution of PTB was not altered (Figure 3, left). This finding is similar to the transcription-insensitive distribution of hnRNP K (Figure 3, middle) (Michael et al., 1997), and is in contrast to the transcription-sensitive distribution of endogenous hnRNP A1, which accumulates in the cytoplasm upon transcriptional inhibition (Figure 3, right) (Pinol-Roma and Dreyfuss, 1991, 1992). Similar localizations of PTB, hnRNP A1, and hnRNP K were also observed in MG63 (osteosarcoma) and Wacar (normal human skin fibroblast) cells during transcriptional inhibition (our unpublished data), suggesting that our findings in HeLa cells were not due to cell type biases. These results, therefore, categorize PTB in the transcription-insensitive group of hnRNPs as classified by Dreyfuss et al. (1993) because the steady state nuclear distribution pattern of this class of protein is independent of transcriptional activity. However, the steady-state distributions of hnRNP proteins during transcriptional inhibition does not address whether there are any changes in the kinetics of the nucleocytoplasmic shuttling of these proteins.
Figure 3.

Nuclear localization of PTB is not affected by transcriptional inhibition. HeLa cells were incubated in the absence (top) or in the presence of actinomycin D (4 μg/ml) (second panel) for 4 h, DRB (25 μg/ml) (third panel) for 4 h, or α-amanitin (50 μg/ml) (bottom) for 5 h. The cells were fixed and the localization of PTB, hnRNP K, and hnRNP A1 was examined by indirect immunofluorescence with anti-PTB mAb (left), anti-hnRNP K polyclonal antibody (middle), or anti-hnRNP A1 mAb (right) and fluorescein isothiocyanate-conjugated secondary antibody. Bar, 10 μm.
RRM2 Is Required for Nucleocytoplasmic Shuttling of PTB
To begin to understand the mechanisms that regulate the nucleocytoplasmic shuttling of PTB, we first analyzed the domains of PTB that are required for shuttling. A series of deletion mutant fusion proteins were generated and their shuttling kinetics was determined by calculating Fr values at various time points postfusion in heterokaryon assays. PTB consists primarily of an N-terminal NLS and four RRM domains (Figure 5A). Successive deletions from the C terminus were made to remove specific RRM domains while retaining the NLS. Because protein fragments <50 kDa may undergo passive diffusion through nuclear pores (Talcott and Moore, 1999), we designed mutant fusion protein constructs to increase the molecular sizes of the smallest mutant to >50 kDa. GFP (27 kDa) was fused to the N termini and GST (29 kDa) was fused to the C termini of the full-length and all PTB mutant fragments to maintain uniformity (Figure 5A).
Figure 5.

RRM2 domain is required for nucleocytoplasmic shuttling of PTB and deletion of RNA binding domains accelerates PTB shuttling. (A) Diagram of the full-length and deletion mutants. (B) Localization of PTB mutant fusion proteins was examined 2 h post fusion. Top panels are the images of HeLa cells were transfected with GFP-PTB-GST and the bottom panels show DNA staining by DAPI. (C) Heterokaryon shuttling kinetics of GFP-PTB-GST mutants were quantitative are analyzed as the ratios of fluorescence intensities between the HeLa cell nuclei and the NIH 3T3 cell nuclei 2 h after fusion. Bar, 10 μm.
Due to the presence of two large fusion tags, it was important to verify that GFP-PTB-GST construct behaved similarly to the endogenous protein. To do so, we first examined the cellular localization of GFP-PTB-GST in HeLa cells by immunolabeling with the use of SH54, which recognized both the endogenous and fusion protein. Both the endogenous PTB and GFP-PTB-GST localized to the same nuclear regions, including the PNC and the nucleoplasm (Figure 4A). A direct visualization of GFP-PTB alone also showed PNC and nuclear localization (Huang et al., 1997). In addition, we compared the nucleocytoplasmic shuttling kinetics of GFP-PTB-GST and endogenous PTB in time-course heterokaryon assays. As observed with endogenous PTB (Figure 2, A and B), the fluorescence intensity of GFP-PTB-GST in human-mouse heterokaryon nuclei reaches equilibrium at 4 h postfusion (Figure 4, B and C). Furthermore, GFP-PTB-GST, similarly to PTB, remains predominantly nuclear localized upon transcriptional inhibition (our unpublished data). These results demonstrate that the addition of the GFP-GST tags to PTB does not alter its intranuclear localization and its nuclear export/import kinetics, suggesting that GFP-PTB-GST behaves similarly to the endogenous PTB.
Figure 4.

GFP-GST–tagged PTB behaves similarly to the endogenous PTB. (A) Both GFP-GST–tagged and endogenous PTB were similarly localized to PNCs (arrowheads) and nuclei as visualized in transfected HeLa cells with the use of the anti-PTB antibody SH54. (B) Shuttling kinetics of GFP-PTB-GST was analyzed in heterokaryon assays as a function of time. Localization of the fusion protein in the heterokaryons was visualized by direct immunofluorescence (top). The cells were stained with DAPI for differential staining of human and mouse nuclei (bottom). Arrows indicate mouse nuclei. (C) The Fr (y-axis) between mouse and human nuclei over time (x-axis) indicates that GFP-GST PTB exhibits shuttling kinetics similar to endogenous PTB. Bar, 10 μm.
HeLa cells were transfected with each of the GFP-GST–tagged PTB mutants (Figure 5A) and the shuttling kinetics of the mutants was evaluated with the use of heterokaryon assays. Cells were fixed and examined at various time points after fusion with the use of fluorescence microscopy. The Fr value between the mouse and human nuclei at 2 h after fusion is nearly identical for the endogenous PTB, full-length GFP-PTB-GST, and the GFP-PTB-GST-1 mutant that lacks RRM4 (Figure 5, B and C), suggesting that RRM4 is not required for the nucleocytoplasmic shuttling of PTB. In comparison, removing both RRM3 and RRM4 (GFP-PTB-GST-2) dramatically altered the kinetics of the nucleocytoplasmic shuttling. The GFP-PTB-GST-2 mutant exhibited a fourfold higher Fr value between mouse and human nucleoplasm than that observed for endogenous PTB, full-length GFP-GST–tagged PTB and GFP-PTB-GST-1 mutant, 2 h after fusion (Figure 5, B and C). This finding indicates that PTB shuttles faster in the absence of RRM3 and 4. In comparison, deletion of RRM2, 3, and 4 (GFP-PTB-GST-3) resulted in a complete abolition of PTB shuttling and the mutant PTB remains nuclear (Figure 5, B and C). These findings indicate that the truncation of RRM3 and 4, domains required for efficient RNA binding, accelerates nucleocytoplasmic shuttling, and that PTB requires a minimum of the N-terminal NLS sequence along with the RRM1 and 2 domains for the shuttling.
RNA Binding Retards Nucleocytoplasmic Shuttling of PTB
The acceleration of PTB shuttling upon the deletion of RRM3, the major RNA binding domain of PTB (Perez et al., 1997; Oh et al., 1998; Conte et al., 2000), suggested that RNA binding might modulate the nucleocytoplasmic shuttling of PTB. To further address this possibility, we examined the shuttling kinetics of PTB in the absence of nascent RNA synthesis. Untransfected or transfected HeLa cells were treated with RNA polymerase inhibitors (α-amanitin, actinomycin D, or DRB) and cycloheximide to inhibit both RNA and protein synthesis 2 h before fusion with NIH 3T3 cells. After fusion, heterokaryons were continuously incubated in the presence of RNA and protein synthesis inhibitors for an additional 1 or 2 h and the shuttling kinetics of PTB and mutants were evaluated by calculating the Fr of mouse over human nuclei. Endogenous PTB reached equilibrium in the transcriptionally inhibited heterokaryon nuclei faster than in transcriptionally active cells (Figure 6, A and B). The alteration of PTB shuttling kinetics is mainly due to the inhibition of RNA pol II and/or III because selective inhibition of RNA pol I with a lower concentration of actinomycin D at 0.04 μg/ml did not result in detectable changes (our unpublished data). These findings suggest that the absence of nascent RNA in the nucleus accelerates PTB shuttling. The GFP-PTB-GST-2 mutant, which lacks RRM3 and RRM4 and efficient RNA binding, showed a further acceleration in its shuttling kinetics (Figure 5, B and C). It reaches equilibrium between human and mouse nuclei within 1 h in the presence of RNA pol II and/or III inhibitors (Figure 6, A and B) as opposed to 2 h in the absence of the same drugs (Figure 5, B and C). A PTB mutant lacking RRM3 and 4 has been shown to weakly bind nascent RNA (Perez et al., 1997) and these weak interactions in GFP-PTB-GST-2 mutant could have been abolished upon transcriptional inhibition, resulting in further acceleration of shuttling. Altogether, these observations demonstrate that a reduction of RNA binding due to either mutation or transcriptional inhibition does not prevent PTB from shuttling. On the contrary, it accelerates this process, suggesting that RNA binding is not required for the nuclear export of PTB, and that binding to nascent RNAs retards PTB export from the nucleus.
Figure 6.

RNA binding is not required for and retards the nuclear export of PTB. (A) Untransfected or transfected HeLa cells were fused with NIH 3T3 cells and incubated in presence of cycloheximide and actinomycin D (4 μg/ml) to inhibit RNA polymerase I, II, and III for 2 h before and after fusion (except where indicated). Top, distribution of endogenous PTB, as detected with the use of human specific anti-PTB antibody SH54, or mutant fusion proteins. Bottom, DNA staining with the use of DAPI. (B) Quantitative analyses of shuttling kinetics in heterokaryon assays were performed by calculating Fr (y-axis) between mouse and human nuclei. These data indicated that all PTB mutants shuttle at a faster rate in presence of transcription inhibitors. Bar, 10 μm.
To determine whether the inhibition of RNA pol II transcription has similar effects on another shuttling RNP protein whose steady-state nuclear localization is not altered during transcription inhibition, we examined the shuttling properties of hnRNP K. HeLa cells were transfected with GFP-hnRNP K and heterokaryon assays were performed in the presence or absence of actinomycin D at a concentration that inhibits all RNA polymerases. As seen in Figure 7, A and B, no significant changes were detected in the nucleocytoplasmic shuttling kinetics of hnRNP K when transcription was inhibited (Figure 7, A and B). Similar results were obtained when cells were treated with specific RNA pol II inhibitors DRB and α-amanitin (our unpublished data). These findings demonstrate that, in contrast to PTB, the nucleocytoplasmic shuttling kinetics of hnRNP K is independent of nascent RNA synthesis.
Figure 7.

Transcription is not required and has no effect on nucleocytoplasmic shuttling kinetics of hnRNP K. (A) Transfected HeLa and NIH 3T3 cells were incubated in the presence of cycloheximide and actinomycin D (4 μg/ml) for 2 h before and 30 min after fusion. The distribution of GFP-hnRNP K was examined (top). The cells were stained with DAPI to differentiate staining of human and mouse nuclei (bottom). Arrows indicate mouse nuclei. (B) Shuttling kinetics of GFP-hnRNP K was quantitatively analyzed by calculating ratio of fluorescent intensity between the mouse and human nuclei 30 min after fusion (Fr, y-axis). Bar, 10 μm.
Mobility of Nucleoplasmic PTB Protein Increases upon Inhibition of Transcription
Because the absence of RNA binding accelerated PTB shuttling kinetics in heterokaryon assays, we were interested in whether similar acceleration of PTB mobility could be observed in single living cells. To do so, we examined the nucleoplasmic mobility of GFP-PTB-GST as measured by FRAP analyses, which provide information regarding the rate of replacement of bleached GFP-PTB-GST with emission-competent proteins from outside of the bleached zone. HeLa cells were transfected with GFP-PTB-GST and FRAP analyses were performed on the live cells by bleaching a small region of the nucleoplasm with a 488-nm laser pulse (Chen and Huang, 2001). A series of images was acquired at 1-s intervals up to 2 min. Changes in relative fluorescence intensity within the bleached areas were quantitatively measured at each time point for 10 or more representative cells (see MATERIALS AND METHODS).
The fluorescence recovery of GFP-PTB-GST was rapid (Figure 8A) and the maximal recovery was complete after ∼75 s (Figure 8B). When transfected cells were treated with transcription inhibitors that inhibit RNA pol II and/or pol III, the recovery after photobleaching was significantly accelerated (Figure 8B) and was complete after ∼30 s (Figure 8B). In contrast, the GFP-GST-PTB in cells under selective RNA pol I transcription inhibition showed a similar recovery to untreated cells (Figure 8B). These findings in live cells demonstrate that the inhibition of RNA pol II and/or III accelerates the nucleoplasmic movement of PTB, whereas RNA pol I transcription has little effect on PTB dynamics in the nucleoplasm.
Figure 8.

FRAP analyses demonstrate that the absence of RNA pol II and/or pol III transcripts increases the nucleoplasmic mobility of PTB. (A) Small regions of the nucleoplasm of HeLa cells transfected with GFP-PTB were photobleached and images were taken immediately before, after, and at 1-s intervals after bleaching. Arrows indicate the sites of bleaching and numbers represent the time(s) after photobleaching. (B) Quantitative analyses of FRAP demonstrate the FRAP rate of PTB increases upon inhibition of RNA pol II and/or pol III transcription, whereas inhibition of RNA pol I has no significant effect on FRAP rate of PTB.
DISCUSSION
Nuclear Localization of PTB Is Independent of Transcription and PTB Shuttles between Nucleus and Cytoplasm in an Energy-dependent Manner
Using human-mouse heterokaryon assays, we analyzed the kinetics of PTB shuttling between the nucleus and cytoplasm by monitoring the changes in the ratios (Fr) of fluorescence intensities of mouse over human nuclei at various time points after fusion. When Fr reaches 1, the nucleocytoplasmic shuttling protein has reached equilibrium in the heterokaryon assay. The time period required to reach equilibrium is indicative of the rate of the nucleocytoplasmic shuttling of the protein. We found that PTB reaches equilibrium between the mouse and human nuclei at 4 h after fusion, a rate that is significantly slower than hnRNP A1 or K, both of which reach equilibrium within 1 h. Another splicing protein, SR protein, reaches equilibrium within 2 h postfusion (Caceres et al., 1997). In comparison, the shuttling of the nucleolar protein nucleolin, involved in rRNA processing and assembly, occurs much more slowly and does not reach equilibrium until 72 h after fusion (Borer et al., 1989). These observations demonstrate that individual RNA binding proteins have different nucleocytoplasmic shuttling dynamics in mammalian cells, probably indicative of differences in their functions, or differences in mechanisms that facilitate the shuttling. For example, although both PTB and hnRNP A1 have been implicated in pre-mRNA processing, the differences in their shuttling dynamics are consistent with that they play very different roles in these processes as previously demonstrated in pre-mRNA splicing analyses. Furthermore, the shuttling of PTB requires energy because incubation at 4°C prevents PTB from leaving the human nucleus. Export of many other RNA binding proteins, including SR proteins (Borer et al., 1989), and hnRNP A1 (Michael et al., 1995a), have been shown to require energy and the nuclear import of classical NLS bearing proteins have also been blocked at low temperature (Breeuwer and Goldfarb, 1990). However, at this time it remains to be determined at which steps energy is required for PTB shuttling.
Although PTB does move through the cytoplasm, it, like most nuclear shuttling proteins, is predominantly detected in the nucleus at steady state. This distribution is probably due to a balanced regulation of its rate of nuclear import and export (reviewed by Nigg, 1997). The exclusive nuclear staining patterns for some shuttling proteins are dependent upon active RNA pol II transcription, because inhibition of pol II transcription causes these proteins to accumulate to the cytoplasm (Pinol-Roma and Dreyfuss, 1991, 1992). Shuttling RNPs were therefore classified into two categories, the transcription-sensitive and -insensitive groups, according to their steady-state cellular distribution upon transcription inhibition. An early report showed that PTB becomes more cytoplasmically localized upon transcription inhibition and, thus, belongs to the transcription-sensitive group (Michael et al., 1995b). However, in our studies, inhibition of RNA pol II transcription with the use of any of the three drugs individually did not result in any changes in the predominantly nuclear distribution of PTB either in HeLa or in two other cell types. The same observations were made either by immunolabeling with the use of two different monoclonal antibodies recognizing PTB (data for the second antibody not shown) or in GFP-PTB transfected cells. These findings suggest that PTB, like hnRNP K, is a member of the shuttling nuclear protein family whose steady-state cellular distribution is not affected by transcription. The differences from the previous studies could be due to the specific differences within the HeLa cell line used.
RRM2 Plays a Key Role in PTB Export
The steady state nuclear localization of PTB is probably the result of a stronger nuclear association than cytoplasmic association of the protein and nuclear import-export regulation. Sequence analysis of PTB reveals that the N terminus of the protein contains a NLS, but a nuclear export sequence has not been identified. Deletion mutagenesis and domain combination studies showed that a combination of the NLS with either RRMs 2, 3, or 4 results in cytoplasmic accumulation, whereas the combination of the NLS and RRM1 shows the nuclear localization pattern (Huang et al., 1997; Perez et al., 1997). These studies suggest that the NLS alone is not sufficient for the steady-state nuclear localization, but instead that the combination of the NLS and RRM1 constitutes the nuclear targeting and association domain. In this report, we show that a PTB mutant containing the N terminus and RRM1 stays exclusively in the nucleus and does not shuttle. However, the mechanism by which RRM1 and NLS constitute the nuclear association force remains unclear. RNA binding of PTB has been shown not to involve RRM1 (Perez et al., 1997), suggesting that its principal nuclear association is not via RNA binding and instead, may be due to protein–protein interactions. Interestingly, binding to nascent RNA does increase the nuclear retention of PTB (as discussed below). Addition of RRM2 to the NLS plus RRM1 restores nucleocytoplasmic shuttling, suggesting that RRM2 is the key domain that facilitates the nuclear export of PTB. RRM2 has been shown to be necessary for PTB dimerization and protein–protein interactions (Perez et al., 1997; Oh et al., 1998; Kim et al., 2000b), whereas RRM3 and 4 have been shown to be involved in RNA binding (Perez et al., 1997; Oh et al., 1998; Conte et al., 2000). A mutant with RRM3 and 4 alone binds to RNA with affinity similar to the wild-type protein (Perez et al., 1997; Conte et al., 2000), suggesting that RRM1 and 2 are not essential for specific and efficient RNA binding. Therefore, our observation that the N-terminal NLS and RRM1 and 2 are necessary and sufficient for PTB shuttling, suggests that the export of PTB is probably through protein–protein interactions rather RNA binding. Furthermore, hnRNP L, a close homolog of PTB, has been shown to interact with PTB in yeast two-hybrid system and in vitro pull down assays. The results indicate that RRM 1 and 2 domains of PTB are required for its interaction with hnRNP L (Hahm et al., 1998; Kim et al., 2000b). This interaction might play a role in nucleocytoplasmic shuttling of PTB.
RNA Binding Is not Required for Nucleocytoplasmic Shuttling of PTB
hnRNP proteins have been implicated in a variety of functions in RNA metabolism, including pre-mRNA processing, transport, localization, translation, and degradation. Several of these proteins, including hnRNP A1, A2, D, E, I (PTB), and K, shuttle between the nucleus and the cytoplasm (Nakielny and Dreyfuss, 1999). These findings have led to a model in which these hnRNPs are either active participants in the nuclear export of mRNA or are exported as passengers on mRNA-containing RNP complexes and thus follow RNAs into the cytoplasm where they carry out further functions.
To analyze the coupling of RNA export and PTB shuttling, we have examined the effect of RNA binding upon the dynamics of PTB with the use of three different approaches. Deletion mutagenesis that abolishes the key RNA binding domains or transcriptional inhibition that blocks nascent RNA synthesis resulted in acceleration of nucleocytoplasmic shuttling of PTB. Furthermore, transcriptional inhibition significantly increases the nucleoplasmic mobility of PTB in living cells as measured by fluorescence recovery after photobleaching. These observations demonstrate that the lack of binding to nascent RNA in the nucleus allows a more rapid nucleoplasmic movement and nucleocytoplasmic shuttling of PTB, suggesting that RNA binding increases the nuclear retention of PTB and that mRNA export is not required for nucleocytoplasmic shuttling of the protein. However, it remains a distinct possibility that PTB is associated with exporting RNA in normal circumstances. The continuous shuttling of PTB in the absence of nascent RNA may represent a way of regulating the activation or availability of the protein in the nuclear and cytoplasmic compartments. These findings are consistent with recent reports showing that the export of hnRNP A1 from the nucleus is independent of mRNA synthesis and processing (Lichtenstein et al., 2001; Vautier et al., 2001). An increasing number of transcription factors and cell cycle-related proteins have been shown to shuttle between the nucleus and cytoplasm. It has been proposed that shuttling helps regulate the activation of these proteins (Gama-Carvalho and Carmo-Fonseca, 2001), a model that could also apply to PTB.
In contrast to PTB and hnRNP A1, the nucleocytoplasmic shuttling kinetics of hnRNP K was not significantly affected by inhibition of RNA pol II transcription. The function of hnRNP K in RNA metabolism in the nucleus is much less understood than those of PTB and hnRNP A1.
Although hnRNP K was identified as a component of hnRNP particles (Swanson and Dreyfuss, 1988), it actually binds more tightly to DNA than RNA (Tomonaga and Levens, 1995). It has been implicated in the transcription of c-myc (Hobert et al., 1994), and Sp1 and Sp2 (Du et al., 1998), and in the interconversion of double- and single-stranded DNA during transcription (Tomonaga and Levens, 1996). Thus, hnRNP K appears to act more as a DNA-binding than RNA-binding protein, although hnRNP K has recently been shown to bind 15-lipooxygenase mRNA and regulate its translation in cytoplasm (Ostareck-Lederer et al., 1998). It remains to be understood why the nucleocytoplasmic shuttling of hnRNP K is not affected by RNA synthesis.
In summary, we have shown that PTB shuttles between the nucleus and cytoplasm in an energy-dependent manner. At steady state, PTB remains predominantly nuclear during transcriptional inhibition. The nuclear localization of the protein is mainly attributable to RRM1 and is independent of RNA binding, although the presence of nascent RNA increases the nuclear residence time of PTB. RRM2 plays a key role in nuclear export, most probably through protein–protein interactions. Furthermore, the nucleocytoplasmic shuttling of PTB does not require RNA binding. This uncoupling of RNA export and PTB shuttling suggests that the nucleocytoplasmic exchange of PTB may also play regulatory roles by controlling the protein proportions between the nucleus, where PTB plays numerous roles in pre-RNA processing, and the cytoplasm, where it is involved in RNA localization and translation.
ACKNOWLEDGMENTS
We thank Dr. S. Adam for helpful discussions. We are also grateful to Dr. S. Pinol-Roma for anti-hnRNP K antibody. This work is supported by grants to S.H. from the National Cancer Institute (1 RO1 CA-77560-01A1 and 5 K01 CA-74988-03).
REFERENCES
- Borer RA, Lehner CF, Eppenberger HM, Nigg EA. Major nucleolar proteins shuttle between nucleus and cytoplasm. Cell. 1989;56:379–390. doi: 10.1016/0092-8674(89)90241-9. [DOI] [PubMed] [Google Scholar]
- Breeuwer M, Goldfarb DS. Facilitated nuclear transport or histone H1 and other small nucleophilic proteins. Cell. 1990;60:999–1008. doi: 10.1016/0092-8674(90)90348-i. [DOI] [PubMed] [Google Scholar]
- Caceres JF, Misteli T, Screaton GR, Spector DL, Krainer AR. Role of the modular domains of SR proteins in subnuclear localization and alternative splicing specificity. J Cell Biol. 1997;138:225–238. doi: 10.1083/jcb.138.2.225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen D, Huang S. Nucleolar components involved in ribosome biogenesis cycle between the nucleolus and nucleoplasm in interphase cells. J Cell Biol. 2001;153:169–176. doi: 10.1083/jcb.153.1.169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conte MR, Grune T, Ghuman J, Kelly G, Ladas A, Matthews S, Curry S. Structure of tandem RNA recognition motifs from polypyrimidine tract binding protein reveals novel features of the RRM fold. EMBO J. 2000;19:3132–3141. doi: 10.1093/emboj/19.12.3132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cote CA, Gautreau D, Denegre JM, Kress TL, Terry NA, Mowry KL. A Xenopus protein related to hnRNP I has a role in cytoplasmic RNA localization. Mol Cell. 1999;4:431–437. doi: 10.1016/s1097-2765(00)80345-7. [DOI] [PubMed] [Google Scholar]
- Dreyfuss G, Matunis MJ, Pinol-Roma S, Burd CG. hnRNP proteins and the biogenesis of mRNA. Annu Rev Biochem. 1993;62:289–321. doi: 10.1146/annurev.bi.62.070193.001445. [DOI] [PubMed] [Google Scholar]
- Du Q, Melnikova IN, Gardner PD. Differential effects of heterogeneous nuclear ribonucleoprotein K on Sp1- and Sp3-mediated transcriptional activation of a neuronal nicotinic acetylcholine receptor promoter. J Biol Chem. 1998;273:19877–19883. doi: 10.1074/jbc.273.31.19877. [DOI] [PubMed] [Google Scholar]
- Dubois MF, Nguyen VT, Bellier S, Bensaude O. Inhibitors of transcription such as 5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole and isoquinoline sulfonamide derivatives (H-8 and H-7) promote dephosphorylation of the carboxyl-terminal domain of RNA polymerase II largest subunit. J Biol Chem. 1994;269:13331–13316. [PubMed] [Google Scholar]
- Gama-Carvalho M, Carmo-Fonseca M. The rules and roles of nucleocytoplasmic shuttling proteins. FEBS Lett. 2001;498:157–163. doi: 10.1016/s0014-5793(01)02487-5. [DOI] [PubMed] [Google Scholar]
- Ghetti A, Pinol-Roma S, Michael WM, Morandi C, Dreyfuss G. hnRNP I, the polypyrimidine tract-binding protein: distinct nuclear localization and association with hnRNAs. Nucleic Acids Res. 1992;20:3671–3678. doi: 10.1093/nar/20.14.3671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gil A, Sharp PA, Jamison SF, Garcia-Blanco MA. Characterization of cDNAs encoding the polypyrimidine tract-binding protein. Genes Dev. 1991;5:1224–1236. doi: 10.1101/gad.5.7.1224. [DOI] [PubMed] [Google Scholar]
- Giraud S, Greco A, Brink M, Diaz JJ, Delafontaine P. Translation initiation of the insulin-like growth factor I receptor mRNA is mediated by an internal ribosome entry site. J Biol Chem. 2001;276:5668–5675. doi: 10.1074/jbc.M005928200. [DOI] [PubMed] [Google Scholar]
- Gosert R, Chang KH, Rijnbrand R, Yi M, Sangar DV, Lemon SM. Transient expression of cellular polypyrimidine-tract binding protein stimulates cap-independent translation directed by both picornaviral and flaviviral internal ribosome entry sites in vivo. Mol Cell Biol. 2000;20:1583–1595. doi: 10.1128/mcb.20.5.1583-1595.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahm B, Cho OH, Kim JE, Kim YK, Kim JH, Oh YL, Jang SK. Polypyrimidine tract-binding protein interacts with hnRNP L. FEBS Lett. 1998;425:401–406. doi: 10.1016/s0014-5793(98)00269-5. [DOI] [PubMed] [Google Scholar]
- Hellen CU, Pestova TV, Litterst M, Wimmer E. The cellular polypeptide p57 (pyrimidine tract-binding protein) binds to multiple sites in the poliovirus 5′ nontranslated region. J Virol. 1994;68:941–950. doi: 10.1128/jvi.68.2.941-950.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hobert O, Jallal B, Schlessinger J, Ullrich A. Novel signaling pathway suggested by SH3 domain-mediated p95vav/heterogeneous ribonucleoprotein K interaction. J Biol Chem. 1994;269:20225–20228. [PubMed] [Google Scholar]
- Huang S, Deerinck TJ, Ellisman MH, Spector DL. The dynamic organization of the perinucleolar compartment in the cell nucleus, J. Cell Biol. 1997;137:965–974. doi: 10.1083/jcb.137.5.965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hunt SL, Jackson RJ. Polypyrimidine-tract binding protein (PTB) is necessary, but not sufficient, for efficient internal initiation of translation of human rhinovirus-2 RNA. RNA. 1999;5:344–359. doi: 10.1017/s1355838299981414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ito T, Lai MM. An internal polypyrimidine-tract-binding protein-binding site in the hepatitis C virus RNA attenuates translation, which is relieved by the 3′-untranslated sequence. Virology. 1999;254:288–296. doi: 10.1006/viro.1998.9541. [DOI] [PubMed] [Google Scholar]
- Ito T, Tahara SM, Lai MM. The 3′-untranslated region of hepatitis C virus RNA enhances translation from an internal ribosomal entry site. J Virol. 1998;72:8789–8796. doi: 10.1128/jvi.72.11.8789-8796.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izaurralde E, Jarmolowski A, Beisel C, Mattaj IW, Dreyfuss G, Fischer U. A role for the M9 transport signal of hnRNP A1 in mRNA nuclear export. J Cell Biol. 1997;137:27–35. doi: 10.1083/jcb.137.1.27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang SK, Wimmer E. Cap-independent translation of encephalomyocarditis virus RNA: structural elements of the internal ribosomal entry site and involvement of a cellular 57-kD RNA-binding protein. Genes Dev. 1990;4:1560–1572. doi: 10.1101/gad.4.9.1560. [DOI] [PubMed] [Google Scholar]
- Kaminski A, Hunt SL, Patton JG, Jackson RJ. Direct evidence that polypyrimidine tract binding protein (PTB) is essential for internal initiation of translation of encephalomyocarditis virus RNA. RNA. 1995;1:924–938. [PMC free article] [PubMed] [Google Scholar]
- Kim YK, Hahm B, Jang SK. Polypyrimidine tract-binding protein inhibits translation of bip mRNA. J Mol Biol. 2000a;304:119–133. doi: 10.1006/jmbi.2000.4179. [DOI] [PubMed] [Google Scholar]
- Kim JH, Hahm B, Kim YK, Choi M, Jang SK. Protein-protein interaction among hnRNPs shuttling between nucleus and cytoplasm. J Mol Biol. 2000b;298:395–405. doi: 10.1006/jmbi.2000.3687. [DOI] [PubMed] [Google Scholar]
- Kim YK, Jang SK. La protein is required for efficient translation driven by encephalomyocarditis virus internal ribosomal entry site. J Gen Virol. 1999;80:3159–3166. doi: 10.1099/0022-1317-80-12-3159. [DOI] [PubMed] [Google Scholar]
- Krecic AM, Swanson MS. hnRNP complexes: composition, structure, and function. Curr Opin Cell Biol. 1999;11:363–371. doi: 10.1016/S0955-0674(99)80051-9. [DOI] [PubMed] [Google Scholar]
- Kruhlak MJ, Lever MA, Fischle W, Verdin E, Bazett-Jones DP, Hendzel MJ. Reduced mobility of the alternate splicing factor (ASF) through the nucleoplasm and steady state speckle compartments. J Cell Biol. 2000;150:41–51. doi: 10.1083/jcb.150.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lichtenstein M, Guo W, Tartakoff AM. Control of nuclear export of hnRNP A1. Traffic. 2001;2:261–267. doi: 10.1034/j.1600-0854.2001.1o002.x. [DOI] [PubMed] [Google Scholar]
- Lou H, Gagel RF, Berget SM. An intron enhancer recognized by splicing factors activates polyadenylation. Genes Dev. 1996;10:208–219. doi: 10.1101/gad.10.2.208. [DOI] [PubMed] [Google Scholar]
- Lou H, Helfman DM, Gagel RF, Berget SM. Polypyrimidine tract-binding protein positively regulates inclusion of an alternative 3′-terminal exon. Mol Cell Biol. 1999;19:78–85. doi: 10.1128/mcb.19.1.78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lou H, Neugebauer KM, Gagel RF, Berget SM. Regulation of alternative polyadenylation by U1 snRNPs and SRp20. Mol Cell Biol. 1998;18:4977–4985. doi: 10.1128/mcb.18.9.4977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matera AG, Frey MR, Margelot K, Wolin SL. A perinucleolar compartment contains several RNA polymerase III transcripts as well as the polypyrimidine tract-binding protein, hnRNP I. J Cell Biol. 1995;129:1181–1193. doi: 10.1083/jcb.129.5.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michael WM, Choi M, Dreyfuss G. A nuclear export signal in hnRNP A1: a signal-mediated, temperature-dependent nuclear protein export pathway. Cell. 1995a;83:415–422. doi: 10.1016/0092-8674(95)90119-1. [DOI] [PubMed] [Google Scholar]
- Michael WM, Eder PS, Dreyfuss G. The K nuclear shuttling domain: a novel signal for nuclear import and nuclear export in the hnRNP K protein. EMBO J. 1997;16:3587–3598. doi: 10.1093/emboj/16.12.3587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Michael WM, Siomi H, Choi M, Pinol-Roma S, Nakielny S, Liu Q, Dreyfuss G. Signal sequences that target nuclear import and nuclear export of pre-mRNA-binding proteins. Cold Spring Harbor Symp Quant Biol. 1995b;60:663–668. doi: 10.1101/sqb.1995.060.01.071. [DOI] [PubMed] [Google Scholar]
- Moreira A, Takagaki Y, Brackenridge S, Wollerton M, Manley JL, Proudfoot NJ. The upstream sequence element of the C2 complement poly(A) signal activates mRNA 3′ end formation by two distinct mechanisms. Genes Dev. 1998;12:2522–2534. doi: 10.1101/gad.12.16.2522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakielny S, Dreyfuss G. Transport of proteins and RNAs in and out of the nucleus. Cell. 1999;99:677–690. doi: 10.1016/s0092-8674(00)81666-9. [DOI] [PubMed] [Google Scholar]
- Nguyen VT, Giannoni F, Dubois MF, Seo SJ, Vigneron M, Kedinger C, Bensaude O. In vivo degradation of RNA polymerase II largest subunit triggered by alpha-amanitin. Nucleic Acids Res. 1996;24:2924–2929. doi: 10.1093/nar/24.15.2924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nigg EA. Nucleocytoplasmic transport: signals, mechanisms and regulation. Nature. 1997;386:779–787. doi: 10.1038/386779a0. [DOI] [PubMed] [Google Scholar]
- Oh YL, Hahm B, Kim YK, Lee HK, Lee JW, Song O, Tsukiyama-Kohara K, Kohara M, Nomoto A, Jang SK. Determination of functional domains in polypyrimidine-tract-binding protein. Biochem J. 1998;331:169–175. doi: 10.1042/bj3310169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ostareck-Lederer A, Ostareck DH, Hentze MW. Cytoplasmic regulatory functions of the KH-domain proteins hnRNPs K and E1/E2. Trends Biochem Sci. 1998;23:409–411. doi: 10.1016/s0968-0004(98)01301-2. [DOI] [PubMed] [Google Scholar]
- Patton JG, Mayer SA, Tempst P, Nadal-Ginard B. Characterization and molecular cloning of polypyrimidine tract-binding protein: a component of a complex necessary for pre-mRNA splicing. Genes Dev. 1991;5:1237–1251. doi: 10.1101/gad.5.7.1237. [DOI] [PubMed] [Google Scholar]
- Perez I, McAfee JG, Patton JG. Multiple RRMs contribute to RNA binding specificity and affinity for polypyrimidine tract binding protein. Biochemistry. 1997;36:11881–11890. doi: 10.1021/bi9711745. [DOI] [PubMed] [Google Scholar]
- Perry R. Selective effects of actinomycin D on the intracellular distribution of RNA synthesis in tissue culture cells. Exp Cell Res. 1963;29:400–406. [Google Scholar]
- Phair RD, Misteli T. High mobility of proteins in the mammalian cell nucleus. Nature. 2000;404:604–609. doi: 10.1038/35007077. [DOI] [PubMed] [Google Scholar]
- Pinol-Roma S, Dreyfuss G. Transcription-dependent and transcription-independent nuclear transport of hnRNP proteins. Science. 1991;253:312–314. doi: 10.1126/science.1857966. [DOI] [PubMed] [Google Scholar]
- Pinol-Roma S, Dreyfuss G. Shuttling of pre-mRNA binding proteins between nucleus and cytoplasm. Nature. 1992;355:730–732. doi: 10.1038/355730a0. [DOI] [PubMed] [Google Scholar]
- Romanelli MG, Weighardt F, Biamonti G, Riva S, Morandi C. Sequence determinants for hnRNP I protein nuclear localization. Exp Cell Res. 1997;235:300–304. doi: 10.1006/excr.1997.3677. [DOI] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 1989. [Google Scholar]
- Sehgal PB, Darnell JE, Jr, Tamm I. The inhibition by DRB (5,6-dichloro-1-beta-D-ribofuranosylbenzimidazole) of hnRNA and mRNA production in HeLa cells. Cell. 1976;9:473–480. doi: 10.1016/0092-8674(76)90092-1. [DOI] [PubMed] [Google Scholar]
- Swanson MS, Dreyfuss G. Classification and purification of proteins of heterogeneous nuclear ribonucleoprotein particles by RNA-binding specificities. Mol Cell Biol. 1988;8:2237–2241. doi: 10.1128/mcb.8.5.2237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talcott B, Moore MS. Getting across the nuclear pore complex. Trends Cell Biol. 1999;9:312–318. doi: 10.1016/s0962-8924(99)01608-6. [DOI] [PubMed] [Google Scholar]
- Tomonaga T, Levens D. Heterogeneous nuclear ribonucleoprotein K is a DNA-binding transactivator. J Biol Chem. 1995;270:4875–4881. doi: 10.1074/jbc.270.9.4875. [DOI] [PubMed] [Google Scholar]
- Tomonaga T, Levens D. Activating transcription from single stranded DNA. Proc Natl Acad Sci USA. 1996;93:5830–5835. doi: 10.1073/pnas.93.12.5830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vautier D, Chesne P, Cunha C, Calado A, Renard JP, Carmo-Fonseca M. Transcription-dependent nucleocytoplasmic distribution of hnRNP A1 protein in early mouse embryos. J Cell Sci. 2001;114:1521–1531. doi: 10.1242/jcs.114.8.1521. [DOI] [PubMed] [Google Scholar]
- Visa N, Alzhanova-Ericsson AT, Sun X, Kiseleva E, Bjorkroth B, Wurtz T, Daneholt B. A pre-mRNA-binding protein accompanies the RNA from the gene through the nuclear pores and into polysomes. Cell. 1996;84:253–264. doi: 10.1016/s0092-8674(00)80980-0. [DOI] [PubMed] [Google Scholar]
- Wagner EJ, Garcia-Blanco MA. Polypyrimidine tract binding protein antagonizes exon definition. Mol Cell Biol. 2001;21:3281–3288. doi: 10.1128/MCB.21.10.3281-3288.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- White J, Stelzer E. Photobleaching GFP reveals protein dynamics inside live cells. Trends Cell Biol. 1999;9:61–65. doi: 10.1016/s0962-8924(98)01433-0. [DOI] [PubMed] [Google Scholar]
- Witherell GW, Wimmer E. Encephalomyocarditis virus internal ribosomal entry site RNA-protein interactions. J Virol. 1994;68:3183–3192. doi: 10.1128/jvi.68.5.3183-3192.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]