

Abstract
Substantial genetic differentiation, as great as among species, exists between populations of Drosophila melanogaster inhabiting opposite slopes of a small canyon. Previous work has shown that prezygotic sexual isolation and numerous differences in stress-related phenotypes have evolved between D. melanogaster populations in “Evolution Canyon,” Israel, in which slopes 100–400 m apart differ dramatically in aridity, solar radiation, and associated vegetation. Because the canyon's width is well within flies' dispersal capabilities, we examined genetic changes associated with local adaptation and incipient speciation in the absence of geographical isolation. Here we report remarkable genetic differentiation of microsatellites and divergence in the regulatory region of hsp70Ba which encodes the major inducible heat shock protein of Drosophila, in the two populations. Additionally, an analysis of microsatellites suggests a limited exchange of migrants and lack of recent population bottlenecks. We hypothesize that adaptation to the contrasting microclimates overwhelms gene flow and is responsible for the genetic and phenotypic divergence between the populations.
Keywords: thermotolerance‖hsp70‖P element‖genetic distance‖premating isolation
A recurrent issue in evolutionary biology is the amount of genetic isolation required for incipient species to diverge from a common ancestor, with geographical isolation and its impact on gene flow a prominent source of genetic isolation (1–3). Indeed, Mayr (4, 5) has stressed repeatedly the role of geographical separation of populations in the origin of species. Although several evolutionary mechanisms may give rise to new species in contiguous populations, in the absence of these mechanisms gene flow would seem to exert a potent homogenizing force on incipient isolates (6, 7). Nonetheless, profound adaptive radiation of species can occur without large scale geographical isolation, as in the >450 species of haplochromine cichlids from Lake Malawi (8–11). Critics reasonably may dispute that such cases are evidence for speciation without geographic separation, because in Lake Malawi (≈500-km long) and other such instances, potential spatial and ecological separation are large relative to the dispersal abilities of the speciating populations.
Accordingly, we have investigated “Evolution Canyon,” Israel, where Drosophila melanogaster occurs on north- and south-facing slopes with greatly differing climatic regimes and in the intervening region; the slopes are 400 m apart at the top and 100 m apart at the bottom (12–14). Although adult Drosophila can traverse several kilometers in a single day (15), populations on each slope have diverged in body size, heat and desiccation tolerance, oviposition thermal preference, fluctuating asymmetry, rates of mutation and recombination, and mate preference (16–18). We thus ask: Have the populations likewise diverged genetically according to microsatellite markers and a candidate gene strongly linked to resistance to environmental stress? Is this divergence consistent with genetic isolation despite the contiguity of the D. melanogaster populations in the canyon? Finally, does the divergence implicate any evolutionary mechanism(s) as its cause?
To address these questions, we analyzed both putatively neutral (microsatellites) and non-neutral (hsp70Ba, a heat shock gene) markers in D. melanogaster collected from the middle elevation of each slope. Microsatellites, abundant repetitive DNA sequences with motifs 2–6-nucleotides long, are ideal tools for the study of population history. In particular, they can elucidate migration rates between populations and detect recent population growth or decline (19–23). In D. melanogaster, hsp70Ba is one of five genes encoding Hsp70, a heat-inducible molecular chaperone that plays a crucial role in inducible thermotolerance and resistance to other stresses (24). Because thermotolerance is related to Hsp70 levels, nucleotide variation affecting Hsp70 levels can be a target of selection. Such variation potentially occurs in the hsp70Ba promoter, where D. melanogaster from Evolution Canyon are polymorphic for a 1.2-kb P element insertion that interrupts a regulatory region essential for high hsp70 transcription. We report remarkable differences in the frequencies of both microsatellites and the P element-bearing hsp70Ba allele (henceforth hsp70BaP) between D. melanogaster inhabiting the two slopes <400 m apart. These differences are consistent with local adaptation to contrasting microclimates, overwhelming gene flow between the adjacent habitats.
Methods
Flies.
Flies were collected from the two midslope stations 90 m above sea level on the opposite slopes of Evolution Canyon (Lower Nahal Oren, Mt. Carmel, Israel) during August-September, 1997. First, isofemale lines were established from the samples with 25 lines per slope. Next, 10 females and 10 males of each isofemale line were combined in population cages to construct north-facing slope (NFS) and south-facing slope (SFS) synthetic populations. The populations were maintained as mass cultures with random mating for 50–55 generations on standard cornmeal-sugar-yeast-agar medium in half-pint bottles at 24 ± 1°C and on a 12/12 light/dark cycle.
Microsatellite Detection.
We sampled 39 females from NFS and 39 females from SFS. DNA extracts were prepared with a standard proteinase K protocol according to Gloor et al. (25). PCR primers were designed for 15 loci according to Schug et al. (26). Primer sequences and amplification conditions are available also at http://www.mbg.cornell.edu/aquadro/microsatellite.html. Primers were labeled with a carbocyanine dye (Cy-5) on the 5′ end. Each 20-μl reaction mixture contained 2 μg of template DNA, 100 pmol/μl of each primer, 1.5–2.5 mM MgCl2, 2 mM dNTPs, 2 μl of magnesium-free buffer and 0.1 μl of 500U Taq polymerase (Sigma). The thermocycling profile was 45 cycles of 94°C for 30 s, 52–60°C for 75 s, and 72°C for 6 min. PCR products were visualized by electrophoresis on denaturing sequencing gels with an ALFexpress II DNA automated sequencer and protocols of APBiotech (27). Samples were loaded with an internal lane standard, usually PstI-cut lambda, and labeled with ROX, a red fluorescent dye. Fragment sizes were determined with ALFWIN FRAGMENT ANALYZER 1.03 software (APBiotech).
hsp70BaP Screening.
Single-fly DNA preparations were prepared from individual flies as described above. The hsp70Ba promoter region was amplified from individual flies by adding 2 μl of template DNA to buffer [10 mM Tris⋅HCl (pH 9.0)/50 mM KCl/0.1% Triton X-100] with 1.5–3.0 mM MgCl2, 0.2 mM each dNTP, 5 pmol of each primer, and 1.25 units of Taq DNA polymerase (Promega) per 25-μl reaction. The primers were: upper, 5′-GCAAGCAATCATCATCCAAT-3′, and lower, 5′-ACTGTGTTTCTGGGGTTCAT-3′. These primers flank a polymorphic P element insertion site. To determine P element frequency, this amplification was performed on individual males and females in conjunction with an additional PCR that contained the same lower primer as described above and a P element-specific upper primer (5′-GCCTTCTTTTATCTTTTCTGG-3′). For PCR amplification of hsp70Ba promoters to be cloned and sequenced, 1 μl of template DNA from individuals was added to buffer [50 mM KCl/50 mM Tris⋅HCl (pH 8.3)] with 1.5 mM MgCl2, 0.2 mM of each dNTP, 5 pmol of each primer (flanking primers as described above), and 2.5 units MasterAmp Extra Long DNA polymerase mix (Epicentre Technologies, Madison, WI) per 100-μl reaction. Reaction conditions for all PCRs consisted of an initial 1.5 min at 94°C, 35 cycles of 92°C for 1 min, 54°C for 1 min, and 72°C for 2.5 min and a final step at 72°C for 5 min.
For cloning and sequencing, PCR products were amplified from single-fly DNA template, cleaned with Qiagen spin columns, resuspended in water, and cloned into the pGEM-T Easy vector (Promega) according to manufacturer recommendations. Individual plasmid clones were prepared with Qiagen Miniprep spin columns, suspended in water, and sequenced. Sequencing reactions were performed with ABI Prism cycle sequencing kits (Perkin–Elmer) according to manufacturer instructions. All samples were sequenced by using primers based on vector sequence (pUC/M13 forward and reverse). Internal sequencing primers (available on request) were used to provide complete coverage. Sequencing was conducted on an ABI 377 sequencer (Perkin–Elmer).
Statistical Analysis of Microsatellites.
Each locus was examined for gene diversity, heterozygosity, and genetic differentiation measured by Wright's F statistics (FST; ref. 28), by using POPGENE (http://www.ualberta.ca/∼fyeh/index.htm) and GENEPOP (29). The significance of FST values was assessed with allelic permutations generated by fstat (5,000 resamplings; ref. 30). Because no direct sequence data for microsatellites were available in this study, we determined approximate (maximal) repeat numbers for each allele by dividing the PCR product size by the number of base pairs in a single repeat (according to ref. 26). We investigated the relationship between (i) gene diversity, (ii) heterozygosity, (iii) variance of the maximal repeat number, and (iv) number of alleles as dependent variables and (i) slope, (ii) marker chromosomal location, and (iii) recombination rate per chromosome physical unit length as independent variables. Whenever feasible, P values from exact as opposed to asymptotic or Monte Carlo methods are reported (31). These were computed mainly with statxact 4 (Cytel; http://www.cytel.com; ref. 32).
Because a bottleneck in one or both populations can affect the estimation of genetic distance (33), the program BOTTLENECK (http://www.ensam.inra.fr/URLB/bottleneck/pub.html; refs. 34 and 35) was used to assess whether studied populations were subject to a demographic decline. This program detects a deficiency in the number of alleles observed given the expected heterozygosity and the principle that under mutation-drift equilibrium allelic diversity is reduced faster than heterozygosity (36). Three significance tests (sign test, standardized differences test, and Wilcoxon sign-rank test) and three mutation models (infinite allele, two-phased, and stepwise mutation) were implemented.
To obtain maximum likelihood estimates of migration rates between slopes and effective population sizes of NFS and SFS, we analyzed a coalescence/Monte Carlo Markov Chain process, generated by MIGRATE (http://evolution.genetics.washington.edu/lamarc/migrate.html; refs. 22, 37) and run on SunOS 5.6. The estimation process uses an expansion of the coalescent theory that includes migration and Metropolis–Hastings importance sampling. The following assumptions are made: populations have constant sizes through time such that they do not grow or decline, and the mutation rate μ is constant (but may vary across loci according to a gamma distribution). Additionally, suppositions of the coalescence theory (sensu Kingman, ref. 38) need to be valid. The parameters P to estimate using genealogies G with data D were θ = 4Neμ and γ = 4Nem, where Ne is the effective population size and m is migration rate per generation. The likelihood formula is based on the product between the genealogy likelihood (D|G) and the prior probability of the coalescent genealogy
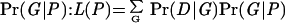 |
(ref. 22). We set the Markov Chain Monte Carlo parameters as follows: 10 short chains with 1,000 genealogies and 2 long chains with 10,000 genealogies sampled and burn-in of 10,000. Beerli and Felsenstein (22) found that these numbers are sufficient to produce a convergence to a value independent of the initial parameter values.
Results
Microsatellites.
Despite a small number of total alleles and low heterozygosity, many of the alleles in both the NFS and SFS populations were private (Tables 1 and 2). Thus, because other allele frequencies were also very different, Nei's (39) genetic distance (D = 0.566) and the FST statistic of genetic differentiation between slopes are remarkably high (Table 3). The two populations did not differ significantly in gene diversity (HST), heterozygosity (H), variance in the maximal repeat number (Var), or number of alleles (exact Mann–Whitney test, P > 0.05 in each case). Microsatellite variability depended on chromosome location (Spearman rank correlation = −0.41, P < 0.025 for HST and distance (in cM) to centromere, and Spearman rank correlation = −0.32, P < 0.1 for H and distance). The number of alleles was correlated (Rs = 0.45, P < 0.02) with recombination rate per DNA unit length (40). Although DMHSP82 (chromosome III, 63B9) was the most variable locus, on average chromosome III did not vary more than chromosomes X and II. Of the loci analyzed, DMPROSPER (III 86E1) is located nearest the hsp70 clusters (III 87A7, 87C1), but its allelic frequencies did not deviate significantly from the average.
Table 1.
Numbers of private alleles in two populations of D. melanogaster from the opposite slopes of the Evolution Canyon
| Population | Frequency, f
|
Total | ||
|---|---|---|---|---|
| <0.05 | 0.05 ≤ f ≤ 0.5 | >0.5 | ||
| NFS | 2 | 7 | 2 | 11 |
| SFS | 7 | 5 | 5 | 17 |
Three arbitrary categories based on their frequencies (f) are distinguished.
Table 2.
Measures of genetic variation in D. melanogaster from Evolution Canyon by chromosome for 15 microsatellite loci: gene diversity (Hst), observed heterozygosity (H), variance in maximal repeat number (Var), and the number of alleles (the repeat sequence and cytological location according to ref. 26)
| Chromosome | Locus | Repeat | Cyt. location | d*, cM | r† |
Hst
|
H
|
Var
|
No. of alleles
|
||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NSF | SFS | NFS | SFS | NFS | SFS | NFS | SFS | ||||||
| X | DELTEX | (AGTT)6 | 6A1-2 | 49 | 3.83 | 0.214 | 0.646 | 0.231 | 0.526 | 2.333 | 6.667 | 3 | 4 |
| DROACS2 | (CT)7 | 1B1-5 | 66 | 0.00 | 0.138 | 0.233 | 0.147 | 0.265 | 2.000 | 18.00 | 2 | 2 | |
| DROSEV2 | (TTG)9 | 10A1-2 | 33 | 3.44 | 0.401 | 0.187 | 0.371 | 0.133 | 2.333 | 4.333 | 3 | 3 | |
| II | DROEXPAND | (CAG)8 | 21C3 | 55 | 1.63 | 0.356 | 0.626 | 0.333 | 0.333 | 1.000 | 1.000 | 3 | 3 |
| DROGPAD | (GT)19 | 47A | 5 | 1.60 | 0.527 | 0.288 | 0.454 | 0.333 | 0.500 | 8.000 | 2 | 2 | |
| DROGPDHA | (CT)7 | 25F5-6A | 37 | 4.95 | 0.483 | 0.291 | 0.600 | 0.226 | 2.917 | 1.667 | 4 | 4 | |
| ELF1 | (CAG)6 | 54F | 32 | 3.41 | 0.716 | 0.723 | 0.607 | 0.484 | 1.667 | 2.500 | 4 | 5 | |
| ODD | (CAG)4 | 24A | 47 | 4.22 | 0.366 | 0.299 | 0.061 | 0.083 | 1.000 | 1.000 | 3 | 3 | |
| TWI | (CAG)3 | 59C-D | 47 | 2.71 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 1 | 1 | |
| III | DMCATHPO | (ACC)6 | 75D-E1 | 9 | 0.48 | 0.528 | 0.719 | 0.297 | 0.500 | 1.000 | 1.667 | 3 | 4 |
| DMHSP82 | (TA)5 | 63B9 | 41 | 4.29 | 0.767 | 0.833 | 0.650 | 0.389 | 43.23 | 49.611 | 7 | 9 | |
| DMPROSPER | (GA)12 | 86E1 | 4 | 1.35 | 0.606 | 0.518 | 0.136 | 0.350 | 2.333 | 2.333 | 3 | 3 | |
| DMSIDNA | (GC)6 | 96E1-4 | 54 | 3.88 | 0.593 | 0.000 | 0.250 | 0.000 | 2.500 | 0.000 | 5 | 1 | |
| DMU1951 | (TA)16 | 93C | 36 | 3.53 | 0.608 | 0.774 | 0.067 | 0.243 | 1.667 | 11.333 | 4 | 7 | |
| DROLMALK | (CAA)6 | 98 | 56 | 3.81 | 0.316 | 0.106 | 0.270 | 0.111 | 1.000 | 0.500 | 3 | 2 | |
d, distance to centromere.
r, recombination rate per DNA unit length (40).
Table 3.
Estimates of FST values and results of tests for interslope differentiation in D. melanogaster from Evolution Canyon (the assumption of random mating within populations is relaxed)
| Chromosome | Locus | FST | P* |
|---|---|---|---|
| X | DELTEX | 0.516 | <0.0001 |
| DROACS2 | 0.789 | <0.0001 | |
| DROSEV2 | 0.691 | <0.0001 | |
| II | DROEXPAND | 0.116 | 0.0014 |
| DROGPAD | 0.305 | <0.0001 | |
| DROGPDHA | 0.629 | <0.0001 | |
| ELF1 | 0.036 | 0.0029 | |
| ODD | −0.026 | >0.5 | |
| TWI | −0.066 | >0.5 | |
| III | DMCATHPHO | 0.075 | <0.0001 |
| DMHSP82 | 0.012 | >0.5 | |
| DMPROSPER | 0.282 | <0.0001 | |
| DMSIDNA | 0.197 | <0.0002 | |
| DMU1951 | 0.064 | <0.0001 | |
| DROLMALK | 0.731 | <0.0001 | |
| Across all loci | 0.365† | <0.0001 |
The P values refer to the exact G test (ref. 30).
Jacknifed value; its 95% confidence interval (bootstrapping over loci) is equal to (0.193, 0.513).
We used the BOTTLENECK program (34, 35) to test for a potential effective population size reduction that may have occurred either during the colonization of the canyon or due to unfavorable weather. None of the three mutation models produced results unequivocally suggesting bottlenecks in either population in the recent past (Table 4).
Table 4.
Tests for Cornuet and Luikart's (34) bottleneck effects under three models of microsatellite evolution
| Population | Exp. H exc. | H exc. | H def | Probability
|
Interpretation | ||
|---|---|---|---|---|---|---|---|
| Sign test | SD test | Wilcoxon | |||||
| Infinite allele model (see ref. 36) | |||||||
| NFS | 7.46 | 9 | 5 | 0.289 | 0.07 | 0.119 | No deviation |
| SFS | 7.37 | 9 | 5 | 0.273 | 0.04 | 0.241 | No deviation |
| Two-phased model of mutation | |||||||
| NFS | 7.82 | 6 | 8 | 0.237 | 0.425 | 0.951 | No deviation |
| SFS | 7.83 | 8 | 6 | 0.575 | 0.372 | 0.801 | No deviation |
| Stepwise mutation model | |||||||
| NFS | 8.15 | 5 | 9 | 0.080 | 0.060 | 0.193 | No deviation |
| SFS | 7.86 | 5 | 9 | 0.101 | 0.040 | 0.241 | No deviation |
The table includes: Expected number of loci with excess heterozygosity (Exp. H exc.), observed number of loci exhibiting heterozygosity excess (H exc.), observed number of loci with heterozygosity deficiency (H def.), and the probability values for each test. The number of loci expected to have an excess of heterozygosity depends on the number of chromosomes sampled, the number of alleles observed, and heterozygosity expected from the observed allele frequencies under the assumption that the populations are at the mutation drift equilibrium and that all loci mutate according to the model specified. Probabilities from three tests [sign test, standard differences (SD) test, and Wilcoxon test] were calculated using the bottleneck program.
Estimated 0.95 HPD intervals of the parameter θ are NFS (0.782 < θ < 0.921) and SFS (0.818 < θ < 0.943). For the migration rate, 4Nem, the 0.95 HPD intervals are: from NFS into SFS (1.620 < γ < 2.249) and from SFS into NFS (1.252 < γ < 1.818). For a comparison, the average gene flow M estimated directly from FST, M = (1/FST − 1)/4 (41), was 0.835.
hsp70BaP Frequency.
Sequencing of the highly conserved transcribed region of the hsp70Ba gene in the Evolution Canyon lines expectedly revealed little polymorphism (not presented here, but see GenBank accession nos. AF 385405–385408). By contrast, the lines were polymorphic for a 1,222-bp insertion in the hsp70Ba promoter. Sequencing identified this insertion as a nonautonomous P element at position −184 relative to transcription start (GenBank accession no. AF377341). This P element intervenes between the second and third of the four heat shock elements (HSEs) located in the hsp70Ba promoter. Ninety-six individuals from NFS included five hsp70BaP homozygotes and 55 heterozygotes (an allelic frequency of 33.9%), and 124 SFS individuals included three heterozygotes (a frequency of 1.2%). This interslope difference in genotype frequencies is highly significant (P < 0.0001 by using exact permutation test).
Discussion
Interslope Genetic Divergence.
Our assay of microsatellite variability revealed striking genetic differentiation between populations derived from the opposite canyon slopes, with genetic distance (D = 0.566) as large as that between sibling Drosophila species (allozyme data, refs. 42–44). For example, D values obtained from allozyme data and ranging from 0.5 to 0.6 have been recorded for the D. melanogaster-Drosophila simulans species pair and Drosophila willistoni group [summarized by Coyne and Orr (43, 44)]. Clearly, comparisons between microsatellite- and allozyme-based distances should be taken with caution because of the higher microsatellite mutation rate (45). Nonetheless, to our knowledge this is the highest intraspecies genetic distance ever reported for Drosophila, approached only by a D of 0.44 between D. melanogaster populations 15 km apart from one another in the Brazzaville area of the Congo (46).
Similarly, the FST value for NFS and SFS populations was also high in comparison with other reports (Table 5), even when the relatively large standard error of the estimate is considered. If microsatellite size homoplasy, which may occur both among and within species, is taken into account, even higher coefficients of genetic divergence are expected (49, 50). Numerous private alleles and several near-private alleles in both populations strongly affect the D and FST values. Although other genetic distance measures have been developed specifically for microsatellite data, e.g., RST or δμ2 (33, 51, 52), we chose to use D and FST for two reasons: (i) the ease of comparing our results with others' and (ii) the behavior of other measures is not obvious when the assumptions of the stepwise mutation model are violated. Indeed, microsatellites may follow a complex nonstepwise mutation pattern (49).
Table 5.
Comparison of microsatellite differentiation for various populations of D. melanogaster, D. simulans, and D. pseudoobscura.
Chromosome Ecology.
In our data, intraslope genetic variation (HST) decreased with marker distance from the centromere, a strong suppressor of recombination. This may reflect a negative association between allelic variation and recombination, which is contrary to the trend in Drosophila (and other taxa) and potentially caused by background selection and selective sweeping (53). The association between allele number and distance from the centromere was negative but not significant (data not shown), whereas the correlation between allele number and recombination rate (40) was positive and significant at P < 0.02. This inconsistency resists simple explanation, but could be caused by high variation in recombination rate (54, 55) or background/sweeping selection and diversifying/balanced selection interacting with recombination in opposite ways (54, 56, 57).
hsp70Ba.
The hsp70BaP allele was 28 times more frequent in NFS than in SFS. These large differences in microsatellite and hsp70BaP frequencies are consistent with low but nonzero gene flow between NFS and SFS populations. Estimated migrants from NFS into SFS and from SFS into NFS do not exceed three and two individuals per generation, respectively. Calculations of migration rate from microsatellite frequencies likely underestimate actual migration, because genetic methods describe only migration events resulting in gene pool changes. Whatever degree of sympatry Drosophila populations in Evolution Canyon represent, even a largely reduced gene flow is expected eventually to homogenize allele frequencies between populations (6–7). Yet, the interslope genetic distance we report is equivalent to those from allozyme data associated with complete reproductive isolation (43, 44). We hypothesize that divergent natural selection acting in the two microclimates overwhelmed the homogenizing effect of migration with a subsequent shift in mating preferences (18, 58), in turn yielding a correlated differentiation of genetic markers. Indeed, reproductive isolation is a frequent byproduct of adaptive response to different selection regimes (ref. 59 and references therein). Because reproductive isolation is an essential component of incipient speciation within the range of a common ancestor (60), we emphasize that Drosophila in Evolution Canyon prefer mates native to their own slope; although the different lines from the same slope did not deviate more than 1.3% from random mating, the deviation between lines from opposite slopes exceeded 15% (18, 58).
The between slope difference in hsp70BaP frequency is consistent with differential thermal selection. During and after heat shock, the heat shock transcription factor (HSF) trimerizes and binds to HSEs in the hsp70 promoter to release the paused transcriptional apparatus and induce transcription (61). Although HSEs 1 and 2 are sufficient for some transcriptional activation, the additional HSEs increase transcription. The P element insertion disrupts the hsp70Ba promoter, intervening between the proximal and distal HSE pairs, and may therefore alter cooperative binding and transcription. Indeed, strains with the P element insertion in the hsp70Ba promoter have lower inducible thermotolerance and lower Hsp70 protein levels than strains lacking the insertion (unpublished data). Thus, the difference in hsp70BaP allele frequency may result from stronger selection against the insertion in the SFS.
Although neither our data nor prior work suggest such alternative explanations for the divergence in genetic markers, the divergence could have arisen via (re)colonization of the canyon by individuals from genetically distant ancestral populations, perhaps accompanied by a habitat preference, genetic drift, and/or evolution of the synthetic populations while in laboratory culture. Microsatellite frequencies indicate no bottlenecks, suggesting that genetic drift has not have played a major role in the diversification. Laboratory evolution seems unlikely because microsatellite frequencies in D. melanogaster change slowly in laboratory culture due to relatively low mutation rates (62, 63), and hsp70BaP allele frequency did not change with time.
Our findings and hypotheses on the genetic and phenotypic divergence in D. melanogaster are consistent also with patterns both for other drosophilids and unrelated species in Evolution Canyon. Another drosophilid, D. simulans varies in thermotolerance in the same way as does D. melanogaster (17). In addition, their allozyme diversity indices significantly differ across the canyon (14). As with the P element in the hsp70Ba promoter, the frequency of a BARE-1 retrotransposon in the wild barley (Hordeum spontaneum; ref. 64) varies with the height and dryness of the slope on both slopes but especially on the drier SFS. In fact, a 3-fold difference in BARE-1 abundance significantly affects genome and cell size, thereby presumably leading to adaptive changes in the plants' growth rates. Recently, Nevo has hypothesized that incipient sympatric speciation may be ongoing in a soil fungus (Sordaria fimicola) and other taxa (Nostoc, Lotus, etc.) inhabiting Evolution Canyon (see ref. 65 and references therein). The D. melanogaster case presented here, however, is unique not only in detecting massive fine scale genetic differentiation in a very mobile organism but in revealing genetic changes associated with incipient sympatric or near sympatric speciation driven by adaptation to local environments.
Acknowledgments
We thank Jerry A. Coyne and the four reviewers for constructive criticism, Kostas Iliadi and Eugenia Rashkovetsky for help during preparation of this manuscript, Katarzyna Michalak for PCR, Peter Beerli for computing, and Tomáš Pavlic̨ek for field assistance. We acknowledge Margaret G. Kidwell, Francisco J. Ayala, Hampton L. Carson, and James F. Crow for constructive comments that improved the paper. The work was supported by Israel Science Foundation Grant 02198, United States-Israel Binational Science Foundation Grant 4556, National Science Foundation Grants DGE-0072944, IBN99-72678, and 99-86158, Foundation for Polish Science Grant KBN 6 PO4F 011 16, and the Ancell-Teicher Research Foundation for Molecular Genetics and Evolution.
Abbreviations
- NFS
north-facing slope
- SFS
south-facing slope
- HSE
heat shock element
References
- 1.Kondrashov A S. Theor Popul Biol. 1986;29:1–15. doi: 10.1016/0040-5809(86)90002-x. [DOI] [PubMed] [Google Scholar]
- 2.Coyne J A, Orr H A. Philos Trans R Soc London B. 1998;28:287–305. doi: 10.1098/rstb.1998.0210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kondrashov A S, Kondrashov F A. Nature (London) 1999;6742:351–354. doi: 10.1038/22514. [DOI] [PubMed] [Google Scholar]
- 4.Mayr E. Animal Species and Evolution. Cambridge, MA: Harvard Univ. Press; 1963. [Google Scholar]
- 5.Mayr E. In: Mechanisms of Speciation. Barigozzi C, editor. New York: Liss; 1982. pp. 1–19. [Google Scholar]
- 6.Crow J F, Kimura M. An Introduction to Population Genetics Theory. New York: Harper & Row; 1970. [Google Scholar]
- 7.King R B. Evolution (Lawrence, Kans) 1993;47:1819–1833. doi: 10.1111/j.1558-5646.1993.tb01272.x. [DOI] [PubMed] [Google Scholar]
- 8.Meyer A. Trends Ecol Evol. 1993;8:279–284. doi: 10.1016/0169-5347(93)90255-N. [DOI] [PubMed] [Google Scholar]
- 9.van Oppen M H L, Duetsch J C, Turner G F, Rico C, Ibrahim K M, Robinson R L, Hewitt G M. Proc R Soc London Ser B. 1997;264:1803–1812. [Google Scholar]
- 10.Markert J A, Arnegard M E, Danley P D, Kocher T D. Mol Ecol. 1999;8:1013–1026. [Google Scholar]
- 11.Danley P D, Markert J A, Arnegard M E, Kocher T D. Evolution (Lawrence, Kans) 2000;54:1725–1737. doi: 10.1111/j.0014-3820.2000.tb00716.x. [DOI] [PubMed] [Google Scholar]
- 12.Nevo E. Proc R Soc London Ser B. 1995;262:149–155. [Google Scholar]
- 13.Nevo E. Theor Popul Biol. 1997;52:231–243. doi: 10.1006/tpbi.1997.1330. [DOI] [PubMed] [Google Scholar]
- 14.Harry M, Rashkovetsky E, Pavlicek T, Baker S, Derzhavets E M, Capy P, Cariou M-L, Lachaise D, Asada N, Nevo E. Biologia (Bratisl) 1999;54:685–705. [Google Scholar]
- 15.Coyne J A, Milstead B. Am Nat. 1987;130:70–82. [Google Scholar]
- 16.Rashkovetsky E, Korol A B, Pavlicek T, Nevo E. Drosoph Inf Serv. 1997;80:83–85. [Google Scholar]
- 17.Nevo E, Rashkovetsky E, Pavlicek T, Korol A. Heredity. 1998;80:9–16. doi: 10.1046/j.1365-2540.1998.00274.x. [DOI] [PubMed] [Google Scholar]
- 18.Korol A, Rashkovetsky E, Iliadi K, Michalak P, Ronin Y, Nevo E. Proc Natl Acad Sci USA. 2000;97:12637–12642. doi: 10.1073/pnas.220041397. . (First Published October 24, 2000; 10.1073/pnas.220041397) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Kimmel M R, Chakraborty R, King J P, Bamshad M, Watkins W S, Watkins W S, Jorde L B. Genetics. 1998;148:1921–1930. doi: 10.1093/genetics/148.4.1921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Wilson I J, Balding D J. Genetics. 1998;150:499–510. doi: 10.1093/genetics/150.1.499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Beaumont M A. Genetics. 1999;153:2013–2029. doi: 10.1093/genetics/153.4.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Beerli P, Felsenstein J. Genetics. 1999;152:763–773. doi: 10.1093/genetics/152.2.763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Gonser R, Donnelly P, Nicholson G, Di Rienzo A. Genetics. 2000;154:1793–1807. doi: 10.1093/genetics/154.4.1793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Feder M E, Hofmann G E. Annu Rev Physiol. 1999;61:243–282. doi: 10.1146/annurev.physiol.61.1.243. [DOI] [PubMed] [Google Scholar]
- 25.Gloor G B, Preston C R, Johnson-Schlitz D M, Nassif N A, Phillips R W, Benz W K, Robertson H M, Engels W R. Genetics. 1993;135:81–95. doi: 10.1093/genetics/135.1.81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Schug M D, Wetterstrand K A, Gaudette M S, Lim R H, Hutter C M. Mol Ecol. 1998;7:57–70. doi: 10.1046/j.1365-294x.1998.00304.x. [DOI] [PubMed] [Google Scholar]
- 27.Amersham Pharmacia. alfwintm fragment analyser 1.03, User Manual. UK, Uppsala, Sweden: Amersham Pharmacia; 1998. [Google Scholar]
- 28.Weir B S, Cockerham C C. Evolution (Lawrence, Kans) 1984;38:1358–1370. doi: 10.1111/j.1558-5646.1984.tb05657.x. [DOI] [PubMed] [Google Scholar]
- 29.Raymond M, Rousset F. J Hered. 1995;86:248–249. [Google Scholar]
- 30.Goudet J. J Hered. 1995;86:485–486. [Google Scholar]
- 31.Weerahandi S. Exact Statistical Methods for Data Analysis. New York: Springer; 1995. [Google Scholar]
- 32.Cytel software Corp. statxact, User Manual. MA: Cambridge; 2000. [Google Scholar]
- 33.Hedrick P. Evolution (Lawrence, Kans) 1999;53:313–318. [Google Scholar]
- 34.Cornuet J M, Luikart G. Genetics. 1996;144:2001–2014. doi: 10.1093/genetics/144.4.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Piry S G, Luikart G, Cornuet J M. bottleneck, A Program for Detecting Recent Effective Population Size Reductions from Allele Data Frequencies. Montpellier, France: INRA; 1996. [Google Scholar]
- 36.Maruyama T, Fuerst P A. Genetics. 1985;111:675–689. doi: 10.1093/genetics/111.3.675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Beerli P. migrate, Documentation and Program, part of lamarc 0.9. Seattle: University of Washington; 1997–2000. [Google Scholar]
- 38.Kingman J. Stochastic Proc Appl. 1982;13:235–248. [Google Scholar]
- 39.Nei M. Genetics. 1978;89:583–590. doi: 10.1093/genetics/89.3.583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Kliman R M, Hey J. Mol Biol Evol. 1993;10:1239–1258. doi: 10.1093/oxfordjournals.molbev.a040074. [DOI] [PubMed] [Google Scholar]
- 41.Slatkin M. Evolution (Lawrence, Kans) 1993;47:264–279. doi: 10.1111/j.1558-5646.1993.tb01215.x. [DOI] [PubMed] [Google Scholar]
- 42.Ayala F J. Evol Biol. 1975;8:1–78. [Google Scholar]
- 43.Coyne J A, Orr H A. Evolution (Lawrence, Kans) 1989;43:362–381. doi: 10.1111/j.1558-5646.1989.tb04233.x. [DOI] [PubMed] [Google Scholar]
- 44.Coyne J A, Orr H A. Evolution (Lawrence, Kans) 1997;51:295–303. doi: 10.1111/j.1558-5646.1997.tb02412.x. [DOI] [PubMed] [Google Scholar]
- 45.Balloux F, Brünner H, Lugon-Moulin N, Hausser J, Goudet J. Evolution (Lawrence, Kans) 2000;54:1414–1422. doi: 10.1111/j.0014-3820.2000.tb00573.x. [DOI] [PubMed] [Google Scholar]
- 46.Capy P, Veuille M, Paillette M, Jallon J-M, Vouidibio J, David J R. Heredity. 2000;84:468–475. doi: 10.1046/j.1365-2540.2000.00711.x. [DOI] [PubMed] [Google Scholar]
- 47.Irvin S D, Wetterstrand K A, Hutter C M, Aquadro C F. Genetics. 1998;150:777–790. doi: 10.1093/genetics/150.2.777. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Noor M A F, Schug M D, Aquadro C F. Genet Res. 2000;75:25–35. doi: 10.1017/s0016672399004024. [DOI] [PubMed] [Google Scholar]
- 49.Colson I, Goldstein D B. Genetics. 1999;152:617–627. doi: 10.1093/genetics/152.2.617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.van Oppen M J H, Rico C, Turner G F, Hewitt G M. Mol Biol Evol. 2000;17:489–498. doi: 10.1093/oxfordjournals.molbev.a026329. [DOI] [PubMed] [Google Scholar]
- 51.Slatkin M. Genetics. 1995;139:457–462. doi: 10.1093/genetics/139.1.457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Goldstein D B, Linares A R, Cavalli-Sforza L, Feldman M V. Proc Natl Acad Sci USA. 1995;92:11549–11552. [Google Scholar]
- 53.Charlesworth B. Genet Res. 1996;68:131–149. doi: 10.1017/s0016672300034029. [DOI] [PubMed] [Google Scholar]
- 54.Korol A B, Preygel I A, Preygel S I. Recombination Variability and Evolution. London: Chapman & Hall; 1994. [Google Scholar]
- 55.Andolfatto P, Przeworski M. Genetics. 2000;156:257–268. doi: 10.1093/genetics/156.1.257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Stephan W, Xing L, Kirby D A, Braverman J M. Proc Natl Acad Sci USA. 1998;95:5649–5654. doi: 10.1073/pnas.95.10.5649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kelly J K, Wade M J. J Theor Biol. 2000;204:83–101. doi: 10.1006/jtbi.2000.2003. [DOI] [PubMed] [Google Scholar]
- 58.Iliadi, K., Iliadi, N., Rashkovetsky, E., Minkov, I., Nevo, E. & Korol, A. (2001) Proc. R. Soc. London Ser. B, in press. [DOI] [PMC free article] [PubMed]
- 59.Powell J R. Progress and Prospects in Evolutionary Biology: The Drosophila Model. London: Oxford Univ. Press; 1997. [Google Scholar]
- 60.Howard D J, Marshall J L, Braswell W E, Coyne J A. Science. 2001;291:1853. doi: 10.1126/science.291.5510.1853a. [DOI] [PubMed] [Google Scholar]
- 61.Fernandes M, O'Brien T, Lis J T. In: The Biology of Heat Shock Proteins and Molecular Chaperons. Morimoto R I, Tissieres A, Georgopoulos C, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1994. [Google Scholar]
- 62.Schug M D, Mackay T F C, Aquadro C F. Nat Genet. 1997;15:99–102. doi: 10.1038/ng0197-99. [DOI] [PubMed] [Google Scholar]
- 63.Schug M D, Hutter C M, Noor M A F, Aquadro C F. Genetica. 1998;103:359–367. [PubMed] [Google Scholar]
- 64.Kalender R, Tanskanen J, Immonen S, Nevo E, Schulman A H. Proc Natl Acad Sci USA. 2000;97:6603–6607. doi: 10.1073/pnas.110587497. . (First Published May 23, 2000; 10.1073/pnas.110587497) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Nevo E. Proc Natl Acad Sci USA. 2001;98:6233–6240. doi: 10.1073/pnas.101109298. [DOI] [PMC free article] [PubMed] [Google Scholar]