

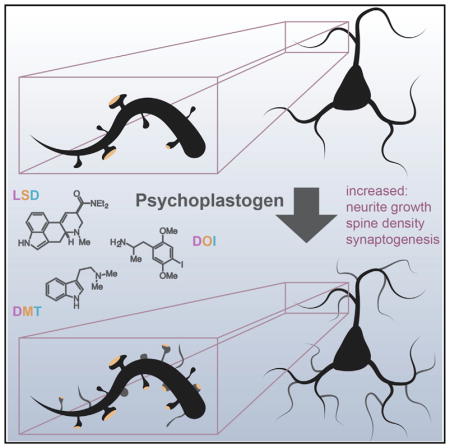
SUMMARY
Atrophy of neurons in the prefrontal cortex (PFC) plays a key role in the pathophysiology of depression and related disorders. The ability to promote both structural and functional plasticity in the PFC has been hypothesized to underlie the fast-acting antidepressant properties of the dissociative anesthetic ketamine. Here, we report that, like ketamine, serotonergic psychedelics are capable of robustly increasing neuritogenesis and/or spinogenesis both in vitro and in vivo. These changes in neuronal structure are accompanied by increased synapse number and function, as measured by fluorescence microscopy and electrophysiology. The structural changes induced by psychedelics appear to result from stimulation of the TrkB, mTOR, and 5-HT2A signaling pathways and could possibly explain the clinical effectiveness of these compounds. Our results underscore the therapeutic potential of psychedelics and, importantly, identify several lead scaffolds for medicinal chemistry efforts focused on developing plasticity-promoting compounds as safe, effective, and fast-acting treatments for depression and related disorders.
In Brief
Ly et al. demonstrate that psychedelic compounds such as LSD, DMT, and DOI increase dendritic arbor complexity, promote dendritic spine growth, and stimulate synapse formation. These cellular effects are similar to those produced by the fast-acting antidepressant ketamine and highlight the potential of psychedelics for treating depression and related disorders.
INTRODUCTION
Neuropsychiatric diseases, including mood and anxiety disorders, are some of the leading causes of disability worldwide and place an enormous economic burden on society (Gustavsson et al., 2011; Whiteford et al., 2013). Approximately one-third of patients will not respond to current antidepressant drugs, and those who do will usually require at least 2–4 weeks of treatment before they experience any beneficial effects (Rush et al., 2006). Depression, post-traumatic stress disorder (PTSD), and addiction share common neural circuitry (Arnsten, 2009; Russo et al., 2009; Peters et al., 2010; Russo and Nestler, 2013) and have high comorbidity (Kelly and Daley, 2013). A preponderance of evidence from a combination of human imaging, postmortem studies, and animal models suggests that atrophy of neurons in the prefrontal cortex (PFC) plays a key role in the pathophysiology of depression and related disorders and is precipitated and/or exacerbated by stress (Arnsten, 2009; Autry and Monteggia, 2012; Christoffel et al., 2011; Duman and Aghajanian, 2012; Duman et al., 2016; Izquierdo et al., 2006; Pittenger and Duman, 2008; Qiao et al., 2016; Russo and Nestler, 2013). These structural changes, such as the retraction of neurites, loss of dendritic spines, and elimination of synapses, can potentially be counteracted by compounds capable of promoting structural and functional neural plasticity in the PFC (Castrén and Antila, 2017; Cramer et al., 2011; Duman, 2002; Hayley and Litteljohn, 2013; Kolb and Muhammad, 2014; Krystal et al., 2009; Mathew et al., 2008), providing a general solution to treating all of these related diseases. However, only a relatively small number of compounds capable of promoting plasticity in the PFC have been identified so far, each with significant drawbacks (Castrén and Antila, 2017). Of these, the dissociative anesthetic ketamine has shown the most promise, revitalizing the field of molecular psychiatry in recent years.
Ketamine has demonstrated remarkable clinical potential as a fast-acting antidepressant (Berman et al., 2000; Ionescu et al., 2016; Zarate et al., 2012), even exhibiting efficacy in treatment-resistant populations (DiazGranados et al., 2010; Murrough et al., 2013; Zarate et al., 2006). Additionally, it has shown promise for treating PTSD (Feder et al., 2014) and heroin addiction (Krupitsky et al., 2002). Animal models suggest that its therapeutic effects stem from its ability to promote the growth of dendritic spines, increase the synthesis of synaptic proteins, and strengthen synaptic responses (Autry et al., 2011; Browne and Lucki, 2013; Li et al., 2010).
Like ketamine, serotonergic psychedelics and entactogens have demonstrated rapid and long-lasting antidepressant and anxiolytic effects in the clinic after a single dose (Bouso et al., 2008; Carhart-Harris and Goodwin, 2017; Grob et al., 2011; Mithoefer et al., 2013, 2016; Nichols et al., 2017; Sanches et al., 2016; Osório et al., 2015), including in treatment-resistant populations (Carhart-Harris et al., 2016, 2017; Mithoefer et al., 2011; Oehen et al., 2013; Rucker et al., 2016). In fact, there have been numerous clinical trials in the past 30 years examining the therapeutic effects of these drugs (Dos Santos et al., 2016), with 3,4-methylenedioxymethamphetamine (MDMA) recently receiving the “breakthrough therapy” designation by the Food and Drug Administration for treating PTSD. Furthermore, classical psychedelics and entactogens produce antidepressant and anxiolytic responses in rodent behavioral tests, such as the forced swim test (Cameron et al., 2018) and fear extinction learning (Cameron et al., 2018; Catlow et al., 2013; Young et al., 2015), paradigms for which ketamine has also been shown to be effective (Autry et al., 2011; Girgenti et al., 2017; Li et al., 2010). Despite the promising antidepressant, anxiolytic, and anti-addictive properties of serotonergic psychedelics, their therapeutic mechanism of action remains poorly understood, and concerns about safety have severely limited their clinical usefulness.
Because of the similarities between classical serotonergic psychedelics and ketamine in both preclinical models and clinical studies, we reasoned that their therapeutic effects might result from a shared ability to promote structural and functional neural plasticity in cortical neurons. Here, we report that serotonergic psychedelics and entactogens from a variety of chemical classes (e.g., amphetamine, tryptamine, and ergoline) display plasticity-promoting properties comparable to or greater than ketamine. Like ketamine, these compounds stimulate structural plasticity by activating the mammalian target of rapamycin (mTOR). To classify the growing number of compounds capable of rapidly promoting induced plasticity (Castrén and Antila, 2017), we introduce the term “psychoplastogen, ” from the Greek roots psych- (mind), -plast (molded), and -gen (producing). Our work strengthens the growing body of literature indicating that psychoplastogens capable of promoting plasticity in the PFC might have value as fast-acting antidepressants and anxiolytics with efficacy in treatment-resistant populations and suggests that it may be possible to use classical psychedelics as lead structures for identifying safer alternatives.
RESULTS
Psychedelics Promote Neuritogenesis
Because atrophy of cortical neurons is believed to be a contributing factor to the development of mood and anxiety disorders (Christoffel et al., 2011; Duman and Aghajanian, 2012), we first treated cultured cortical neurons with psychedelics from a variety of structural classes (Figures 1A and S1A) and measured the resulting changes in various morphological features. Using Sholl analysis (Ristanovic et al., 2006), we observed that several psychedelics increased dendritic arbor complexity comparably to ketamine, as measured by the area under the curve of the Sholl plots as well as the maximum number of crossings (Figures 1B–1E and S1B–S1E). This increase in arbor complexity appeared to result from large changes in both the number of dendritic branches and the total length of the arbors (Figures 1F, 1H, S1F, and S1H). Psychedelics had a limited effect on the number of primary dendrites and did not alter the length of the longest dendrite (Figures 1G, 1I, S1G, and S1I).
Figure 1. Psychedelics Promote Neuritogenesis both In Vitro and In Vivo.

(A) Chemical structures of psychedelics.
(B) Representative tracings of cortical neurons (DIV6) treated with compounds.
(C) Sholl analysis demonstrates that psychedelics increase dendritic arbor complexity (n = 39–41 neurons).
(D) Area under the curve (AUC) of the Sholl plots in (C).
(E) Maximum number of crossings (Nmax) of the Sholl plots in (C).
(F–I) Cortical neurons treated with psychedelics display an increase in the number of branches (F), the number of primary dendrites (G), and the total length of the dendritic arbor (H) but not the length of the longest dendrite (I).
(J and K) Class I neurons from Drosophila larvae treated with psychedelics during the first instar display increased branching (J) but not total length of the dendritic arbor (K) (n = 3 neurons).
(L) Representative images of neurons from (J) and (K).
(M and N) Class I neurons from Drosophila larvae treated with psychedelics during the third instar display increased branching (M) but not total length of the dendritic arbor (N) (n = 3 neurons).
(O) Representative images from (M) and (N).
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, as compared to vehicle control (VEH). Scale bars, 30 μm. Data are represented as mean ± SEM. See also Figures S1–S5.
Nearly all psychedelic compounds tested were capable of robustly promoting neuritogenesis, with comparable effects being produced by tryptamines (N,N-dimethyltryptamine [DMT] and psilocin), amphetamines (2,5-dimethoxy-4-iodoamphetamine [DOI] and MDMA), and ergolines (lysergic acid diethylamide [LSD]). As a positive control, we treated cells with 7,8-dihydroxyflavone (DHF), a psychoplastogen structurally dissimilar to classical psychedelics (Jang et al., 2010), and found that it also increased dendritic arbor complexity (Figure S2). This neurite outgrowth structural phenotype seems to only be induced by select compounds because serotonin and D-amphetamine, molecules that are chemically related to classical psychedelics and entactogens, exerted minimal to no effects on neuritogenesis (Figure S2).
To establish the relative potencies and efficacies of hallucinogens and entactogens for promoting neurite outgrowth, we conducted 8-point dose-response studies (Figure S3). We defined 100% and 0% efficacy as the maximum number of crossings induced by ketamine (10 μM) and vehicle (0.1% DMSO), respectively. We chose the 10 μM concentration of ketamine as the upper limit because this concentration of ketamine is reached in the brain following intraperitoneal administration of an antidepressant dose in rats (Yang et al., 2018). For consistency, we used this same concentration when testing the effects of psychedelics and entactogens, with DMT being the only exception. We used a maximum 90 μM concentration of DMT in our studies to more closely mimic the brain concentration of DMT in rats treated with an antidepressant dose (Cohen and Vogel, 1972; Cameron et al., 2018). In this neuritogenesis assay, ketamine’s half maximal effective concentration (EC50) value was 132 nM. Surprisingly, the majority of the psychedelics and entactogens we tested exhibited significantly greater potency than ketamine, with LSD being particularly potent (EC50 = 0.409 nM). In fact, LSD exhibited activity across 8 orders of magnitude into the low picomolar range (Figure S3).
Notably, the anti-addictive alkaloid ibogaine (Alper, 2001; Belgers et al., 2016) was the only psychedelic tested that had absolutely no effect (Figure S4). This was a surprising result because we hypothesized that ibogaine’s long-lasting anti-addictive properties might result from its psychoplastogenic properties. Previous work by He et al. (2005) clearly demonstrated that ibogaine increases the expression of glial cell line-derived neurotrophic factor (GDNF) and that this plasticity-promoting protein is critical to ibogaine’s anti-addictive mechanism of action. Because several reports have suggested that noribogaine, a metabolite of ibogaine, might actually be the active compound in vivo (Zubaran et al., 1999; Baumann et al., 2000, 2001), we decided to test its ability to promote neuritogenesis in cultured cortical neurons. Gratifyingly, noribogaine robustly increased dendritic arbor complexity with an EC50 value comparable to ketamine (Figure S3), providing additional evidence suggesting that it may be the active compound in vivo.
To assess the in vivo effects of classical psychedelics on neuritogenesis, we started treating Drosophila larvae during the first instar with LSD and DOI. As observed in rodent cortical cultures, both LSD and DOI significantly increased dendritic branching of class I sensory neurons; however, they did not increase the total length of the dendritic arbors (Figures 1J–1L). Because of the striking effects of psychedelics on the structures of immature neurons, we hypothesized that they might influence neurodevelopment. To test this, we chronically treated zebrafish embryos with compounds for 6 days immediately following dechorionation and assessed gross morphological changes and behavior. We did not observe any differences in head sizes between the treatment groups, nor did we detect any statistically significant differences in activity levels (Figure S5). Next we assessed the ability of psychedelics to promote neuritogenesis in more mature neurons by starting to treat Drosophila larvae during the late second instar. Again, psychedelics increased the branching of class I neurons, although the effect was less dramatic than that observed when treatment was started during the first instar (Figure 1M–1O). Although different developmental stages might be more or less susceptible to the effects of psychedelics, it is also possible that the smaller effect size observed after administering compounds starting at the later time point was simply the result of treating the larvae for a shorter period of time. Regardless, it was quite surprising to observe compound-induced changes in neuronal structure after initiating treatment during the late second instar because class I neurons are stereotyped and typically possess relatively few higher-order branches (Grueber et al., 2002). Moreover, our results demonstrate that psychedelics can promote changes in neuronal structure across vertebrate (rats) and invertebrate (Drosophila) species, suggesting that they act through an evolutionarily conserved mechanism.
Psychedelics Promote Spinogenesis and Synaptogenesis
In addition to dendritic atrophy, loss of dendritic spines is a hallmark of depression and other neuropsychiatric disorders (Christoffel et al., 2011; Duman and Aghajanian, 2012), so we next assessed the effects of psychedelics on spinogenesis. We treated mature rat cortical cultures for 24 hr with DOI, DMT, and LSD as representative compounds from the amphetamine, tryptamine, and ergoline classes of psychedelics, respectively. All three compounds increased the number of dendritic spines per unit length, as measured by super-resolution structured illumination microscopy (SIM) (Figures 2A, 2B, and S6), with LSD nearly doubling the number of spines per 10 μm. Additionally, treatment caused a shift in spine morphology, favoring immature (thin and filopodium) over more mature (mushroom) spine types (Figure 2C). Colocalization of pre- and postsynaptic markers following treatment demonstrated that psychedelics promoted synaptogenesis by increasing the density, but not the size of synapses (Figure 2D–2F). This increase in synapse density was accompanied by an increase in the density of VGLUT1 puncta, but not PSD-95 puncta, following compound administration (Figures 2G and 2H).
Figure 2. Psychedelics Promote Spinogenesis, Synaptogenesis, and Functional Plasticity.

(A) Representative images of cortical neurons (DIV19) treated with compounds for 24 hr, demonstrating that psychedelics increase the number of dendritic spines (blue, MAP2; orange, F-actin).
(B) Quantification of spine density (n = 56–65 neurons).
(C) Relative proportions of spine types following treatment of cortical cultures with psychedelics (n = 16–21 neurons).
(D) Representative images of cortical neurons (DIV19) treated for 24 hr, demonstrating that psychedelics increase synaptogenesis (green, VGLUT1; magenta, PSD-95; yellow, MAP2). White areas in the VGLUT1 + PSD-95 images indicate colocalization of pre- and postsynaptic makers and are indicated by gray arrows.
(E–H) Quantification of synapse density (E), synapse size (F), presynaptic density (VGLUT1) (G), and postsynaptic density (PSD-95) (H) following 24-hr treatment of cortical neurons (DIV19) (n = 39–42 neurons).
(I) Representative images of Golgi-Cox-stained pyramidal neurons from the PFC of rats 24 hr after receiving a 10 mg/kg dose of DMT.
(J) Quantification of spines from (I), demonstrating that DMT (10 mg/kg) increases spinogenesis in vivo to a comparable extent as ketamine (10 mg/kg) (n = 11–17 neurons).
(K and L) Whole-cell voltage-clamp recordings of layer V pyramidal neurons from slices obtained 24 hr after DMT treatment (10 mg/kg and 1 mg/kg) demonstrate that DMT increases both spontaneous excitatory postsynaptic current (sEPSC) frequency (K) and amplitude (L) (n = 11–38 neurons from 3 animals).
(M) Representative traces for the 10 mg/kg experiments quantified in (K) and (L).
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, as compared to vehicle control (VEH). Data are represented as mean ± SEM. See also Figure S6.
Encouraged by our in vitro results, we next assessed the effects of a single intraperitoneal dose of DMT on spinogenesis in the PFC of adult rats using Golgi-Cox staining. We chose to administer a 10 mg/kg dose of DMT for three reasons. First, all available data suggested that this dose would produce hallucinogenic effects in rats with minimal safety risks (Glennon et al., 1980, 1983; Glennon, 1999; Gatch et al., 2009; Smith et al., 1998; Appel et al., 1999; Winter et al., 2007; Carbonaro et al., 2015; Helsley et al., 1998; Strassman et al., 1994; Nair and Jacob, 2016). Second, we have previously shown that a 10 mg/kg dose of DMT produces positive effects in rat behavioral tests relevant to depression and PTSD (Cameron et al., 2018). Finally, we wanted to directly compare the effects of DMT with ketamine, and seminal studies conducted by Li et al. (2010) had previously demonstrated that a 10 mg/kg dose of ketamine produced a robust increase in dendritic spine density in the PFC of rats. We observed a significant increase in the density of dendritic spines on cortical pyramidal neurons 24 hr after dosing with DMT (Figures 2I and 2J). This effect was comparable with that produced by ketamine at the same dose (Figure 2J). Importantly, this DMT-induced increase in dendritic spine density was accompanied by functional effects. Ex vivo slice recordings revealed that both the frequency and amplitude of spontaneous excitatory postsynaptic currents (EPSCs) were increased following DMT treatment (Figures 2K–2M). Interestingly, 10 mg/kg and 1 mg/kg doses produced similar responses despite the fact that they are predicted to be hallucinogenic and subhallucinogenic, respectively (Strassman et al., 1994; Nair and Jacob, 2016).
Because the half-life of DMT is exceedingly short (~15 min), these results confirm that structural and functional changes induced by DMT persist for hours after the compound has been cleared from the body. Moreover, they demonstrate that DMT produces functional effects on pyramidal neurons of the PFC that mirror those produced by ketamine (Li et al., 2010). Because the PFC is a key brain region involved in extinction learning (Quirk et al., 2006), and both ketamine and DMT have been shown to facilitate fear extinction (Cameron et al., 2018; Girgenti et al., 2017), our results suggest a link between the plasticity-promoting and behavioral effects of these drugs. Because fear extinction can be enhanced by increasing levels of brain-derived neurotrophic factor (BDNF) in the PFC (Peters et al., 2010), and ketamine’s behavioral effects have been shown to be BDNF-dependent (Lepack et al., 2014), we next sought to determine the role of BDNF signaling in the plasticity-promoting effects of classical psychedelics.
Psychedelics Promote Plasticity through a TrkB-and mTOR-Dependent Mechanism
The role of BDNF in both neuritogenesis and spinogenesis is well known (Cohen-Cory et al., 2010), and several reports suggest that psychedelics are capable of increasing levels of neurotrophic factors (He et al., 2005; Martin et al., 2014; Nichols and Sanders-Bush, 2002; Vaidya et al., 1997). Therefore, we treated cortical neurons with BDNF, DOI, and a combination of the two to see whether they had any additive or synergistic effects. Dose-response studies using recombinant BDNF (Figures 3A–3C) revealed that a 50 ng/mL treatment increased neuritogenesis to a comparable extent as DOI (10 μM). Moreover, a combination of the two did not confer any added benefit, suggesting that they operate through a related mechanism (Figures 3D–3F). Next, we treated cortical neurons with DOI, DMT, and LSD for 24 hr before measuring BDNF gene and protein expression using droplet digital PCR (ddPCR) and ELISA, respectively. Although psychedelics did not increase the expression of BDNF transcript (Figure 3G), they did result in a 2-fold increase in BDNF protein levels, although this effect was not statistically significant (Figure 3H). When cortical cultures were co-treated with ANA-12 (Cazorla et al., 2011), a selective antagonist of BDNF’s high-affinity receptor TrkB, the ability of psychedelics or BDNF to stimulate neuritogenesis and spinogenesis was completely blocked (Figure 4).
Figure 3. Psychedelics and BDNF Promote Neuritogenesis via a Related Mechanism.

(A–C) Dose response of recombinant BDNF on neuritogenesis. AUC of the Sholl plots (A), Nmax of the Sholl plots (B), and total number of branches (C) of treated cortical neurons (n = 11–12 neurons per treatment, DIV6) indicate that the highest concentration of BDNF (50 ng/mL) is more effective at promoting neuritogenesis than lower concentrations (5.0 and 0.5 ng/mL).
(D) Sholl analysis (n = 5–10 neurons) demonstrating that DOI (10 μM) increases neuritogenesis to a comparable extent as recombinant BDNF (50 ng/mL). A combination of DOI (10 μM) and BDNF (50 ng/mL) did not have any additive or synergistic effects.
(E) AUC of the Sholl plots in (D).
(F) Nmax of the Sholl plots in (D).
(G and H) Cultured cortical neurons (DIV18) were treated with compounds for 24 hr, and then BDNF gene (G) and protein (H) expression was assessed via ddPCR (n = 4) and ELISA (n = 3–4), respectively.
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, as compared to vehicle control (VEH). Data are represented as mean ± SEM.
Figure 4. Psychedelic-Induced Changes in Neuronal Structure Are Mediated by TrkB.

(A–D) The effects of psychedelics on dendritic arbor complexity are blocked by ANA-12, a selective inhibitor of TrkB, as measured by Sholl analysis of cultured cortical neurons (A) (DIV6). Compound-induced increases in the AUC of the Sholl plots (B), the Nmax of the Sholl plots (C), and the number of dendritic branches (D) are completely blocked by ANA-12 (n = 8–10 neurons).
(E) The spinogenesis-promoting properties of psychedelics are blocked by ANA-12 (n = 19–21 neurons, DIV19).
(F) Control experiment demonstrating that ANA-12 blocks the effects of BDNF on neuritogenesis (n = 11–15 neurons).
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, as compared to vehicle control (VEH) or vehicle + antagonist. Data are represented as mean ± SEM.
Activation of TrkB is known to promote signaling through mTOR (Takei et al., 2004), which plays a key role in structural plasticity (Jaworski et al., 2005; Kumar et al., 2005), the production of proteins necessary for synaptogenesis (Hoeffer and Klann, 2010), and the effects of ketamine (Dwyer and Duman, 2013; Li et al., 2010). Treatment with rapamycin, an mTOR inhibitor, completely blocked psychedelic-induced neuritogenesis (Figure 5), thus confirming that mTOR activation plays a role in the plasticity-promoting effects of classical serotonergic psychedelics.
Figure 5. Psychedelic-Induced Changes in Neuronal Structure Are Mediated by mTOR.

(A–D) The effects of psychedelics on dendritic arbor complexity are blocked by rapamycin, an inhibitor of mTOR, as measured by Sholl analysis of cultured cortical neurons (A) (DIV6). Compound-induced increases in the AUC of the Sholl plots (B), the Nmax of the Sholl plots (C), and the number of dendritic branches (D) are completely blocked by rapamycin (n = 9–12 neurons).
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, as compared to vehicle control (VEH) or vehicle + antagonist. Data are represented as mean ± SEM.
The 5-HT2A Receptor Mediates the Effects of Psychedelics on Structural Plasticity
Finally, we sought to determine whether the 5-HT2A receptor played any role in the plasticity-promoting effects of DOI, DMT, and LSD because this receptor is known to be primarily responsible for the hallucinogenic effects of classical psychedelics (Nichols, 2004, 2016). Furthermore, the psychoplastogenic potencies of these and related compounds correlate well with their 5-HT2A receptor affinities (Figure S3) (i.e., a higher 5-HT2A binding affinity generally predicts more potent psychoplastogenic effects). Control experiments demonstrated that 5-HT2A receptors were expressed on cultured rat cortical neurons at both 6 days in vitro (DIV6) and DIV19 (Figure 6A). Next we found that co-treatment with ketanserin, a selective 5-HT2A antagonist, completely abrogated the ability of DMT, LSD, and DOI to promote both neuritogenesis and spinogenesis (Figures 6B–6F). Ketanserin was also able to block the effects of psilocin as well as the non-classical psychedelic noribogaine and enactogen MDMA (Figure 6G).
Figure 6. The 5-HT2A Receptor Mediates the Effects of Psychedelics on Structural Plasticity.

(A) Rat embryonic cortical neurons express 5-HT2A receptors at both DIV6 and DIV19 (scale bar, 10 μm).
(B) The effects of psychedelics on increasing dendritic arbor complexity are blocked by co-treating with ketanserin, a selective antagonist of 5-HT2A receptors, as measured by Sholl analysis of cultured cortical neurons (DIV6).
(C–E) Compound-induced increases in the AUC of the Sholl plots (C), the Nmax of the Sholl plots (D), and the number of dendritic branches (E) are completely blocked by ketanserin (n = 10–11 neurons, DIV6).
(F) The spinogenesis-promoting properties of psychedelics are blocked by ketanserin (n = 19–20 neurons, DIV19).
(G) Ketanserin also blocks the increased Nmax induced by psilocin, noribogaine, and MDMA.
(H) Ketanserin dose-dependently blocks the psychoplastogenic effects of 10 nM LSD (n = 9–38 neurons, DIV6).
*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, as compared to vehicle control (VEH) or vehicle + antagonist. Data are represented as mean ± SEM.
These initial experiments were performed using doses of psychoplastogens that produced maximal effects on structural plasticity (circa 10 μM) in combination with a 10-fold excess of ketanserin (100 μM). At these concentrations, we could not rule out the possibility of other receptors contributing to the antagonistic effects of ketanserin. Therefore, we treated cultured cortical neurons with a significantly lower dose of LSD (10 nM) and attempted to block its ability to promote neurite outgrowth using increasing doses of ketanserin (Figure 6H). We found that ketanserin blocks the psychoplastogenic effects of LSD by ~50% when treated at 10 nM. This is consistent with the fact that the binding affinities of ketanserin and LSD for the 5-HT2A receptor are roughly equivalent (low nanomolar). Increasing the concentration of ketanserin to 100 nM, 10-fold higher than the concentration of LSD used in this experiment, completely prevented LSD-induced neuritogenesis. At 100 nM, ketanserin is relatively selective for the 5-HT2A receptor, although, at this concentration, we cannot rule out the possible involvement of 5-HT2C, adrenergic, or histamine receptors.
As a final note, the concentration responses of most psychoplastogens had Hill slopes that deviated from 1.0 (Figure S3), implying polypharmacology. Because psychedelics have relatively high affinities for 5-HT2A receptors, it is likely that the effects of psychedelics are mediated primarily through 5-HT2A receptors at low concentrations and modulated by other targets at high concentrations. Interestingly, the concentration response of DMT was the only one to exhibit a Hill slope greater than 1.0, indicating some form of cooperativity.
DISCUSSION
Classical serotonergic psychedelics are known to cause changes in mood (Griffiths et al., 2006, 2008, 2011) and brain function (Carhart-Harris et al., 2017) that persist long after the acute effects of the drugs have subsided. Moreover, several psychedelics elevate glutamate levels in the cortex (Nichols, 2004, 2016) and increase gene expression in vivo of the neurotrophin BDNF as well as immediate-early genes associated with plasticity (Martin et al., 2014; Nichols and Sanders-Bush, 2002; Vaidya et al., 1997). This indirect evidence has led to the reasonable hypothesis that psychedelics promote structural and functional neural plasticity, although this assumption had never been rigorously tested (Bogenschutz and Pommy, 2012; Vollenweider and Kometer, 2010). The data presented here provide direct evidence for this hypothesis, demonstrating that psychedelics cause both structural and functional changes in cortical neurons.
Prior to this study, two reports suggested that psychedelics might be able to produce changes in neuronal structure. Jones et al. (2009) demonstrated that DOI was capable of transiently increasing the size of dendritic spines on cortical neurons, but no change in spine density was observed. The second study showed that DOI promoted neurite extension in a cell line of neuronal lineage (Marinova et al., 2017). Both of these reports utilized DOI, a psychedelic of the amphetamine class. Here we demonstrate that the ability to change neuronal structure is not a unique property of amphetamines like DOI because psychedelics from the ergo-line, tryptamine, and iboga classes of compounds also promote structural plasticity. Additionally, D-amphetamine does not increase the complexity of cortical dendritic arbors in culture, and therefore, these morphological changes cannot be simply attributed to an increase in monoamine neurotransmission.
The identification of psychoplastogens belonging to distinct chemical families is an important aspect of this work because it suggests that ketamine is not unique in its ability to promote structural and functional plasticity. In addition to ketamine, the prototypical psychoplastogen, only a relatively small number of plasticity-promoting small molecules have been identified previously. Such compounds include the N-methyl-D-aspartate (NMDA) receptor ligand GLYX-13 (i.e., rapastinel), the mGlu2/3 antagonist LY341495, the TrkB agonist 7,8-DHF, and the muscarinic receptor antagonist scopolamine (Lepack et al., 2016; Castello et al., 2014; Zeng et al., 2012; Voleti et al., 2013). We observe that hallucinogens from four distinct structural classes (i.e., tryptamine, amphetamine, ergoline, and iboga) are also potent psychoplastogens, providing additional lead scaffolds for medicinal chemistry efforts aimed at identifying neurotherapeutics. Furthermore, our cellular assays revealed that several of these compounds were more efficacious (e.g., MDMA) or more potent (e.g., LSD) than ketamine. In fact, the plasticity-promoting properties of psychedelics and entactogens rivaled that of BDNF (Figures 3A–3C and S3). The extreme potency of LSD in particular might be due to slow off kinetics, as recently proposed following the disclosure of the LSD-bound 5-HT2B crystal structure (Wacker et al., 2017).
Importantly, the psychoplastogenic effects of psychedelics in cortical cultures were also observed in vivo using both vertebrate and invertebrate models, demonstrating that they act through an evolutionarily conserved mechanism. Furthermore, the concentrations of psychedelics utilized in our in vitro cell culture assays were consistent with those reached in the brain following systemic administration of therapeutic doses in rodents (Yang et al., 2018; Cohen and Vogel, 1972). This suggests that neuritogenesis, spinogenesis, and/or synaptogenesis assays performed using cortical cultures might have value for identifying psychoplastogens and fast-acting antidepressants. It should be noted that our structural plasticity studies performed in vitro utilized neurons exposed to psychedelics for extended periods of time. Because brain exposure to these compounds is often of short duration due to rapid metabolism, it will be interesting to assess the kinetics of psychedelic-induced plasticity.
A key question in the field of psychedelic medicine has been whether or not psychedelics promote changes in the density of dendritic spines (Kyzar et al., 2017). Using super-resolution SIM, we clearly demonstrate that psychedelics do, in fact, increase the density of dendritic spines on cortical neurons, an effect that is not restricted to a particular structural class of compounds. Using DMT, we verified that cortical neuron spine density increases in vivo and that these changes in structural plasticity are accompanied by functional effects such as increased amplitude and frequency of spontaneous EPSCs. We specifically designed these experiments to mimic previous studies of ketamine (Li et al., 2010) so that we might directly compare these two compounds, and, to a first approximation, they appear to be remarkably similar. Not only do they both increase spine density and neuronal excitability in the cortex, they seem to have similar behavioral effects. We have shown previously that, like ketamine, DMT promotes fear extinction learning and has antidepressant effects in the forced swim test (Cameron et al., 2018). These results, coupled with the fact that ayahuasca, a DMT-containing concoction, has potent antidepressant effects in humans (Osório et al., 2015; Sanches et al., 2016; Santos et al., 2007), suggests that classical psychedelics and ketamine might share a related therapeutic mechanism.
Although the molecular targets of ketamine and psychedelics are different (NMDA and 5-HT2A receptors, respectively), they appear to cause similar downstream effects on structural plasticity by activating mTOR. This finding is significant because ketamine is known to be addictive whereas many classical psychedelics are not (Nutt et al., 2007, 2010). The exact mechanisms by which these compounds stimulate mTOR is still not entirely understood, but our data suggest that, at least for classical psychedelics, TrkB and 5-HT2A receptors are involved. Although most classical psychedelics are not considered to be addictive, there are still significant safety concerns with their use in medicine because they cause profound perceptual disturbances and still have the potential to be abused. Therefore, the identification of non-hallucinogenic analogs capable of promoting plasticity in the PFC could facilitate a paradigm shift in our approach to treating neuropsychiatric diseases. Moreover, such compounds could be critical to resolving the long-standing debate in the field concerning whether the subjective effects of psychedelics are necessary for their therapeutic effects (Majić et al., 2015). Although our group is actively investigating the psychoplastogenic properties of non-hallucinogenic analogs of psychedelics, others have reported the therapeutic potential of safer structural and functional analogs of ketamine (Moskal et al., 2017; Yang et al., 2015; Zanos et al., 2016).
Our data demonstrate that classical psychedelics from several distinct chemical classes are capable of robustly promoting the growth of both neurites and dendritic spines in vitro, in vivo, and across species. Importantly, our studies highlight the similarities between the effects of ketamine and those of classical serotonergic psychedelics, supporting the hypothesis that the clinical antidepressant and anxiolytic effects of these molecules might result from their ability to promote structural and functional plasticity in prefrontal cortical neurons. We have demonstrated that the plasticity-promoting properties of psychedelics require TrkB, mTOR, and 5-HT2A signaling, suggesting that these key signaling hubs may serve as potential targets for the development of psychoplastogens, fast-acting antidepressants, and anxiolytics. Taken together, our results suggest that psychedelics may be used as lead structures to identify next-generation neurotherapeutics with improved efficacy and safety profiles.
EXPERIMENTAL PROCEDURES
Drugs
For in vitro studies, all compounds were dissolved in DMSO and diluted 1:1,000, with the exception that BDNF was dissolved in water. Cells were treated with DMT at a final concentration of 90 μM (0.1% DMSO), whereas all other compounds were used at 10 μM (0.1% DMSO) unless noted otherwise. For in vivo studies, DMT or ketamine was dissolved in sterile 0.9% saline and administered intraperitoneally at a dose of 10 mg/kg and an injection volume of 1 mL/kg.
Animals
Sprague-Dawley rats were obtained from Charles River Laboratories (Wilmington, MA, USA). The age and sex of the animals used are noted under each individual experimental subheading. All experimental procedures involving animals were approved by the University of California, Davis Institutional Animal Care and Use Committee (IACUC) and adhered to the principles described in the NIH Guide for the Care and Use of Laboratory Animals. The University of California, Davis is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC) and has an Animal Welfare Assurance number (A3433-01) on file with the Office of Laboratory Animal Welfare (OLAW).
Cell Culture
Primary cortical cultures were prepared using tissue from embryonic day 18 (E18) Sprague-Dawley rats. Cells were plated at various densities on poly-D-lysine-coated plates depending on the specific experiment (Supplemental Experimental Procedures). Plating medium consisted of 10% heat-inactivated fetal bovine serum (FBS) (Life Technologies), 1% penicillin-streptomycin (Life Technologies), and 0.5 mM glutamine (Life Technologies) in Neurobasal (Life Technologies). After 15–24 hr, the medium was removed and exchanged for replacement medium containing 1 × B27 supplement (Life Technologies), 1% penicillin-streptomycin, 0.5 mM glutamine, 12.5 μM glutamate, and Neuro-basal. After 96 hr, 50% of the medium was removed and replaced with feed medium containing 1 × B27 supplement, 1% penicillin-streptomycin, and 0.5 mM glutamine. Once per week, until the cultures had reached sufficient maturity for experiments, 50% of the culture medium was removed and replaced with feed medium, with an additional 20% by volume being added to account for evaporation. For experiments using antagonists or inhibitors, cells were pretreated with ANA-12 (10 μM), rapamycin (100 nM), and ketanserin (100 μM) for 10 min prior to addition of test compounds unless otherwise noted. The final DMSO concentration of these experiments was 0.2%. The neurons used in each cellular experiment were taken from at least two different treatment wells, and the wells were randomized to account for plate effects. All of the cellular experiments were replicated on at least two occasions by two or more experimenters.
Statistical Analysis
Appropriate samples sizes were estimated based on our previous experiences performing similar experiments. Data are represented as mean ± SEM. Statistical analyses were performed using GraphPad Prism (version 7.0a). For analyses involving comparison of three or more groups, a one-way analysis of variance (Dunnett’s post hoc test) was utilized. No statistics were calculated for the individual points of the Sholl plots. Instead, statistical analyses were performed on the aggregate data (i.e., the area under the curve of the Sholl plot). Probability distributions from electrophysiology experiments were compared using a Kolmogorov-Smirnov test. *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001 compared with vehicle control or vehicle + antagonist.
Supplementary Material
Highlights.
Serotonergic psychedelics increase neuritogenesis, spinogenesis, and synaptogenesis
Psychedelics promote plasticity via an evolutionarily conserved mechanism
TrkB, mTOR, and 5-HT2A signaling underlie psychedelic-induced plasticity
Noribogaine, but not ibogaine, is capable of promoting structural neural plasticity
Acknowledgments
We thank Lee E. Dunlap for synthesizing DMT, Aurora Martinez-Horta for assistance with Golgi-Cox staining experiments, the Light Microscopy Imaging Facility in the Department of Molecular and Cellular Biology for the use of their Nikon N-SIM, Javier González-Maeso for providing 198A2 cells, and members of the Olson laboratory for discussions regarding the manuscript. We also thank Valentina Popescu in the Department of Classics at UC Davis for assistance with coining the term “psychoplastogen. ” Several antibodies were generously provided by the Stanley Center for Psychiatric Research at the Broad Institute. This work was supported by funds from the UC Davis Department of Chemistry and Department of Biochemistry and Molecular Medicine, a UC Davis Provost’s Undergraduate Fellowship (to A.C.G.), a UC Davis MIND Institute IDDRC grant U54 HD079125 (to M.Y.D. and K.B.), an Alfred P. Sloan Fellowship (FG-2016-6814 to M.Y.D.), and NIH grants T32GM113770 (to C.L.) and 5T32MH082174-09 (to L.P.C. and E.B.).
Footnotes
Supplemental Information includes Supplemental Experimental Procedures and six figures and can be found with this article online at https://doi.org/10.1016/j.celrep.2018.05.022.
AUTHOR CONTRIBUTIONS
D.E.O., C.L., and A.K.M. were responsible for the overall experimental design. C.L. performed the majority of the neuritogenesis, spinogenesis, synaptogenesis, and biochemical experiments described, with A.C.G. and S.S.Z. providing assistance. C.L., L.P.C., and D.E.O. performed the Golgi-Cox staining experiments. L.P.C., J.M.W., and E.V.B. performed the electrophysiology experiments, with J.A.G. supervising these studies. K.M.O.-M. and P.C.W. performed the experiments in Drosophila. M.Y.D. and K.F.B. performed the zebrafish experiments. A.S. developed the program for analyzing synapse number and size. M.R.P. supervised the SIM experiments. W.C.D. performed 5-HT2A expression studies and assisted with fluorescence microscopy data analysis. D.E.O. conceived the project, supervised the experiments, and wrote the manuscript with input from all authors.
DECLARATION OF INTERESTS
D.E.O. has submitted a patent application related to this work (PCT/US2017/054277).
References
- Alper KR. Ibogaine: a review. Alkaloids Chem Biol. 2001;56:1–38. doi: 10.1016/s0099-9598(01)56005-8. [DOI] [PubMed] [Google Scholar]
- Appel JB, West WB, Rolandi WG, Alici T, Pechersky K. Increasing the selectivity of drug discrimination procedures. Pharmacol Biochem Behav. 1999;64:353–358. doi: 10.1016/s0091-3057(99)00089-1. [DOI] [PubMed] [Google Scholar]
- Arnsten AFT. Stress signalling pathways that impair prefrontal cortex structure and function. Nat Rev Neurosci. 2009;10:410–422. doi: 10.1038/nrn2648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Autry AE, Monteggia LM. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol Rev. 2012;64:238–258. doi: 10.1124/pr.111.005108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Autry AE, Adachi M, Nosyreva E, Na ES, Los MF, Cheng PF, Kavalali ET, Monteggia LM. NMDA receptor blockade at rest triggers rapid behavioural antidepressant responses. Nature. 2011;475:91–95. doi: 10.1038/nature10130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baumann MH, Pablo JP, Ali SF, Rothman RB, Mash DC. Noribogaine (12-hydroxyibogamine): a biologically active metabolite of the antiaddictive drug ibogaine. Ann N Y Acad Sci. 2000;914:354–368. doi: 10.1111/j.1749-6632.2000.tb05210.x. [DOI] [PubMed] [Google Scholar]
- Baumann MH, Pablo J, Ali SF, Rothman RB, Mash DC. Comparative neuropharmacology of ibogaine and its O-desmethyl metabolite, noribogaine. Alkaloids Chem Biol. 2001;56:79–113. doi: 10.1016/s0099-9598(01)56009-5. [DOI] [PubMed] [Google Scholar]
- Belgers M, Leenaars M, Homberg JR, Ritskes-Hoitinga M, Schellekens AF, Hooijmans CR. Ibogaine and addiction in the animal model, a systematic review and meta-analysis. Transl Psychiatry. 2016;6:e826. doi: 10.1038/tp.2016.71. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berman RM, Cappiello A, Anand A, Oren DA, Heninger GR, Charney DS, Krystal JH. Antidepressant effects of ketamine in depressed patients. Biol Psychiatry. 2000;47:351–354. doi: 10.1016/s0006-3223(99)00230-9. [DOI] [PubMed] [Google Scholar]
- Bogenschutz MP, Pommy JM. Therapeutic mechanisms of classic hallucinogens in the treatment of addictions: from indirect evidence to testable hypotheses. Drug Test Anal. 2012;4:543–555. doi: 10.1002/dta.1376. [DOI] [PubMed] [Google Scholar]
- Bouso JC, Doblin R, Farré M, Alcázar MA, Gómez-Jarabo G. MDMA-assisted psychotherapy using low doses in a small sample of women with chronic posttraumatic stress disorder. J Psychoactive Drugs. 2008;40:225–236. doi: 10.1080/02791072.2008.10400637. [DOI] [PubMed] [Google Scholar]
- Browne CA, Lucki I. Antidepressant effects of ketamine: mechanisms underlying fast-acting novel antidepressants. Front Pharmacol. 2013;4:161. doi: 10.3389/fphar.2013.00161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cameron LP, Benson CJ, Dunlap LE, Olson DE. Effects of N, N-dimethyltryptamine on rat behaviors relevant to anxiety and depression. ACS Chem Neurosci. 2018 doi: 10.1021/acschemneuro.8b00134. Published online April 24, 2018. [DOI] [PMC free article] [PubMed]
- Carbonaro TM, Eshleman AJ, Forster MJ, Cheng K, Rice KC, Gatch MB. The role of 5-HT2A, 5-HT2C and mGlu2 receptors in the behavioral effects of tryptamine hallucinogens N,N-dimethyltryptamine and N,N-diisopropyltryptamine in rats and mice. Psychopharmacology (Berl) 2015;232:275–284. doi: 10.1007/s00213-014-3658-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carhart-Harris RL, Goodwin GM. The therapeutic potential of psychedelic drugs: past, present, and future. Neuropsychopharmacology. 2017;42:2105–2113. doi: 10.1038/npp.2017.84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carhart-Harris RL, Bolstridge M, Rucker J, Day CM, Erritzoe D, Kaelen M, Bloomfield M, Rickard JA, Forbes B, Feilding A, et al. Psilocybin with psychological support for treatment-resistant depression: an open-label feasibility study. Lancet Psychiatry. 2016;3:619–627. doi: 10.1016/S2215-0366(16)30065-7. [DOI] [PubMed] [Google Scholar]
- Carhart-Harris RL, Roseman L, Bolstridge M, Demetriou L, Pannekoek JN, Wall MB, Tanner M, Kaelen M, McGonigle J, Murphy K, et al. Psilocybin for treatment-resistant depression: fMRI-measured brain mechanisms. Sci Rep. 2017;7:13187. doi: 10.1038/s41598-017-13282-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castello NA, Nguyen MH, Tran JD, Cheng D, Green KN, LaFerla FM. 7,8-Dihydroxyflavone, a small molecule TrkB agonist, improves spatial memory and increases thin spine density in a mouse model of Alzheimer disease-like neuronal loss. PLoS ONE. 2014;9:e91453. doi: 10.1371/journal.pone.0091453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castrén E, Antila H. Neuronal plasticity and neurotrophic factors in drug responses. Mol Psychiatry. 2017;22:1085–1095. doi: 10.1038/mp.2017.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Catlow BJ, Song S, Paredes DA, Kirstein CL, Sanchez-Ramos J. Effects of psilocybin on hippocampal neurogenesis and extinction of trace fear conditioning. Exp Brain Res. 2013;228:481–491. doi: 10.1007/s00221-013-3579-0. [DOI] [PubMed] [Google Scholar]
- Cazorla M, Prémont J, Mann A, Girard N, Kellendonk C, Rognan D. Identification of a low-molecular weight TrkB antagonist with anxiolytic and antidepressant activity in mice. J Clin Invest. 2011;121:1846–1857. doi: 10.1172/JCI43992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christoffel DJ, Golden SA, Russo SJ. Structural and synaptic plasticity in stress-related disorders. Rev Neurosci. 2011;22:535–549. doi: 10.1515/RNS.2011.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen I, Vogel WH. Determination and physiological disposition of dimethyltryptamine and diethyltryptamine in rat brain, liver and plasma. Biochem Pharmacol. 1972;21:1214–1216. doi: 10.1016/0006-2952(72)90119-0. [DOI] [PubMed] [Google Scholar]
- Cohen-Cory S, Kidane AH, Shirkey NJ, Marshak S. Brain-derived neurotrophic factor and the development of structural neuronal connectivity. Dev Neurobiol. 2010;70:271–288. doi: 10.1002/dneu.20774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cramer SC, Sur M, Dobkin BH, O’Brien C, Sanger TD, Trojanowski JQ, Rumsey JM, Hicks R, Cameron J, Chen D, et al. Harnessing neuroplasticity for clinical applications. Brain. 2011;134:1591–1609. doi: 10.1093/brain/awr039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiazGranados N, Ibrahim LA, Brutsche NE, Ameli R, Henter ID, Luckenbaugh DA, Machado-Vieira R, Zarate CA., Jr Rapid resolution of suicidal ideation after a single infusion of an N-methyl-D-aspartate antagonist in patients with treatment-resistant major depressive disorder. J Clin Psychiatry. 2010;71:1605–1611. doi: 10.4088/JCP.09m05327blu. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dos Santos RG, Osório FL, Crippa JA, Riba J, Zuardi AW, Hallak JE. Antidepressive, anxiolytic, and antiaddictive effects of ayahuasca, psilocybin and lysergic acid diethylamide (LSD): a systematic review of clinical trials published in the last 25 years. Ther Adv Psychopharmacol. 2016;6:193–213. doi: 10.1177/2045125316638008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duman RS. Synaptic plasticity and mood disorders. Mol Psychiatry. 2002;7(Suppl 1):S29–S34. doi: 10.1038/sj.mp.4001016. [DOI] [PubMed] [Google Scholar]
- Duman RS, Aghajanian GK. Synaptic dysfunction in depression: potential therapeutic targets. Science. 2012;338:68–72. doi: 10.1126/science.1222939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duman RS, Aghajanian GK, Sanacora G, Krystal JH. Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat Med. 2016;22:238–249. doi: 10.1038/nm.4050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dwyer JM, Duman RS. Activation of mammalian target of rapamycin and synaptogenesis: role in the actions of rapid-acting antidepressants. Biol Psychiatry. 2013;73:1189–1198. doi: 10.1016/j.biopsych.2012.11.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feder A, Parides MK, Murrough JW, Perez AM, Morgan JE, Saxena S, Kirkwood K, Aan Het Rot M, Lapidus KA, Wan LB, et al. Efficacy of intravenous ketamine for treatment of chronic posttraumatic stress disorder: a randomized clinical trial. JAMA Psychiatry. 2014;71:681–688. doi: 10.1001/jamapsychiatry.2014.62. [DOI] [PubMed] [Google Scholar]
- Gatch MB, Rutledge MA, Carbonaro T, Forster MJ. Comparison of the discriminative stimulus effects of dimethyltryptamine with different classes of psychoactive compounds in rats. Psychopharmacology (Berl) 2009;204:715–724. doi: 10.1007/s00213-009-1501-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Girgenti MJ, Ghosal S, LoPresto D, Taylor JR, Duman RS. Ketamine accelerates fear extinction via mTORC1 signaling. Neurobiol Dis. 2017;100:1–8. doi: 10.1016/j.nbd.2016.12.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glennon RA. Arylalkylamine drugs of abuse: an overview of drug discrimination studies. Pharmacol Biochem Behav. 1999;64:251–256. doi: 10.1016/s0091-3057(99)00045-3. [DOI] [PubMed] [Google Scholar]
- Glennon RA, Young R, Rosecrans JA, Kallman MJ. Hallucinogenic agents as discriminative stimuli: a correlation with serotonin receptor affinities. Psychopharmacology (Berl) 1980;68:155–158. doi: 10.1007/BF00432133. [DOI] [PubMed] [Google Scholar]
- Glennon RA, Young R, Jacyno JM, Slusher M, Rosecrans JA. DOM-stimulus generalization to LSD and other hallucinogenic indo-lealkylamines. Eur J Pharmacol. 1983;86:453–459. doi: 10.1016/0014-2999(83)90196-6. [DOI] [PubMed] [Google Scholar]
- Griffiths RR, Richards WA, McCann U, Jesse R. Psilocybin can occasion mystical-type experiences having substantial and sustained personal meaning and spiritual significance. Psychopharmacology (Berl) 2006;187:268–283. doi: 10.1007/s00213-006-0457-5. discussion 284–292. [DOI] [PubMed] [Google Scholar]
- Griffiths RR, Richards WA, Johnson M, McCann U, Jesse R. Mystical-type experiences occasioned by psilocybin mediate the attribution of personal meaning and spiritual significance 14 months later. J Psychopharmacol (Oxford) 2008;22:621–632. doi: 10.1177/0269881108094300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths RR, Johnson MW, Richards WA, Richards BD, McCann U, Jesse R. Psilocybin occasioned mystical-type experiences: immediate and persisting dose-related effects. Psychopharmacology (Berl) 2011;218:649–665. doi: 10.1007/s00213-011-2358-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grob CS, Danforth AL, Chopra GS, Hagerty M, McKay CR, Halberstadt AL, Greer GR. Pilot study of psilocybin treatment for anxiety in patients with advanced-stage cancer. Arch Gen Psychiatry. 2011;68:71–78. doi: 10.1001/archgenpsychiatry.2010.116. [DOI] [PubMed] [Google Scholar]
- Grueber WB, Jan LY, Jan YN. Tiling of the Drosophila epidermis by multidendritic sensory neurons. Development. 2002;129:2867–2878. doi: 10.1242/dev.129.12.2867. [DOI] [PubMed] [Google Scholar]
- Gustavsson A, Svensson M, Jacobi F, Allgulander C, Alonso J, Beghi E, Dodel R, Ekman M, Faravelli C, Fratiglioni L, et al. CDBE 2010 Study Group. Cost of disorders of the brain in Europe 2010. Eur Neuropsychopharmacol. 2011;21:718–779. doi: 10.1016/j.euroneuro.2011.08.008. [DOI] [PubMed] [Google Scholar]
- Hayley S, Litteljohn D. Neuroplasticity and the next wave of antidepressant strategies. Front Cell Neurosci. 2013;7:218. doi: 10.3389/fncel.2013.00218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He DY, McGough NN, Ravindranathan A, Jeanblanc J, Logrip ML, Phamluong K, Janak PH, Ron D. Glial cell line-derived neurotrophic factor mediates the desirable actions of the anti-addiction drug ibogaine against alcohol consumption. J Neurosci. 2005;25:619–628. doi: 10.1523/JNEUROSCI.3959-04.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helsley S, Fiorella D, Rabin RA, Winter JC. A comparison of N,N-dimethyltryptamine, harmaline, and selected congeners in rats trained with LSD as a discriminative stimulus. Prog Neuropsychopharmacol Biol Psychiatry. 1998;22:649–663. doi: 10.1016/s0278-5846(98)00031-1. [DOI] [PubMed] [Google Scholar]
- Hoeffer CA, Klann E. mTOR signaling: at the crossroads of plasticity, memory and disease. Trends Neurosci. 2010;33:67–75. doi: 10.1016/j.tins.2009.11.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ionescu DF, Swee MB, Pavone KJ, Taylor N, Akeju O, Baer L, Nyer M, Cassano P, Mischoulon D, Alpert JE, et al. Rapid and Sustained Reductions in Current Suicidal Ideation Following Repeated Doses of Intravenous Ketamine: Secondary Analysis of an Open-Label Study. J Clin Psychiatry. 2016;77:e719–e725. doi: 10.4088/JCP.15m10056. [DOI] [PubMed] [Google Scholar]
- Izquierdo A, Wellman CL, Holmes A. Brief uncontrollable stress causes dendritic retraction in infralimbic cortex and resistance to fear extinction in mice. J Neurosci. 2006;26:5733–5738. doi: 10.1523/JNEUROSCI.0474-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jang SW, Liu X, Yepes M, Shepherd KR, Miller GW, Liu Y, Wilson WD, Xiao G, Blanchi B, Sun YE, Ye K. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc Natl Acad Sci USA. 2010;107:2687–2692. doi: 10.1073/pnas.0913572107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaworski J, Spangler S, Seeburg DP, Hoogenraad CC, Sheng M. Control of dendritic arborization by the phosphoinositide-3′-kinase-Akt-mammalian target of rapamycin pathway. J Neurosci. 2005;25:11300–11312. doi: 10.1523/JNEUROSCI.2270-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones KA, Srivastava DP, Allen JA, Strachan RT, Roth BL, Penzes P. Rapid modulation of spine morphology by the 5-HT2A serotonin receptor through kalirin-7 signaling. Proc Natl Acad Sci USA. 2009;106:19575–19580. doi: 10.1073/pnas.0905884106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly TM, Daley DC. Integrated treatment of substance use and psychiatric disorders. Soc Work Public Health. 2013;28:388–406. doi: 10.1080/19371918.2013.774673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolb B, Muhammad A. Harnessing the power of neuroplasticity for intervention. Front Hum Neurosci. 2014;8:377. doi: 10.3389/fnhum.2014.00377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krupitsky E, Burakov A, Romanova T, Dunaevsky I, Strassman R, Grinenko A. Ketamine psychotherapy for heroin addiction: immediate effects and two-year follow-up. J Subst Abuse Treat. 2002;23:273–283. doi: 10.1016/s0740-5472(02)00275-1. [DOI] [PubMed] [Google Scholar]
- Krystal JH, Tolin DF, Sanacora G, Castner SA, Williams GV, Aikins DE, Hoffman RE, D’Souza DC. Neuroplasticity as a target for the pharmacotherapy of anxiety disorders, mood disorders, and schizophrenia. Drug Discov Today. 2009;14:690–697. doi: 10.1016/j.drudis.2009.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar V, Zhang MX, Swank MW, Kunz J, Wu GY. Regulation of dendritic morphogenesis by Ras-PI3K-Akt-mTOR and Ras-MAPK signaling pathways. J Neurosci. 2005;25:11288–11299. doi: 10.1523/JNEUROSCI.2284-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kyzar EJ, Nichols CD, Gainetdinov RR, Nichols DE, Kalueff AV. Psychedelic Drugs in Biomedicine. Trends Pharmacol Sci. 2017;38:992–1005. doi: 10.1016/j.tips.2017.08.003. [DOI] [PubMed] [Google Scholar]
- Lepack AE, Fuchikami M, Dwyer JM, Banasr M, Duman RS. BDNF release is required for the behavioral actions of ketamine. Int J Neuropsychopharmacol. 2014;18:pyu033. doi: 10.1093/ijnp/pyu033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lepack AE, Bang E, Lee B, Dwyer JM, Duman RS. Fast-acting antidepressants rapidly stimulate ERK signaling and BDNF release in primary neuronal cultures. Neuropharmacology. 2016;111:242–252. doi: 10.1016/j.neuropharm.2016.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li N, Lee B, Liu RJ, Banasr M, Dwyer JM, Iwata M, Li XY, Aghajanian G, Duman RS. mTOR-dependent synapse formation underlies the rapid antidepressant effects of NMDA antagonists. Science. 2010;329:959–964. doi: 10.1126/science.1190287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Majić T, Schmidt TT, Gallinat J. Peak experiences and the afterglow phenomenon: when and how do therapeutic effects of hallucinogens depend on psychedelic experiences? J Psychopharmacol (Oxford) 2015;29:241–253. doi: 10.1177/0269881114568040. [DOI] [PubMed] [Google Scholar]
- Marinova Z, Walitza S, Grunblatt E. The hallucinogen 2,5-dime-thoxy-4-iodoamphetamine hydrochloride activates neurotrophin receptors in a neuronal cell line and promotes neurites extension. J Neural Transm (Vienna) 2017;124:749–759. doi: 10.1007/s00702-017-1706-y. [DOI] [PubMed] [Google Scholar]
- Martin DA, Marona-Lewicka D, Nichols DE, Nichols CD. Chronic LSD alters gene expression profiles in the mPFC relevant to schizophrenia. Neuropharmacology. 2014;83:1–8. doi: 10.1016/j.neuropharm.2014.03.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mathew SJ, Manji HK, Charney DS. Novel drugs and therapeutic targets for severe mood disorders. Neuropsychopharmacology. 2008;33:2080–2092. doi: 10.1038/sj.npp.1301652. [DOI] [PubMed] [Google Scholar]
- Mithoefer MC, Wagner MT, Mithoefer AT, Jerome L, Doblin R. The safety and efficacy of +/−3,4-methylenedioxymethamphetamine-assisted psychotherapy in subjects with chronic, treatment-resistant posttraumatic stress disorder: the first randomized controlled pilot study. J Psychopharmacol (Oxford) 2011;25:439–452. doi: 10.1177/0269881110378371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mithoefer MC, Wagner MT, Mithoefer AT, Jerome L, Martin SF, Yazar-Klosinski B, Michel Y, Brewerton TD, Doblin R. Durability of improvement in post-traumatic stress disorder symptoms and absence of harmful effects or drug dependency after 3,4-methylenedioxymethamphet-amine-assisted psychotherapy: a prospective long-term follow-up study. J Psychopharmacol (Oxford) 2013;27:28–39. doi: 10.1177/0269881112456611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mithoefer MC, Grob CS, Brewerton TD. Novel psychopharmacological therapies for psychiatric disorders: psilocybin and MDMA. Lancet Psychiatry. 2016;3:481–488. doi: 10.1016/S2215-0366(15)00576-3. [DOI] [PubMed] [Google Scholar]
- Moskal JR, Burgdorf JS, Stanton PK, Kroes RA, Disterhoft JF, Burch RM, Khan MA. The development of rapastinel (formerly GLYX-13); a rapid acting and long lasting antidepressant. Curr Neuropharmacol. 2017;15:47–56. doi: 10.2174/1570159X14666160321122703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murrough JW, Perez AM, Pillemer S, Stern J, Parides MK, aan het Rot M, Collins KA, Mathew SJ, Charney DS, Iosifescu DV. Rapid and longer-term antidepressant effects of repeated ketamine infusions in treatment-resistant major depression. Biol Psychiatry. 2013;74:250–256. doi: 10.1016/j.biopsych.2012.06.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nair AB, Jacob S. A simple practice guide for dose conversion between animals and human. J Basic Clin Pharm. 2016;7:27–31. doi: 10.4103/0976-0105.177703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichols DE. Hallucinogens. Pharmacol Ther. 2004;101:131–181. doi: 10.1016/j.pharmthera.2003.11.002. [DOI] [PubMed] [Google Scholar]
- Nichols DE. Psychedelics. Pharmacol Rev. 2016;68:264–355. doi: 10.1124/pr.115.011478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichols CD, Sanders-Bush E. A single dose of lysergic acid diethylamide influences gene expression patterns within the mammalian brain. Neuropsychopharmacology. 2002;26:634–642. doi: 10.1016/S0893-133X(01)00405-5. [DOI] [PubMed] [Google Scholar]
- Nichols DE, Johnson MW, Nichols CD. Psychedelics as medicines: an emerging new paradigm. Clin Pharmacol Ther. 2017;101:209–219. doi: 10.1002/cpt.557. [DOI] [PubMed] [Google Scholar]
- Nutt D, King LA, Saulsbury W, Blakemore C. Development of a rational scale to assess the harm of drugs of potential misuse. Lancet. 2007;369:1047–1053. doi: 10.1016/S0140-6736(07)60464-4. [DOI] [PubMed] [Google Scholar]
- Nutt DJ, King LA, Phillips LD Independent Scientific Committee on Drugs. Drug harms in the UK: a multicriteria decision analysis. Lancet. 2010;376:1558–1565. doi: 10.1016/S0140-6736(10)61462-6. [DOI] [PubMed] [Google Scholar]
- Oehen P, Traber R, Widmer V, Schnyder U. A randomized, controlled pilot study of MDMA (±3,4-Methylenedioxymethamphetamine)-assisted psychotherapy for treatment of resistant, chronic Post-Traumatic Stress Disorder (PTSD) J Psychopharmacol (Oxford) 2013;27:40–52. doi: 10.1177/0269881112464827. [DOI] [PubMed] [Google Scholar]
- de Osório FL, Sanches RF, Macedo LR, Santos RG, Maia-de-Oliveira JP, Wichert-Ana L, Araujo DB, Riba J, Crippa JA, Hallak JE. Antidepressant effects of a single dose of ayahuasca in patients with recurrent depression: a preliminary report. Rev Bras Psiquiatr. 2015;37:13–20. doi: 10.1590/1516-4446-2014-1496. [DOI] [PubMed] [Google Scholar]
- Peters J, Dieppa-Perea LM, Melendez LM, Quirk GJ. Induction of fear extinction with hippocampal-infralimbic BDNF. Science. 2010;328:1288–1290. doi: 10.1126/science.1186909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pittenger C, Duman RS. Stress, depression, and neuroplasticity: a convergence of mechanisms. Neuropsychopharmacology. 2008;33:88–109. doi: 10.1038/sj.npp.1301574. [DOI] [PubMed] [Google Scholar]
- Qiao H, Li MX, Xu C, Chen HB, An SC, Ma XM. Dendritic Spines in Depression: What We Learned from Animal Models. Neural Plast. 2016;2016:8056370. doi: 10.1155/2016/8056370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quirk GJ, Garcia R, González-Lima F. Prefrontal mechanisms in extinction of conditioned fear. Biol Psychiatry. 2006;60:337–343. doi: 10.1016/j.biopsych.2006.03.010. [DOI] [PubMed] [Google Scholar]
- Ristanović D, Milosević NT, Stulić V. Application of modified Sholl analysis to neuronal dendritic arborization of the cat spinal cord. J Neurosci Methods. 2006;158:212–218. doi: 10.1016/j.jneumeth.2006.05.030. [DOI] [PubMed] [Google Scholar]
- Rucker JJ, Jelen LA, Flynn S, Frowde KD, Young AH. Psychedelics in the treatment of unipolar mood disorders: a systematic review. J Psychopharmacol (Oxford) 2016;30:1220–1229. doi: 10.1177/0269881116679368. [DOI] [PubMed] [Google Scholar]
- Rush AJ, Trivedi MH, Wisniewski SR, Nierenberg AA, Stewart JW, Warden D, Niederehe G, Thase ME, Lavori PW, Lebowitz BD, et al. Acute and longer-term outcomes in depressed outpatients requiring one or several treatment steps: a STAR*D report. Am J Psychiatry. 2006;163:1905–1917. doi: 10.1176/ajp.2006.163.11.1905. [DOI] [PubMed] [Google Scholar]
- Russo SJ, Nestler EJ. The brain reward circuitry in mood disorders. Nat Rev Neurosci. 2013;14:609–625. doi: 10.1038/nrn3381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo SJ, Mazei-Robison MS, Ables JL, Nestler EJ. Neurotrophic factors and structural plasticity in addiction. Neuropharmacology. 2009;56(Suppl 1):73–82. doi: 10.1016/j.neuropharm.2008.06.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanches RF, de Lima Osório F, Dos Santos RG, Macedo LR, Maia-de-Oliveira JP, Wichert-Ana L, de Araujo DB, Riba J, Crippa JA, Hallak JE. Antidepressant effects of a single dose of ayahuasca in patients with recurrent depression: a SPECT study. J Clin Psychopharmacol. 2016;36:77–81. doi: 10.1097/JCP.0000000000000436. [DOI] [PubMed] [Google Scholar]
- Santos RG, Landeira-Fernandez J, Strassman RJ, Motta V, Cruz AP. Effects of ayahuasca on psychometric measures of anxiety, panic-like and hopelessness in Santo Daime members. J Ethnopharmacol. 2007;112:507–513. doi: 10.1016/j.jep.2007.04.012. [DOI] [PubMed] [Google Scholar]
- Smith RL, Canton H, Barrett RJ, Sanders-Bush E. Agonist properties of N,N-dimethyltryptamine at serotonin 5-HT2A and 5-HT2C receptors. Pharmacol Biochem Behav. 1998;61:323–330. doi: 10.1016/s0091-3057(98)00110-5. [DOI] [PubMed] [Google Scholar]
- Strassman RJ, Qualls CR, Uhlenhuth EH, Kellner R. Dose-response study of N,N-dimethyltryptamine in humans. II Subjective effects and preliminary results of a new rating scale. Arch Gen Psychiatry. 1994;51:98–108. doi: 10.1001/archpsyc.1994.03950020022002. [DOI] [PubMed] [Google Scholar]
- Takei N, Inamura N, Kawamura M, Namba H, Hara K, Yonezawa K, Nawa H. Brain-derived neurotrophic factor induces mammalian target of rapamycin-dependent local activation of translation machinery and protein synthesis in neuronal dendrites. J Neurosci. 2004;24:9760–9769. doi: 10.1523/JNEUROSCI.1427-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vaidya VA, Marek GJ, Aghajanian GK, Duman RS. 5-HT2A receptor-mediated regulation of brain-derived neurotrophic factor mRNA in the hippocampus and the neocortex. J Neurosci. 1997;17:2785–2795. doi: 10.1523/JNEUROSCI.17-08-02785.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Voleti B, Navarria A, Liu RJ, Banasr M, Li N, Terwilliger R, Sanacora G, Eid T, Aghajanian G, Duman RS. Scopolamine rapidly increases mammalian target of rapamycin complex 1 signaling, synaptogenesis, and antidepressant behavioral responses. Biol Psychiatry. 2013;74:742–749. doi: 10.1016/j.biopsych.2013.04.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollenweider FX, Kometer M. The neurobiology of psychedelic drugs: implications for the treatment of mood disorders. Nat Rev Neurosci. 2010;11:642–651. doi: 10.1038/nrn2884. [DOI] [PubMed] [Google Scholar]
- Wacker D, Wang S, McCorvy JD, Betz RM, Venkatakrishnan AJ, Levit A, Lansu K, Schools ZL, Che T, Nichols DE, et al. Crystal Structure of an LSD-Bound Human Serotonin Receptor. Cell. 2017;168:377–389 e12. doi: 10.1016/j.cell.2016.12.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whiteford HA, Degenhardt L, Rehm J, Baxter AJ, Ferrari AJ, Erskine HE, Charlson FJ, Norman RE, Flaxman AD, Johns N, et al. Global burden of disease attributable to mental and substance use disorders: findings from the Global Burden of Disease Study 2010. Lancet. 2013;382:1575–1586. doi: 10.1016/S0140-6736(13)61611-6. [DOI] [PubMed] [Google Scholar]
- Winter JC, Rice KC, Amorosi DJ, Rabin RA. Psilocybin-induced stimulus control in the rat. Pharmacol Biochem Behav. 2007;87:472–480. doi: 10.1016/j.pbb.2007.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang C, Shirayama Y, Zhang JC, Ren Q, Yao W, Ma M, Dong C, Hashimoto K. R-ketamine: a rapid-onset and sustained antidepressant without psychotomimetic side effects. Transl Psychiatry. 2015;5:e632. doi: 10.1038/tp.2015.136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang Y, Cui Y, Sang K, Dong Y, Ni Z, Ma S, Hu H. Ketamine blocks bursting in the lateral habenula to rapidly relieve depression. Nature. 2018;554:317–322. doi: 10.1038/nature25509. [DOI] [PubMed] [Google Scholar]
- Young MB, Andero R, Ressler KJ, Howell LL. 3,4-Methyle-nedioxymethamphetamine facilitates fear extinction learning. Transl Psychiatry. 2015;5:e634. doi: 10.1038/tp.2015.138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zanos P, Moaddel R, Morris PJ, Georgiou P, Fischell J, Elmer GI, Alkondon M, Yuan P, Pribut HJ, Singh NS, et al. NMDAR inhibition-independent antidepressant actions of ketamine metabolites. Nature. 2016;533:481–486. doi: 10.1038/nature17998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zarate CA, Jr, Singh JB, Carlson PJ, Brutsche NE, Ameli R, Luckenbaugh DA, Charney DS, Manji HK. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch Gen Psychiatry. 2006;63:856–864. doi: 10.1001/archpsyc.63.8.856. [DOI] [PubMed] [Google Scholar]
- Zarate CA, Jr, Brutsche NE, Ibrahim L, Franco-Chaves J, Diazgranados N, Cravchik A, Selter J, Marquardt CA, Liberty V, Luckenbaugh DA. Replication of ketamine’s antidepressant efficacy in bipolar depression: a randomized controlled add-on trial. Biol Psychiatry. 2012;71:939–946. doi: 10.1016/j.biopsych.2011.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng Y, Lv F, Li L, Yu H, Dong M, Fu Q. 7,8-dihydroxyflavone rescues spatial memory and synaptic plasticity in cognitively impaired aged rats. J Neurochem. 2012;122:800–811. doi: 10.1111/j.1471-4159.2012.07830.x. [DOI] [PubMed] [Google Scholar]
- Zubaran C, Shoaib M, Stolerman IP, Pablo J, Mash DC. Noribogaine generalization to the ibogaine stimulus: correlation with noribogaine concentration in rat brain. Neuropsychopharmacology. 1999;21:119–126. doi: 10.1016/S0893-133X(99)00003-2. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.