

Abstract
Chicken erythroid ankyrin undergoes a fairly rapid cycle of cytoskeletal association, dissociation, and turnover. In addition, the cytoskeletal association of ankyrin is regulated by phosphorylation. Treatment of erythroid cells with serine and threonine phosphatase inhibitors stimulated the hyperphosphorylation of the 225- and 205-kDa ankyrin isoforms, and dissociated the bulk of these isoforms from cytoskeletal spectrin. In vitro binding studies have shown that this dissociation of ankyrin from spectrin in vivo can be attributed to a reduced ability of hyperphosphorylated ankyrin to bind spectrin. Interestingly, a significant fraction of detergent insoluble ankyrin accumulates in a spectrin-independent pool. At least some of this spectrin-independent pool of ankyrin is complexed with the AE1 anion exchanger, and the solubility properties of this pool are also regulated by phosphorylation. Treatment of cells with serine and threonine phosphatase inhibitors had no effect on ankyrin/AE1 complex formation. However, these inhibitors were sufficient to shift ankyrin/AE1 complexes from the detergent insoluble to the soluble pool. These analyses, which are the first to document the in vivo consequences of ankyrin phosphorylation, indicate that erythroid ankyrin-containing complexes can undergo dynamic rearrangements in response to changes in phosphorylation.
INTRODUCTION
Ankyrins constitute a family of adapter proteins that mediate interactions of multiple plasma membrane proteins with the spectrin-based cytoskeleton (Bennett, 1992). This family of proteins associates with a variety of membrane compartments in cells (Bennett and Stenbuck, 1979; Devarajan et al., 1996; Zhou et al., 1997) and has been implicated in the stabilization (Hammerton et al., 1991) and trafficking (Devarajan et al., 1997; Zhou et al., 1998) of membrane proteins. A well-characterized interaction of erythroid ankyrin is its association with the N-terminal cytoplasmic domain of the AE1 anion exchanger (Bennett and Stenbuck, 1980). This interaction provides a link between the plasma membrane and the submembranous spectrin scaffold in red blood cells. The critical role of this member of the ankyrin family in maintaining erythroid cell shape and stability is illustrated by the fact that deficiencies in the human Ank 1 gene are often observed in patients with hereditary spherocytosis (Tse and Lux, 1999).
Although ankyrin family members are involved in diverse processes associated with membrane protein trafficking and localization, the composition of ankyrin-containing complexes within cells and the biochemical mechanisms that regulate their formation are not well understood. In this study, we attempted to characterize the precise nature of erythroid ankyrin-containing complexes, and we investigated whether the properties of these complexes are regulated by phosphorylation. Other investigators have shown that the phosphorylation of human erythroid ankyrin in vitro reduces its affinity for both the AE1 anion exchanger and spectrin (Cianci et al., 1988). However, a role for phosphorylation in regulating the association of ankyrin with AE1 and spectrin in vivo has not been established.
For these studies, chicken embryonic erythroid cells were used as a model system. Although these embryonic red cells retain many properties that are similar to mammalian erythroid cells, the presence of a nucleus and other membrane organelles in these cells makes this a very useful model for studying the role of ankyrin in general cellular processes. Our analyses revealed that detergent-insoluble ankyrin polypeptides accumulate in spectrin-dependent and spectrin-independent pools, and some of the spectrin-independent pool of ankyrin exists in a complex with AE1. Treatment of erythroid cells with serine and threonine phosphatase inhibitors stimulates the hyperphosphorylation of the 225- and 205-kDa chicken ankyrin isoforms. In addition, treatment of cells with phosphatase inhibitors shifts both the spectrin-dependent and spectrin-independent pools of detergent-insoluble ankyrin to the soluble pool. In vitro binding studies have shown that the dissociation of ankyrin from the spectrin cytoskeleton in cells treated with phosphatase inhibitors is at least in part due to a reduced ability of hyperphosphorylated ankyrin to bind spectrin. These results are consistent with a role for chicken erythroid ankyrin as a multifunctional adapter, whose interactions are dynamically regulated in response to changes in phosphorylation.
MATERIALS AND METHODS
Immunoprecipitation
Erythroid cells from 10-d-old chicken embryos were incubated in methionine-free DMEM containing 0.25mCi/ml 35S-Translabel (ICN, Costa Mesa, CA) at 37°C for various times. After labeling, the cells were lysed by incubation in hypotonic lysis buffer (5 mM Tris-HCl pH 7.5, 5 mM MgCl2, 1 mM dithiothreitol, 1 mM EGTA). Alternatively, the cells were chased in DMEM containing 10% fetal calf serum before hypotonic lysis. The hypotonic lysate was centrifuged for 5 min in an Eppendorf centrifuge. After centrifugation, the hypotonic soluble fraction was collected. The hypotonic pellet was resuspended in isotonic buffer (150 mM NaCl, 10 mM Tris pH 7.5, 5 mM MgCl2, 2 mM EGTA, 6 mM β-mercaptoethanol) containing 1% Triton X-100 and separated into detergent-soluble and -insoluble fractions as described previously (Ghosh et al., 1999). In some instances, cells were directly lysed in isotonic buffer containing 1% Triton X-100, or in a low salt buffer (10 mM NaCl, 10 mM Tris pH 7.5, 5 mM MgCl2, 2 mM EGTA, 6 mM β-mercaptoethanol) containing 1% Triton X-100 and separated into soluble and insoluble fractions by centrifugation. The insoluble pellets were sonicated in the same buffer used for lysis, and insoluble material was pelleted by centrifugation and discarded. Immunoblotting analysis has shown that the discarded material is devoid of AE1, ankyrin, and α-spectrin (our unpublished data). Protein A-agarose beads preloaded with AE1-specific (Cox et al., 1995) or ankyrin-specific (Ghosh et al., 1999) peptide antibodies, or with monoclonal antibodies (mAbs) specific for α-spectrin (ICN) were added to the various fractions and incubated overnight at 4°C. The protein A beads were washed with 20 mM Tris pH7.5, 1% (wt/vol) NaCl, 5 mM EGTA, 5 mM EDTA, 0.1% (wt/vol) SDS, 1% (vol/vol) Triton X-100, and 1% (wt/vol) sodium deoxycholate (immunoprecipitation buffer) or with detergent lysis buffer, and the immune complexes were released by incubation in SDS sample buffer. Precipitates were analyzed on a 6% SDS polyacrylamide gel. The gels were stained with GelCode Blue (Pierce Chemical, Rockford, IL), and subsequently fixed and treated with 20% 2,5-diphenyloxazole in dimethyl sulfoxide. The gels were then dried and exposed to Kodak Biomax MR film. The immunoprecipitation profiles observed when the precipitates were washed with immunoprecipitation buffer or with isotonic lysis buffer were the same.
[32P]Orthophosphate Labeling of Chicken Erythroid Cells
Erythroid cells isolated from 10-d-old chicken embryos were washed with phosphate-free DMEM. The cells were then resuspended in phosphate-free DMEM containing 1mCi/ml [32P]orthophosphate and incubated at 37°C for 4 h in the presence or absence of phosphatase inhibitors. At this time, the cells were detergent fractionated and immunoprecipitates were prepared and analyzed on a 6% SDS polyacrylamide gel. In some instances, the precipitates were digested with 20 U of calf intestine alkaline phosphatase (BRL, Rockville, MD) at 37°C for 45 min before gel analysis. The gels were dried and exposed to Kodak Biomax MS film.
One-Dimensional Peptide Mapping
Erythroid cells isolated from 10-d-old chicken embryos were incubated in methionine-free DMEM containing 0.25mCi/ml 35S-Translabel for 4 h. At this time, the cells were lysed in isotonic lysis buffer containing 1% Triton X-100 and fractionated. Ankyrin immunoprecipitates prepared from the detergent-insoluble fraction were electrophoresed on a 6% SDS polyacrylamide gel. The gel was stained with GelCode Blue, and regions of the gel corresponding to the 225-, 220-, and 205-kDa polypeptides were excised. The proteins were electroeluted from the gel slices in a buffer composed of 125 mM Tris-HCl pH 6.8, 0.1% SDS, 1.0 mM EDTA, and 30 mM dithiothreitol. The eluted proteins were digested with 10 ng of Staphylococcus aureus strain V8 endoproteinase at 37°C for 30 min (Cleveland et al., 1977). The addition of fivefold more V8 protease to these reactions did not alter the digestion profiles. The digested protein was analyzed on a 15% SDS polyacrylamide gel. After electrophoresis, the gel was fixed and treated with 20% 2,5-diphenyloxazole in dimethyl sulfoxide, dried, and exposed to Kodak Biomax MR film.
Immunoblotting Analysis
Erythroid cells from 10-d-old chicken embryos were incubated in the absence or presence of 1 μM okadaic acid for 2 h before detergent fractionation in isotonic buffer. The detergent-soluble and -insoluble fractions were electrophoresed on a 6% SDS polyacrylamide gel, and electrophoretically transferred to nitrocellulose. The filter was incubated with a 1:3000 dilution of the ankyrin-specific antibody in Tris-buffered saline containing 0.25% gelatin. The filter was then washed in Tris-buffered saline containing 0.05% Tween 20, and incubated with goat anti-rabbit (GAR) IgG conjugated to horseradish peroxidase. After washing, immunoreactive species were detected by enhanced chemiluminescence. Similar immunoblotting analyses were carried out with chicken AE1-specific antibodies at a 1:20,000 dilution.
In some instances, erythroid cells that had been incubated in the absence or presence of 100 nM calyculin A for 2 h were lysed in low salt buffer containing 1% Triton X-100. Soluble and insoluble fractions from these cells were processed for immunoblotting with a 1:500 dilution of an α-spectrin-specific mAb (ICN), a 1:2500 dilution of an affinity-purified rabbit antibody specific for β-spectrin (Zhou et al., 1998), or a 1:3000 dilution of the ankyrin-specific antibody. Alternatively, immunoprecipitates prepared from the soluble and insoluble fractions with the α-spectrin monoclonal or AE1-specific antibodies were subjected to immunoblotting analysis with ankyrin-specific antibodies.
In Vitro Binding Assay
An immunoprecipitate was prepared with the α-spectrin mAb from the detergent-insoluble fraction of erythroid cells that were lysed in isotonic buffer containing 1% Triton X-100. Analysis of α-spectrin immunoprecipitates prepared from 35S-labeled cells under these conditions revealed that β-spectrin coprecipitates with α-spectrin in a stoichiometric complex (our unpublished data). The α-spectrin immunoprecipitate was washed into low salt buffer containing 1% Triton X-100 and incubated overnight at 4°C with ankyrin that was immunopurified from untreated or calyculin A-treated erythroid cells as described below. After extensive washing in low salt buffer, the α-spectrin immunoprecipitates were processed for immunoblotting analysis with ankyrin-specific antibodies.
Erythroid cells that had been incubated in the absence or presence of 100 nM calyculin A for 2 h were lysed in isotonic buffer containing 1% Triton X-100. These cells were separated into soluble and insoluble fractions by centrifugation, and ankyrin antibodies directly conjugated to CNBr-activated Sepharose 4B beads were used to immunoprecipitate ankyrin from the detergent-insoluble fraction. After washing in isotonic buffer containing 1% Triton X-100, immunoprecipitated ankyrin was eluted from the beads in 0.2 M glycine pH 2.3. The eluted ankyrin was dialyzed against low salt buffer containing 1% Triton X-100, and subsequently incubated with an α-spectrin immunoprecipitate in a total volume of 500 μl.
Quantitative Densitometry
X-ray films were scanned with the use of DeskScan II 2.2 software (Hewlett Packard, Palo Alto, CA), and quantitative densitometry was performed with NIH Image. Each value presented in the text represents the average from at least three independent experiments. The relative incorporation of 32P into the ankyrin isoforms was quantitated by phosphoimager analysis with the use of ImageQuant software (Molecular Dynamics, Sunnyvale, CA).
Immunolocalization Analysis
Erythroid cells isolated from 10-d-old chicken embryos were fixed by incubating the cells in 1% paraformaldehyde in phosphate-buffered saline (PBS) for 15 min. After fixation, the cells were permeabilized with acetone. The cells were washed with PBS and incubated with a 1:100 dilution of the α-spectrin-specific mAb in PBS. The cells were then washed and incubated with donkey anti-mouse (DAM) IgG conjugated to fluorescein isothiocyanate (FITC). Alternatively, the cells were incubated with a 1:200 dilution of the affinity-purified β-spectrin-specific antibody followed by donkey anti-rabbit (DAR) IgG conjugated to lissamine. After washing, immunoreactive polypeptides were visualized with the use of a Zeiss Axiophot microscope.
The cellular distribution of detergent insoluble ankyrin, α-spectrin, and β-spectrin was examined by extracting erythroid cells from 10-d-old embryos with ice-cold low salt buffer containing 1% Triton X-100 for 30 s. The preextracted cells were fixed by incubation in 1% paraformaldehyde in PBS for 15 min, and subsequently incubated with a 1:100 dilution of the α-spectrin monoclonal and a 1:1000 dilution of the rabbit polyclonal antibody specific for ankyrin. After washing, the cells were incubated with DAM-IgG conjugated to FITC and DAR-IgG conjugated to lissamine. Alternatively, fixed cells were incubated with a 1:200 dilution of the affinity-purified β-spectrin-specific antibody followed by GAR-IgG Fab fragments conjugated to tetramethylrhodamine isothiocyanate. The cells were then washed and incubated with the ankyrin-specific antibody that had been directly conjugated to Alexa 488 (Molecular Probes, Eugene, OR) according to manufacturer's instructions. After washing, immunoreactive polypeptides were visualized with the use of a Zeiss Axiophot microscope.
RESULTS
Properties of Newly Synthesized Chicken Erythroid Ankyrin
Pulse-chase analyses have investigated the mechanisms involved in the assembly of ankyrin-containing complexes in chicken embryonic erythroid cells. Erythroid cells isolated from 10-d-old chicken embryos were pulsed for 2 min with 35S-Translabel and chased for 5 min. At this time, the cells were lysed in hypotonic lysis buffer, and separated into soluble and insoluble fractions by centrifugation. The soluble fraction was saved, and the insoluble pellet was resuspended in isotonic buffer containing 1% Triton X-100 and separated into detergent-soluble and -insoluble fractions. Immunoprecipitates were prepared from each fraction with an ankyrin-specific peptide antibody. Immunoblotting analysis with this antibody has detected polypeptides of 225- and 205-kDa from 10-d-old embryonic erythroid cells that are almost entirely detergent insoluble (Ghosh et al., 1999). This analysis revealed that the newly synthesized 225-kDa ankyrin isoform was primarily present in the soluble fractions at the end of the 5-min chase, whereas the newly synthesized 205-kDa ankyrin isoform was faintly detected in the detergent-insoluble fraction (Figure 1B, lanes 1–3). In contrast to the profile observed for the newly synthesized polypeptides, Coomassie Blue staining of the precipitates revealed that the 225- and 205-kDa ankyrin isoforms could only be detected in the detergent-insoluble fraction (Figure 1A, lane 3). A Coomassie-stained polypeptide of 220 kDa also coprecipitated with the detergent-insoluble 225- and 205-kDa ankyrin isoforms (Figure 1A, lane 3). None of these polypeptides was observed in immunoprecipitates prepared in the presence of the peptide the antibody was raised against (Figure 1, A and B, lanes 4–6).
Figure 1.

Acquisition of detergent insolubility by newly synthesized ankyrin. Chicken erythroid cells were pulsed with 35S-Translabel in methionine-free DMEM for 2 min, and chased for 5 min in DMEM containing 10% fetal calf serum. At this time, cells were lysed in hypotonic lysis buffer, and the lysate was centrifuged for 5 min. After centrifugation, the hypotonic soluble (H) fraction was collected, and the pellet was resuspended in isotonic buffer containing 1% Triton X-100 and separated into detergent-soluble (S) and -insoluble (I) fractions by centrifugation. Immunoprecipitates were prepared from each fraction with ankyrin-specific peptide antibodies in the absence (A and B, lanes 1–3) or presence (A and B, lanes 4–6) of 25 μg of the peptide antigen. Immune complexes were washed with immunoprecipitation buffer and analyzed on a 6% SDS-polyacrylamide gel. The steady-state distribution of ankyrin was determined by staining the gel with GelCode Blue (A), whereas labeled ankyrin was detected by fluorography (B). Alternatively, erythroid cells were pulsed for 15 min (C, lanes 1–3), and chased for 1 (C, lanes 4–6) or 4 (C, lanes 7–9) h. At each time point, cells were fractionated and ankyrin immunoprecipitates were prepared and analyzed as described above. In similar pulse-chase analyses, cells were directly lysed by incubation in isotonic buffer containing 1% Triton X-100. The lysate was separated into soluble and insoluble fractions, and AE1 immunoprecipitates were prepared (D). The region of the gel containing coimmunoprecipitating ankyrin polypeptides is shown in each panel. Dashes adjacent to each panel mark the 225-, 220-, and 205-kDa polypeptides detected in these analyses.
Similar analyses with erythroid cells that were pulsed for 15 min and chased for 1 or 4 h revealed that the 225- and 205-kDa ankyrin isoforms were detectable in all fractions at the end of a 15-min pulse (Figure 1C, lanes 1–3). Furthermore, a faint labeled species of 220 kDa was observed in the detergent-insoluble ankyrin precipitate after a 15-min labeling (Figure 1C, lane 3). During the chase, there was a dramatic reduction in the soluble forms of ankyrin. The depletion of ankyrin from the soluble pools correlated with an increase in ankyrin in the detergent-insoluble fraction (Figure 1C, lanes 6 and 9). In addition, the prevalence of the 220-kDa polypeptide in the detergent-insoluble ankyrin precipitate increased during the chase (Figure 1C). These data indicate that chicken erythroid ankyrin is primarily synthesized as a soluble polypeptide. Quantitation of multiple experiments has revealed that 27.4 ± 6.2% (n = 4) of the soluble 225-kDa polypeptides associate with the detergent-insoluble fraction during the course of the chase. The remainder of the newly synthesized 225-kDa polypeptides turns over.
One-dimensional peptide mapping analysis investigated whether the 220-kDa polypeptide that coprecipitated with detergent-insoluble ankyrin corresponds to a form of ankyrin not recognized by the peptide antibody, or to a distinct component of the cytoskeleton. Ankyrin precipitates were prepared from the detergent-insoluble fraction of 35S-labeled erythroid cells. After SDS gel electrophoresis, the 225- and 205-kDa ankyrin isoforms and the 220-kDa polypeptide were excised from the gel and subjected to proteolytic digestion with V8 protease. This analysis revealed virtually identical peptide maps for each polypeptide (Figure 2). The peptides marked with asterisks are the only ones that differ among the three polypeptides. These results strongly suggest that the 225-, 220-, and 205-kDa polypeptides are derived from the same gene.
Figure 2.
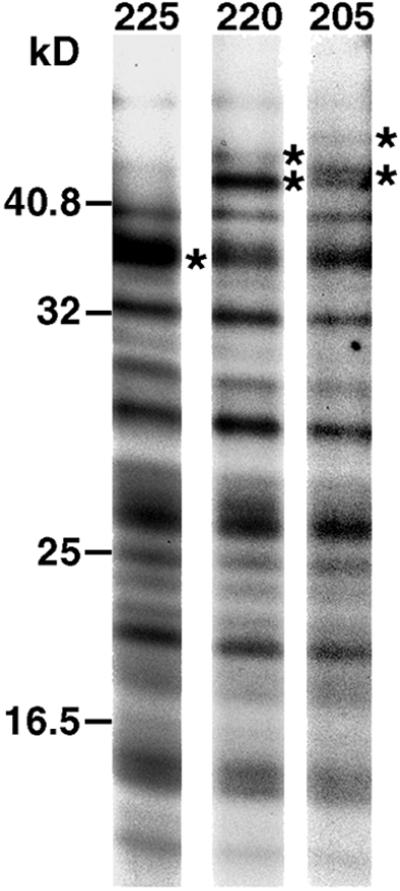
One-dimensional peptide mapping analysis of polypeptides in the ankyrin-containing complex. Erythroid cells were metabolically labeled with 35S-Translabel for 4 h. At this time, cells were detergent fractionated, and an ankyrin immunoprecipitate was prepared from the detergent-insoluble fraction. The immune complex was washed with immunoprecipitation buffer and electrophoresed on a 6% SDS polyacrylamide gel, which was stained with GelCode Blue. Regions of the gel corresponding to the 225-, 220-, and 205-kDa polypeptides were excised. The polypeptides were electroeluted and digested with S. aureus strain V8 endoproteinase. The resulting peptides were analyzed on a 15% SDS polyacrylamide gel and detected by fluorography. Peptides that differ between the 225-, 220-, and 205-kDa polypeptides are marked with asterisks. Molecular weight markers are indicated to the left of the figure.
It was possible that the 220-kDa polypeptide detected in these assays arose by postlysis processing of the 225-kDa polypeptide by a protease exclusively present in the detergent-insoluble fraction of erythroid cells. To address this possibility, an ankyrin immunoprecipitate prepared from the detergent-soluble fraction of 35S-labeled cells was mixed with the detergent-insoluble fraction prepared from unlabeled cells. After SDS gel analysis, the only labeled polypeptides detected in the immunoprecipitate were the 205- and 225-kDa isoforms (our unpublished data). This suggests the 220-kDa polypeptide does not arise by the postlysis proteolysis of the 225-kDa polypeptide. The precise mechanism(s) involved in generating the 205- and 220-kDa polypeptides remains to be elucidated.
Chicken Erythroid Ankyrin Exists in AE1-containing and AE1-deficient Complexes
Previous studies have shown that newly synthesized chicken AE1 anion exchangers associate with the detergent-insoluble fraction of erythroid cells during recycling to the Golgi. This acquisition of insolubility by AE1 correlated with its association with detergent-insoluble ankyrin (Ghosh and Cox, 1999). To determine whether the entire cellular pool of ankyrin exists in this AE1-containing complex, erythroid cells isolated from 10-d-old embryos were pulsed with 35S-Translabel for 15 min and chased for 4 h. At this time, the cells were detergent lysed and immunoprecipitates were prepared from the detergent-soluble and -insoluble fractions with AE1 or ankyrin-specific antibodies (Figure 3A, lanes 1–4). After immunoprecipitation, the supernatant from the AE1 precipitate was split in half and reprecipitated with AE1 (Figure 3A, lanes 5 and 6) or ankyrin-specific (Figure 3A, lanes 7 and 8) antibodies. This analysis revealed that the 205- and 225-kDa ankyrin isoforms coprecipitated with detergent-soluble and -insoluble forms of AE1 in the initial round of precipitation. In addition, the 220-kDa isoform coprecipitated with detergent-insoluble AE1 (Figure 3A, lanes 1 and 2). Although ankyrin was essentially absent in the reprecipitation with AE1 antibodies (Figure 3A, lanes 5 and 6), each isoform was detected in the ankyrin reprecipitate from the insoluble fraction, and the 225-kDa isoform was faintly detected in the ankyrin reprecipitate from the soluble fraction (Figure 3A, lanes 7 and 8). Quantitation of multiple immunodepletion experiments has indicated that 46.5 ± 1.9% (n = 3) of detergent-insoluble ankyrin exists in a stable complex with AE1. It is possible that the remaining population of detergent-insoluble ankyrin is dissociated from AE1 during cell lysis and immunoprecipitation. However, this seems unlikely because detergent-soluble ankyrin almost quantitatively coprecipitates with AE1 under the same conditions (Figure 3A, lane 1). Regardless of the origin of the AE1-free pool of ankyrin, these data indicate that two separate populations of detergent-insoluble ankyrin are present in these embryonic erythroid cells, only one of which is tightly associated with AE1. Although previous studies have shown that ∼50% of AE1 is detergent insoluble (Ghosh and Cox, 1999), similar coprecipitation experiments have demonstrated that only 11.7 ± 0.7% (n = 4) of detergent-insoluble AE1 exists in a complex with ankyrin (Figure 3B).
Figure 3.

Analysis of ankyrin and AE1-containing complexes in chicken erythroid cells. Erythroid cells were pulsed with 35S-Translabel for 15 min and chased for 4 h. After detergent lysis, immunoprecipitates were prepared from the detergent-soluble and -insoluble fractions with AE1-specific (A, lanes 1 and 2) or ankyrin-specific (A, lanes 3 and 4) antibodies. The supernatants from the AE1 precipitates were split in half and reprecipitated with AE1 (A, lanes 5 and 6) or ankyrin (A, lanes 7 and 8) antibodies. Additional AE1 (B, lane 1) and ankyrin (B, lane 2) immunoprecipitates were prepared from the detergent-insoluble fraction of cells labeled as described above. The supernatant from this ankyrin immunoprecipitate was split in half and reprecipitated with ankyrin (B, lane 3) or AE1 (B, lane 4) antibodies. Immune complexes from these analyses were electrophoresed on 6% SDS polyacrylamide gels and labeled proteins were detected by fluorography. A only shows the portion of the gel containing labeled ankyrin polypeptides, whereas B only shows the portion of the gel containing labeled AE1. Dashes to the left of A indicate the 225-, 220-, and 205-kDa ankyrin isoforms.
Detergent-insoluble Ankyrin Is Unstable
The pulse-chase analysis in Figure 1C revealed that the prevalence of the 225- and 205-kDa ankyrin isoforms in the detergent-insoluble fraction peaked at the 1-h chase point and declined by 4 h of chase. Quantitation of multiple experiments of this type has indicated that there is a 45.6 ± 9.3% (n = 4) reduction in insoluble ankyrin by 4–6 h of chase. A similar result was observed for the population of ankyrin that coprecipitated with detergent-insoluble AE1 (Figure 1D). After dissociation from the insoluble pool, neither total ankyrin (Figure 1C) nor AE1-associated ankyrin (Figure 1D) accumulate in the soluble pool; rather these polypeptides turn over. In contrast to ankyrin, both the detergent-soluble and detergent-insoluble forms of AE1 are stable in similar pulse-chase analyses (Ghosh and Cox, 1999).
Effect of Serine and Threonine Phosphatase Inhibitors on Fractionation Properties of Chicken Erythroid Ankyrin and AE1
Other investigators have shown that turkey red cell ankyrin is phosphorylated in vivo (Alper et al., 1980). To investigate whether the phosphorylation status of chicken erythroid ankyrin is involved in regulating its cytoskeletal association and/or its turnover in vivo, erythroid cells from 10-d-old embryos were incubated in the absence or presence of okadaic acid, a serine and threonine phosphatase inhibitor. After detergent lysis of the cells in isotonic buffer, the detergent-soluble and -insoluble fractions were subjected to immunoblotting analysis with ankyrin or AE1-specific antibodies. This analysis revealed that okadaic acid treatment shifted the solubility properties of ankyrin (Figure 4). Quantitation of multiple experiments revealed that ankyrin shifted from 87.4 ± 4.4% (n = 4) detergent insoluble in untreated cells to 21.4 ± 3.8% (n = 4) detergent insoluble in okadaic acid-treated cells. In addition to its effect on the detergent solubility properties of ankyrin, okadaic acid treatment resulted in an altered mobility for ankyrin on SDS gels (Figure 4). In contrast, okadaic acid did not affect the SDS gel mobility or the detergent solubility properties of AE1 (Figure 4). Similar experiments revealed that the serine and threonine phosphatase inhibitor calyculin A had an identical effect on the detergent solubility properties and SDS-gel mobility of erythroid ankyrin (Figure 5B).
Figure 4.

Effect of okadaic acid on the detergent solubility properties of ankyrin. Chicken erythroid cells were incubated in complete media for 2 h at 37°C in the absence (lanes 1 and 2) or presence (lanes 3 and 4) of 1 μ M okadaic acid. The cells were then detergent lysed and separated into soluble (S, lanes 1 and 3) and insoluble (I, lanes 2 and 4) fractions. Equivalent amounts of the resulting fractions were electrophoresed on a 6% SDS polyacrylamide gel, and transferred to nitrocellulose. The filters were incubated with ankyrin or AE1-specific antibodies, followed by GAR-IgG conjugated to horseradish peroxidase. Immunoreactive species were detected by enhanced chemiluminescence.
Figure 5.

Ankyrin is hyperphosphorylated in cells treated with okadaic acid. Erythroid cells were washed in phosphate-free DMEM. The cells were then incubated in phosphate-free DMEM containing 1mCi/ml [32P]orthophosphate for 4 h in the absence (A, lanes 1–4) or presence (A, lanes 5–8) of 1 μM okadaic acid. After detergent lysis, immunoprecipitates were prepared from the detergent-soluble (S) and -insoluble (I) fractions with ankyrin (A, lanes 1, 2, 5, and 6) or AE1-specific (A, lanes 3, 4, 7, and 8) antibodies. Immune complexes were washed with immunoprecipitation buffer and analyzed on a 6% SDS polyacrylamide gel, and 32P-labeled polypeptides were detected by autoradiography. Erythroid cells were also labeled for 4 h with [32P]orthophosphate in the absence (B, lanes 1 and 2) or presence (B, lanes 3–6) of 100 nM calyculin A. Ankyrin immunoprecipitates were prepared from the detergent-soluble and -insoluble fractions and analyzed on a 6% SDS polyacrylamide gel. Some of the immunoprecipitates (B, lanes 3 and 4) were digested with calf-intestinal alkaline phosphatase before electrophoresis. Immunoprecipitated polypeptides were visualized by staining with GelCode Blue and 32P-labeled species were detected by autoradiography. Dashes to the left of each panel indicate the 225-, 220-, and 205-kDa ankyrin isoforms. Dashes to the right of B indicate the hyperphosphorylated 225- (225*) and 205-kDa (205*) ankyrin isoforms.
To determine whether the effects of these phosphatase inhibitors on ankyrin may be due to increased levels of ankyrin phosphorylation, ankyrin immunoprecipitates were prepared from erythroid cells labeled with [32P]orthophosphate in the presence or absence of okadaic acid. This analysis revealed that phosphorylated forms of the 225- and 205-kDa ankyrin isoforms could be detected in both the detergent-soluble and -insoluble fractions of untreated cells (Figure 5A, lanes 1 and 2). This basal level of ankyrin phosphorylation was stimulated in the presence of okadaic acid (Figure 5A, lanes 5 and 6). Quantitation of several experiments revealed that okadaic acid treatment stimulated approximately a 12-fold increase in the phosphorylation of the 225-kDa ankyrin isoform. In addition, phosphoimager analysis of the 32P-labeled ankyrin from okadaic acid-treated cells revealed the 225-kDa ankyrin isoform contained ∼15-fold more counts than the 205-kDa isoform. Because immunoblotting analysis has indicated that the 225-kDa isoform is ∼15-fold more abundant than the 205-kDa isoform (Ghosh and Cox, 1999), these data suggest that each isoform is phosphorylated to the same extent in the presence of okadaic acid. Interestingly, there was no detectable 32P incorporation into the 220-kDa ankyrin isoform in either untreated or okadaic acid-treated cells.
Additional studies investigated the basis for the altered SDS-gel mobility of ankyrin immunoprecipitated from cells treated with phosphatase inhibitors. The 32P-labeled ankyrin precipitates prepared from calyculin A-treated cells were incubated in the presence (Figure 5B, lanes 3 and 4) or absence (Figure 5B, lanes 5 and 6) of alkaline phosphatase before SDS-gel analysis. As shown in Figure 5B, alkaline phosphatase treatment of the precipitate almost entirely removed the labeled phosphate from ankyrin. In addition, the migration of both the 205- and 225-kDa isoforms after alkaline phosphatase digestion was similar to that seen for ankyrin from untreated cells (Figure 5B, lanes 1 and 2). These results suggest that the altered migration of ankyrin isolated from cells treated with phosphatase inhibitors is primarily due to the increased level of phosphorylation of these polypeptides.
AE1 immunoprecipitates prepared from 32P-labeled cells revealed that both detergent-soluble and -insoluble AE1 were phosphorylated. However, the extent of AE1 phosphorylation was not stimulated by okadaic acid treatment (Figure 5A). A subset of phosphorylated ankyrin coprecipitated with detergent-soluble and -insoluble AE1 in both untreated and okadaic acid-treated cells (Figure 5A). Although okadaic acid treatment had no significant effect on the solubility properties of AE1 (Figure 4), treatment of cells with this reagent shifted the bulk of phosphorylated ankyrin that coprecipitated with AE1 from the insoluble to the soluble pool (Figure 5A). This result is consistent with the coprecipitation results that indicated a small percentage of AE1 insolubility is dependent upon its association with ankyrin.
Serine and Threonine Phosphatase Inhibitors Stimulate Dissociation of Ankyrin from Detergent-insoluble Pool
To examine whether okadaic acid stimulated the dissociation of ankyrin from the detergent-insoluble cytoskeleton, erythroid cells were pulsed for 15 min and chased for 4 h. At this time, an aliquot of cells was detergent lysed and ankyrin precipitates were prepared from the detergent-soluble and -insoluble fractions (Figure 6, lanes 1 and 2). The remainder of the cells was chased an additional 2 h in the absence or presence of okadaic acid and processed for immunoprecipitation. Consistent with the observation that detergent-insoluble ankyrin is unstable (Figure 1C), there was a slight reduction in the amount of insoluble ankyrin between 4 and 6 h of chase in control cells (Figure 6, compare lanes 2 and 4). This loss of ankyrin from the detergent-insoluble pool was further stimulated by the addition of okadaic acid to the media (Figure 6, lane 6). However, in contrast to control cells, the decrease in detergent-insoluble ankyrin that occurred in the presence of okadaic acid correlated with a substantial increase in the detergent-soluble form of the polypeptide (Figure 6, lane 5).
Figure 6.

Okadaic acid stimulates the dissociation of ankyrin from the detergent-insoluble pool of chicken erythroid cells. Erythroid cells were pulsed with 35S-Translabel for 15 min and chased for 4 h. At this time, an aliquot of cells was detergent lysed, and immunoprecipitates were prepared from the detergent-soluble (S) and -insoluble (I) fractions with ankyrin-specific antibodies (lanes 1 and 2). The remaining cells were chased an additional 2 h in the absence (lanes 3 and 4) or presence (lanes 5 and 6) of 1 μM okadaic acid before detergent lysis and processing for immunoprecipitation. Samples from each time point were electrophoresed on a 6% SDS polyacrylamide gel and labeled polypeptides were detected by fluorography. The top panel in the figure shows the region of the gel where the ankyrin isoforms migrate. The bottom panel shows the region of the gel where coprecipitating AE1 migrates. The autoradiogram in the bottom panel was exposed 5 times longer than the autoradiogram in the top panel. Dashes to the left of the figure indicate the 225- and 205-kDa ankyrin isoforms. Dashes to the right of the figure indicate the hyperphosphorylated 225- (225*) and 205-kDa (205*) ankyrin isoforms.
The shift in the solubility properties of ankyrin in cells treated with okadaic acid was also observed for labeled anion exchangers that coprecipitated with ankyrin. Quantitation of multiple experiments revealed that 67.4 ± 2.6% (n = 3) of the AE1 polypeptides that coprecipitated with ankyrin in untreated cells after a 4-h chase were associated with the detergent-insoluble form of ankyrin (Figure 6, lanes 1 and 2). A similar profile was observed in untreated cells after a 6-h chase (Figure 6, lanes 3 and 4). However, only 24.9 ± 1.2% (n = 3) of the AE1 that coprecipitated with ankyrin in okadaic acid-treated cells was associated with the detergent-insoluble form of the polypeptide (Figure 6, lanes 5 and 6). These results suggest that okadaic acid treatment dissociated a large fraction of AE1/ankyrin-containing complexes from the detergent-insoluble pool, and in the presence of okadaic acid these detergent-soluble complexes were stable. The fact that okadaic acid treatment shifted the majority of AE1/ankyrin complexes to the soluble pool yet had no discernible effect on the steady-state detergent solubility properties of AE1 is again consistent with the notion that a small percentage of AE1 insolubility is due to its association with ankyrin.
Treatment of Cells with Calyculin A Stimulates Dissociation of Ankyrin from Spectrin Cytoskeleton and Alters Solubility Properties of Spectrin-independent Pool of Ankyrin
Because spectrin was not present in stoichiometric amounts in the ankyrin immunoprecipitates described above, the molecular basis for the detergent insolubility of ankyrin was unclear. Other investigators have shown that in vitro interactions between spectrin and ankyrin are sensitive to salt concentration (Bennett and Branton, 1977). Therefore, it was possible that spectrin–ankyrin interactions were disrupted during our immunoprecipitation protocol, which was carried out in isotonic buffer (150 mM NaCl) containing 1% Triton X-100. To investigate this possibility, cell fractionation and immunoprecipitations with α-spectrin antibodies were performed with the use of a low salt buffer (10 mM NaCl) containing 1% Triton X-100. After precipitation, the immune complexes were subjected to immunoblotting analysis with ankyrin-specific antibodies. These studies demonstrated that the 225-kDa ankyrin isoform exists in an α-spectrin–containing complex that almost exclusively accumulates in the detergent-insoluble fraction of cells lysed in low salt buffer (Figure 7A, bottom, lane 2). Longer exposure of the immunoblot revealed that the 205-kDa ankyrin isoform behaves identically to the 225-kDa isoform (our unpublished data). Treatment of cells with calyculin A altered the interaction of ankyrin with spectrin. Quantitation of multiple experiments has indicated there is a 49.2 ± 5.1% (n = 3) reduction in the total amount of ankyrin that coprecipitated with spectrin in calyculin A-treated cells (Figure 7A, bottom, lanes 3 and 4). Furthermore, the ankyrin polypeptides that still coprecipitated with spectrin in calyculin A-treated cells were primarily present in detergent-soluble complexes. Additional immunoblotting analyses revealed that the majority of α-spectrin and β-spectrin accumulate in the detergent-insoluble or cytoskeletal fraction of these embryonic erythroid cells, and calyculin A treatment resulted in a slight but reproducible shift of spectrin to the soluble pool (Figure 7A). These results indicate that calyculin A treatment of cells stimulated the dissociation of a significant fraction of ankyrin from the detergent-insoluble spectrin cytoskeleton. The accumulation of soluble ankyrin/spectrin complexes observed in calyculin A-treated cells may result from the release of intact complexes from the cytoskeleton or may be due to the formation of new ankyrin/spectrin complexes in the soluble pool. Regardless of their origin, these soluble complexes are stable in the presence of calyculin A.
Figure 7.

Effect of calyculin A on the solubility properties of ankyrin/spectrin and ankyrin/AE1 complexes. Erythroid cells were incubated in complete media for 2 h at 37°C in the absence (A, lanes 1 and 2) or presence (A, lanes 3 and 4) of 100 nM calyculin A. The cells were then lysed in low salt buffer containing 1% Triton X-100, and separated into soluble (S) and insoluble (I) fractions by centrifugation. Equivalent amounts of the resulting fractions were electrophoresed on a 6% SDS polyacrylamide gel, and transferred to nitrocellulose. The filters were incubated with an α-spectrin-specific mAb; an affinity-purified rabbit antibody specific for β-spectrin, or with the ankyrin-specific antibody. Immunoprecipitates prepared from the soluble and insoluble fractions with the α-spectrin antibody were washed with the low salt lysis buffer and processed for immunoblotting analysis with ankyrin-specific antibodies. After washing, blots were incubated with horseradish peroxidase-conjugated secondary antibodies, and immunoreactive species were detected by enhanced chemiluminescence. Alternatively, erythroid cells incubated in the absence (B, lanes 1, 2, 5, and 6) or presence (B, lanes 3, 4, 7, and 8) of 100 nM calyculin A were lysed in low salt buffer containing 1% Triton X-100 (B, lanes 1–4) or isotonic buffer containing 1% Triton X-100 (B, lanes 5–8). AE1 immunoprecipitates prepared from the resulting detergent soluble and insoluble fractions were washed with lysis buffer and processed for immunoblotting with ankyrin-specific antibodies. Dashes to the left of each panel indicate the basally phosphorylated (225) and hyperphosphorylated (225*) 225-kDa ankyrin isoform.
Interestingly, even though the majority of cytoskeletal ankyrin/spectrin complexes were either disrupted or shifted to the soluble pool as a result of calyculin treatment, quantitation of multiple experiments indicated that 69.6 ± 2.1% (n = 3) of total ankyrin still fractionated in the detergent-insoluble pool when calyculin A-treated cells were lysed in low salt buffer (Figure 7A). This suggested that ankyrin can fractionate in the detergent-insoluble pool independent of its association with cytoskeletal spectrin. To determine whether this spectrin-independent insoluble pool of ankyrin is associated with AE1, cells incubated in the absence or presence of calyculin A were detergent lysed in low salt buffer containing 1% Triton X-100. AE1 immunoprecipitates prepared from the detergent-soluble and -insoluble fractions from these cells were blotted and probed with ankyrin antibodies. In contrast to what was observed with complexes containing ankyrin and spectrin (Figure 7A, bottom), calyculin A treatment had little effect on the fractionation properties of complexes containing AE1 and ankyrin when cells were fractionated in low salt buffer (Figure 7B, lanes 1–4). However, calyculin A treatment was sufficient to shift most of the insoluble ankyrin and AE1 containing complexes to the soluble pool if cells were fractionated in isotonic buffer containing 1% Triton X-100 (Figure 7B, lanes 5–8), consistent with the results shown in Figure 6. These analyses indicated that the solubility properties of both the spectrin-dependent and spectrin-independent pools of ankyrin can be altered by phosphorylation. Furthermore, the different salt sensitivity of the ankyrin/spectrin and ankyrin/AE1 complexes after calyculin A treatment indicated that some ankyrin/AE1 complexes acquire detergent insolubility by a mechanism independent of their association with the spectrin cytoskeleton.
Phosphorylation Status of Ankyrin Regulates Its Ability to Bind Spectrin In Vitro
The majority of spectrin dissociates from ankyrin in cells lysed in isotonic buffer containing 1% Triton X-100 (our unpublished data). This property has allowed us to investigate whether the phosphorylation state of ankyrin directly affects its ability to associate with spectrin in vitro. Erythroid cells were lysed in isotonic buffer containing 1% Triton X-100, and an α-spectrin immunoprecipitate was prepared from the detergent-insoluble fraction in the same buffer. This precipitate was washed into low salt buffer containing 1% Triton X-100 and incubated with ankyrin that was immunopurified from untreated or calyculin A-treated erythroid cells that had been lysed in isotonic buffer containing 1% Triton X-100. After extensive washing, the precipitates were blotted and probed with ankyrin antibodies. Incubation of the α-spectrin immunoprecipitate with buffer alone revealed there was a background level of the 225-kDa ankyrin isoform still associated with spectrin immunoprecipitated from cells lysed in isotonic buffer (Figure 8, lane 3). However, the amount of the 225-kDa ankyrin isoform associated with spectrin significantly increased when the spectrin immunoprecipitate was incubated with ankyrin immunopurified from control cells (Figure 8, lane 1). Longer exposure of this immunoblot indicated that the 205-kDa ankyrin isoform from control cells also bound to spectrin at levels significantly above background (our unpublished data). In contrast, spectrin bound very little of the hyperphosphorylated ankyrin that had been immunopurified from calyculin A-treated cells (Figure 8, lane 2). This result, which was obtained in two independent experiments, indicates that ankyrin hyperphosphorylation reduces its ability to associate with spectrin in vitro. Furthermore, these data provide a molecular explanation for the partial dissociation of hyperphosphorylated ankyrin from the spectrin cytoskeleton in calyculin A-treated cells.
Figure 8.

Ankyrin hyperphosphorylation reduces its ability to associate with spectrin in vitro. Erythroid cells were lysed in isotonic buffer (150 mM NaCl) containing 1% Triton X-100, and an α-spectrin immunoprecipitate was prepared from the resulting detergent-insoluble fraction. This immunoprecipitate was washed into low salt buffer (10 mM NaCl) containing 1% Triton X-100 and subsequently incubated with low salt buffer alone (lane 3), or with ankyrin that had been immunopurified from control (lane 1) or calyculin A-treated (lane 2) cells. After extensive washing, the immunoprecipitates were split in half and processed for immunoblotting with ankyrin-specific or α-spectrin-specific antibodies. Lanes showing 10% of the input ankyrin from control (Ank) and calyculin A-treated cells (Ank*) are included. Dashes to the left of the ankyrin immunoblot indicate the basally phosphorylated (225) and hyperphosphorylated (225*) 225-kDa ankyrin isoform.
Erythroid Ankyrin Colocalizes with Spectrin in Perinuclear Compartment
Localization studies have shown that chicken erythroid AE1 anion exchangers and ankyrin accumulate in the plasma membrane and in a perinuclear compartment that overlaps the distribution of the Golgi (Ghosh and Cox, 1999). In addition, these analyses indicated that a substantial fraction of the perinuclear pool of these polypeptides is detergent insoluble. Similar immunolocalization studies have indicated that α-spectrin (Figure 9A) and β-spectrin (Figure 9B) also accumulate in the plasma membrane and in a perinuclear compartment of chicken embryonic erythroid cells. This pattern of localization has previously been observed for α-spectrin in human erythroid cells that have not yet undergone terminal differentiation (Nehls et al., 1993). Extraction of chicken erythroid cells with a low salt buffer containing 1% Triton X-100 before fixation revealed that cytoskeletal α-spectrin is present both on the cell surface and in the perinuclear compartment of these cells (our unpublished data). However, the cytoskeletal pool of α-spectrin often resided solely in the perinuclear compartment where it overlapped the distribution of ankyrin (Figure 9, C–E). A similar result was obtained when preextracted cells were double stained with β-spectrin and ankyrin antibodies (Figure 9, F–H). The observation that a subset of detergent-insoluble ankyrin accumulates in regions of the cell devoid of spectrin (Figure 9, C and F, arrowheads) lends support to the coprecipitation data that indicated a spectrin-independent pool of detergent-insoluble ankyrin exists in these embryonic erythroid cells.
Figure 9.

α-Spectrin and β-spectrin colocalize with a subset of detergent-insoluble ankyrin in chicken erythroid cells. Chicken erythroid cells were fixed in 1% paraformaldehyde in PBS, and permeabilized with acetone. The cells were then incubated with the α-spectrin-specific mAb followed by DAM-IgG conjugated to FITC (A). Alternatively, cells were incubated with an affinity purified β-spectrin-specific antibody followed by DAR-IgG conjugated to lissamine (B). In some instances, erythroid cells were extracted in low salt buffer containing 1% Triton X-100 before fixation. These preextracted cells were incubated with the α-spectrin monoclonal (D) and the rabbit antibody specific for ankyrin (C) followed by DAM-IgG conjugated to FITC and DAR-IgG conjugated to lissamine. Alternatively, preextracted cells were incubated with the affinity-purified β-spectrin–specific antibody (G) followed by GAR-IgG Fab fragments conjugated to tetramethylrhodamine isothiocyanate. These cells were then washed and incubated with the ankyrin-specific antibody that had been directly conjugated to Alexa 488 (F). After washing, immunoreactive polypeptides were visualized with the use of a Zeiss Axiophot microscope. E and H correspond to the merged images of C and D, and F and G, respectively. Arrowheads in C and F indicate ankyrin-staining regions that are devoid of spectrin. Bars, 2 μm.
DISCUSSION
Our studies have revealed that the association of chicken erythroid ankyrin isoforms with the detergent-insoluble fraction of erythroid cells occurs via spectrin-dependent and spectrin-independent pathways (Figure 10). Furthermore, some of the spectrin-independent pool of detergent-insoluble ankyrin exists in a complex with the AE1 anion exchanger. Other investigators have shown that the interaction of human erythroid ankyrin with AE1 in vitro is complex, involving two separate sites in ankyrin (repeats 7–12 and 13–24) (Michaely and Bennett, 1995), and two regions in the cytoplasmic domain of AE1 that are distant in the primary structure (Davis et al., 1989; Willardson et al., 1989; Ding et al., 1994, 1996). In addition to this structural complexity, the association of ankyrin with AE1 is characterized by interactions of both low and high affinity (Van Dort et al., 1998). The consequence of these distinct in vitro interactions on the detergent solubility properties of these polypeptides in vivo is not understood. To begin to address this issue, kinetic analyses coupled with quantitative immunoprecipitations have examined how interactions between ankyrin and AE1 correlate with changes in the detergent solubility properties of these polypeptides in chicken embryonic erythroid cells. These studies have shown that ∼45% of detergent-insoluble ankyrin exists in a stable complex with AE1. This result is consistent with observations made with AE1 knockout mice (Peters et al., 1996). Red cells from these animals exhibit approximately a 50% reduction in their steady-state levels of ankyrin, suggesting that the stable accumulation of approximately half of the cellular pool of erythroid ankyrin is dependent upon its interaction with AE1.
Figure 10.

Model depicting the fate of ankyrin/AE1 complexes in chicken erythroid cells. Thick arrows in the model indicate the predominant pathways. In the absence of serine and threonine phosphatase inhibitors (pathways indicated by purple arrows), detergent-soluble complexes of ankyrin and AE1 associate with the detergent-insoluble or cytoskeletal fraction of erythroid cells in a spectrin-dependent and spectrin-independent manner. These complexes then dissociate from the cytoskeletal fraction, and ankyrin turns over. Whether phosphorylation plays a role in the cytoskeletal dissociation and turn over of ankyrin under these conditions is not known. When cells are incubated in the presence of serine and threonine phosphatase inhibitors (pathways indicated by orange arrows), ankyrin becomes hyperphosphorylated. Ankyrin hyperphosphorylation does not block ankyrin/AE1 complex formation. However, hyperphosphorylation of ankyrin significantly blocks the association of soluble ankyrin/AE1 complexes with the spectrin-dependent and spectrin-independent cytoskeletal fractions. These phosphatase inhibitors also stimulate the dissociation of ankyrin/AE1 complexes from the cytoskeletal pool. After dissociation, ankyrin/AE1 complexes containing hyperphosphorylated ankyrin stably accumulate in the soluble pool.
The analyses described above have further indicated that ∼45% of total detergent-insoluble ankyrin is turned over by 4–6 h postsynthesis. This rate of turnover is also observed for ankyrin molecules associated with the AE1 anion exchanger, whereas AE1 itself is stable during this time frame (Figure 10). Given that the steady-state pool of detergent-insoluble ankyrin is relatively stable, these results imply that “cytoskeletal” ankyrin is undergoing a fairly rapid cycle of association, dissociation, and turnover. The transient nature of ankyrin in these complexes suggests these embryonic erythroid cells possess a dynamic membrane cytoskeleton that is continuously being remodeled. Other isoforms of ankyrin have been proposed to transiently associate with membrane proteins. Polypeptides derived from the Ank B gene are required for the proper localization of the calcium homeostasis machinery in muscle, including ryanodine receptors, IP3 receptors and SERCA. Yet, these ank B isoforms do not continuously associate with these membrane proteins in stoichiometric complexes (Tuvia et al., 1999).
To investigate the role of phosphorylation in regulating the dynamic properties of chicken erythroid ankyrin, we examined the effect of serine and threonine phosphatase inhibitors on ankyrin solubility and stability. Treatment of erythroid cells with these reagents resulted in the hyperphosphorylation of the 225- and 205-kDa ankyrin isoforms, and stimulated the dissociation of ankyrin-containing complexes from the detergent-insoluble pool. Additional pulse-chase studies have indicated that these reagents also significantly blocked the shift of newly synthesized ankyrin from the soluble to the insoluble pool (our unpublished data). The combined effect of these inhibitors on the fractionation properties of both the newly synthesized and preexisting pools of ankyrin (Figure 10), coupled with the fact that soluble ankyrin did not turn over in the presence of these reagents resulted in a dramatic shift of total ankyrin from the detergent-insoluble to the -soluble pool.
The observation that the in vitro phosphorylation of ankyrin reduces its affinity for both spectrin and AE1 (Cianci et al., 1988) suggested potential mechanisms whereby ankyrin hyperphosphorylation could regulate its detergent solubility properties in vivo. Our analyses did not indicate that ankyrin hyperphosphorylation affected its capacity to complex with AE1 in vivo. However, there was a substantial reduction in the amount of ankyrin that coprecipitated with spectrin after treatment of cells with calyculin A. In vitro binding studies suggest that the decrease in the amount of ankyrin associated with spectrin is at least in part due to a reduced ability of hyperphosphorylated ankyrin to associate with spectrin. These results, which are the first to document the in vivo consequences of ankyrin phosphorylation, suggest that the ankyrin/spectrin cytoskeleton of red cells can undergo dynamic reorganization in response to changes in ankyrin phosphorylation.
Surprisingly, our studies have shown that a significant fraction of detergent-insoluble ankyrin is not associated with spectrin in chicken embryonic erythroid cells. The coprecipitation data further indicate that at least some of this spectrin-independent pool of detergent-insoluble ankyrin is complexed with AE1 (Figure 10). The acquisition of detergent insolubilty by this spectrin-independent pool of ankyrin/AE1 complexes may be the result of their association with alternative cytoskeletal receptors, such as filamin, which coprecipitates with ankyrin from embryonic erythroid cells (our unpublished data). Alternatively, the acquisition of detergent insolubility by ankyrin/AE1 complexes may be the result of conformational changes in one or both of these proteins after complex formation.
In conclusion, our coprecipitation and localization data suggest that chicken erythroid ankyrin functions as a versatile adapter, linking elements of the cytoskeleton to various membrane compartments in the cell. Future studies will characterize the specific phosphorylation/dephosphorylation events that regulate complex formation and stability of this multifunctional adapter protein. In addition, these analyses will identify the kinases and phosphatases that control these processes in vivo.
ACKNOWLEDGMENTS
We thank Dr. R. Bloch for kindly providing β-spectrin antibodies. We also thank F.C. Dorsey and Dr. K.H. Cox for critical evaluation of the manuscript. This research was supported by a grant from the National Chapter of the American Heart Association (96–008610). S.G. was supported by the Alma and Hal Reagan Predoctoral Research Fellowship.
REFERENCES
- Alper SL, Palfrey HC, DeRiemer SA, Greengard P. Hormonal control of protein phosphorylation in turkey erythrocytes. Phosphorylation by cAMP-dependent and Ca2+-dependent protein kinases of distinct sites in goblin, a high molecular weight protein of the plasma membrane. J Biol Chem. 1980;255:11029–11039. [PubMed] [Google Scholar]
- Bennett V. Ankyrins. Adaptors between diverse plasma membrane proteins and the cytoplasm. J Biol Chem. 1992;267:8703–8706. [PubMed] [Google Scholar]
- Bennett V, Branton D. Selective association of spectrin with the cytoplasmic surface of human erythrocyte plasma membranes. J Biol Chem. 1977;252:2753–2763. [PubMed] [Google Scholar]
- Bennett V, Stenbuck PJ. The membrane attachment protein for spectrin is associated with band 3 in human erythrocyte membranes. Nature. 1979;280:468–473. doi: 10.1038/280468a0. [DOI] [PubMed] [Google Scholar]
- Bennett V, Stenbuck PJ. Association between ankyrin and the cytoplasmic domain of band 3 isolated from the human erythrocyte membrane. J Biol Chem. 1980;255:6424–6432. [PubMed] [Google Scholar]
- Cianci CD, Giorgi M, Morrow JS. Phosphorylation of ankyrin down-regulates its cooperative interaction with spectrin and protein 3. J Cell Biochem. 1988;37:301–315. doi: 10.1002/jcb.240370305. [DOI] [PubMed] [Google Scholar]
- Cleveland DW, Fischer SG, Kirschner MW, Laemmli UK. Peptide mapping by limited proteolysis in sodium dodecyl sulfate and analysis by gel electrophoresis. J Biol Chem. 1977;252:1102–1106. [PubMed] [Google Scholar]
- Cox KH, Adair-Kirk TL, Cox JV. Four variant chicken erythroid AE1 anion exchangers. Role of the alternative N-terminal sequences in intracellular targeting in transfected human erythroleukemia cells. J Biol Chem. 1995;270:19752–19760. doi: 10.1074/jbc.270.34.19752. [DOI] [PubMed] [Google Scholar]
- Davis L, Lux SE, Bennett V. Mapping the ankyrin-binding site of the human erythrocyte anion exchanger. J Biol Chem. 1989;264:9665–9672. [PubMed] [Google Scholar]
- Devarajan P, Stabach PR, De Matteis MA, Morrow JS. Na+,K+-ATPase transport from endoplasmic reticulum to Golgi requires the Golgi spectrin-ankyrin G119 skeleton in Madin Darby canine kidney cells. Proc Natl Acad Sci USA. 1997;94:10711–10716. doi: 10.1073/pnas.94.20.10711. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devarajan P, Stabach PR, Mann AS, Ardito T, Kashgarian M, Morrow JS. Identification of a small cytoplasmic ankyrin (AnkG119) in the kidney and muscle that binds beta I sigma spectrin and associates with the Golgi apparatus. J Cell Biol. 1996;133:819–830. doi: 10.1083/jcb.133.4.819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ding Y, Casey JR, Kopito RR. The major kidney AE1 isoform does not bind ankyrin (Ank1) in vitro. An essential role for the 79 NH2-terminal amino acid residues of band 3. J Biol Chem. 1994;269:32201–32208. [PubMed] [Google Scholar]
- Ding Y, Kobayashi S, Kopito R. Mapping of ankyrin binding determinants on the erythroid anion exchanger, AE1. J Biol Chem. 1996;271:22494–22498. doi: 10.1074/jbc.271.37.22494. [DOI] [PubMed] [Google Scholar]
- Ghosh S, Cox KH, Cox JV. Chicken erythroid AE1 anion exchangers associate with the cytoskeleton during recycling to the Golgi. Mol Biol Cell. 1999;10:455–469. doi: 10.1091/mbc.10.2.455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammerton RW, Krzeminski KA, Mays RW, Ryan TA, Wollner DA, Nelson WJ. Mechanism for regulating cell surface distribution of Na+,K+-ATPase in polarized epithelial cells. Science. 1991;254:847–850. doi: 10.1126/science.1658934. [DOI] [PubMed] [Google Scholar]
- Michaely P, Bennett V. The ANK repeats of erythrocyte ankyrin form two distinct but cooperative binding sites for the erythrocyte anion exchanger. J Biol Chem. 1995;270:22050–22057. doi: 10.1074/jbc.270.37.22050. [DOI] [PubMed] [Google Scholar]
- Nehls V, Zeitler-Zapf P, Drenckhahn D. Different sequences of expression of band 3, spectrin, and ankyrin during normal erythropoiesis and erythroleukemia. Am J Pathol. 1993;142:1565–1573. [PMC free article] [PubMed] [Google Scholar]
- Peters LL, et al. Anion exchanger 1 (band 3) is required to prevent erythrocyte membrane surface loss but not to form the membrane skeleton. Cell. 1996;86:917–927. doi: 10.1016/s0092-8674(00)80167-1. [DOI] [PubMed] [Google Scholar]
- Tse WT, Lux SE. Red blood cell membrane disorders. Br J Hematol. 1999;104:2–13. doi: 10.1111/j.1365-2141.1999.01130.x. [DOI] [PubMed] [Google Scholar]
- Tuvia S, Buhusi M, Davis L, Reedy M, Bennett V. Ankyrin-B is required for intracellular sorting of structurally diverse Ca+2 homeostasis proteins. J Cell Biol. 1999;147:995–1008. doi: 10.1083/jcb.147.5.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Dort HM, Moriyama R, Low PS. Effect of band 3 subunit equilibrium on the kinetics and affinity of ankyrin binding to erythrocyte membrane vesicles. J Biol Chem. 1998;273:14819–14826. doi: 10.1074/jbc.273.24.14819. [DOI] [PubMed] [Google Scholar]
- Willardson BM, Thevenin BJ, Harrison ML, Kuster WM, Benson MD, Low PS. Localization of the ankyrin-binding site on erythrocyte membrane protein, band 3. J Biol Chem. 1989;264:15893–15899. [PubMed] [Google Scholar]
- Zhou D, Birkenmeier CS, Williams MW, Sharp JJ, Barker JE, Bloch RJ. Small, membrane-bound, alternatively spliced forms of ankyrin 1 associated with the sarcoplasmic reticulum of mammalian skeletal muscle. J Cell Biol. 1997;136:621–631. doi: 10.1083/jcb.136.3.621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou D, Lambert S, Malen PL, Carpenter S, Boland LM, Bennett V. AnkyrinG is required for clustering of voltage-gated Na+ channels at axon initial segments and for normal action potential firing. J Cell Biol. 1998;143:1295–304. doi: 10.1083/jcb.143.5.1295. [DOI] [PMC free article] [PubMed] [Google Scholar]