

Summary
Plant molecular farming offers a cost‐effective and scalable approach to the expression of recombinant proteins which has been proposed as an alternative to conventional production platforms for developing countries. In recent years, numerous proofs of concept have established that plants can produce biologically active recombinant proteins and immunologically relevant vaccine antigens that are comparable to those made in conventional expression systems. Driving many of these advances is the remarkable plasticity of the plant proteome which enables extensive engineering of the host cell, as well as the development of improved expression vectors facilitating higher levels of protein production. To date, the only plant‐derived viral glycoprotein to be tested in humans is the influenza haemagglutinin which expresses at ~50 mg/kg. However, many other viral glycoproteins that have potential as vaccine immunogens only accumulate at low levels in planta. A critical consideration for the production of many of these proteins in heterologous expression systems is the complexity of post‐translational modifications, such as control of folding, glycosylation and disulphide bridging, which is required to reproduce the native glycoprotein structure. In this review, we will address potential shortcomings of plant expression systems and discuss strategies to optimally exploit the technology for the production of immunologically relevant and structurally authentic glycoproteins for use as vaccine immunogens.
Keywords: glycoprotein, biopharming, vaccine, chaperone
Introduction
The development of plant‐based expression systems for the scalable and economical production of recombinant proteins constitutes a major paradigm shift in the manufacturing of pharmaceuticals. This technology is fast becoming established in several niche areas where traditional manufacturing approaches fall short—such as where a rapid response is required, or where production scalability is limited (Rybicki, 2009; Stoger et al., 2014; Streatfield et al., 2015). A major driving force for molecular farming is the lower costs involved; this is for both the production of biomass and the less stringent infrastructure requirements than for typical fermentation systems (Edgue et al., 2017; Rybicki, 2009; Tschofen et al., 2016). These unique features are particularly appealing for developing countries which typically suffer a greater burden of infectious disease, and where the infrastructure for traditional pharmaceutical production is often largely or completely absent (Hefferon, 2013; Ma et al., 2013; Rybicki et al., 2013). In addition, plants are safer than traditional expression systems as they do not produce endotoxins or support the growth of potentially infectious viruses or prions (Moustafa et al., 2016). Plants can also mediate complex post‐translational modifications, including both glycosylation and disulphide bond formation (Faye et al., 2005; Tschofen et al., 2016). Furthermore, the remarkable plasticity of the plant proteome enables the simultaneous co‐expression of multiple proteins, enabling the carefully regulated manipulation of the secretory pathway for the production of proteins with homogenous mammalian‐like glycosylation (Castilho and Steinkellner, 2012; Castilho et al., 2010). Lastly, plant‐derived vaccine antigens offer the potential for oral immunization, although it is expected that some degree of formulation would be necessary at the least to ensure dose consistency (Rybicki, 2009).
Immunization is unarguably the most effective biomedical intervention against infectious diseases and has resulted in the successful licensing of over 70 vaccines for use in humans (Nabel, 2013). The most consistent biomarker of vaccine efficacy is the induction of neutralizing antibodies, which in the case of many enveloped viruses target the glycoprotein spikes on their surfaces (Nabel, 2013). Given this observation, the development of glycoprotein‐based antigens is a major focal point in the development of vaccines against many viral pathogens (summarized in Figure 1) (Hajj Hussein et al., 2015). Although not a focus of this review, it is also noteworthy that, in addition to their use as vaccine antigens, recombinant glycoproteins also represent potential diagnostic reagents, particularly in the One Health initiative (Rybicki, 2017).
Figure 1.

Production of recombinant glycoproteins as subunit vaccines to protect against viral infections. Virus‐derived glycoproteins often represent the major antigenic determinants of viral infections, and therefore, the production of recombinant glycoproteins in heterologous expression systems is of considerable interest for vaccine development. This diagram shows a typical enveloped virion where the surface is decorated with trimeric glycoproteins which facilitate entry into susceptible target cells. The glycoprotein gene is cloned into a heterologous expression vector and the protein overexpressed (for example, transiently by Agrobacterium‐mediated infiltration). The glycoprotein is purified from contaminating host proteins and formulated for immunization to elicit neutralizing antibodies against the virus.
Whilst recombinant viral structural proteins generally accumulate at reasonable levels in plants, many viral glycoproteins express poorly, or even below, the threshold of detection by standard methods, for reasons that are not completely understood. This has been reported for a number of different viral glycoproteins and has most likely been observed for others in work that has not been published. In our experience, this is associated with severe tissue necrosis that is indicative of endoplasmic reticulum (ER) stress arising from the accumulation of misfolded proteins and the associated unfolded protein response (Howell, 2013; Pera et al., 2015; Phoolcharoen et al., 2011). This phenomenon has been described in the literature for the extracellular subunit of the Ebola glycoprotein (GP1), rotavirus VP7 (Pera et al., 2015; Phoolcharoen et al., 2011) and observed in our laboratory for the full‐length GnGc proteins of both Rift Valley fever virus and Crimean–Congo haemorrhagic fever virus, as well as the premembrane and envelope proteins of Zika virus. It is worth noting, however, that tissue necrosis is not limited to viral glycoproteins and may arise from the overexpression of any protein that exceeds the folding capacity of the system (Liu and Howell, 2010).
Although a minimum expression yield of 1% of the total soluble protein is generally regarded as a prerequisite to justify industrial production, this is rarely achieved and especially not for viral glycoproteins (Rybicki, 2009). However, many of the influenza haemagglutinin (HA) glycoproteins express at unusually high levels in planta, and candidate vaccine antigens have been reported to accumulate at ~50 mg/kg (D'Aoust et al., 2008; Shoji et al., 2011). Amongst the most impressive demonstrations of plant molecular farming is the pioneering work of Medicago Inc. which have developed a highly scalable expression platform for the production of the influenza virus glycoprotein. Their lead vaccine candidate, plant‐derived influenza virus haemagglutinin (HA) virus‐like particles, is currently being evaluated in phase 3 efficacy trials which, if successful, should lead to FDA approval by 2020. At the time of writing, influenza HA remains the only viral glycoprotein expressed in plants that has been evaluated in human trials (Pillet et al., 2016).
It is unclear why many viral glycoproteins only accumulate at low levels in planta although we have surmised that this may be due to either incompatibility with the endogenous plant chaperones or inadequate levels of crucial chaperones required to facilitate folding of the overexpressed protein. The lack of adequate levels of molecular chaperones has been acknowledged as a bottleneck in other heterologous expression systems (Hsu and Betenbaugh, 1997; Rudolph and Lilie, 1996). If this is taken to be true for plants, a major challenge which will need to be addressed is the identification of which chaperone or even combination of chaperones is rate‐limiting for the expression of any given glycoprotein. In this review, we will discuss recent successes in the production of plant‐produced viral glycoproteins and critically discuss considerations for the production of authentic, vaccine‐relevant mimetics of native glycoproteins in plants.
Factors influencing expression of functional glycoproteins
Despite the well‐documented successes in certain instances of expression of virus glycoprotein vaccine candidates in plants, in many cases, yields are too low for generating sufficiently large batches for vaccination programmes. However, there are a number of different strategies that can be taken into consideration which may not only improve yields, but could also improve functionality of the antigens. These include factors such as targeting proteins to the secretory pathway, N‐glycosylation, proteolytic maturation along the secretory pathway and the influence of chaperones in the endoplasmic reticulum.
Targeting recombinant glycoproteins into the secretory pathway
In eukaryotes, the translocation of glycoproteins through the ER is essential to impart authentic glycosylation and other post‐translational modifications (PTMs), as well as to impose quality control prior to further progress along the secretory pathway. In the ER, newly synthesized proteins interact with molecular chaperones and glycosylation enzymes, which act in concert to facilitate the folding and maturation of the protein (Braakman and Hebert, 2013). Terminally misfolded proteins or incompletely assembled polypeptides do not progress beyond the ER but instead are targeted into the ER‐associated protein degradation (ERAD) pathway for proteosomal degradation (Hiller et al., 1996; Jensen et al., 1995; Lippincott‐Schwartz et al., 1988; Sommer and Jentsch, 1993; Ward et al., 1995). Given the critical role of the secretory pathway to facilitate proper folding and glycosylation, targeting recombinant proteins to different subcellular localizations may improve their accumulation at the expense of the native structure.
Glycoproteins naturally encode an N‐terminal signal sequence (typically 20–30 amino acid residues) that targets the newly synthesized protein into the ER (Hegde and Bernstein, 2006). The nature of this signal peptide determines the efficiency of translocation into the ER and subsequent cleavage of the leader peptide from the glycoprotein (Braakman and Hebert, 2013). To illustrate this point, it has been reported that in mammalian cells the native HIV Env signal peptide is inefficiently cleaved, resulting in prolonged retention of the protein in the ER and contamination of virions with aberrantly folded Env species (Crooks et al., 2011; Land et al., 2003; Li et al., 1996, 2000). The use of highly efficient foreign signal peptides is often exploited for the production of glycoproteins in heterologous systems, such as the honeybee melittin signal sequence in insect cells or the tissue plasminogen activator leader sequence in mammalian expression systems (Sanders et al., 2013; Tessier et al., 1991). Several examples of heterologous signal peptides used for glycoprotein expression in plants are listed in Table 1. We routinely make use of the LPH leader sequence, a murine monoclonal antibody‐derived signal peptide, to target recombinant proteins into the plant secretory pathway. This approach has shown promise for the production of trimeric Env glycoproteins from HIV and of a soluble Gn antigen from RVFV (Mbewana, 2017).
Table 1.
Notable examples of heterologous signal peptides used for the production of viral glycoproteins in plants
| Signal peptide | Origin of signal peptide | Antigen | Expression host | Reference |
|---|---|---|---|---|
| LPH | Signal peptide from murine monoclonal antibody mAb24 heavy chain (Fischer et al., 1999) | Rift Valley fever virus Gn | N. benthamiana | Mbewana (2017) |
| Chimeric Gn/influenza HA particle‐forming fusion protein | N. benthamiana | Mbewana (2017) | ||
| HIV Env gp140 | N. benthamiana | Margolin et al. (submitted for publication) | ||
| PDI | Signal sequence from protein disulphide isomerase of Medicago sativa GenBank CAA77575.1 | Influenza H1 HA | N. benthamiana | D'Aoust et al. (2008) |
| Chimeric HIV gp140/influenza HA particle‐forming fusion protein | N. benthamiana | WO 2012/083 445 A1 | ||
| PR‐1a | Signal peptide derived from pathogenesis‐related protein 1a of Nicotiana tabacum NCBI BAA14220 | Influenza H5 | N. benthamiana | Shoji et al. (2009a) |
| Yellow fever E protein | N. benthamiana | Tottey et al. (2018) | ||
| Barley α‐amylase | Signal peptide from barley α‐amylase GenBank CAX51374 | Truncated HIV‐1 gp41 | N. benthamiana | Kessans et al. (2013) |
| SIV gp130 | Zea mays | Horn et al. (2003) | ||
| PR‐S | Signal peptide from extracellular tobacco protein PR‐S (Sijmons et al., 1990) | Rabies G protein | N. tabacum | Ashraf et al. (2005) |
TSP, total soluble protein.
Another commonly used signal peptide is the protein disulphide isomerase (PDI) signal sequence which was used by Medicago Inc. for the production of recombinant influenza H1 HA, but not H5 HA which was expressed under the control of the native signal peptide (D'Aoust et al., 2008). In contrast, Fraunhofer USA has reported the use of the pathogenesis‐related protein (PRP)‐1a signal peptide to produce soluble influenza H5 HA in plants (Shoji et al., 2009a). We have found that a soluble RVFV Gn glycoprotein was only expressed when fused to the LPH signal sequence but not the PDI signal sequence (Mbewana, 2017). Other pertinent examples include the rabies virus glycoprotein which has been produced in transgenic tomatoes using the native signal peptide and in transgenic Nicotiana tabacum under the control of the PRP signal peptide (Ashraf et al., 2005; McGarvey et al., 1995). The latter antigen, which was further modified to contain a C‐terminal ER retention signal, protected mice against lethal rabies challenge (Ashraf et al., 2005).
It appears that the impact of different signal peptides on heterologous protein expression can only be determined empirically. To illustrate this point further, it was reported that detectable expression of a truncated HIV gp41 antigen could be achieved in tobacco plants using the barley alpha‐amylase signal peptide, but not when the signal sequences from apple pectinase, N. plumbaginifolia calreticulin or rice alpha‐amylase were used (Kessans et al., 2013). A recent study has suggested that tissue necrosis during heterologous protein expression in planta could be linked to improper cleavage of the signal peptide. The authors reported that substituting the Arabidopsis thaliana chitinase signal peptide for the native signal sequence of transforming growth factor‐β1 resulted in high levels of protein production whilst still maintaining a healthy phenotype in N. benthamiana plants (Wilbers et al., 2016).
It is also worth noting that the targeting of the recombinant H5 HA protein into the plant secretory pathway resulted in the formation of enveloped VLPs which budded from the plasma membrane in the absence of other accessory proteins (D'Aoust et al., 2008). This phenomenon appears to be unique to plants, as other heterologous expression systems require neuraminidase to sever the sialic acid linkages that tether budding virions to the host cell (Chen et al., 2007). Plants, however, lack endogenous sialic acid activity, enabling HA‐containing particles to bud from the plasma membrane into the apoplastic spaces between plant leaves (D'Aoust et al., 2008; Seveno et al., 2004). Our group has also observed the assembly of similar enveloped H5 HA VLPs (Rybicki 2014), although early attempts to recover the recombinant protein that included the use of Triton‐X100 precluded the recovery of enveloped VLPs (Mortimer et al., 2012). A similar phenomenon has been described by Fraunhofer USA for the production of enveloped Ebola virus VLPs (eVLP) composed of the full‐length glycoprotein (Karczewski and Ysibov, 2016). This is a useful way of improving the immunogenicity of small soluble proteins which may otherwise be poorly immunogenic (Bachmann and Jennings, 2010; Zabel et al., 2013).
The potential impact of plant‐derived N‐glycosylation
Whilst the ability of plants to mediate N‐glycosylation is regarded as an advantage over more primitive prokaryotic expression systems, complex glycans derived from plants are unique in that they contain core β(1,2)‐xylose and α(1,3)‐fucose but lack both the β(1,4)‐galactose and sialic acid that is characteristic of mammalian cell glycosylation (Strasser et al., 2008; Figure 2). The observation that these glyco‐epitopes were immunogenic in small animals, and reports that serum antibodies against these epitopes can be found in humans, raised concerns of possible allergic reactions and rapid clearance following administration (Bardor et al., 2003; Bosch and Schots, 2010; Jin et al., 2008). In spite of this trepidation, Medicago's HA‐based vaccine candidates were reported to be safe, even in volunteers with pre‐existing plant allergies, suggesting that these concerns were largely unwarranted (Ward et al., 2014). However, it is acknowledged that some transient antibody responses to the plant‐derived glyco‐epitopes were observed, although these were generally weak (Ward et al., 2014). The potential negative implications for use of proteins with these plant‐specific glyco‐epitopes have prompted extensive efforts to humanize the glycosylation of proteins in plants by elimination of plant‐specific glycans and the introduction of human glycosyltransferases [reviewed by Strasser et al. (2014)]. It is also interesting to note that a number of different plant polysaccharides are even being explored as adjuvants, with several having advanced into clinical trials [reviewed by Rosales‐Mendoza et al. (2016)].
Figure 2.
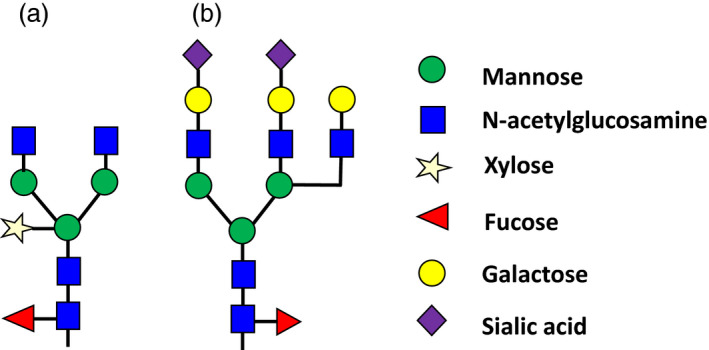
Comparison of typical N‐glycan structures from glycoproteins produced in (a) plants and (b) mammalian expression hosts. Whilst the core glycan machinery is preserved amongst eukaryotes, species‐specific modifications occur in the Golgi apparatus. Plant‐produced glycoproteins contain characteristic α‐(1,3) fucose and (β1,2) xylose residues, whereas mammalian‐derived proteins contain α(1,6)‐fucose moieties and terminal sialic acid residues. Sialylation does not occur naturally in plants.
The HIV Env glycoprotein is amongst the most heavily glycosylated proteins described, with approximately half the molecular mass of gp120 being attributed to glycans (Joyce et al., 2016; Lasky et al., 1986). These glycans are host‐derived and are considered ‘self‐antigens’, enabling the virus to mask vulnerable epitopes with a poorly immunogenic shield (Jardine et al., 2013; Julien et al., 2013; McGuire et al., 2013; Sok et al., 2016; Wei et al., 2003). Similarly, influenza HA also contains multiple glycosylation sites which appear to have been increasing since 1918, presumably to enable the virus to escape from neutralizing antibodies (Herve et al., 2015; Job et al., 2013; Skehel et al., 1984; Wei et al., 2010). Despite reports that glycan occupancy may be lower in plant‐produced proteins than in other production platforms, Medicago's plant‐produced HA was reported to contain glycans at all six potential N‐glycan sites in the extracellular portion of the protein (Le Mauff et al., 2015; Ward et al., 2014). It is unclear if potential underglycosylation will have an impact on recombinant HIV Env glycoproteins expressed in plants which are expected to contain ~90 glycans on each trimer (Cao et al., 2017). Underglycosylation could potentially compromise the folding of the glycoproteins as N‐glycans are critical for the recruitment of the lectin chaperones: calnexin and calreticulin (Wada et al., 1991). Furthermore, in the case of HIV‐1 Env, many of the broadly neutralizing antibodies isolated from natural infection target epitopes that contain a glycan component suggesting that a recombinant immunogen should reproduce these structures (Crispin et al., 2018). Encouragingly, a recent publication by Castilho and colleagues has suggested the co‐expression of a Leishmania major oligosaccaryltransferase can be used to improve the glycan occupancy of plant‐derived proteins (Castilho et al., 2018). Furthermore, ongoing efforts to humanize the glycosylation of recombinant plant‐produced proteins have demonstrated the successful removal of plant‐specific N‐glycan moieties and the introduction of human glycosyltransferases in planta (Strasser et al., 2014). Although this approach has been developed primarily for the production of recombinant antibodies, it is expected that as the range of plant‐produced viral glycoproteins expands that similar approaches will be implemented (Kallolimath and Steinkellner, 2015). A particularly interesting aspect of this work will be to determine the impact on the antigenicity and immunogenicity of plant‐produced viral glycoproteins expressed in wild‐type and glycoengineered plants.
The HIV Env glycoprotein appears to be unique in that many of the epitopes targeted by broadly neutralizing antibodies in viruses isolated from natural infections incorporate a glycan component (Andrabi et al., 2015; Balla‐Jhagjhoorsingh et al., 2013; Blattner et al., 2014; Falkowska et al., 2014; Gorman et al., 2016; Julien et al., 2013; Kong et al., 2013; Lyumkis et al., 2013; Sanders et al., 2002; Scanlan et al., 2002; Sok et al., 2014; Trkola et al., 1996). Encouragingly, the recombinant HIV Env gp140 produced by Rosenberg and colleagues in N. benthamiana was reported to react with several prototype monoclonal antibodies including 2G12, which specifically targets a glycan‐dependent epitope on the outer domain of the glycoprotein (Rosenberg et al., 2013). Glycoprofiling of the purified HIV Env protein revealed predominantly high‐mannose‐type glycans, with 21.5% containing plant‐specific α(1,3)‐fucose and β(1,2)‐xylose residues (Rosenberg et al., 2013). In natural infections, the virion‐associated Env comprises of predominantly unprocessed oligomannose moieties (Bonomelli et al., 2011; Doores et al., 2010). This is considered an important hallmark of the native structure, as the steric constraints of the trimer limit access of the glycan processing enzymes along the secretory pathway (Bonomelli et al., 2011; Doores et al., 2010; Pritchard et al., 2015). Whilst it is encouraging that plant‐produced HIV Env antigens can reproduce the structure of complex, glycan‐dependent epitopes, a recent study has suggested that sialic acid residues may contribute to the epitopes targeted by broadly neutralizing antibodies (Andrabi et al., 2017). Given this observation, wild‐type plants may not be ideal for the production of authentic mimetics of the glycoprotein as sialylation does not naturally occur in plants (Seveno et al., 2004). Recent studies have reported the artificial engineering of sialic acid biosynthetic pathways in plants which could be used to address this potential limitation (Castilho et al., 2010, 2013; Kallolimath et al., 2016; Paccalet et al., 2007). In apparent contradiction to this, a study conducted by Kong et al. comparing the influence of expression system‐dependent glycosylation concluded that cells lacking sialic acid may improve the immunogenicity of recombinant HIV Env immunogens, though not necessarily for neutralizing antibody responses (Kong et al., 2010).
Proteolytic maturation of viral glycoproteins along the secretory pathway
Many glycoproteins require proteolytic cleavage to assume their mature fusion‐entry conformation required for infection; therefore, the ability of a heterologous expression system to mediate authentic processing of the protein is an important consideration. The HIV Env glycoprotein, for example, undergoes proteolytic cleavage by furin and other subtilisin‐like proteases during its translocation along the secretory pathway (Decroly et al., 1994, 1996; Gu et al., 1995; Hallenberger et al., 1992; Vollenweider et al., 1996). Proteolytic maturation of the glycoprotein is necessary for it to assume its correct quaternary structure (Ringe et al., 2013). This has important implications for vaccine design, as cleavage‐defective Env trimers have been reported to assume aberrant conformations which induce antibodies against epitopes that are sterically inaccessible in the context of the functional virion‐bound trimer and are therefore unable to neutralize the virus (Ringe et al., 2013; Tran et al., 2014).
Given these observations, trimeric HIV Env immunogens are often produced in mammalian cell culture by replacing the native cleavage site with a hexa‐arginine motif and co‐expressing heterologous furin to promote optimal cleavage of the recombinant glycoprotein (Binley et al., 2002). Furin activity does not naturally occur along the plant secretory pathway, although a precedent exists for the co‐expression of the protease to produce functional human transforming growth factor β1 in planta, suggesting that theoretically it may be possible to produce cleaved HIV Env glycoproteins in plants (Wilbers et al., 2016). Another potential complication with the production of a proteolytically cleaved glycoprotein is the labile association between the respective subunits. In the case of cleaved HIV Env trimers, an artificial disulphide bond is typically introduced at the interface of gp120 and the ectodomain of gp41 to ensure that the two subunits remain associated after cleavage (Binley et al., 2000). During natural infection, in the absence of any stabilizing mutations between the subunits, the gp120 subunit may be shed leaving gp41 stumps on the surface of the virion, which are proposed to serve as immunological decoys (Moore et al., 2006). Accordingly, it is important that a HIV Env‐derived vaccine antigen should be stable and that the gp120 and gp41 components remain associated for maximum immunogenicity.
Recent studies in mammalian cell cultures have managed to circumvent this requirement for furin cleavage by employing a flexible glycine‐rich linker in place of the native HIV Env cleavage site (Georgiev et al., 2015; Sharma et al., 2015). These cleavage‐independent trimers have been reported to be authentic mimics of the virion‐bound trimer, to retain native‐like antigenicity and to elicit neutralizing antibodies in preclinical studies (Georgiev et al., 2015; Pauthner et al., 2017; Sharma et al., 2015). A similar approach has also been reported for the production of respiratory syncytial virus fusion glycoprotein in mammalian cells, where the furin cleavage site was replaced with a flexible linker. This suggests that this strategy may be broadly applicable to other viral Type I glycoproteins, such as Ebolavirus GP (Joyce et al., 2016). We have exploited this design strategy to produce soluble cleavage‐independent HIV‐1 Env trimers in plants, facilitating the first immunogenicity studies of trimeric plant‐produced HIV‐1 Env proteins (Margolin et al., submitted for publication).
The structurally analogous influenza virus HA glycoprotein also requires proteolytic cleavage for viral infectivity, resulting in structural rearrangements that expose the fusion peptide (Bullough et al., 1994; Owens and Compans, 1990). Cleavage of H0 (the immature precursor glycoprotein) is mediated by ubiquitous cellular proteases, including the endoprotease furin, and may even be mediated by the proteases of co‐infecting bacteria (Galloway et al., 2013; Steinhauer, 1999; Stieneke‐Grober et al., 1992). The expression of HA in plants, however, is reported to yield a single product of 72 kDa, consistent with the size expected for uncleaved HA (D'Aoust et al., 2008; Mortimer et al., 2012; Shoji et al., 2011). A subsequent report confirmed the presence of low levels of HA1 and HA2 (the extracellular and transmembrane subunits of HA, respectively) suggesting that some level of precursor cleavage does occur, albeit very inefficiently (Le Mauff et al., 2015). Unlike with HIV, cleavage is not a requirement to elicit protective immunity against the influenza glycoprotein, as uncleaved H5 HA VLPs confer protection against lethal challenge (D'Aoust et al., 2008; Mett et al., 2008; Pillet et al., 2015; Shoji et al., 2009a).
The chaperone folding pathway in the plant endoplasmic reticulum
The endoplasmic reticulum in eukaryotes is a major site of protein folding and quality control, where nascent proteins interact with molecular chaperones (Saibil, 2013). Protein folding in the ER is directed by two major chaperone systems: these are the classical chaperone system and the carbohydrate‐binding system, although the two systems are not mutually exclusive (Braakman and Hebert, 2013). The folding of glycoproteins is intrinsically linked to N‐glycosylation (recently reviewed by Richard Strasser) (Strasser, 2018), as trimming of the glycan moiety signals entry and exit from the calnexin/calreticulin quality control cycle (Lannoo and Van Damme, 2015). Misfolded glycoproteins are retained in the ER and will cycle through the calnexin/calreticulin folding cycle until the correct conformation is achieved, or they will be targeted for degradation (Figure 3). In the absence of proper glycosylation, nascent glycoproteins may be prone to misfolding as a result of mispaired cysteine residues. This is best illustrated by the work of Daniels and colleagues, who provided evidence that the strategic placement of glycans on influenza HA serves to shield susceptible cysteine residues during folding (Daniels et al., 2003). Calnexin and calreticulin further form complexes with the oxidoreductase ERp57 which catalyses the formation of disulphide bonds (Frickel et al., 2004; Oliver et al., 1997, 1999; Zapun et al., 1998). In addition to ERp57, other members of the protein disulphide isomerase family are also responsible for catalysing disulphide bonds resulting in a level of built‐in redundancy into the system (Rutkevich et al., 2010).
Figure 3.

Carbohydrate‐mediated folding in the endoplasmic reticulum. Following translation, the nascent protein (red) enters into the endoplasmic reticulum (ER) through a translocon pore (SEC61) where it sequentially interacts with a network of chaperones and enzymes to acquire its correct higher order structure. During entry into the ER, the oligosaccharyltransferase complex (OST) transfers a preformed Glc3Man9 NAc2 glycan precursor to the Asn of the Asn‐X‐Ser/Thr motif in the polypeptide chain. The two outermost glucose residues are then sequentially removed by glucosidase I (G‐I) and glucosidase II (G‐II) signalling entry of the resulting monoglucosylated (GlcMan9 NAc2) protein into the calnexin (CNX)/calreticulin (CRT) folding cycle. Calnexin is the membrane‐bound homologue of calreticulin and preferentially interacts with nascent proteins associated with the ER membrane (only calreticulin is indicated in this diagram for the sake of simplicity). This interaction with CNX/CRT facilitates the co‐ordinated activities of other chaperones such as the oxidoreductase ERp57 to further assist with folding. Incorrectly folded proteins are reglucosylated by UDP‐glucose:glycoprotein glucosyltransferase (UGGT) signalling their re‐entry into the CNX/CRT folding cycle to undergo another round of chaperone‐mediated folding. In contrast, correctly folded proteins are acted on by G‐II, resulting in the removal of their remaining glucose residue. This signals release from the CNX/CRT pathway enabling export of the protein into the Golgi. Glycoproteins may undergo several rounds of chaperone‐mediated folding to assume the correct quaternary structure before continuing along the secretory pathway. In contrast, terminally misfolded glycoproteins are targeted for proteolytic degradation via the endoplasmic reticulin‐associated degradation (ERAD) pathway.
Given the extended period of divergent evolution of mammals and plants, the low expression levels reported for many viral glycoproteins may be due to incompatibility with the endogenous plant chaperone machinery. In support of this, putative homologues of key chaperones known to interact with glycoproteins exhibit low‐level sequence identity to their human counterparts. We are currently exploring the co‐expression of human molecular chaperones in planta (patent application: PA167643/P) to improve the production of heterologous glycoproteins, based on the hypothesis that the endogenous plant chaperones may represent a critical bottleneck for high levels of expression.
The co‐expression of various protein disulphide isomerases may improve the disulphide bond formation of soluble HIV Env antigens produced in plants. The gp120 subunit of HIV Env contains nine intrachain disulphide bonds with an additional disulphide bond present in the transmembrane subunit (gp41) (Go et al., 2014; Leonard et al., 1990). Given the presence of 20 cysteine residues in each gp160 subunit, and the obvious potential for aberrant disulphide bond formation, reports of cross‐linked dimers and aggregates in the literature are unsurprising (Leonard et al., 1990; Owens and Compans, 1990). We and others have reported the presence of unresolved higher molecular weight products following Western blotting of plant‐produced HIV Env gp140 proteins (Rosenberg et al., 2013) (Margolin et al., submitted for publication). Rosenberg and colleagues speculated that these comprised of higher order oligomers that were incompletely resolved by SDS‐PAGE (Rosenberg et al., 2013). We hypothesize that these unresolved higher molecular weight products are actually aggregates of misfolded protein with scrambled disulphide bridges. We have observed that these higher molecular weight products do not dissociate in the presence of detergent or under varying conditions of pH and salt concentrations. However, boiling the recombinant proteins for extended lengths of time in the presence of β‐mercaptoethanol results in improved dissociation of these higher order molecular weight protein species, suggesting that improving the disulphide bond formation may improve protein production.
Current candidate viral glycoproteins produced in plants
A number of viral glycoprotein vaccine candidates have been successfully expressed in plants to date; either transiently or by generation of transgenic or transplastomic plants (Table 2). These include antigens against high‐impact human diseases such as influenza and HIV, as well as those against vector‐borne and other zoonotic viral diseases such as Zika, dengue and West Nile fevers, Japanese encephalitis, coronavirus infections, Crimean–Congo haemorrhagic and Rift Valley fevers, rabies, infectious bronchitis, Newcastle disease and Ebola. Many of these have been tested for specific immunogenicity in animals, and some including infectious bronchitis, influenza, Newcastle disease and rabies have been tested for efficacy using animal challenge models, with encouraging results.
Table 2.
Summary of recombinant glycoprotein vaccine antigens produced in plants
| Virus | Glycoprotein (antigen) | Expression host (leaves unless otherwise specified) | Transient (T)/Transgenic (TG)/Transplastomic (TP) yields | Antigen tested in animals | Animal testing | Immunogenicity | Challenge efficacy | Reference |
|---|---|---|---|---|---|---|---|---|
| CCHFV | GnGc | N. tabacum | TG 0.9–1.4 mg/kg FW | Tobacco leaf pellets | BALB/c mice oral | IgG; IgA | ND | Ghiasi et al. (2011) |
| N. tabacum hairy roots | TG 1.8 mg/kg FW | Hairy root pellets | BALB/c mice oral | IgG; IgA | ND | |||
| Coronavirus | S1‐GFP fusion (his‐tagged) | N. tabacum | T NQ | NT | ND | – | – | Li et al. (2006) |
| N. tabacum | TP 0.2% TSP | NT | ND | – | – | |||
| S1 | N. tabacum root | TG NQ | Lyophilized tobacco root | ALB/c mice oral | IgG | ND | Pogrebnyak et al. (2005) | |
| Lycopersicon esculentum (tomato) fruit | TG NQ | Lyophilized tomato fruit | BALB/c mice oral | IgA | ND | |||
| Dengue virus | pRM/E | Lactuca sativa (lettuce) | TP NQ | NT | ND | – | – | Kanagaraj et al. (2011) |
| D2EIII | N. benthamiana | T 5.6 mg/kg FW | Ni+ affinity‐purified D2III | C3H mice IM | IgG and PRNT neutralization | ND | Saejung et al. (2007) | |
| Et (truncated) | N. benthamiana | T 600 mg/kg FW | NT | ND | – | – | Martinez et al. (2010) | |
| pRM/Et | N benthamiana | T 500 mg/kg FW | NT | ND | – | – | ||
| HBCag‐DIII | N. benthamiana | T 400 mg/kg FW | NT | ND | – | – | ||
| CTB‐EIII | N. tabacum | TG 0.0053%–0.019% TSP | NT | ND | – | – | Kim et al. (2010) | |
| EDIII (consensus) | Oryza sativa (rice) callus | TG 450 mg/kg FW | NT | ND | – | – | Kim et al. (2012) | |
| Ebola virus | EIC (GP1 fusion with heavy chain monoclonal ab) | N. benthamiana | T 50 mg/kg FW | Protein G affinity‐purified EIC | BALB/c mice SC | IgG | ND | Phoolcharoen et al. (2011) |
| IBV | S (spike protein) | Solanum tuberosum (potato) tubers | TG 2.39–2.53 mg/kg FW | Fresh potato tuber; potato tuber extract | Chickens oral; IM | IL‐2; Neutralization | 66.7% survival oral; 100% survival IM | Zhou et al. (2004) |
| JEV | EDIII‐BaMV CP | N. benthamiana | T NQ | NT | ND | – | – | Chen et al. (2017) |
| Chenopodium quinoa | T 4300–8900 mg/kg | Purified chimeric virus particles | BALB/c mice IP | IgG; PRNT neutralization | ND | |||
| HIV | gp140 ∆CFI Env | N. benthamiana | T 80 mg/kg FW | NT | ND | – | – | Rosenberg et al. (2013) |
| dgp41 (deconstructed gp41) | N. benthamiana | T 9 mg/kg FW | NT | ND | – | – | Kessans et al. (2013) | |
| Consensus Env/HA chimaera | N. benthamiana | T NQ | NT | ND | – | – | WO 212/083445A1 | |
| gp140 NFL | N. benthamiana | T 5–6 mg/kg | Lectin‐column‐purified gp140 NFL | Rabbits IM | Binding and neutralizing antibodies | ND | Margolin et al. (submitted for publication) | |
| Influenza virus | H1N1 HA (haemagglutinin) | N. benthamiana | T 90 mg/kg FW | Ni+ affinity‐purified HA | Mice IM; rabbits IM; ferrets IM | HAI titres and virus neutralization | ND | Shoji et al. (2011) |
| H5N1 HA | N. benthamiana | T 50 mg/kg FW | Ni+ affinity‐purified HA | Mice IM; rabbits IM; ferrets IM | HAI titres and virus neutralization | ND | ||
| H5N1 HA | N. benthamiana | T NQ | Ni+ affinity‐purified HA | BALB/c mice IN; ferrets IN | IgA, IgG; HAI titres and virus neutralization | Mice 100% survival; Ferrets 100% survival | Major et al. (2015) | |
| H5N1 HA (VLPs) | N. benthamiana | T 50 mg/kg FW | Fetuin affinity‐purified HA VLPs | BALB/c mice IM | HAI titres | 100% protection | D'Aoust et al. (2008) | |
| H1N1 HA (VLPs) | N. benthamiana | T 50 mg/kg FW | Fetuin affinity‐purified HA VLPs | BALB/c mice IM | HAI titres | 100% protection | ||
| Newcastle disease virus | F (fusion protein) | Zea mays (maize) kernels | TG 0.9%–0.17% TSP | Ground kernels | Chicken oral | IgG | 100% protection | Guerrero‐Andrade et al. (2006) |
| HN | Solanum tuberosum tubers | TG 300–600 mg/kg | Crude leaf extracts | BALB/c mice IP; oral | IgG; IgG + IgA | ND | Berinstein et al. (2005) | |
| F | S. tuberosum | TG 300–600 mg/kg | Crude leaf extracts | BALB/c mice IP; oral | IgG; IgG + IgA | ND | ||
| HN | N. benthamiana | T 3000 mg/kg FW | NT | ND | – | – | Gomez et al. (2009) | |
| Rabies virus | Surface protein G | N. tabacum | TG 0.1%–0.38% TSP | Human antirabies IgG affinity‐purified G | BALB/C mice IP | IgG | 100% protection | Ashraf et al. (2005) |
| Surface protein G | Z. mays kernels | TG 25 mg/kg FW | Ground kernels | Sheep oral | RFFIT neutralization | 66% survival | Loza‐Rubio et al. (2012) | |
| RVFV | Gn deletion mutant | Arabidopsis thaliana whole plants | TG NQ | Fresh leaves | C57/B1 mice oral | IgG | ND | Kalbina et al. (2016) |
| WNV | E domain III (DIII) (his‐tagged) | N. benthamiana | T 1.16–73 mg/kg FW | Ni+ affinity‐purified DIII | BALB/c mice SC | IgG | ND | He et al. (2014) |
| Zika virus | E glycoprotein (his‐tagged) | N. benthamiana | T 160 mg/kg FW | Ni+ affinity‐purified E | C57/BL6 mice SC | IgG; IFNƳ, IL‐4, IL6; PRNT neutralization | ND | Yang et al. (2017b) |
| HBCag‐DIII (VLPs) | N. benthamiana | T 1.8 mg/kg FW | Sucrose gradient‐purified VLPs | C57/BL6 mice SC | IgG; IFNƳ; PRNT neutralization | ND | Yang et al. (2017a) |
CCHFV, Crimean–Congo haemorrhagic fever virus; HAI, haemagglutinin inhibition; IBV, infectious bronchitis virus; IM, intramuscular; IN, intranasal; IP, intraperitoneal; JEV, Japanese encephalitis virus; mg/kg FW, mg/kg fresh weight; ND, not done; NQ, no quantitation; NT, not tested; PRNT, plaque reduction neutralization test; RFFIT, rapid fluorescent focus inhibition test; RVFV, Rift Valley fever virus; SC, subcutaneous; WNV, West Nile virus.
Influenza
Human seasonal influenza disease is presently caused by three influenza virus A subtypes (H1N1, H1N1pdm09 and H3N2) and two influenza virus B subtypes (Yamagata and Victoria). Annually 5%–10% of adults and 20%–30% of children are infected, with 3–5 million cases of severe illness, and about 250 000–500 000 deaths (WHO, 2014), although this figure has recently been increased to 300 000–650 000 deaths, based on new modelling studies (Iuliano et al., 2017). The conventional chicken egg‐based vaccine technology has a production capacity as of 2011 of 1.4 billion doses of trivalent vaccine and a production level of 620 million doses. However, it takes 6 months to make and produce the vaccines, and thus, vaccine antigens are chosen based on the viruses circulating at that time, which in the Northern Hemisphere 2017–2018 influenza season has been predicted to cause problems due to H3N2 vaccine mismatch with the circulating type (Dugan, 2017).
The potential for rapid response and scalability of plant‐based expression is best illustrated by the pioneering work of Medicago Inc. and the Fraunhofer USA group in the development of candidate vaccines for pandemic influenza strains (D'Aoust et al., 2008; Shoji et al., 2011; Stoger et al., 2014; Streatfield et al., 2015). Given the slow production time and limited production capacity of using embryonated hen's eggs, traditional influenza vaccine manufacturers are not equipped to respond timeously, or in sufficient volume, to a pandemic outbreak (D'Aoust et al., 2010; Quan et al., 2016). Typical egg‐based production of split virion vaccines can take up to 6 months due to the limitations inherent in the production platform which can be further hampered by variable production yields (Manini et al., 2017). In contrast, Medicago Inc in the USA has reported the production of a fully formulated HA VLP vaccine within 21 days from the release of the sequence for the pandemic A/H1N1(A/California/04/09) strain (D'Aoust et al., 2010). Furthermore, their production platform is rapidly scalable and has been used to successfully produce 10 million doses of a monovalent H1N1 VLP vaccine within 30 days under the Defense Advanced Research Projects Agency (DARPA) Blue Angel programme [https://www.rt.com/usa/future-vaccine-darpa-research-255/].
The Fraunhofer Centre for Molecular Biotechnology (USA) has described the development of a similar transient plant expression platform for the production of soluble HA antigens which has been reported to yield purified vaccines in just over a month (Shoji et al., 2012). Both of these platforms are highly flexible and have been used to produce HA from several seasonal and pandemic influenza subtypes (D'Aoust et al., 2008; Mett et al., 2008; Pillet et al., 2015; Shoji et al., 2008, 2009a,b, 2011, 2012). These antigens have demonstrated promising immunogenicity in preclinical animal models with several reports of protection against lethal virus challenge (D'Aoust et al., 2008; Mett et al., 2008; Pillet et al., 2015; Shoji et al., 2008, 2009a,b, 2011). Consistent with these studies, clinical trials have reported that these vaccines are safe and highly immunogenic in human volunteers (Chichester et al., 2012; Cummings et al., 2014; Landry et al., 2010, 2014; Ward et al., 2014). Prompted by the limited access to vaccines in South Africa during the 2009 H1N1 pandemic, our group has developed a similar production platform for the expression of both full‐length and soluble HA antigens (Mortimer et al., 2012).
HIV
The human immunodeficiency virus is responsible for an unprecedented global pandemic, particularly in sub‐Saharan Africa which bears a disproportionate burden of global infections relative to its size (Shao and Williamson, 2012). A broadly protective vaccine is urgently needed to combat the spread of HIV, and even a partially effective vaccine is expected to make a large impact on the epidemic (Medlock et al., 2017). Current state‐of‐the‐art subunit vaccines are based on rationally engineered soluble Env trimer mimetics produced in mammalian cell culture systems, with the intention of eliciting neutralizing antibodies (Sanders and Moore, 2017). The high costs and limited production capacity available are expected to be particularly problematic, considering the likelihood that repeat immunizations will be required (Shao and Williamson, 2012). Given the structural similarity between influenza HA and the HIV Env glycoprotein, it is reasonable to assume that plants may have the capacity to emulate the structure of the HIV Env glycoprotein trimer (Karlsson Hedestam et al., 2008). This has prompted us to explore the potential of N. benthamiana for the production by transient agroinfection of trimeric Env glycoproteins based on HIV subtypes circulating in sub‐Saharan Africa (patent application: PPA UK 1617480.7; Margolin et al., submitted for publication).
Very few studies have reported the successful expression of a significant portion of the HIV Env glycoproteins in plants. The first published study describing the expression of the majority of the HIV Env glycoprotein was by Rosenberg and colleagues, who produced a soluble gp140 protein (protein lacks the transmembrane and cytoplasmic domains) in N. benthamiana as a reagent to evaluate the reactivity of plant‐produced monoclonal antibodies (Rosenberg et al., 2013). The authors reported that the recombinant Env protein was produced at ~80 mg/kg by both transient and stable transgenic expression (Rosenberg et al., 2013). Preceding this, a patent filed by Medicago Inc. described the production of chimeric virus‐like particles comprising of a synthetic consensus group M Env protein fused to the transmembrane and cytoplasmic domains of influenza HA. The patent application further reported that expression of the native HIV Env protein could not be detected (international patent application number: WO 2012/083445 A1). The antigens produced in both of these studies were modified to remove the cleavage site which is now known to result in an aberrant structure, unless a flexible linker peptide is introduced at the interface of the gp120 and gp41 to retain the conformational mobility of the two subunits (Georgiev et al., 2015; Ringe et al., 2013; Sharma et al., 2015). It is also noteworthy that the major surface glycoprotein from simian immunodeficiency virus (SIV) (gp130) has been successfully produced in transgenic corn with the aim of developing an orally immunogenic vaccine, although no immunogenicity was reported (Horn et al., 2003).
We have recently described the development of a platform for the expression of soluble HIV‐1 Env gp140 trimers in N. benthamiana plants (patent application: PPA UK 1617480.7). These antigens exploit a 10 residue glycine‐rich linker peptide at the interface of the gp120 and gp41 subunits to eliminate the requirement for furin‐mediated cleavage, which does not naturally occur in plants (Wilbers et al., 2016). Preliminary immunogenicity studies confirmed that the proteins were immunogenic in rabbits; however, despite eliciting high titres of binding antibodies, the vaccines only induced low levels of neutralizing antibodies. The removal, by size exclusion chromatography, of nontrimeric Env species improved the induction of neutralizing antibodies which improved further when animals were primed with recombinant modified vaccinia Ankara (MVA) virus expressing Gag and Env antigens. To our knowledge, this is the first study to report the immunogenicity of a plant‐produced HIV‐1 Env trimer in an animal model (Margolin et al., submitted for publication).
Flaviviruses
The flaviviruses West Nile (WNV), Zika (ZIKV), yellow fever (YFV), Japanese encephalitis (JEV) and dengue (DENV) comprise a group of specifically mosquito‐borne viruses, within the family Flaviriridae. The recent emergence and re‐emergence of WNV, YFV and ZIKV in the Americas (Zumla et al., 2016), and of WNV and tick‐borne encephalitis in Europe (Kaaijk and Luytjes, 2017), have resulted in renewed interest in the development of prophylactic vaccines against these pathogens. These concerns have been further compounded by the finding that infection with ZIKV during pregnancy was associated with birth defects during a recent outbreak in Brazil (Yakob and Walker, 2016). Similar limitations in terms of the scalability and production costs of conventional production platforms as for influenza vaccines have driven interest in the potential of molecular farming to produce vaccines for these emerging flaviruses (Cardona‐Ospina et al., 2016; He et al., 2014; Yap and Smith, 2010). Several dengue envelope antigens have been produced in plants, although there is a paucity of immunogenicity data to support their utility as vaccine immunogens (Kim et al., 2010, 2012; Martinez et al., 2010; Saejung et al., 2007).
The domain III of the flavivirus Env glycoprotein is targeted by neutralizing antibodies during natural infection and is therefore an important target for vaccine‐mediated protection (Beasley and Barrett, 2002; Crill and Roehrig, 2001; Zhao et al., 2016). Martínez and colleagues have reported the successful production of both a truncated envelope protein and a chimeric fusion between the hepatitis B core antigen and the dengue virus envelope domain III (EDIII), for potential use as a vaccine or diagnostic reagent (Martinez et al., 2010). Dengue EDIII has also been produced as a fusion protein with Vibrio cholera toxin B subunit in transgenic potatoes—presumably for use as an oral vaccine, although no immunogenicity was reported (Kim et al., 2010). Similarly, EDIII has been expressed as a soluble protein in both transgenic tobacco and rice callus. The dengue virus glycoprotein has also been produced as a modified polyprotein in transplastomic lettuce, containing part of the capsid in addition to the premembrane and Env proteins (Kanagaraj et al., 2011). In spite of the lack of immunogenicity reports for many of these studies, immunization of mice with N. benthamiana‐derived dengue EDIII has been shown to elicit neutralizing antibodies in mice, warranting further development of these antigens (Saejung et al., 2007). Similarly, a transiently expressed West Nile virus EDIII antigen was reported to be immunogenic in mice (He et al., 2014). Chimeric bamboo mosaic virus particles presenting EDIII from Japanese encephalitis virus have also been produced in plants and were shown to elicit neutralizing antibodies in mice (Chen et al., 2017). More recently, increases in demand and reports of adverse effects against the current live attenuated YFV vaccine have prompted the development of a plant‐produced subunit vaccine as an alternative (Tottey et al., 2018). Tottey and colleagues successfully produced the envelope glycoprotein, as a soluble protein and as a chimeric fusion to bacterial lichenase, both of which protected mice from lethal challenge. It should be noted, however, that these were comparatively less immunogenic than the 17DD live attenuated vaccine (Tottey et al., 2018). Many of these soluble vaccine candidates would benefit from multimerization in the form of synthetic nanoparticles to improve their immunogenicity. The large size of nanoparticles facilitates their entry into lymphatics and enables drainage to lymph nodes to interact with professional antigen‐presenting cells (Manolova et al., 2008). In addition, repeating arrays of antigens promote the cross‐linking of B cells, resulting in sustained and durable antibody responses even in the absence of T‐cell stimulation (Bachmann and Jennings, 2010; Zabel et al., 2013).
Motivated by the recent outbreaks in the Americas, two studies from Arizona State University have described the production of Zika virus glycoprotein antigens in plants. The first successfully expressed a soluble E protein which retained native antigenicity based on the binding of a panel of monoclonal antibodies and elicited both cellular and humoral responses in immunized mice. Furthermore, the levels of neutralizing antibodies in immunized animals exceeded the threshold required for protective immunity (Yang et al., 2017b). The same group has also developed a VLP vaccine comprised of the hepatitis B core antigen presenting the Zika virus E protein domain III, to circumvent potentially inducing antibody‐dependent enhancement (ADE) of infection against dengue infection (Yang et al., 2017a).
Other vector‐borne and zoonotic viruses
Several vector‐borne and zoonotic diseases have recently been identified as prime candidates for the development of vaccines and diagnostics under the One Health initiative: notable examples are the bunyaviruses Crimean–Congo haemorrhagic fever and Rift Valley fever viruses (Rybicki, 2017). Many of these pathogens affect livestock and cause zoonotic infections through direct contact with humans. It is also worth noting that the less stringent regulatory requirements for the commercialization of veterinary vaccines make for a comparatively quicker road to market than human vaccines—and that a successful animal vaccine could quickly become a human vaccine for these viruses (Rybicki, 2010). To this end, a soluble RVFV Gn glycoprotein has been expressed at low levels in stably transformed Arabidopsis thaliana. Although the recombinant protein could not be detected by Western blotting, mRNA expression was confirmed by RT‐PCR and fresh plant material was reported to be orally immunogenic in mice (Kalbina et al., 2016). Our group has expressed and purified a chimeric particle‐forming Gn antigen from N. benthamiana plants which contains the transmembrane and cytoplasmic domains of influenza HA (Mbewana, 2017). This vaccine candidate is of particular interest as immunity against Gn is sufficient to confer protective immunity (de Boer et al., 2010). The full‐length Crimean–Congo haemorrhagic fever virus GnGc protein has also been successfully produced in tobacco leaves and hairy root cultures. When mice were fed with the transgenic material, both IgG and IgA antibodies were elicited, although expression levels of the viral proteins were very low (Ghiasi et al., 2011).
Another plant‐produced viral glycoprotein with potential as a vaccine antigen is the rabies virus glycoprotein G, which has been expressed in transgenic maize, tobacco and tomatoes (Ashraf et al., 2005; Loza‐Rubio et al., 2008; McGarvey et al., 1995). Transgenic maize expressing the G protein was shown to protect sheep from virulent challenge, although the level of protection was lower than the commercial vaccine (Loza‐Rubio et al., 2012). The full‐length glycoprotein from infectious bronchitis virus (IBV) glycoprotein has also been produced in transgenic potatoes and tuber extract was sufficient to protect chickens against virulent challenge following both oral and intramuscular immunizations (Zhou et al., 2004). Similarly, an orally immunogenic Newcastle disease virus fusion protein produced in transgenic maize was reported to protect chickens against challenge (Guerrero‐Andrade et al., 2006).
The extracellular subunit of the Ebola virus glycoprotein (GP1) has been expressed in tobacco plants as an immune complex with a human monoclonal antibody (Phoolcharoen et al., 2011). Interestingly, the authors reported that GP1 could not be produced in sufficient quantities without the heavy chain fusion, due to extensive leaf necrosis. They hypothesized that the presence of the monoclonal antibody segment enabled improved recruitment of molecular chaperones, once again suggesting that the plant chaperone machinery may represent a critical bottleneck for glycoprotein expression. The GP1 component of the fusion protein was properly folded, based on reactivity of a conformational monoclonal antibody. The authors further reported that subcutaneous immunization of mice with the immune complex elicited immune responses comparable to a protective Venezuelan equine encephalitis virus vaccine, although the antibody titres were admittedly lower. A truncated form of the SARS Coronavirus spike glycoprotein has been expressed by transplastomic and stable nuclear transformation of both tobacco and lettuce leaves, for use as an oral vaccine (Li et al., 2006).
Conclusion
Molecular farming is on the cusp of being accepted as a mainstream technology for the production of heterologous pharmaceuticals, with particular appeal for resource‐limited regions. Recent years have borne witness to the licensure of the first plant‐made pharmaceutical for use in humans—taliglucerase alpha—with several more presently advancing through clinical trials. Plant‐made pharmaceuticals have proven to be highly scalable and safe in humans, despite unfounded concerns regarding the impact of plant‐specific glycosylation. Limitations of expression yields have been addressed with improved expression vectors enabling high levels of expression of many pharmaceutically relevant proteins.
Although historically glycoproteins have been difficult to produce in planta, a number of different viral glycoproteins have been successfully expressed in plants, including most notably influenza HA and HIV Env, and several glycoproteins from emerging pathogens (D'Aoust et al., 2008). Fundamental differences in plants have been addressed by the manipulation of the secretory pathway, such as by glycoengineering, or the co‐expression of various other proteins to modify the host cell environment (Goulet et al., 2012; Jutras et al., 2015; Steinkellner and Castilho, 2015). Whilst many of the considerations discussed in this review are important for the production of viral glycoproteins in plants, it is likely that additional bottlenecks still remain and that these bottlenecks may even vary for different viral glycoproteins. Once these factors are identified, appropriate strategies can be conceived to expand the range of viral glycoproteins produced in planta. In conclusion, the use of molecular farming for the production of viral glycoproteins is an exciting and rapidly growing application of the technology with the potential to provide urgently needed vaccines and diagnostics for regions where they are needed the most.
Conflict of interest
The authors have filed patent applications protecting the production of HIV envelope trimers in plants (PA166256P) and the co‐expression of human molecular chaperones in plants to improve the production of heterologous proteins (PA167643_P).
Acknowledgements
The authors acknowledge the South African Medical Research Council Strategic Health Innovation Partnerships initiative for funding.
References
- Andrabi, R. , Voss, J.E. , Liang, C.H. , Briney, B. , McCoy, L.E. , Wu, C.Y. , Wong, C.H. et al. (2015) Identification of common features in prototype broadly neutralizing antibodies to HIV envelope V2 apex to facilitate vaccine design. Immunity, 43, 959–973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andrabi, R. , Su, C.Y. , Liang, C.H. , Shivatare, S.S. , Briney, B. , Voss, J.E. , Nawazi, S.K. et al. (2017) Glycans function as anchors for antibodies and help drive HIV broadly neutralizing antibody development. Immunity, 47, 524–537. e523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashraf, S. , Singh, P.K. , Yadav, D.K. , Shahnawaz, M. , Mishra, S. , Sawant, S.V. and Tuli, R. (2005) High level expression of surface glycoprotein of rabies virus in tobacco leaves and its immunoprotective activity in mice. J. Biotechnol. 119, 1–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bachmann, M.F. and Jennings, G.T. (2010) Vaccine delivery: a matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 10, 787–796. [DOI] [PubMed] [Google Scholar]
- Balla‐Jhagjhoorsingh, S.S. , Corti, D. , Heyndrickx, L. , Willems, E. , Vereecken, K. , Davis, D. and Vanham, G. (2013) The N276 glycosylation site is required for HIV‐1 neutralization by the CD4 binding site specific HJ16 monoclonal antibody. PLoS ONE, 8, e68863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bardor, M. , Faveeuw, C. , Fitchette, A.C. , Gilbert, D. , Galas, L. , Trottein, F. , Faye, L. et al. (2003) Immunoreactivity in mammals of two typical plant glyco‐epitopes, core alpha(1,3)‐fucose and core xylose. Glycobiology, 13, 427–434. [DOI] [PubMed] [Google Scholar]
- Beasley, D.W. and Barrett, A.D. (2002) Identification of neutralizing epitopes within structural domain III of the West Nile virus envelope protein. J. Virol. 76, 13097–13100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berinstein, A. , Vazquez‐Rovere, C. , Asurmendi, S. , Gomez, E. , Zanetti, F. , Zabal, O. , Tozzini, A. et al. (2005) Mucosal and systemic immunization elicited by Newcastle disease virus (NDV) transgenic plants as antigens. Vaccine, 23, 5583–5589. [DOI] [PubMed] [Google Scholar]
- Binley, J.M. , Sanders, R.W. , Clas, B. , Schuelke, N. , Master, A. , Guo, Y. , Kajumo, F. et al. (2000) A recombinant human immunodeficiency virus type 1 envelope glycoprotein complex stabilized by an intermolecular disulfide bond between the gp120 and gp41 subunits is an antigenic mimic of the trimeric virion‐associated structure. J. Virol. 74, 627–643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Binley, J.M. , Sanders, R.W. , Master, A. , Cayanan, C.S. , Wiley, C.L. , Schiffner, L. , Travis, B. et al. (2002) Enhancing the proteolytic maturation of human immunodeficiency virus type 1 envelope glycoproteins. J. Virol. 76, 2606–2616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blattner, C. , Lee, J.H. , Sliepen, K. , Derking, R. , Falkowska, E. , de la Pena, A.T. , Cupo, A. et al. (2014) Structural delineation of a quaternary, cleavage‐dependent epitope at the gp41‐gp120 interface on intact HIV‐1 Env trimers. Immunity, 40, 669–680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Boer, S.M. , Kortekaas, J. , Antonis, A.F. , Kant, J. , van Oploo, J.L. , Rottier, P.J. , Moormann, R.J. et al. (2010) Rift Valley fever virus subunit vaccines confer complete protection against a lethal virus challenge. Vaccine, 28, 2330–2339. [DOI] [PubMed] [Google Scholar]
- Bonomelli, C. , Doores, K.J. , Dunlop, D.C. , Thaney, V. , Dwek, R.A. , Burton, D.R. , Crispin, M. et al. (2011) The glycan shield of HIV is predominantly oligomannose independently of production system or viral clade. PLoS ONE, 6, e23521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bosch, D. and Schots, A. (2010) Plant glycans: friend or foe in vaccine development? Expert. Rev. Vaccines, 9, 835–842. [DOI] [PubMed] [Google Scholar]
- Braakman, I. and Hebert, D.N. (2013) Protein folding in the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 5, a013201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bullough, P.A. , Hughson, F.M. , Skehel, J.J. and Wiley, D.C. (1994) Structure of influenza haemagglutinin at the pH of membrane fusion. Nature, 371, 37–43. [DOI] [PubMed] [Google Scholar]
- Cao, L. , Diedrich, J.K. , Kulp, D.W. , Pauthner, M. , He, L. , Park, S.R. , Sok, D. et al. (2017) Global site‐specific N‐glycosylation analysis of HIV envelope glycoprotein. Nat. Commun. 8, 14954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardona‐Ospina, J.A. , Sepulveda‐Arias, J.C. , Mancilla, L. and Gutierrez‐Lopez, L.G. (2016) Plant expression systems, a budding way to confront chikungunya and Zika in developing countries? F1000Res, 5, 2121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castilho, A. and Steinkellner, H. (2012) Glyco‐engineering in plants to produce human‐like N‐glycan structures. Biotechnol. J. 7, 1088–1098. [DOI] [PubMed] [Google Scholar]
- Castilho, A. , Strasser, R. , Stadlmann, J. , Grass, J. , Jez, J. , Gattinger, P. , Kunert, R. et al. (2010) In planta protein sialylation through overexpression of the respective mammalian pathway. J. Biol. Chem. 285, 15923–15930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castilho, A. , Neumann, L. , Gattinger, P. , Strasser, R. , Vorauer‐Uhl, K. , Sterovsky, T. , Altmann, F. et al. (2013) Generation of biologically active multi‐sialylated recombinant human EPOFc in plants. PLoS ONE, 8, e54836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Castilho, A. , Beihammer, G. , Pfeiffer, C. , Goritzer, K. , Montero‐Morales, L. , Vavra, U. , Moresch, D. et al. (2018) An oligosacchyaryl transferase from Leishmania major increases the N‐glycan occupancy on recombinant glycoproteins produced in Nicotiana benthamiana . Plant Biotechnol. J. 10.1111/pbi.12906, In press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, B.J. , Leser, G.P. , Morita, E. and Lamb, R.A. (2007) Influenza virus hemagglutinin and neuraminidase, but not the matrix protein, are required for assembly and budding of plasmid‐derived virus‐like particles. J. Virol. 81, 7111–7123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen, T.‐H. , Hu, C.‐C. , Liao, J.‐T. , Lee, Y.‐L. , Huang, Y.‐W. , Lin, N.‐S. , Lin, Y.‐L. et al. (2017) Production of Japanese encephalitis virus antigens in plants using bamboo mosaic virus‐based vector. Front. Microbiol. 8, 788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chichester, J.A. , Jones, R.M. , Green, B.J. , Stow, M. , Miao, F. , Moonsammy, G. , Streatfield, S.J. et al. (2012) Safety and immunogenicity of a plant‐produced recombinant hemagglutinin‐based influenza vaccine (HAI‐05) derived from A/Indonesia/05/2005 (H5N1) influenza virus: a phase 1 randomized, double‐blind, placebo‐controlled, dose‐escalation study in healthy adults. Viruses, 4, 3227–3244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crill, W.D. and Roehrig, J.T. (2001) Monoclonal antibodies that bind to domain III of dengue virus E glycoprotein are the most efficient blockers of virus adsorption to Vero cells. J. Virol. 75, 7769–7773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crispin, M. , Ward, A.B. and Wilson, I.A. (2018) Structure and immune recognition of the HIV Glycan shield. Annu. Rev. Biophys. 47, 499–523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crooks, E.T. , Tong, T. , Osawa, K. and Binley, J.M. (2011) Enzyme digests eliminate nonfunctional Env from HIV‐1 particle surfaces, leaving native Env trimers intact and viral infectivity unaffected. J. Virol. 85, 5825–5839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cummings, J.F. , Guerrero, M.L. , Moon, J.E. , Waterman, P. , Nielsen, R.K. , Jefferson, S. , Gross, F.L. et al. (2014) Safety and immunogenicity of a plant‐produced recombinant monomer hemagglutinin‐based influenza vaccine derived from influenza A (H1N1)pdm09 virus: a Phase 1 dose‐escalation study in healthy adults. Vaccine, 32, 2251–2259. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Daniels, R. , Kurowski, B. , Johnson, A.E. and Hebert, D.N. (2003) N‐linked glycans direct the cotranslational folding pathway of influenza hemagglutinin. Mol. Cell. 11, 79–90. [DOI] [PubMed] [Google Scholar]
- D'Aoust, M.A. , Lavoie, P.O. , Couture, M.M. , Trepanier, S. , Guay, J.M. , Dargis, M. , Mongrand, S. et al. (2008) Influenza virus‐like particles produced by transient expression in Nicotiana benthamiana induce a protective immune response against a lethal viral challenge in mice. Plant Biotechnol. J. 6, 930–940. [DOI] [PubMed] [Google Scholar]
- D'Aoust, M.A. , Couture, M.M. , Charland, N. , Trepanier, S. , Landry, N. , Ors, F. and Vezina, L.P. (2010) The production of hemagglutinin‐based virus‐like particles in plants: a rapid, efficient and safe response to pandemic influenza. Plant Biotechnol. J. 8, 607–619. [DOI] [PubMed] [Google Scholar]
- Decroly, E. , Vandenbranden, M. , Ruysschaert, J.M. , Cogniaux, J. , Jacob, G.S. , Howard, S.C. , Marshall, G. et al. (1994) The convertases furin and PC1 can both cleave the human immunodeficiency virus (HIV)‐1 envelope glycoprotein gp160 into gp120 (HIV‐1 SU) and gp41 (HIV‐I TM). J. Biol. Chem. 269, 12240–12247. [PubMed] [Google Scholar]
- Decroly, E. , Wouters, S. , Di Bello, C. , Lazure, C. , Ruysschaert, J.M. and Seidah, N.G. (1996) Identification of the paired basic convertases implicated in HIV gp160 processing based on in vitro assays and expression in CD4(+) cell lines. J. Biol. Chem. 271, 30442–30450. [DOI] [PubMed] [Google Scholar]
- Doores, K.J. , Bonomelli, C. , Harvey, D.J. , Vasiljevic, S. , Dwek, R.A. , Burton, D.R. , Crispin, M. et al. (2010) Envelope glycans of immunodeficiency virions are almost entirely oligomannose antigens. Proc. Natl Acad. Sci. U. S. A. 107, 13800–13805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dugan, V. (2017) Update: Influenza Activity — United States, October 1–November 25, 2017. In: Morbidity and Mortality Weekly Report (MMWR). Centers for Disease Control and Prevention. [DOI] [PMC free article] [PubMed]
- Edgue, G. , Twyman, R.M. , Beiss, V. , Fischer, R. and Sack, M. (2017) Antibodies from plants for bionanomaterials. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 9, c1462. [DOI] [PubMed] [Google Scholar]
- Falkowska, E. , Le, K.M. , Ramos, A. , Doores, K.J. , Lee, J.H. , Blattner, C. , Ramirez, A. et al. (2014) Broadly neutralizing HIV antibodies define a glycan‐dependent epitope on the prefusion conformation of gp41 on cleaved envelope trimers. Immunity, 40, 657–668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faye, L. , Boulaflous, A. , Benchabane, M. , Gomord, V. and Michaud, D. (2005) Protein modifications in the plant secretory pathway: current status and practical implications in molecular pharming. Vaccine, 23, 1770–1778. [DOI] [PubMed] [Google Scholar]
- Fischer, R. , Schumann, D. , Zimmermann, S. , Drossard, J. , Sack, M. and Schillberg, S. (1999) Expression and characterization of bispecific single‐chain Fv fragments produced in transgenic plants. Eur. J. Biochem. 262, 810–816. [DOI] [PubMed] [Google Scholar]
- Frickel, E.M. , Frei, P. , Bouvier, M. , Stafford, W.F. , Helenius, A. , Glockshuber, R. and Ellgaard, L. (2004) ERp57 is a multifunctional thiol‐disulfide oxidoreductase. J. Biol. Chem. 279, 18277–18287. [DOI] [PubMed] [Google Scholar]
- Galloway, S.E. , Reed, M.L. , Russell, C.J. and Steinhauer, D.A. (2013) Influenza HA subtypes demonstrate divergent phenotypes for cleavage activation and pH of fusion: implications for host range and adaptation. PLoS Pathog. 9, e1003151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Georgiev, I.S. , Joyce, M.G. , Yang, Y. , Sastry, M. , Zhang, B. , Baxa, U. , Chen, R.E. et al. (2015) Single‐chain soluble BG505.SOSIP gp140 trimers as structural and antigenic mimics of mature closed HIV‐1 Env. J. Virol. 89, 5318–5329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghiasi, S.M. , Salmanian, A.H. , Chinikar, S. and Zakeri, S. (2011) Mice orally immunized with a transgenic plant expressing the glycoprotein of Crimean‐Congo hemorrhagic fever virus. Clin. Vaccine Immunol. 18, 2031–2037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Go, E.P. , Hua, D. and Desaire, H. (2014) Glycosylation and disulfide bond analysis of transiently and stably expressed clade C HIV‐1 gp140 trimers in 293T cells identifies disulfide heterogeneity present in both proteins and differences in O‐linked glycosylation. J. Proteome Res. 13, 4012–4027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez, E. , Zoth, S.C. , Asurmendi, S. , Rovere, C.V. and Berinstein, A. (2009) Expression of Hemagglutinin‐Neuraminidase glycoprotein of Newcastle Disease Virus in agroinfiltrated Nicotiana benthamiana plants. J. Biotechnol. 144, 337–340. [DOI] [PubMed] [Google Scholar]
- Gorman, J. , Soto, C. , Yang, M.M. , Davenport, T.M. , Guttman, M. , Bailer, R.T. , Chambers, M. et al. (2016) Structures of HIV‐1 Env V1V2 with broadly neutralizing antibodies reveal commonalities that enable vaccine design. Nat. Struct. Mol. Biol. 23, 81–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goulet, C. , Khalf, M. , Sainsbury, F. , D'Aoust, M.A. and Michaud, D. (2012) A protease activity‐depleted environment for heterologous proteins migrating towards the leaf cell apoplast. Plant Biotechnol. J. 10, 83–94. [DOI] [PubMed] [Google Scholar]
- Gu, M. , Rappaport, J. and Leppla, S.H. (1995) Furin is important but not essential for the proteolytic maturation of gp160 of HIV‐1. FEBS Lett. 365, 95–97. [DOI] [PubMed] [Google Scholar]
- Guerrero‐Andrade, O. , Loza‐Rubio, E. , Olivera‐Flores, T. , Fehervari‐Bone, T. and Gomez‐Lim, M.A. (2006) Expression of the Newcastle disease virus fusion protein in transgenic maize and immunological studies. Transgenic Res. 15, 455–463. [DOI] [PubMed] [Google Scholar]
- Hajj Hussein, I. , Chams, N. , Chams, S. , El Sayegh, S. , Badran, R. , Raad, M. , Gerges‐Geagea, A. et al. (2015) Vaccines through centuries: major cornerstones of global health. Front. Public Health, 3, 269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hallenberger, S. , Bosch, V. , Angliker, H. , Shaw, E. , Klenk, H.D. and Garten, W. (1992) Inhibition of furin‐mediated cleavage activation of HIV‐1 glycoprotein gp160. Nature, 360, 358–361. [DOI] [PubMed] [Google Scholar]
- He, J. , Peng, L. , Lai, H. , Hurtado, J. , Stahnke, J. and Chen, Q. (2014) A plant‐produced antigen elicits potent immune responses against West Nile virus in mice. Biomed. Res. Int. 2014, 952865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hefferon, K. (2013) Plant‐derived pharmaceuticals for the developing world. Biotechnol. J. 8, 1193–1202. [DOI] [PubMed] [Google Scholar]
- Hegde, R.S. and Bernstein, H.D. (2006) The surprising complexity of signal sequences. Trends Biochem. Sci. 31, 563–571. [DOI] [PubMed] [Google Scholar]
- Herve, P.L. , Lorin, V. , Jouvion, G. , Da Costa, B. and Escriou, N. (2015) Addition of N‐glycosylation sites on the globular head of the H5 hemagglutinin induces the escape of highly pathogenic avian influenza A H5N1 viruses from vaccine‐induced immunity. Virology, 486, 134–145. [DOI] [PubMed] [Google Scholar]
- Hiller, M.M. , Finger, A. , Schweiger, M. and Wolf, D.H. (1996) ER degradation of a misfolded luminal protein by the cytosolic ubiquitin‐proteasome pathway. Science, 273, 1725–1728. [DOI] [PubMed] [Google Scholar]
- Horn, M.E. , Pappu, K.M. , Bailey, M.R. , Clough, R.C. , Barker, M. , Jilka, J.M. , Howard, J.A. et al. (2003) Advantageous features of plant‐based systems for the development of HIV vaccines. J. Drug Target. 11, 539–545. [DOI] [PubMed] [Google Scholar]
- Howell, S.H. (2013) Endoplasmic reticulum stress responses in plants. Annu. Rev. Plant Biol. 64, 477–499. [DOI] [PubMed] [Google Scholar]
- Hsu, T.A. and Betenbaugh, M.J. (1997) Coexpression of molecular chaperone BiP improves immunoglobulin solubility and IgG secretion from Trichoplusia ni insect cells. Biotechnol. Prog. 13, 96–104. [DOI] [PubMed] [Google Scholar]
- Iuliano, A.D. , Roguski, K.M. , Chang, H.H. , Muscatello, D.J. , Palekar, R. , Tempia, S. , Cohen, C. et al. (2017) Estimates of global seasonal influenza‐associated respiratory mortality: a modelling study. Lancet, 391, 1285–1300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jardine, J. , Julien, J.P. , Menis, S. , Ota, T. , Kalyuzhniy, O. , McGuire, A. , Sok, D. et al. (2013) Rational HIV immunogen design to target specific germline B cell receptors. Science, 340, 711–716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen, T.J. , Loo, M.A. , Pind, S. , Williams, D.B. , Goldberg, A.L. and Riordan, J.R. (1995) Multiple proteolytic systems, including the proteasome, contribute to CFTR processing. Cell, 83, 129–135. [DOI] [PubMed] [Google Scholar]
- Jin, C. , Altmann, F. , Strasser, R. , Mach, L. , Schahs, M. , Kunert, R. , Rademacher, T. et al. (2008) A plant‐derived human monoclonal antibody induces an anticarbohydrate immune response in rabbits. Glycobiology, 18, 235–241. [DOI] [PubMed] [Google Scholar]
- Job, E.R. , Deng, Y.M. , Barfod, K.K. , Tate, M.D. , Caldwell, N. , Reddiex, S. , Maurer‐Stroh, S. et al. (2013) Addition of glycosylation to influenza A virus hemagglutinin modulates antibody‐mediated recognition of H1N1 2009 pandemic viruses. J. Immunol. 190, 2169–2177. [DOI] [PubMed] [Google Scholar]
- Joyce, M.G. , Zhang, B. , Ou, L. , Chen, M. , Chuang, G.Y. , Druz, A. , Kong, W.P. et al. (2016) Iterative structure‐based improvement of a fusion‐glycoprotein vaccine against RSV. Nat. Struct. Mol. Biol. 23, 811–820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Julien, J.P. , Cupo, A. , Sok, D. , Stanfield, R.L. , Lyumkis, D. , Deller, M.C. , Klasse, P.J. et al. (2013) Crystal structure of a soluble cleaved HIV‐1 envelope trimer. Science, 342, 1477–1483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jutras, P.V. , D'Aoust, M.A. , Couture, M.M. , Vezina, L.P. , Goulet, M.C. , Michaud, D. and Sainsbury, F. (2015) Modulating secretory pathway pH by proton channel co‐expression can increase recombinant protein stability in plants. Biotechnol. J. 10, 1478–1486. [DOI] [PubMed] [Google Scholar]
- Kaaijk, P. and Luytjes, W. (2017) Are we prepared for emerging flaviviruses in Europe? Challenges for vaccination. Hum. Vaccin. Immunother. 14, 337–344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalbina, I. , Lagerqvist, N. , Moiane, B. , Ahlm, C. , Andersson, S. , Strid, A. and Falk, K.I. (2016) Arabidopsis thaliana plants expressing Rift Valley fever virus antigens: mice exhibit systemic immune responses as the result of oral administration of the transgenic plants. Protein Expr. Purif. 127, 61–67. [DOI] [PubMed] [Google Scholar]
- Kallolimath, S. and Steinkellner, H. (2015) Glycosylation of plant produced human antibodies. Hum. Antibodies, 23, 45–48. [DOI] [PubMed] [Google Scholar]
- Kallolimath, S. , Castilho, A. , Strasser, R. , Grunwald‐Gruber, C. , Altmann, F. , Strubl, S. , Galuska, C.E. et al. (2016) Engineering of complex protein sialylation in plants. Proc. Natl Acad. Sci. U. S. A. 113, 9498–9503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanagaraj, A.P. , Verma, D. and Daniell, H. (2011) Expression of dengue‐3 premembrane and envelope polyprotein in lettuce chloroplasts. Plant Mol. Biol. 76, 323–333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Karczewski, J. and Ysibov, V. (2016) Plant‐based technologies to enable rapid response to Ebola outbreak.
- Karlsson Hedestam, G.B. , Fouchier, R.A. , Phogat, S. , Burton, D.R. , Sodroski, J. and Wyatt, R.T. (2008) The challenges of eliciting neutralizing antibodies to HIV‐1 and to influenza virus. Nat. Rev. Microbiol. 6, 143–155. [DOI] [PubMed] [Google Scholar]
- Kessans, S.A. , Linhart, M.D. , Matoba, N. and Mor, T. (2013) Biological and biochemical characterization of HIV‐1 Gag/dgp41 virus‐like particles expressed in Nicotiana benthamiana. Plant Biotechnol. J. 11, 681–690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, T.G. , Kim, M.Y. and Yang, M.S. (2010) Cholera toxin B subunit‐domain III of dengue virus envelope glycoprotein E fusion protein production in transgenic plants. Protein Expr. Purif. 74, 236–241. [DOI] [PubMed] [Google Scholar]
- Kim, M.Y. , Yang, M.S. and Kim, T.G. (2012) Expression of a consensus dengue virus envelope protein domain III in transgenic callus of rice. Plant Cell. Tiss. Org. 109, 509–515. [Google Scholar]
- Kong, L. , Sheppard, N.C. , Stewart‐Jones, G.B. , Robson, C.L. , Chen, H. , Xu, X. , Krashias, G. et al. (2010) Expression‐system‐dependent modulation of HIV‐1 envelope glycoprotein antigenicity and immunogenicity. J. Mol. Biol. 403, 131–147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kong, L. , Lee, J.H. , Doores, K.J. , Murin, C.D. , Julien, J.P. , McBride, R. , Liu, Y. et al. (2013) Supersite of immune vulnerability on the glycosylated face of HIV‐1 envelope glycoprotein gp120. Nat. Struct. Mol. Biol. 20, 796–803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Land, A. , Zonneveld, D. and Braakman, I. (2003) Folding of HIV‐1 envelope glycoprotein involves extensive isomerization of disulfide bonds and conformation‐dependent leader peptide cleavage. FASEB J. 17, 1058–1067. [DOI] [PubMed] [Google Scholar]
- Landry, N. , Ward, B.J. , Trepanier, S. , Montomoli, E. , Dargis, M. , Lapini, G. and Vezina, L.P. (2010) Preclinical and clinical development of plant‐made virus‐like particle vaccine against avian H5N1 influenza. PLoS ONE, 5, e15559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landry, N. , Pillet, S. , Favre, D. , Poulin, J.F. , Trepanier, S. , Yassine‐Diab, B. and Ward, B.J. (2014) Influenza virus‐like particle vaccines made in Nicotiana benthamiana elicit durable, poly‐functional and cross‐reactive T cell responses to influenza HA antigens. Clin. Immunol. 154, 164–177. [DOI] [PubMed] [Google Scholar]
- Lannoo, N. and Van Damme, E.J. (2015) Review/N‐glycans: the making of a varied toolbox. Plant Sci. 239, 67–83. [DOI] [PubMed] [Google Scholar]
- Lasky, L.A. , Groopman, J.E. , Fennie, C.W. , Benz, P.M. , Capon, D.J. , Dowbenko, D.J. , Nakamura, G.R. et al. (1986) Neutralization of the AIDS retrovirus by antibodies to a recombinant envelope glycoprotein. Science, 233, 209–212. [DOI] [PubMed] [Google Scholar]
- Le Mauff, F. , Mercier, G. , Chan, P. , Burel, C. , Vaudry, D. , Bardor, M. , Vezina, L.P. et al. (2015) Biochemical composition of haemagglutinin‐based influenza virus‐like particle vaccine produced by transient expression in tobacco plants. Plant Biotechnol. J. 13, 717–725. [DOI] [PubMed] [Google Scholar]
- Leonard, C.K. , Spellman, M.W. , Riddle, L. , Harris, R.J. , Thomas, J.N. and Gregory, T.J. (1990) Assignment of intrachain disulfide bonds and characterization of potential glycosylation sites of the type 1 recombinant human immunodeficiency virus envelope glycoprotein (gp120) expressed in Chinese hamster ovary cells. J. Biol. Chem. 265, 10373–10382. [PubMed] [Google Scholar]
- Li, Y. , Bergeron, J.J. , Luo, L. , Ou, W.J. , Thomas, D.Y. and Kang, C.Y. (1996) Effects of inefficient cleavage of the signal sequence of HIV‐1 gp 120 on its association with calnexin, folding, and intracellular transport. Proc. Natl Acad. Sci. U. S. A. 93, 9606–9611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, Y. , Luo, L. , Thomas, D.Y. and Kang, C.Y. (2000) The HIV‐1 Env protein signal sequence retards its cleavage and down‐regulates the glycoprotein folding. Virology, 272, 417–428. [DOI] [PubMed] [Google Scholar]
- Li, H.Y. , Ramalingam, S. and Chye, M.L. (2006) Accumulation of recombinant SARS‐CoV spike protein in plant cytosol and chloroplasts indicate potential for development of plant‐derived oral vaccines. Exp. Biol. Med. (Maywood) 231, 1346–1352. [DOI] [PubMed] [Google Scholar]
- Lippincott‐Schwartz, J. , Bonifacino, J.S. , Yuan, L.C. and Klausner, R.D. (1988) Degradation from the endoplasmic reticulum: disposing of newly synthesized proteins. Cell, 54, 209–220. [DOI] [PubMed] [Google Scholar]
- Liu, J.X. and Howell, S.H. (2010) Endoplasmic reticulum protein quality control and its relationship to environmental stress responses in plants. Plant Cell. 22, 2930–2942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loza‐Rubio, E. , Rojas, E. , Gomez, L. , Olivera, M.T. and Gomez‐Lim, M.A. (2008) Development of an edible rabies vaccine in maize using the Vnukovo strain. Dev. Biol. (Basel) 131, 477–482. [PubMed] [Google Scholar]
- Loza‐Rubio, E. , Rojas‐Anaya, E. , Lopez, J. , Olivera‐Flores, M.T. , Gomez‐Lim, M. and Tapia‐Perez, G. (2012) Induction of a protective immune response to rabies virus in sheep after oral immunization with transgenic maize, expressing the rabies virus glycoprotein. Vaccine, 30, 5551–5556. [DOI] [PubMed] [Google Scholar]
- Lyumkis, D. , Julien, J.P. , de Val, N. , Cupo, A. , Potter, C.S. , Klasse, P.J. , Burton, D.R. et al. (2013) Cryo‐EM structure of a fully glycosylated soluble cleaved HIV‐1 envelope trimer. Science, 342, 1484–1490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ma, J.K. , Christou, P. , Chikwamba, R. , Haydon, H. , Paul, M. , Ferrer, M.P. , Ramalingam, S. et al. (2013) Realising the value of plant molecular pharming to benefit the poor in developing countries and emerging economies. Plant Biotechnol. J. 11, 1029–1033. [DOI] [PubMed] [Google Scholar]
- Major, D. , Chichester, J.A. , Pathirana, R.D. , Guilfoyle, K. , Shoji, Y. , Guzman, C.A. , Yusibov, V. et al. (2015) Intranasal vaccination with a plant‐derived H5 HA vaccine protects mice and ferrets against highly pathogenic avian influenza virus challenge. Hum. Vacc. Immunother. 11, 1235–1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manini, I. , Trombetta, C. , Lazzeri, G. , Pozzi, T. , Rossi, S. and Montomoli, E. (2017) Egg‐Independent Influenza Vaccines and Vaccine Candidates. Vaccines, 5, 18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manolova, V. , Flace, A. , Bauer, M. , Schwarz, K. , Saudan, P. and Bachmann, M.F. (2008) Nanoparticles target distinct dendritic cell populations according to their size. Eur. J. Immunol. 38, 1404–1413. [DOI] [PubMed] [Google Scholar]
- Margolin, E. , Chapman, R. , Meyers, A.E. , van Diepen, M. , Ximba, P. , Hermanus, T. , et al. (submitted for publication) Production and immunogenicity of soluble plant‐produced HIV‐1 subtype C Envelope gp140 immunogens. [DOI] [PMC free article] [PubMed]
- Martinez, C.A. , Topal, E. , Giulietti, A.M. , Talou, J.R. and Mason, H. (2010) Exploring different strategies to express Dengue virus envelope protein in a plant system. Biotechnol. Lett. 32, 867–875. [DOI] [PubMed] [Google Scholar]
- Mbewana, S. (2017) The development of Rift Valley fever virus candidate vaccines and reagents produced in Nicotiana benthamiana. Molecular & Cell Biology. University of Cape Town. http://open.UCT.ac.za/handle/11427/25446.
- McGarvey, P.B. , Hammond, J. , Dienelt, M.M. , Hooper, D.C. , Fu, Z.F. , Dietzschold, B. , Koprowski, H. et al. (1995) Expression of the rabies virus glycoprotein in transgenic tomatoes. Biotechnology (N Y) 13, 1484–1487. [DOI] [PubMed] [Google Scholar]
- McGuire, A.T. , Hoot, S. , Dreyer, A.M. , Lippy, A. , Stuart, A. , Cohen, K.W. , Jardine, J. et al. (2013) Engineering HIV envelope protein to activate germline B cell receptors of broadly neutralizing anti‐CD4 binding site antibodies. J. Exp. Med. 210, 655–663. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Medlock, J. , Pandey, A. , Parpia, A.S. , Tang, A. , Skrip, L.A. and Galvani, A.P. (2017) Effectiveness of UNAIDS targets and HIV vaccination across 127 countries. Proc. Natl Acad. Sci. U. S. A. 114, 4017–4022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mett, V. , Musiychuk, K. , Bi, H. , Farrance, C.E. , Horsey, A. , Ugulava, N. , Shoji, Y. et al. (2008) A plant‐produced influenza subunit vaccine protects ferrets against virus challenge. Influenza Other Respir. Viruses, 2, 33–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore, P.L. , Crooks, E.T. , Porter, L. , Zhu, P. , Cayanan, C.S. , Grise, H. , Corcoran, P. et al. (2006) Nature of nonfunctional envelope proteins on the surface of human immunodeficiency virus type 1. J. Virol. 80, 2515–2528. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mortimer, E. , Maclean, J.M. , Mbewana, S. , Buys, A. , Williamson, A.L. , Hitzeroth, I.I. and Rybicki, E.P. (2012) Setting up a platform for plant‐based influenza virus vaccine production in South Africa. BMC Biotechnol. 12, 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moustafa, K. , Makhzoum, A. and Tremouillaux‐Guiller, J. (2016) Molecular farming on rescue of pharma industry for next generations. Crit. Rev. Biotechnol. 36, 840–850. [DOI] [PubMed] [Google Scholar]
- Nabel, G.J. (2013) Designing tomorrow's vaccines. N. Engl. J. Med. 368, 551–560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oliver, J.D. , van der Wal, F.J. , Bulleid, N.J. and High, S. (1997) Interaction of the thiol‐dependent reductase ERp57 with nascent glycoproteins. Science, 275, 86–88. [DOI] [PubMed] [Google Scholar]
- Oliver, J.D. , Roderick, H.L. , Llewellyn, D.H. and High, S. (1999) ERp57 functions as a subunit of specific complexes formed with the ER lectins calreticulin and calnexin. Mol. Biol. Cell. 10, 2573–2582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Owens, R.J. and Compans, R.W. (1990) The human immunodeficiency virus type 1 envelope glycoprotein precursor acquires aberrant intermolecular disulfide bonds that may prevent normal proteolytic processing. Virology, 179, 827–833. [DOI] [PubMed] [Google Scholar]
- Paccalet, T. , Bardor, M. , Rihouey, C. , Delmas, F. , Chevalier, C. , D'Aoust, M.A. , Faye, L. et al. (2007) Engineering of a sialic acid synthesis pathway in transgenic plants by expression of bacterial Neu5Ac‐synthesizing enzymes. Plant Biotechnol. J. 5, 16–25. [DOI] [PubMed] [Google Scholar]
- Pauthner, M. , Havenar‐Daughton, C. , Sok, D. , Nkolola, J.P. , Bastidas, R. , Boopathy, A.V. , Carnathan, D.G. et al. (2017) Elicitation of Robust Tier 2 Neutralizing Antibody Responses in Nonhuman Primates by HIV Envelope Trimer Immunization Using Optimized Approaches. Immunity, 46, 1073–1088. e1076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pera, F.F. , Mutepfa, D.L. , Khan, A.M. , Els, J.H. , Mbewana, S. , van Dijk, A.A. , Rybicki, E.P. et al. (2015) Engineering and expression of a human rotavirus candidate vaccine in Nicotiana benthamiana. Virol. J. 12, 205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phoolcharoen, W. , Bhoo, S.H. , Lai, H. , Ma, J. , Arntzen, C.J. , Chen, Q. and Mason, H.S. (2011) Expression of an immunogenic Ebola immune complex in Nicotiana benthamiana. Plant Biotechnol. J. 9, 807–816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pillet, S. , Racine, T. , Nfon, C. , Di Lenardo, T.Z. , Babiuk, S. , Ward, B.J. , Kobinger, G.P. et al. (2015) Plant‐derived H7 VLP vaccine elicits protective immune response against H7N9 influenza virus in mice and ferrets. Vaccine, 33, 6282–6289. [DOI] [PubMed] [Google Scholar]
- Pillet, S. , Aubin, E. , Trepanier, S. , Bussiere, D. , Dargis, M. , Poulin, J.F. , Yassine‐Diab, B. et al. (2016) A plant‐derived quadrivalent virus like particle influenza vaccine induces cross‐reactive antibody and T cell response in healthy adults. Clin. Immunol. 168, 72–87. [DOI] [PubMed] [Google Scholar]
- Pogrebnyak, N. , Golovkin, M. , Andrianov, V. , Spitsin, S. , Smirnov, Y. , Egolf, R. and Koprowski, H. (2005) Severe acute respiratory syndrome (SARS) S protein production in plants: development of recombinant vaccine. Proc. Natl Acad. Sci. U. S. A. 102, 9062–9067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard, L.K. , Vasiljevic, S. , Ozorowski, G. , Seabright, G.E. , Cupo, A. , Ringe, R. , Kim, H.J. et al. (2015) Structural constraints determine the glycosylation of HIV‐1 envelope trimers. Cell. Rep. 11, 1604–1613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quan, F.S. , Lee, Y.T. , Kim, K.H. , Kim, M.C. and Kang, S.M. (2016) Progress in developing virus‐like particle influenza vaccines. Expert. Rev. Vaccines, 15, 1281–1293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ringe, R.P. , Sanders, R.W. , Yasmeen, A. , Kim, H.J. , Lee, J.H. , Cupo, A. , Korzun, J. et al. (2013) Cleavage strongly influences whether soluble HIV‐1 envelope glycoprotein trimers adopt a native‐like conformation. Proc. Natl Acad. Sci. U. S. A. 110, 18256–18261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosales‐Mendoza, S. , Salazar‐Gonzalez, J.A. , Decker, E.L. and Reski, R. (2016) Implications of plant glycans in the development of innovative vaccines. Expert. Rev. Vaccines, 15, 915–925. [DOI] [PubMed] [Google Scholar]
- Rosenberg, Y. , Sack, M. , Montefiori, D. , Forthal, D. , Mao, L. , Hernandez‐Abanto, S. , Urban, L. et al. (2013) Rapid high‐level production of functional HIV broadly neutralizing monoclonal antibodies in transient plant expression systems. PLoS ONE, 8, e58724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rudolph, R. and Lilie, H. (1996) In vitro folding of inclusion body proteins. FASEB J. 10, 49–56. [PubMed] [Google Scholar]
- Rutkevich, L.A. , Cohen‐Doyle, M.F. , Brockmeier, U. and Williams, D.B. (2010) Functional relationship between protein disulfide isomerase family members during the oxidative folding of human secretory proteins. Mol. Biol. Cell. 21, 3093–3105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rybicki, E.P. (2009) Plant‐produced vaccines: promise and reality. Drug Discov. Today 14, 16–24. [DOI] [PubMed] [Google Scholar]
- Rybicki, E.P. (2010) Plant‐made vaccines for humans and animals. Plant Biotechnol. J. 8, 620–637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rybicki, E.P. (2014) Plant based vaccines against viruses. Virology Journal 11, 205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rybicki, E.P. (2017) Plant‐made vaccines and reagents for the One Health initiative. Hum. Vaccin. Immunother. 13, 2912–2917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rybicki, E.P. , Hitzeroth, I.I. , Meyers, A. , Dus Santos, M.J. and Wigdorovitz, A. (2013) Developing country applications of molecular farming: case studies in South Africa and Argentina. Curr. Pharm. Des. 19, 5612–5621. [DOI] [PubMed] [Google Scholar]
- Saejung, W. , Fujiyama, K. , Takasaki, T. , Ito, M. , Hori, K. , Malasit, P. , Watanabe, Y. et al. (2007) Production of dengue 2 envelope domain III in plant using TMV‐based vector system. Vaccine, 25, 6646–6654. [DOI] [PubMed] [Google Scholar]
- Saibil, H. (2013) Chaperone machines for protein folding, unfolding and disaggregation. Nat. Rev. Mol. Cell. Biol. 14, 630–642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, R.W. and Moore, J.P. (2017) Native‐like Env trimers as a platform for HIV‐1 vaccine design. Immunol. Rev. 275, 161–182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, R.W. , Venturi, M. , Schiffner, L. , Kalyanaraman, R. , Katinger, H. , Lloyd, K.O. , Kwong, P.D. et al. (2002) The mannose‐dependent epitope for neutralizing antibody 2G12 on human immunodeficiency virus type 1 glycoprotein gp120. J. Virol. 76, 7293–7305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, R.W. , Derking, R. , Cupo, A. , Julien, J.P. , Yasmeen, A. , de Val, N. , Kim, H.J. et al. (2013) A next‐generation cleaved, soluble HIV‐1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non‐neutralizing antibodies. PLoS Pathog. 9, e1003618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scanlan, C.N. , Pantophlet, R. , Wormald, M.R. , Ollmann Saphire, E. , Stanfield, R. , Wilson, I.A. , Katinger, H. et al. (2002) The broadly neutralizing anti‐human immunodeficiency virus type 1 antibody 2G12 recognizes a cluster of alpha1–>2 mannose residues on the outer face of gp120. J. Virol. 76, 7306–7321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seveno, M. , Bardor, M. , Paccalet, T. , Gomord, V. , Lerouge, P. and Faye, L. (2004) Glycoprotein sialylation in plants? Nat. Biotechnol. 22, 1351–1352; author reply 1352–1353. [DOI] [PubMed] [Google Scholar]
- Shao, Y. and Williamson, C. (2012) The HIV‐1 epidemic: low‐ to middle‐income countries. Cold Spring Harb. Perspect Med. 2, a007187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharma, S.K. , de Val, N. , Bale, S. , Guenaga, J. , Tran, K. , Feng, Y. , Dubrovskaya, V. et al. (2015) Cleavage‐independent HIV‐1 Env trimers engineered as soluble native spike mimetics for vaccine design. Cell. Rep. 11, 539–550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shoji, Y. , Chichester, J.A. , Bi, H. , Musiychuk, K. , de la Rosa, P. , Goldschmidt, L. , Horsey, A. et al. (2008) Plant‐expressed HA as a seasonal influenza vaccine candidate. Vaccine, 26, 2930–2934. [DOI] [PubMed] [Google Scholar]
- Shoji, Y. , Bi, H. , Musiychuk, K. , Rhee, A. , Horsey, A. , Roy, G. , Green, B. et al. (2009a) Plant‐derived hemagglutinin protects ferrets against challenge infection with the A/Indonesia/05/05 strain of avian influenza. Vaccine, 27, 1087–1092. [DOI] [PubMed] [Google Scholar]
- Shoji, Y. , Farrance, C.E. , Bi, H. , Shamloul, M. , Green, B. , Manceva, S. , Rhee, A. et al. (2009b) Immunogenicity of hemagglutinin from A/Bar‐headed Goose/Qinghai/1A/05 and A/Anhui/1/05 strains of H5N1 influenza viruses produced in Nicotiana benthamiana plants. Vaccine, 27, 3467–3470. [DOI] [PubMed] [Google Scholar]
- Shoji, Y. , Chichester, J.A. , Jones, M. , Manceva, S.D. , Damon, E. , Mett, V. , Musiychuk, K. et al. (2011) Plant‐based rapid production of recombinant subunit hemagglutinin vaccines targeting H1N1 and H5N1 influenza. Hum. Vaccin. 7(Suppl), 41–50. [DOI] [PubMed] [Google Scholar]
- Shoji, Y. , Farrance, C.E. , Bautista, J. , Bi, H. , Musiychuk, K. , Horsey, A. , Park, H. et al. (2012) A plant‐based system for rapid production of influenza vaccine antigens. Influenza Other Respir. Viruses, 6, 204–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sijmons, P.C. , Dekker, B.M. , Schrammeijer, B. , Verwoerd, T.C. , van den Elzen, P.J. and Hoekema, A. (1990) Production of correctly processed human serum albumin in transgenic plants. Biotechnology (N Y) 8, 217–221. [DOI] [PubMed] [Google Scholar]
- Skehel, J.J. , Stevens, D.J. , Daniels, R.S. , Douglas, A.R. , Knossow, M. , Wilson, I.A. and Wiley, D.C. (1984) A carbohydrate side chain on hemagglutinins of Hong Kong influenza viruses inhibits recognition by a monoclonal antibody. Proc. Natl Acad. Sci. U. S. A. 81, 1779–1783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sok, D. , Doores, K.J. , Briney, B. , Le, K.M. , Saye‐Francisco, K.L. , Ramos, A. , Kulp, D.W. et al. (2014) Promiscuous glycan site recognition by antibodies to the high‐mannose patch of gp120 broadens neutralization of HIV. Sci. Transl. Med. 6, 236ra263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sok, D. , Pauthner, M. , Briney, B. , Lee, J.H. , Saye‐Francisco, K.L. , Hsueh, J. , Ramos, A. et al. (2016) A Prominent Site of Antibody Vulnerability on HIV Envelope Incorporates a Motif Associated with CCR5 Binding and Its Camouflaging Glycans. Immunity, 45, 31–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sommer, T. and Jentsch, S. (1993) A protein translocation defect linked to ubiquitin conjugation at the endoplasmic reticulum. Nature, 365, 176–179. [DOI] [PubMed] [Google Scholar]
- Steinhauer, D.A. (1999) Role of hemagglutinin cleavage for the pathogenicity of influenza virus. Virology, 258, 1–20. [DOI] [PubMed] [Google Scholar]
- Steinkellner, H. and Castilho, A. (2015) N‐glyco‐engineering in plants: update on strategies and major achievements. Methods Mol. Biol. 1321, 195–212. [DOI] [PubMed] [Google Scholar]
- Stieneke‐Grober, A. , Vey, M. , Angliker, H. , Shaw, E. , Thomas, G. , Roberts, C. , Klenk, H.D. et al. (1992) Influenza virus hemagglutinin with multibasic cleavage site is activated by furin, a subtilisin‐like endoprotease. EMBO J. 11, 2407–2414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoger, E. , Fischer, R. , Moloney, M. and Ma, J.K. (2014) Plant molecular pharming for the treatment of chronic and infectious diseases. Annu. Rev. Plant Biol. 65, 743–768. [DOI] [PubMed] [Google Scholar]
- Strasser, R. (2018) Protein Quality Control in the Endoplasmic Reticulum of Plants. Annu. Rev. Plant Biol. 69, 147–172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strasser, R. , Stadlmann, J. , Schahs, M. , Stiegler, G. , Quendler, H. , Mach, L. , Glossl, J. et al. (2008) Generation of glyco‐engineered Nicotiana benthamiana for the production of monoclonal antibodies with a homogeneous human‐like N‐glycan structure. Plant Biotechnol. J. 6, 392–402. [DOI] [PubMed] [Google Scholar]
- Strasser, R. , Altmann, F. and Steinkellner, H. (2014) Controlled glycosylation of plant‐produced recombinant proteins. Curr. Opin. Biotechnol. 30, 95–100. [DOI] [PubMed] [Google Scholar]
- Streatfield, S.J. , Kushnir, N. and Yusibov, V. (2015) Plant‐produced candidate countermeasures against emerging and reemerging infections and bioterror agents. Plant Biotechnol. J. 13, 1136–1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tessier, D.C. , Thomas, D.Y. , Khouri, H.E. , Laliberte, F. and Vernet, T. (1991) Enhanced secretion from insect cells of a foreign protein fused to the honeybee melittin signal peptide. Gene, 98, 177–183. [DOI] [PubMed] [Google Scholar]
- Tottey, S. , Shoji, Y. , Jones, R.M. , Chichester, J.A. , Green, B.J. , Musiychuk, K. , Si, H. et al. (2018) Plant‐produced subunit vaccine candidates against yellow fever induce virus neutralizing antibodies and confer protection against viral challenge in animal models. Am. J. Trop. Med. Hyg. 98, 420–431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tran, K. , Poulsen, C. , Guenaga, J. , de Val, N. , Wilson, R. , Sundling, C. , Li, Y. et al. (2014) Vaccine‐elicited primate antibodies use a distinct approach to the HIV‐1 primary receptor binding site informing vaccine redesign. Proc. Natl Acad. Sci. U. S. A. 111, E738–E747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trkola, A. , Purtscher, M. , Muster, T. , Ballaun, C. , Buchacher, A. , Sullivan, N. , Srinivasan, K. et al. (1996) Human monoclonal antibody 2G12 defines a distinctive neutralization epitope on the gp120 glycoprotein of human immunodeficiency virus type 1. J. Virol. 70, 1100–1108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tschofen, M. , Knopp, D. , Hood, E. and Stoger, E. (2016) Plant Molecular Farming: Much More than Medicines. Annu. Rev. Anal. Chem. (Palo Alto Calif) 9, 271–294. [DOI] [PubMed] [Google Scholar]
- Vollenweider, F. , Benjannet, S. , Decroly, E. , Savaria, D. , Lazure, C. , Thomas, G. , Chretien, M. et al. (1996) Comparative cellular processing of the human immunodeficiency virus (HIV‐1) envelope glycoprotein gp160 by the mammalian subtilisin/kexin‐like convertases. Biochem. J. 314(Pt 2), 521–532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada, I. , Rindress, D. , Cameron, P.H. , Ou, W.J. , Doherty, J.J. 2nd , Louvard, D. , Bell, A.W. et al. (1991) SSR alpha and associated calnexin are major calcium binding proteins of the endoplasmic reticulum membrane. J. Biol. Chem. 266, 19599–19610. [PubMed] [Google Scholar]
- Ward, C.L. , Omura, S. and Kopito, R.R. (1995) Degradation of CFTR by the ubiquitin‐proteasome pathway. Cell, 83, 121–127. [DOI] [PubMed] [Google Scholar]
- Ward, B.J. , Landry, N. , Trepanier, S. , Mercier, G. , Dargis, M. , Couture, M. , D'Aoust, M.A. et al. (2014) Human antibody response to N‐glycans present on plant‐made influenza virus‐like particle (VLP) vaccines. Vaccine, 32, 6098–6106. [DOI] [PubMed] [Google Scholar]
- Wei, X. , Decker, J.M. , Wang, S. , Hui, H. , Kappes, J.C. , Wu, X. , Salazar‐Gonzalez, J.F. et al. (2003) Antibody neutralization and escape by HIV‐1. Nature, 422, 307–312. [DOI] [PubMed] [Google Scholar]
- Wei, C.J. , Boyington, J.C. , Dai, K. , Houser, K.V. , Pearce, M.B. , Kong, W.P. , Yang, Z.Y. et al. (2010) Cross‐neutralization of 1918 and 2009 influenza viruses: role of glycans in viral evolution and vaccine design. Sci. Transl. Med. 2, 24ra21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WHO . (2014) Influenza vaccine viruses and reagents. Geneva: World Health Organisation. [Google Scholar]
- Wilbers, R.H. , Westerhof, L.B. , van Raaij, D.R. , van Adrichem, M. , Prakasa, A.D. , Lozano‐Torres, J.L. , Bakker, J. et al. (2016) Co‐expression of the protease furin in Nicotiana benthamiana leads to efficient processing of latent transforming growth factor‐beta1 into a biologically active protein. Plant Biotechnol. J. 14, 1695–1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yakob, L. and Walker, T. (2016) Zika virus outbreak in the Americas: the need for novel mosquito control methods. Lancet Glob. Health, 4, e148–e149. [DOI] [PubMed] [Google Scholar]
- Yang, M. , Lai, H. , Sun, H. and Chen, Q. (2017a) Virus‐like particles that display Zika virus envelope protein domain III induce potent neutralizing immune responses in mice. Sci. Rep. 7, 7679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, M. , Sun, H. , Lai, H. , Hurtado, J. and Chen, Q. (2017b) Plant‐produced Zika virus envelope protein elicits neutralizing immune responses that correlate with protective immunity against Zika virus in mice. Plant Biotechnol. J. 16, 572–580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yap, Y.K. and Smith, D.R. (2010) Strategies for the plant‐based expression of dengue subunit vaccines. Biotechnol. Appl. Biochem. 57, 47–53. [DOI] [PubMed] [Google Scholar]
- Zabel, F. , Kundig, T.M. and Bachmann, M.F. (2013) Virus‐induced humoral immunity: on how B cell responses are initiated. Curr. Opin. Virol. 3, 357–362. [DOI] [PubMed] [Google Scholar]
- Zapun, A. , Darby, N.J. , Tessier, D.C. , Michalak, M. , Bergeron, J.J. and Thomas, D.Y. (1998) Enhanced catalysis of ribonuclease B folding by the interaction of calnexin or calreticulin with ERp57. J. Biol. Chem. 273, 6009–6012. [DOI] [PubMed] [Google Scholar]
- Zhao, H. , Fernandez, E. , Dowd, K.A. , Speer, S.D. , Platt, D.J. , Gorman, M.J. , Govero, J. et al. (2016) Structural Basis of Zika Virus‐Specific Antibody Protection. Cell, 166, 1016–1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou, J.Y. , Cheng, L.Q. , Zheng, X.J. , Wu, J.X. , Shang, S.B. , Wang, J.Y. and Chen, J.G. (2004) Generation of the transgenic potato expressing full‐length spike protein of infectious bronchitis virus. J. Biotechnol. 111, 121–130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zumla, A. , Dar, O. , Kock, R. , Muturi, M. , Ntoumi, F. , Kaleebu, P. , Eusebio, M. et al. (2016) Taking forward a ‘One Health’ approach for turning the tide against the Middle East respiratory syndrome coronavirus and other zoonotic pathogens with epidemic potential. Int. J. Infect. Dis. 47, 5–9. [DOI] [PMC free article] [PubMed] [Google Scholar]