

Abstract
In the hair follicle, the dermal papilla (DP) and dermal sheath (DS) support and maintain proliferation and differentiation of the epithelial stem cells that produce the hair fibre. In view of their regulatory properties, in this study, we investigated the interaction between hair follicle dermal cells (DP and DS) and embryonic stem cells (ESCs); induced pluripotent stem cells (iPSCs); and haematopoietic stem cells. We found that coculture of follicular dermal cells with ESCs or iPSCs supported their prolonged maintenance in an apparently undifferentiated state as established by differentiation assays, immunocytochemistry, and RT-PCR for markers of undifferentiated ESCs. We further showed that cytokines that are involved in ESC support are also expressed by cultured follicle dermal cells, providing a possible explanation for maintenance of ES cell stemness in cocultures. The same cytokines were expressed within follicles in situ in a pattern more consistent with a role in follicle growth activities than stem cell maintenance. Finally, we show that cultured mouse follicle dermal cells provide good stromal support for haematopoiesis in an established coculture model. Human follicular dermal cells represent an accessible and readily propagated source of feeder cells for pluripotent and haematopoietic cells and have potential for use in clinical applications.
1. Introduction
Adult hair follicle dermal cell populations have extensive regenerative, inductive, and supportive capabilities, both within adult and developing hair follicles [1, 2] and in combination with other cell types including cornea and amnion [3, 4]. Experimentally, subpopulations of adult hair follicle dermal cells have demonstrated extensive stem cell capabilities, and multipotency, including generation of bone, fat, and muscle in vitro [5–7]. Additionally, dermal cells can differentiate down a haematopoietic lineage both in vivo and in vitro, [8, 9] and have characteristics similar to embryonic neural crest stem cells [10]. In this respect, they behave similarly to stem cell populations isolated from adult bone marrow, a common source of adult stem cells [11]. It is interesting to note that cells isolated from adult bone marrow also have supportive capabilities, particularly in the support of hematopoietic stem cells and embryonic stem cells (ESCs) in vitro [12–14]. Bone marrow cells support epidermal keratinocytes in in vitro skin reconstitution assays [15] and during cutaneous wound healing [16], demonstrating significant similarities with hair follicle dermal cells [17, 18].
ESCs, derived from the inner cell mass of mammalian blastocysts [19–21], retain their developmental potential after prolonged culture to differentiate down all three germ layer lineages in vivo and in vitro. Induced pluripotent stem cells (iPSCs), which are almost comparable to ESCs, are generated by reprogramming somatic cells, a process initially achieved using virus-mediated gene transduction of a few key factors [22, 23]. While mouse ESCs (mESCs) can be continuously cultured without feeder cells on gelatin-coated plates with the addition of leukemia inhibitory factor (LIF) [24], human ESCs (hESCs) or iPSCs will differentiate in culture in the presence of this cytokine. Propagation of undifferentiated human ESCs or iPSCs is commonly carried out by coculture with a mouse embryonic fibroblast (MEF) feeder layer. To improve the potential clinical utility of human pluripotent cells, considerable progress has been made in establishing defined feeder-free culture systems for hESCs or iPSCs [25–27], with methods that include growing the cells on specific substrates [28, 29], in suspension cultures [30] or in defined serum-free medium [31]. Another approach has been to replace MEFs with human-derived cells. For example, bone marrow stromal cells (BMSC) [13], neonatal skin fibroblasts, stromal cells [32–34], amniotic mesenchymal cells [35], or human foetal cell lines [36] have all been employed as feeder cells. However, the use of human feeder layers requires that the feeder cell type is easily accessible, readily propagated, and efficient at maintaining and amplifying undifferentiated hESCs suitable for clinical use.
Interactions between adjacent cells of different types are a major mechanism of organogenesis in developmental biology. In view of the inductive properties of hair follicle dermal cells [1, 2], we initially set out to investigate their effects on mESCs by coculture in vitro, anticipating that the dermal cells would exert some directive influence on mESC differentiation. However, we found that follicular dermal cells appeared to be effective in maintaining the mESCs in an undifferentiated state. This was confirmed by inducing differentiation of the mESCs along multiple lineages after prolonged coculture and by investigating the expression of markers characteristic of undifferentiated mESCs by RT-PCR and immunofluorescence. We subsequently investigated the mechanism by which dermal cultures may be able to support mESCs by examining the expression of members of the IL-6 family of cytokines, known to be crucial for maintaining murine embryonic stem cell pluripotency in vitro via the gp130 receptor and the JAK/STAT pathway. Parallel investigations were also performed on follicles, based on the hypothesis that follicle epithelial stem cells might be maintained in an undifferentiated state by ES cell-type mechanisms. This was not supported by the observations, but the prevalence of IL-6 family cytokines and the gp130 receptor in follicles did point to a functional role of gp130/JAK/STAT signalling in hair follicle activities. When the ability of human hair follicle dermal cells to maintain hESCs and hiPSCs in an undifferentiated state was assessed, it was confirmed that like their rodent cell counterparts, the follicle dermal cells were superior to skin fibroblasts in their ability to maintain and support hESC and iPSC cultures. Finally, given the apparent similarities between bone marrow stromal cells and hair follicle dermis/mesenchyme [17], we performed coculture experiments to investigate the ability of hair follicle dermal cells to support haematopoietic activity. Here again, the follicle cells were the equal if not better than bone marrow-derived stromal cells under the experimental conditions employed.
These observations have implications for the regulation of both dermal and epithelial stem cells in the hair follicle, as well as confirming that hair follicle dermal cells have the potential to be a useful source of feeder cells for the support and amplification of a range of stem cell types.
2. Materials and Methods
2.1. Hair Follicle DP and DS Cell Isolation and Culture
DP and DS were microdissected from the vibrissa follicles of adult PVG rats or BalbC or Zin40 mice as previously described [37]. Animal tissues were obtained from animals housed in accordance with the institutional guidelines at the University of Durham. Human DP and DS were microdissected from skin biopsies as previously described [2], with skin biopsies obtained as anonymised discarded tissue in accordance with Helsinki guidelines. Skin dermal fibroblast (SF) cultures were established as explants from finely minced rodent footpad or human interfollicular scalp skin. A spontaneously transformed rat dermal papilla cell line, RDP-B [38], was also used as a control line. Once established, cells were maintained in MEM (Sigma) supplemented with 10% FBS (Gibco) and antibiotics (Sigma) (dermal cell medium) at 37°C, 5% CO2, with passaging every 2–4 weeks.
2.2. Mouse ESC Culture
Mouse CGR8 ESCs were routinely cultured on mitomycin C-inactivated MEF feeder layers in Glasgow MEM supplemented with 10% FBS, 100 μM β-mercaptoethanol (Gibco), 2 mM L-glutamine, 1% nonessential amino acids, 0.25% NaHCO3, 1 mM pyruvate (Sigma), and 1000 U/ml LIF (Chemicon) (mESC medium). Cells were grown on 0.1% gelatin (Sigma) coated 6-well plates and split at ratios from 1 : 3 and 1 : 6 to 1 : 8 prior to becoming confluent. Two CGR8 cell lines transfected with GFP-expressing vectors were used in various assays; one designated CGR8-GFP, expressed GFP from a CAGG promoter which escaped silencing in mESCs, and was used to track both differentiated and undifferentiated mESCs in cocultures, while the second designated CGR8 Rex1-EGFP, expressed GFP under the control of the Rex-1 promoter, and was used to localize undifferentiated mESCs in cocultures. mESCs were stably transfected with the Rex 1-EGFP vector (a kind gift from Dr. N. Benvenisty) [39], using TransFast (Promega) in accordance with the manufacturer's protocol. CGR8 Rex1-EGFP cells were routinely maintained on MEF feeder layers in mESC medium with periodic reselection by addition of 400 μg/ml G418 to the culture medium.
2.3. 2D Coculture of mESCs with Rodent Dermal Cells
The CGR8 Rex1-EGFP and CGR8-GFP cells were used for coculture experiments. Initially, rodent DP, DS, or SF cells were plated at 4 × 105 cells per well of 6-well culture plates (the same cell density as MEFs for feeder layers). Subconfluent mESCs were then seeded over the feeder layer in each well. The rodent cocultures were passaged every two days with a splitting ratio of 1 : 6. A second set of experiments was performed in which rodent DP or DS monolayers were established in 35 mm culture dishes, and 500, 3000, 5000, or 10,000 Rex 1-EGFP mESCs were added to each dish when the dermal layer reached 80% confluence. These cocultures were then maintained for up to 4 weeks without splitting. A third set of cocultures was also performed but dermal cells were physically separated from mESCs by seeding them on porous membrane inserts (0.45 μm pore size, Falcon), placed over 6-well plates containing mESCs.
All cocultures were routinely maintained in dermal cell medium, without the addition of LIF, but some samples were also grown in ESC differentiation medium (mESC medium without LIF). At least 3 sets of dermal/ES cell cocultures were established, and in the case of the hair follicle dermal cells, experiments were repeated at least 6 times. Dermal cells used were between passage 2 and passage 7 with over 30% of them at passage 1 to passage 3.
2.4. Culture of mESCs with Dermal Cell-Conditioned Medium
To obtain conditioned medium, rodent dermal cells or dermal/ES cell cocultures were incubated with dermal cell medium for 48 hours. After this, time medium was collected and centrifuged (30 min, 3300 rpm) to remove cell debris and the supernatant were stored at −20°C for future use. mESCs were then cultured for 10 days in conditioned medium (CM) diluted 1 : 1 with ESC differentiation medium. The morphology of these cells was compared to mESCs cultured for the same period in mESC medium with or without LIF.
2.5. 3D Coculture of Rodent Cells
CGR8 mESCs and PVG rat DP or DS cells were mixed in three different ratios of mESC : dermal cells (1 : 1, 1 : 3, and 1 : 10). 3D cultures were generated using a hanging drop method; 10 μl drops of cell suspension in ESC differentiation medium containing approximately 400 mixed cells were placed on the lids of bacteriological Petri (nontissue culture-treated) dishes filled with phosphate buffered saline (PBS). Several sets of cocultures utilized follicular dermal cells stained with DiI and CGR8-GFP mESCs so that the mESC and follicular dermal cells could be distinguished in mixed cultures. After cultivation for 3 days, the aggregates were transferred into bacteriological Petri dishes and maintained in suspension in ESC differentiation medium. Differentiation was investigated by microscopy, immunofluorescence, and RT-PCR of the embryoid-like bodies after a further 9 days of culture.
2.6. Differentiation of mESC Cultures
CGR8-GFP mESCs were isolated from cocultures with DP or DS cells. The medium was removed from the cocultures and the cells were washed twice with PBS. A 30 μl volume of TVP solution (trypsin, versene, and chick plasma; Sigma) was taken up in a 200 μl automatic pipette tip and repeatedly expelled and aspirated over a small area of the coculture until gaps could be seen in the cell layer. The contents of the tip were then expelled into a gelatin-coated 35 mm culture dish, and the cells were briefly expanded in mESC medium prior to being plated at 0.54 × 106 cells per 90 mm bacteriological Petri dish to generate EBs. After a short suspension period, EBs could be observed; medium was changed every 2 days by centrifuging EBs for 3 minutes at 800 rpm and resuspending in fresh ESC differentiation medium. EBs were then exposed to different conditions to promote differentiation.
2.6.1. Neuronal Differentiation Assay
4 days after EB establishment, all-trans-retinoic acid (Sigma), diluted in ESC differentiation medium, was added to the suspension cultures (at a final concentration of 10−7 M). This was repeated on day 6. By day 8, approximately 50 EBs were transferred to gelatin-coated 60 mm dishes and incubated with ESC differentiation medium without β-mercaptoethanol (prohibits differentiation and is left out of the medium from this stage) for 2 days. 18 μM cytosine arabinoside (Ara-C) was added on day 10 and the dishes were incubated for a further 2 days to optimize neuron-like cell numbers [40]. Immunocytochemistry was carried out on differentiated cells (day 12 of the assay) with NF200, a primary antibody specific to neurofilaments.
2.6.2. Adipocyte Differentiation Assay
This was carried out according to established methods [41] and the resulting cultures were stained with oil red O to detect lipid [5].
2.6.3. Endoderm Differentiation Assay
6 days after their formation, EBs were plated onto 35 mm 0.1% gelatin-coated dishes and maintained in ESC differentiation medium. After 12 more days, with medium changes every second day, the cells were fixed and stained with antibodies against albumin and alpha-1-fetoprotein.
2.7. Human ESC and iPSC Culture
The human H9 ESC line and iPSC established and validated in our laboratory were used in this study. Cells were grown on mitomycin-inactivated MEFs with hESC medium containing KnockOut DMEM, 100 μM β-mercaptoethanol, 1 mM L-glutamine, 1% nonessential amino acids, 20% serum replacement (Invitrogen), 1% penicillin-streptomycin, and 8 ng/ml FGF2 (Invitrogen), which was changed daily.
2.8. Culture of hESCs and iPSCs on Dermal Feeder Layers from Human Skin
Initially, mitomycin-inactivated MEF or human DP, DS, and SF were seeded at 4 × 105 cells per well of 6-well culture plates in dermal medium. After 1 day, hESCs or iPSCs were seeded onto the feeders and fed every day with hESC medium. hESC and iPSC cells were passaged every 4-5 days by incubation in 1 mg/ml collagenase IV (Invitrogen) at 37°C or mechanically dissociated and then removed to freshly prepared feeders. Cultures were fixed and incubated with nitro blue tetrazolium and 5-bromo-4-chloro-3′-indolyphosphate (NBT/BCIP) substrate solution to detect alkaline phosphatase activity. For immunocytochemistry analysis, colonies were separated from their feeders and seeded onto chamber slides coated with ESC-qualified matrigel in MEF-conditioned hESC medium for 2 days prior to fixation and analysis.
2.9. Flow Cytometry of hESCs
For flow cytometry analysis, hESCs were collected using collagenase IV treatment (1 mg/ml for 5 minutes) followed by brief accutase incubation. Cells were suspended in staining buffer (PBS + 5% FCS) at 106 cells/ml. 105 cells were stained with TRA-1-60, SSEA-4 (Millipore), or Oct4 (Santa Cruz) antibodies at 10 μg/ml final concentration. Several washes were carried out in staining buffer before proceeding to staining with secondary antibodies. Cells were washed three times and resuspended in staining buffer before being analyzed with FACS Calibur (BD) using CellQuest. 10,000 events were acquired for each sample, and propidium iodide staining (1 μg/ml) was used to distinguish live from dead cells.
2.10. Reverse Transcription Polymerase Chain Reaction
Total RNA from 2D and 3D cocultures, rodent DP and DS cell cultures, mESC cultures with or without LIF, and dissected vibrissa follicles (end bulb, mid-follicle, and upper follicle) were prepared using the ToTALLY RNA Kit (Ambion) as described by the manufacturer.
Contaminating genomic DNA was eliminated by DNase I digestion (DNA-free kit, Ambion). Approximately 1 μg total RNA from each sample was reverse transcribed (Superscript II RT, Invitrogen) using oligo-dT primers. PCR was then performed using Taq polymerase (Invitrogen) with specific primer sets for each gene (Table 1). PCR reactions were carried out as follows: 94°C for 5 min; 20–30 cycles of 94°C for 30 s, gene-specific annealing temperature for 30 s, and 72°C for 60s; and 72°C for 5 min.
Table 1.
Primer sets for reverse transcription polymerase chain reaction.
| Name | Sequence | Annealing temp (°C) | Size (bp) |
|---|---|---|---|
| GAPDH | F: GCC AAA AGG GTC ATC ATC TC | 61 | 379 |
| R: ACG GAT ACA TTG GGG GTA GG | |||
| Oct4 | F: CCC GGA AGA GAA AGC GAA CT | 58 | 362 |
| R: GAC GGG AAC AGA GGG AAA GG | |||
| Nanog | F: AGG GTC TGC TAC TGA GAT GCT CTG | 56 | 363 |
| R: CAA CCA CTG GTT TTT CTG CCA CCG | |||
| LIF | F: ATT GTG CCC TTA CTG CTG CT | 61 | 583 |
| R: GCC TGG ACC ACC ACA CTT AT | |||
| CT-1 | F: GAG GAA TAC GTG CAG CAA CA | 57 | 389 |
| R: AGC ACC TTG GCT GAG AAG AT | |||
| OSM | F: CAC GGC TTC TAA GAA CAC TGC | 59 | 547 |
| R: CGA TGG TAT CCC CAG AGA AA | |||
| CNTF | F: CTT TCG CAG AGC AAT CAC CT | 61 | 579 |
| R: CCC CAT AAT GGC TCT CAT GT | |||
| BMP2 | F: TCC ATC ACG AAG AAG CCG TG | 58 | 465 |
| R: CCA AAA GTC ACT AGC AAT GGC | |||
| BMP4 | F: AGG GCC AGC ACG TCA GAA TC | 57 | 430 |
| R: ACC TTG TCA TAC TCA TCC AGG | |||
| CNTFRα | F: CTG TTT CCA CCG TGA CTC CT | 59 | 802 |
| R: TGG GAC ACT GGT CAA GAA GA | |||
| Nodal | F: GCC AGA CAG AAG CCA ACT GTG | 61 | 324 |
| R: TCA GAG GCA CCC ACA CTC CTC | |||
| GluR6 | F: CTG CAG CAC AGA GAG GAA CCA | 60 | 488 |
| R: ATA ACT TCC TCC ATG TGC CTC AC | |||
| Brachyury | F: GCT GAG ACT TGT AAC AAC CG | 55 | 266 |
| R: GCA AAG GAC TCT GAT TAA CTG C | |||
| TTR | F: CCG TTC CAT GAA TTC GCG GAT | 60 | 240 |
| R: TTC ACG GCA TCT TCC TGA GC | |||
| AFP | F: TTG CCT CCA CGT GCT GCC AGC | 61 | 341 |
| R: CGC CAG CTG CTC CTC TGT CAG |
2.11. Immunocytochemistry
Cultured cells were fixed in either methanol (2 min, −20°C) or 4% paraformaldehyde in PBS (10 min, room temperature), blocked against nonspecific binding, and permeabilized with 0.1% triton X prior to incubating with primary antibodies (Table 2) for 1 hr at room temperature or overnight at 4°C. Following primary antibody incubation, cells were rinsed in PBS and incubated with secondary antibodies for 1 hr at room temperature in the dark. Stained samples were mounted under glass coverslips in Mowiol (Calbiochem) and visualized with a Zeiss Axiovert 135 microscope. Isolated rat vibrissa follicles were frozen in OCT (Agar Scientific) in liquid nitrogen or embedded in paraffin wax after overnight 4°C paraformaldehyde fixation. Antibodies bound on paraffin-embedded tissues were visualized using the VECTASTAIN ABC-AP kit (goat IgG, Vector Laboratories).
Table 2.
Antibodies used during immunochemical analysis.
| Antibody | Manufacturer | Dilution used |
|---|---|---|
| Anti-albumin | Dako | 1 : 100 |
| Anti-alpha-1-fetoprotein | Dako | 1 : 100 |
| Anti-α-smooth muscle actin | Sigma | 1 : 10 |
| Goat anti-CNTF | R&D Systems | 1 : 40 |
| Anti-GFP | Abcam | 1 : 100 |
| Anti-gp130 | Santa Cruz | 1 : 30 |
| Goat anti-LIF | R&D Systems | 1 : 100 |
| Anti-NF200 | Sigma | 1 : 100 |
| Anti-OCT-3/4 | R&D Systems | 1 : 20 |
| Anti-Tra-1-60 | Millipore | 1 : 200 |
| Anti-Tra-1-81 | Millipore | 1 : 200 |
| FITC anti-mouse | DAKO | 1 : 80 |
| TRITC anti-mouse | Jackson Immuno | 1 : 100 |
| Alexa Fluor 546 anti-mouse | Molecular Probes | 1 : 500 |
| FITC anti-rabbit | DAKO | 1 : 100 |
| Alexa Fluor 546 anti-rabbit | Molecular Probes | 1 : 500 |
| TRITC anti-rat | Jackson Immuno | 1 : 100 |
| Alexa Fluor 594 anti-rat | Molecular Probes | 1 : 500 |
2.12. Bone Marrow Stromal Cell Culture
Mouse bone marrow stromal cell (BMSC) primary cultures were established as previously described [42]. The S17 stromal cell line is an immortalized cell line originally isolated from Dexter culture [43]. Cultures were routinely passaged as previously described and were plated into 35 mm dishes at p3 or p4 and allowed to grow to confluence.
2.13. Isolation and Purification of Haematopoietic Progenitors
Mice (6-week-old Balb/c) were killed by cervical dislocation, and bone marrow was collected from their femurs by flushing with PBS using a 25 g needle and 1 ml syringe. Red blood cells were lysed by incubation for 10 minutes in a hypotonic solution of ammonium chloride (7.5%) at room temperature, and the remaining nucleated cells were collected by centrifugation at 2000 rpm and washed 3 times in PBS. Viability was determined by Trypan blue exclusion, and cells were counted and resuspended in PBS. These were separated into c-kit-enriched or depleted populations by the magnetic activated cell sorter (MACS) system (Miltenyi Biotec, Bergisch-Gladbach, Germany).
2.14. Stromal Support Experiments
A summary of the strategy for these experiments is shown in the Supplementary Figure S1. Stromal support experiments used mouse DP and DS between passage 3 and passage 5 as stromal layers, with either primary mouse bone marrow stromal cells or S17 mouse stromal cell line as positive controls. All stromal cells were grown in MEM with 10% FCS. Confluent stromal layers were seeded with105 unfractionated bone marrow nucleated cells. Alternatively, they were seeded with the c-kit-enriched (MACS-bound) population or with the depleted (MACS-run through) population from 105 bone marrow nucleated cells. Cocultures were maintained for 28 days, and nonadherent cells were harvested every seven days and used for CFU-M assays. Experiments were repeated 6 times, cell numbers were entered into Excel, and standard deviations were calculated using Excel.
2.15. CFU-A Assays
CFU-A assays were performed essentially as previously described [44]. Briefly, support layers of 0.5 ml 0.6% agar in Alpha-MEM (Gibco-Invitrogen) containing 100 μl per ml of conditioned medium from the cell lines AF1-19T (a source of GM-CSF) and L929 (a source of M-CSF) were plated into the wells of a 24-well plate. Cells (2000/well) were overlaid in 0.5 ml of plating medium (identical to support layer, but using 0.15% agar). Plates were cultured for 20 days in 10% CO2, in humidified incubators. Experiments were repeated 6 times, colony numbers were entered into Excel, and standard deviations were calculated using Excel.
3. Results
3.1. Expansion of mESCs Cocultured on Rodent DP or DS Cells
When mESCs were cocultured with follicular dermal cells (Z40 or PVG in origin) in dermal cell medium, we observed growth of colonies of undifferentiated mESCs similar to those formed on MEFs when supplemented with LIF. The mESCs settled in discrete colonies on top of the dermal cell monolayer (Figures 1(a), 1(d), and 1(g)) and expressed EGFP under the control of Rex-1 (undifferentiated mESC marker, Figures 1(b), 1(e), and 1(h)). The rodent feeder cells were not mitotically arrested and were either split together with the mESCs every 2 or 3 days or maintained as a confluent layer under the mESC colonies. In multiple experiments, the longest continuous coculture of mESCs with rodent dermal cells was either 8 passages or 4 weeks (depending on the method used) before the cocultures were used for other tests. When mESCs were grown with rodent SF cultures, colonies became flatter and less distinct in appearance (Figure 1(a)), and lost Rex-1-directed GFP expression (Figure 1(b)) and Oct4 expression (Figure 1(c)). However, irrespective of the coculture methodology, mES cells grown on follicular dermal cells (DP or DS) showed no significant loss of colony morphology (Figures 1(d) and 1(g)), or Rex-1-directed GFP expression (Figures 1(e) and 1(h)), or evidence of differentiation as indicated by the pluripotency marker Oct4 (Figures 1(f) and 1(i)). After eight or more passages on rodent DP and DS cells, the expanded mES cells were separated from their cocultures and analyzed using RT-PCR for the gene expression of various ES pluripotency and differentiation markers. High levels of Oct4 and Nanog expression were detected in mESCs when cultured on DP and DS cells, but not when cultured without LIF or feeder layers (Figure 1(j)). mESCs cocultured with DP and DS cells in dermal medium (particularly with DS cells) did not express significant levels of differentiation markers such as Nodal, GLU-R6, Brachyury, AFP, and TTR at any point, whereas these were highly expressed in mESCs cultured in ES differentiation medium for 6–14 days.
Figure 1.

Maintenance of mESCs by rodent hair follicle dermal cells. Rex-1 EGFP-transfected CGR8 mESCs were cocultured with PVG fibroblasts (a–c), DP (d–f), and DS (g–i) cells. The ESCs lost colony morphology (a), Rex-1-directed EGFP expression (b), and Oct4 expression (c) when cultured with skin fibroblasts. However, mESCs were effectively maintained in an undifferentiated state by coculture with follicular dermal cells, both DP and DS, displaying compacted colonies (d, g), eGFP expression under the control of Rex-1 (e, h), and high levels of Oct4 expression (f, i). RT-PCR analysis of CGR8 mESCs cocultured with rat DP and DS cells (j). PCR products obtained using primers specific for mouse GAPDH (control), Oct4, Nanog (markers of undifferentiated ES cells), LIF, BMP4 (involved in ES support), Nodal, Glu-R6, Brachyury, TTR, and AFP (markers of differentiation). Scale bars = 50 μm.
3.2. The Supportive Role of Rodent DP and DS on mESCs Is Mediated via Soluble Factors
Having established that pluripotent mESCs could be maintained in an undifferentiated state by follicular dermal cells in coculture, we next investigated if this process was mediated by soluble factors. As previously observed, there are marked contrasts between mESCs maintained in complete mESC medium (dense colonies with just a few differentiated cells) and those cultured in ESC differentiation medium (numerous morphological changes) (Figures 2(a) and 2(b)). When we maintained mESCs in conditioned dermal medium from DP/ES cocultures, we found that this was most effective at maintaining undifferentiated mESC morphology, with DS/ES-, DP only-, and DS only-conditioned media being progressively less effective. However, in all conditioned medium cultures, a larger proportion of the mESCs maintained a typical mESC morphology than in the culture maintained in ESC differentiation medium (Figure 2(c)), suggesting that soluble factors secreted by the dermal cells (both in the presence and absence of ESCs) prevent differentiation of ESCs.
Figure 2.

Dermal cells provide support for mESCs via soluble factors. mESCs were cultured in complete mESC medium (a), ESC differentiation medium (b), and DP/mESC coculture conditioned medium (c) for 10 days. Cultures maintained in conditioned medium showed maintenance of typical mESC colony morphology and behaviour. (d) Cytokines known to be involved in mESC maintenance are expressed by follicular DP, DS, and BMSC. (e) Several of these cytokines are also expressed in different regions of vibrissa follicles as detected by PCR. However, the localization of LIF (f), CNTF (g), and gp130 (h) by immunohistochemistry (pink staining) in the lower follicle bulb region shows that they are predominantly in the follicle epithelium. (i) gp130 is also expressed in the upper follicle (i) specifically in the bulge epithelium (i∗) (region delineated by a yellow rectangle) (CL: club hair; GH: growing hair). Scale bars (a–i) = 100 μm and (i∗) = 50 μm.
3.3. Expression of ES Cell Supporting Cytokines In Vitro and In Vivo
In an attempt to determine which soluble factors may be involved in mediating mESC maintenance, RT-PCR was used to detect transcripts of potential candidates known to be involved in mESC support; Nanog, Oct4, LIF, CNTF, OSM, and CT-1 expression was examined in DP, DS, and BMSC which has previously been shown to support the expansion of undifferentiated ESCs [13]. Members of the bone morphogenetic family (BMP) have also been implicated in mESC regulation in combination with LIF [45]. BMP2 was detected in DP and DS cells, while BMP4 expression was typically lower in DP cells when compared with DS and BMSC (Figure 2(d)). Cultured rodent DP and DS cells expressed higher levels of LIF, CT-1, Nanog, and CNTF than BMSC, while Oct4 (at low levels) and OSM expression was seen in BMSC, but not in dermal cells. Conversely, Nanog was expressed in DP and DS cells but was not detectable in BMSC (Figure 2(d)).
To examine the in vivo expression of cytokines of interest, mid-anagen vibrissa follicles dissected from three Zin40 mice were used as a source of RNA. CNTF, LIF, CT-1, and the receptor component CNTFRα were expressed in all regions of mid-anagen follicles, while OSM was not expressed at comparable levels (Figure 2(e)). We investigated the localization of a subset of these cytokines in intact follicles by immunohistochemistry, and interestingly, we found that localization of LIF (Figure 2(f)) and CNTF (Figure 2(g)) was not only confined to dermal cell populations. Indeed, rather than being highly expressed in the DP and DS, LIF appeared to be concentrated in the epithelium immediately surrounding the DP, whereas CNTF appeared to have a more general distribution, with localized expression in the upper third of the DP and lower DS, coupled with higher levels of epithelial localization. Since the IL-6 family cytokines investigated here all act through a receptor complex containing gp130, we also examined gp130 distribution in intact follicles (Figure 2(h)). Again, this was present predominantly in the epithelial layers of the follicle, with distinct expression at the DP/epithelial boundary. Specifically in the bulge region higher up the follicle (Figure 2(i)), the epithelial cells showed strong gp130 immunolabelling (Figure 2(i∗)).
3.4. Follicular Dermal Cells Are Unable to Prevent ES Cell Differentiation in 3-Dimensional Cultures
Having demonstrated that follicular dermal cells are able to inhibit mESC differentiation in 2D cultures, we next investigated if they can prevent differentiation of mESCs in 3D cultures. EBs were produced containing either mESCs only or a 1 : 1, 3 : 1, or 10 : 1 ratio of follicular dermal to mESCs by hanging drop culture. During the early stages of EB culture (2–4 days), it was noted that the inclusion of follicular dermal cells tended to modulate EB formation, with EB's containing DP or DS cells being less regularly shaped than those comprising mESCs alone, and often, with more than one EB forming in each hanging drop (Figure 3(a)). Once EBs were transferred to suspension culture, they continued to grow for 9–12 days, often producing large cysts containing cardiomyocyte-like (beating) cells. We found that this remained the case no matter how many dermal cells were included, although a higher proportion of dermal cells generally resulted in smaller cysts. Follicular dermal cells alone were unable to form cysts and instead were maintained as tight clumps of cells with little or no increase in size. Within EBs, it appeared that dermal cells were not well distributed and instead were remaining aggregated (Figure 3(b)). Surprisingly, however, in all cases, the mESCs rapidly outgrew the dermal cells and expressed differentiation markers not present in 2D cocultures (Figure 3(c)). We found that neither the addition of LIF nor the addition of an increasing proportion of dermal cells to the EBs could prevent this (data not shown); so, for all subsequent experiments, we used a 1 : 1 ratio of dermal : ESCs for EB formation. The mESCs rapidly lost expression of Oct4 and Nanog, further confirming that undifferentiated mESCs were not maintained, although LIF expression was not substantially downregulated. We found that the pattern of differentiation markers expressed varied from that shown by differentiated mESCs in 2D culture (compare Figures 1(j) and 3(c), ESC only), with no evidence of GluR6 expression and significant loss of BMP4 expression by mESCs in the 3D cultures.
Figure 3.
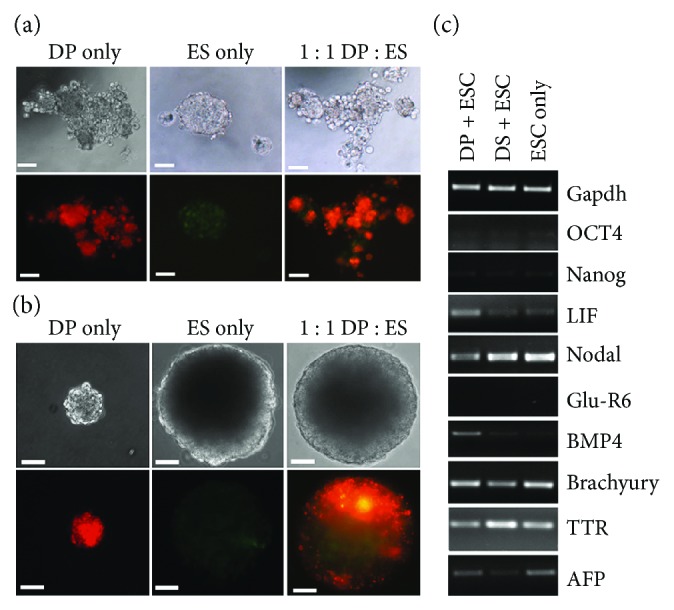
Coculture of dermal cells and mESCs in embryoid bodies does not prevent differentiation. (a) Embryoid-like bodies formed after 3 days in hanging drop culture. Follicular dermal cells were stained with DiI and mESCs expressing GFP under the control of the CAGG promoter. Hanging drops containing follicular dermal cells produced EBs having several foci of aggregation, in contrast to those containing ES cells alone. (b) EBs after 9 days in suspension culture. EBs containing mESCs were much larger than those with only follicular dermal cells. (c) Differentiation markers expressed by mESCs in 3 dimensional cocultures with follicular dermal cells. Scale bars (a) = 50 μm and (b) = 100 μm.
3.5. Cocultured mESCs Maintain Differentiation Potential in Culture
When mESCs were directly differentiated in the neuronal differentiation assay, we observed networks of neuron-like cells after 12 days in culture. Axon-like projections extended in networks from EBs, which in association with other cell types adhered to the culture substrate (Figure 4(a)). Cells originating from cocultures with PVG and Z40, DP, and DS cells all gave positive results in this assay, with a minimum of 65% of EBs producing networks of neuron-like cells.
Figure 4.
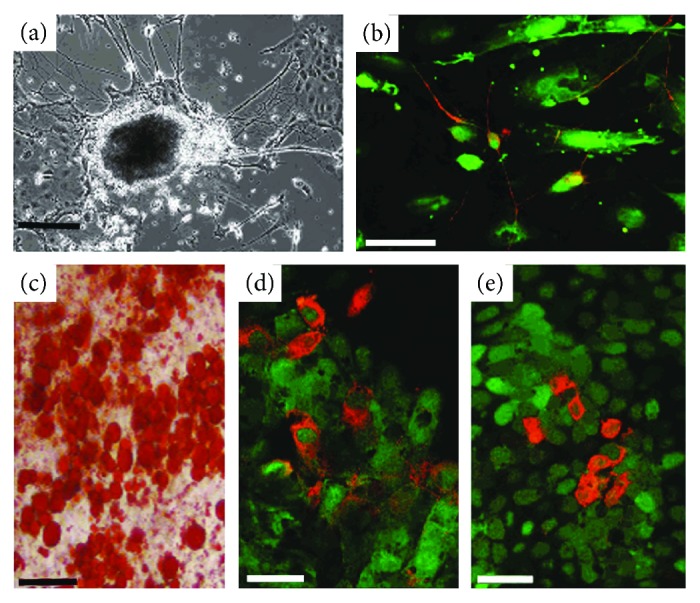
mESCs maintain pluripotency after coculture with dermal cells. (a) Networks of neurons extend from EBs derived from mESCs after 6-day coculture with PVG rat DS cells. (b) GFP mESCs after 6-day coculture with PVG DP cells produce neuron-like cells; the projections of which are visible by fluorescence microscopy. Immunostaining with anti-NF200 antibodies demonstrates expression of neurofilament (red) and the colocalization with the GFP fluorescence (green). (c) ESCs cocultured with PVG DS for 6 days produced high levels of lipid (red). One EB in particular was observed to be producing copious amounts of lipid. Endodermal cells are present in differentiated cells from 6-day cocultures of GFP mESCs with PVG DP (d) and Z40 DP (e). Green fluorescence identifies the cells as being of mESC origin while red fluorescence shows immunoreactivity for albumin (d) and AFP (e). Scale bars (a) = 200 μm, (b–e) = 50 μm, and (d) = 25 μm.
The greatest numbers of neuronal outgrowths were observed in cells originating from 6 day Z40 DS : mESC cocultures where 90% of EBs had neuron-like cells associated with them. Immunocytochemistry for neurofilament markers confirmed these cells to be neuronal, and colocalization with GFP also confirmed that they were derived from mESCs (Figure 4(b)).
In the adipocyte differentiation assay, mESCs isolated from PVG and Z40, DP, and DS cocultures gave consistently positive results. Throughout experimental cultures, oil red O staining showed small lipid droplets and large lipid-filled cells in patches ranging from a few cells to up to 50 cells closely packed together (Figure 4(c)). Approximately 30% of each culture exhibited high levels of lipid deposition.
In the endoderm differentiation assay, mESCs from all cocultures gave positive results. Small clusters of GFP-positive CGR8-GFP cells were immunoreactive for albumin (Figure 4(d)) and alpha-feta-1-protein (Figure 4(e)). As expected, these comprised no more than 1% of the total cell population.
3.6. Maintenance of hESCs and iPSCs by Hair Follicle Dermal Feeder Layers
After demonstrating that mESCs were maintained in an undifferentiated state by coculture with follicular dermal cells, we next asked if human hair follicle dermal cells could act as effective feeders for hESCs and iPSCs. When we cultured hESCs on mitotically inactive human DP or DS feeders for multiple passages, high levels of SSEA-4, TRA-1-60, and OCT4 expression were maintained, as determined by flow cytometry (Figure 5(a)). Although all DP and DS lines tested were able to maintain hESCs expressing high levels of these markers, it was evident that some lines were more effective than others. Immunolabelling of DP-supported hESCs with antibodies to NANOG and OCT4 showed levels of staining equivalent to those shown by hESCs cultured with MEFs (Figures 5(b)–5(e)). Subsequently, when we cultured human iPSCs on mitotically inactivated MEFs (Figures 6(a)–6(d)), human SF (Figures 6(e)–6(h)), DP (Figures 6(i)–6(l)), or DS (Figures 6(m)–6(p)), we found that they maintained a typical hESC-like appearance on the follicular dermal feeders, in discrete colonies with a high nuclear to cytoplasmic ratio. iPSC grown on MEFs, DP, or DS feeders for several passages remained positive for the pluripotency markers alkaline phosphatase, Tra-1-60, and Tra-1-81. Comparatively, expression of these markers was lost in iPSCs after a short period of growth on interfollicular SF feeders.
Figure 5.

Follicular dermal cells support hESCs. hESCs cocultured with either MEFs, human DP cells, or human DS cells showed comparable levels of expression of SSEA-4, TRA-1-60, and Oct4 (a), as determined by flow cytometry indicating that the DP and DS cells were as capable of maintaining hESCs in an undifferentiated state as MEFs. Although all DP and DS cell lines tested were able to maintain hESCs expressing high levels of these markers, it was evident that some lines were more effective than others. hESCs maintained by MEFs and labelled for NANOG (b) and OCT4 (c) have similar staining to hESCs maintained by DP cells and labelled for NANOG (d) and OCT4 (e). Scale bars = 100 μm.
Figure 6.

Hair follicle dermal cells maintain human iPSCs in an undifferentiated state. Human iPSCs were grown on either MEF, human SF, DP, or DS cells. On MEFs, iPSCs maintained normal ESC-like morphology (a), high alkaline phosphatase (b), Tra-1-60 (c), and Tra-1-81 (d) expression. In comparison, when grown on fibroblast feeders, the iPSC colonies became less compact (e), and with increasing passage number lost expression of pluripotency markers and alkaline phosphatase (f–h). When grown on hair follicle dermal cells, DP (i–l), but particularly DS (m–p), the iPSCs maintained normal ESC morphology and high levels of expression of ESC pluripotency markers for several passages in culture. Scale bars = 100 μm.
3.7. Hair Follicle Dermal Cells Support Haematopoietic Progenitors
All stromal cell types appeared to support haematopoietic cells, as indicated by the typical cobblestone morphology of the cells on the surface of the cultures (Supplementary Figure S2).
Cell counts revealed that DP and DS cultures were at least as effective as bone marrow stroma and S17 cell lines in supporting the proliferation of nonadherent cells in coculture experiments (Figure 7(a)). This was particularly the case for unfractionated or c-kit-enriched populations. Flow cytometry with CD45 confirmed that haematopoietic cells were being produced (data not shown), but this could have been due to mitosis of mature cell types. In order to investigate whether haematopoietic progenitors were being supported by the stromal cultures, the nonadherent cells were subjected to colony assay using CFU-A. Figure 7(b) shows that the number of CFU-A colonies produced following coculture with DP or DS cultures appeared greater than that from conventional Dexter-type bone marrow stromal cell culture. This increase in haematopoietic progenitor number was especially marked for the c-kit-enriched population.
Figure 7.

Hair follicle dermal cells support haematopoietic activity in coculture with murine bone marrow cells. (a) Total numbers of nonadherent cells produced by coculture of murine bone marrow cells with different stromal support cells over a 28-day period. N = 6 ± SD. (b) CFU-A colony production by coculture of murine bone marrow cells with stromal cells. Total burst colonies produced per sample over a 28-day period. No colonies were produced from c-kit fractions for any stromal condition. N = 6 ± SD. S-17: an immortalized stromal cell line; BMSC: bone marrow stromal cells; DP: dermal papilla cells; DS: dermal sheath cells.
4. Discussion
The initial hypothesis for the current work was that follicular dermal cells, when cocultured with pluripotent ESCs, might induce differentiation along the lineage of follicular epithelial cells. The inductive capacity of hair follicle dermal cells has been well documented [1, 2]. Nonfollicular epithelium will form follicles when associated with DP cells or embryonic dermis from hairy skin [3, 4], demonstrating the ability of DP cells to direct the differentiation of cells in close proximity, as is believed to be their physiological role in the adult hair follicle [1, 46]. In our hands, rat vibrissa follicle dermal papilla cells lose this inductive capacity around passage 4 in culture [47]. It has been shown that ESCs differentiate along both dermal and epidermal lineages to produce a tissue equivalent to embryonic skin when exposed to factors produced by skin fibroblasts [48]; so, it seemed reasonable to postulate that follicular dermis would induce differentiation along follicular lineages. In contrast to the working hypothesis, follicular dermal cells maintained both rodent and human ESCs and iPSCs in an undifferentiated state after long-term coculture. As both pre- and postinductive rat dermal papilla cultures showed the same influence on ES cells, it appeared that this phenomenon was not linked to loss of DP-inductive properties. The colonies of mESCs produced, either in mixed cocultures or in cultures where the dermal and ESCs were not in contact, were identical to those maintained by LIF or an MEF feeder layer (as confirmed by TEM (data not shown), immunocytochemistry, RT-PCR, and the use of Rex 1-EGFP CGR8 cells). Both human and mouse ESCs retained high levels of intrinsic Oct4 and Nanog expression, which are known to maintain pluripotency both in vivo and in vitro [49–51]. Human iPSCs cultured on follicular dermal cell feeders also retained high levels of the cell surface antigens Tra-1-60 and Tra-1-81, comparable to the levels expressed by iPSCs grown on MEF's, demonstrating that undifferentiated iPSCs can be effectively maintained by follicular dermal cells (both DP and DS). These markers of undifferentiated hESCs disappear rapidly upon differentiation [52], as was seen when the cells were grown on control fibroblast feeder layers. We also showed that mESCs retained their pluripotency after coculture by performing differentiation assays to induce differentiation into cell lineages derived from each of the three germ layers. The observation that in 3D coculture, follicle dermal cells were unable to prevent ES cell differentiation typical of embryoid bodies reflects the powerful influence of the 3D environment on ES behaviour and the fact that there was segregation of the two cell types within the structures. It may also be that in 3D, the dermal cells had a different secretory profile.
The behaviour of mESCs exposed to dermal cell- or coculture-conditioned media in 2D, or in cultures where the cells were physically separated by a 0.45 μM filter, indicated that soluble factors were present in the media secreted by the dermal cells (both in the presence and in the absence of ESCs), with the capacity to inhibit differentiation. A similar study identified secreted factors in MEF-conditioned medium that could maintain undifferentiated hESCs in the absence of feeder cells [53]. Four members of the IL-6 family of cytokines (LIF, CNTF, CT-1, and OSM) have been shown to maintain undifferentiated mESCs in vitro via the LIFR/gp130/STAT pathway [24, 54–58]. Therefore, we interrogated the mRNAs of cultures and found that transcripts of three of these cytokines, LIF, CNTF, and CT-1 were detected in dermal cells. Additionally, BMPs are known to cooperate with LIF to maintain mESC pluripotency [45], and BMP2 and BMP4 were detected in the hair follicle dermal cell culture. We also found that cultured DP and DS expressed Nanog, a downstream effector of the LIF/STAT3 pathway in maintaining mESC pluripotency [49]. Further, undifferentiated hESCs can be maintained without feeder cells by the presence of high levels of Nanog [51]. The expression of these cytokines in vitro raises the intriguing possibility that they may play a functional role in vivo. The localization of LIF and CNTF in the follicle end bulb suggests that they are physiologically relevant. Past studies have cited them as promoters and inhibitors of both differentiation and proliferation [27, 24, 54–57, 59–61], processes that occur predominantly in the follicle end bulb and are key to the cyclic nature of hair follicle activity. Moreover, we noted strong gp130 expression in the bulge region of the follicle outer root sheath, which houses the main epithelial stem cell population of the hair follicle. However, the relative lack of cytokine expression in the dermal papilla cells of the follicle bulb, but the presence of LIF, CNTF, and CT-1 mRNA in all segments of the mid-anagen follicle, indicates a widespread function within follicles, rather than a specific role in follicular epithelial stem cell maintenance. Similarly, the expression of CNTFRα mRNA throughout the follicle suggests that members of the IL-6 family are unlikely to be involved specifically in maintenance of stemness in the surrounding epithelial cells by the follicular dermis. It appears that the mechanisms by which the dermal cells support ES cell maintenance have no obvious parallels with regulation of epithelial stem cell activities in the follicle. The contribution of the above cytokines to various aspects of hair follicle biology remains to be fully defined although there are reported connections between the gp130/JAK/STAT pathway and follicle activities. Interleukin-6 itself has been linked with hair growth inhibition and follicle regression [62, 63]. Moreover, in mice, JAK-STAT3 signalling is needed for the initiation of spontaneous anagen [64]. Stat5 activation in the follicle DP has recently been shown to trigger follicle entry in the growing phase (anagen) [65], and one of us (AMC) has recently shown that pharmacologically inhibiting the JAK/STAT pathway induced anagen from resting (telogen) follicles [66].
The elimination of animal material during both the derivation and long-term culture of hESCs or iPSCs is an important goal prior to application of these cells for clinical therapy. Animal-derived feeders and serum risk the introduction and transfer of nonhuman pathogens to human cells and increase the the risk of graft rejection when cells are introduced into patients [67,68].
A wide range of feeder cell types, conditioned media, and feeder-free systems has been investigated [25, 34], including a xeno-free system for derivation of hESCs using human serum and a human foreskin fibroblast feeder layer [69]. Human hair follicle dermal cells could be similarly utilized for both hESC derivation and long-term maintenance. Moreover, iPSCs have recently been derived from mouse hair follicle DP cells using a single transcription factor [70], and we have derived iPSCs from human hair follicle DP cells [71]. A recent report has shown that human follicle dermal (mesenchymal) cells maintain hES cells in an undifferentiated condition [72]. Our work supports this finding and extends it to human iPS cells. Therefore, follicle dermal cells may parallel the properties of human and mouse adipose-derived cells that can be used both to establish iPSCs in a feeder-independent manner and as feeder cells for supporting different pluripotent stem cells [73].
Previous studies have shown strong parallels between hair follicle dermal cells and bone marrow cells [17]. Therefore, our findings here that hair follicle dermal cells support haematopoiesis are to some extent unsurprising. However, the dermal cells were apparently equal to and possibly superior to bone marrow cells as stromal supports. Since we previously demonstrated that cultured hair follicle dermal cells can produce colonies in CFU-A assays and restore haematopoiesis in irradiated mice [8], this raises the question as to what extent the follicle dermal cells were contributing to the haematopoietic pool. This was not explored here and would need future work. Notwithstanding this, adipose-derived MSCs have also been reported to support haematopoiesis better than bone marrow cells [74]. A feature of the adipose cells was their expression of CXCL12 (SDF-1) a key regulator of haematopoiesis, which is also strongly expressed by cultured hair follicle dermal cells (see Supplementary Figure S2). It would be surprising if hair follicle dermal cells were to be routinely adopted for support of pluripotent human cells given the direction of travel towards feeder-free methods [25–31] and the availability of alternative human candidate cell types [32–35]. However, follicle dermal cells do fulfill important criteria, including being easily accessible, readily propagated, and efficient at maintaining undifferentiated hESCs. Moreover, where there is a clinical role for support cells in transplantation, as in haematopoiesis [74, 75], follicle dermal cells have an advantage. These cells are in the process of being exploited for use in the production of new follicles in the treatment of alopecia and the creation of improved skin grafts. Therefore, work on the safe bioprocessing of these cells for clinical transplantation is already in train, making their eventual use in the context of stem cell support a more plausible proposition.
5. Conclusions
Here, we have demonstrated that coculture of hair follicle dermal cells with ESCs or iPSCs can support their long-term maintenance. We further show that the follicle cells support haematopoietic activity. This could have potential benefit in a clinical context, where the elimination of animal feeder layers is necessary prior to application of pluripotent cells for therapy and where the application of mesenchymal stem cell-like populations goes beyond their own direct therapeutic use, to include a role as support cells for other transplantable cell types.
Acknowledgments
The authors are grateful to Dr. N. Benvenisty from the Department of Genetics in Hebrew University, Jerusalem, Israel, for the gift of the Rex 1-EGFP vector and for the support from the Dermatology Foundation (to Claire A. Higgins) and NYSTEM (to Angela M. Christiano). The authors thank Sun Yung and Xin Zhang for their help and Adam Gilmore for reading the manuscript. This work was funded by a Medical Research Council Grant (G1000846) to Colin A. B. Jahoda, with additional support from the Biotechnology and Biological Sciences Research Council (BBS/B/14458) to Colin A. B. Jahoda and Nicholas Hole.
Abbreviations
- DP:
Dermal papilla
- DS:
Dermal sheath
- ESCs:
Embryonic stem cells
- mESCs:
Mouse ESCs
- iPSCs:
Induced pluripotent stem cells
- EBs:
Embryoid bodies
- CM:
Conditioned medium
- BMSC:
Bone marrow stromal cell
- MEF:
Mouse embryonic fibroblasts.
Conflicts of Interest
The authors declare that they have no conflicts of interest.
Authors' Contributions
Jun Liu, Claire A. Higgins, and Jenna C. Whitehouse contributed equally to the production of the paper.
Supplementary Materials
Supplementary material consists of a schematic illustrating the method and processes used for haematopoietic support assays and RT-PCR showing similarities in marker expression between hair follicle dermal cells and bone marrow cells. In particular, both express stromal cell-derived factor 1 (SDF-1) which has an important role in haematopoiesis. Supplementary Figure S1: schematic showing the strategy used to compare the support of hair follicle dermal cells and bone marrow stromal cells for blood cells. Supplementary Figure S2: RT-PCR demonstrates that important markers of the haematopoietic environment including, thrombospondin-1, V-CAM 1, and SDF-1 are expressed in bone marrow stromal cells (BM), S-17 an immortalized stromal cell culture line, and hair follicle dermal papilla (DP) and dermal sheath (DS) cells (two strains shown).
References
- 1.Morgan B. A. The dermal papilla: an instructive niche for epithelial stem and progenitor cells in development and regeneration of the hair follicle. Cold Spring Harbor Perspectives in Medicine. 2014;4(7, article a015180) doi: 10.1101/cshperspect.a015180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Higgins C. A., Chen J. C., Cerise J. E., Jahoda C. A. B., Christiano A. M. Microenvironmental reprogramming by three-dimensional culture enables dermal papilla cells to induce de novo human hair-follicle growth. Proceedings of the National Academy of Sciences of the United States of America. 2013;110(49):19679–19688. doi: 10.1073/pnas.1309970110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Pearton D. J., Yang Y., Dhouailly D. Transdifferentiation of corneal epithelium into epidermis occurs by means of a multistep process triggered by dermal developmental signals. Proceedings of the National Academy of Sciences of the United States of America. 2005;102(10):3714–3719. doi: 10.1073/pnas.0500344102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Fliniaux I., Viallet J. P., Dhouailly D., Jahoda C. A. B. Transformation of amnion epithelium into skin and hair follicles. Differentiation. 2004;72(9-10):558–565. doi: 10.1111/j.1432-0436.2004.07209009.x. [DOI] [PubMed] [Google Scholar]
- 5.Jahoda C. A. B., Whitehouse C. J., Reynolds A. J., Hole N. Hair follicle dermal cells differentiate into adipogenic and osteogenic lineages. Experimental Dermatology. 2003;12(6):849–859. doi: 10.1111/j.0906-6705.2003.00161.x. [DOI] [PubMed] [Google Scholar]
- 6.Rufaut N. W., Goldthorpe N. T., Wildermoth J. E., Wallace O. A. M. Myogenic differentiation of dermal papilla cells from bovine skin. Journal of Cellular Physiology. 2006;209(3):959–966. doi: 10.1002/jcp.20798. [DOI] [PubMed] [Google Scholar]
- 7.Richardson G. D., Arnott E. C., Jenna Whitehouse C., Lawrence C. M., Hole N., Jahoda C. A. B. Cultured cells from the adult human hair follicle dermis can be directed toward adipogenic and osteogenic differentiation. Journal of Investigative Dermatology. 2005;124(5):1090–1091. doi: 10.1111/j.0022-202X.2005.23734.x. [DOI] [PubMed] [Google Scholar]
- 8.Lako M., Armstrong L., Cairns P. M., Harris S., Hole N., Jahoda C. A. Hair follicle dermal cells repopulate the mouse haematopoietic system. Journal of Cell Science. 2002;115(20):3967–3974. doi: 10.1242/jcs.00060. [DOI] [PubMed] [Google Scholar]
- 9.Shi C., Mai Y., Cheng T. Identification of hematopoietic cell populations from the dermal papillae of human hair follicles. Transplantation Proceedings. 2004;36(10):3208–3211. doi: 10.1016/j.transproceed.2004.11.104. [DOI] [PubMed] [Google Scholar]
- 10.Fernandes K. J. L., McKenzie I. A., Mill P., et al. A dermal niche for multipotent adult skin-derived precursor cells. Nature Cell Biology. 2004;6(11):1082–1093. doi: 10.1038/ncb1181. [DOI] [PubMed] [Google Scholar]
- 11.Herzog E. L., Chai L., Krause D. S. Plasticity of marrow-derived stem cells. Blood. 2003;102(10):3483–3493. doi: 10.1182/blood-2003-05-1664. [DOI] [PubMed] [Google Scholar]
- 12.Itoh K., Tezuka H., Sakoda H., et al. Reproducible establishment of hemopoietic supportive stromal cell lines from murine bone marrow. Experimental Hematology. 1989;17(2):145–153. [PubMed] [Google Scholar]
- 13.Cheng L., Hammond H., Ye Z., Zhan X., Dravid G. Human adult marrow cells support prolonged expansion of human embryonic stem cells in culture. Stem Cells. 2003;21(2):131–142. doi: 10.1634/stemcells.21-2-131. [DOI] [PubMed] [Google Scholar]
- 14.Verfaillie C. M. Can human hematopoietic stem cells be cultured ex vivo? Stem Cells. 1994;12(5):466–476. doi: 10.1002/stem.5530120503. [DOI] [PubMed] [Google Scholar]
- 15.Kataoka K., Medina R. J., Kageyama T., et al. Participation of adult mouse bone marrow cells in reconstitution of skin. The American Journal of Pathology. 2003;163(4):1227–1231. doi: 10.1016/S0002-9440(10)63482-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Fathke C., Wilson L., Hutter J., et al. Contribution of bone marrow-derived cells to skin: collagen deposition and wound repair. Stem Cells. 2004;22(5):812–822. doi: 10.1634/stemcells.22-5-812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Hoogduijn M. J., Gorjup E., Genever P. G. Comparative characterization of hair follicle dermal stem cells and bone marrow mesenchymal stem cells. Stem Cells and Development. 2006;15(1):49–60. doi: 10.1089/scd.2006.15.49. [DOI] [PubMed] [Google Scholar]
- 18.Hill R. P., Gardner A., Crawford H. C., et al. Human hair follicle dermal sheath and papilla cells support keratinocyte growth in monolayer coculture. Experimental Dermatology. 2013;22(3):236–238. doi: 10.1111/exd.12107. [DOI] [PubMed] [Google Scholar]
- 19.Thomson J. A., Itskovitz-Eldor J., Shapiro S. S., et al. Embryonic stem cell lines derived from human blastocysts. Science. 1998;282(5391):1145–1147. doi: 10.1126/science.282.5391.1145. [DOI] [PubMed] [Google Scholar]
- 20.Martin G. R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proceedings of the National Academy of Sciences of the United States of America. 1981;78(12):7634–7638. doi: 10.1073/pnas.78.12.7634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Evans M. J., Kaufman M. H. Establishment in culture of pluripotential cells from mouse embryos. Nature. 1981;292(5819):154–156. doi: 10.1038/292154a0. [DOI] [PubMed] [Google Scholar]
- 22.Takahashi K., Okita K., Nakagawa M., Yamanaka S. Induction of pluripotent stem cells from fibroblast cultures. Nature Protocols. 2007;2(12):3081–3089. doi: 10.1038/nprot.2007.418. [DOI] [PubMed] [Google Scholar]
- 23.Takahashi K., Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126(4):663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 24.Williams R. L., Hilton D. J., Pease S., et al. Myeloid leukaemia inhibitory factor maintains the developmental potential of embryonic stem cells. Nature. 1988;336(6200):684–687. doi: 10.1038/336684a0. [DOI] [PubMed] [Google Scholar]
- 25.Chase L. G., Firpo M. T. Development of serum-free culture systems for human embryonic stem cells. Current Opinion in Chemical Biology. 2007;11(4):367–372. doi: 10.1016/j.cbpa.2007.06.421. [DOI] [PubMed] [Google Scholar]
- 26.Tsutsui H., Valamehr B., Hindoyan A., et al. An optimized small molecule inhibitor cocktail supports long-term maintenance of human embryonic stem cells. Nature Communications. 2011;2(1):p. 167. doi: 10.1038/ncomms1165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Fan Y., Wu J., Ashok P., Hsiung M., Tzanakakis E. S. Production of human pluripotent stem cell therapeutics under defined xeno-free conditions: progress and challenges. Stem Cell Reviews. 2015;11(1):96–109. doi: 10.1007/s12015-014-9544-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nagaoka M., Si-Tayeb K., Akaike T., Duncan S. A. Culture of human pluripotent stem cells using completely defined conditions on a recombinant E-cadherin substratum. BMC Developmental Biology. 2010;10(1):p. 60. doi: 10.1186/1471-213X-10-60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Takenaka C., Miyajima H., Yoda Y., et al. Controlled growth and the maintenance of human pluripotent stem cells by cultivation with defined medium on extracellular matrix- coated micropatterned dishes. PLoS One. 2015;10(6, article e0129855) doi: 10.1371/journal.pone.0129855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Amit M., Chebath J., Margulets V., et al. Suspension culture of undifferentiated human embryonic and induced pluripotent stem cells. Stem Cell Reviews. 2010;6(2):248–259. doi: 10.1007/s12015-010-9149-y. [DOI] [PubMed] [Google Scholar]
- 31.Baharvand H., Totonchi M., Taei A., Seifinejad A., Aghdami N., Salekdeh G. H. Human-induced pluripotent stem cells: derivation, propagation, and freezing in serum- and feeder layer-free culture conditions. Methods in Molecular Biology. 2010;584:425–443. doi: 10.1007/978-1-60761-369-5_23. [DOI] [PubMed] [Google Scholar]
- 32.Richards M., Fong C. Y., Chan W. K., Wong P. C., Bongso A. Human feeders support prolonged undifferentiated growth of human inner cell masses and embryonic stem cells. Nature Biotechnology. 2002;20(9):933–936. doi: 10.1038/nbt726. [DOI] [PubMed] [Google Scholar]
- 33.Mamidi M. K., Pal R., Mori N. A. B., et al. Co-culture of mesenchymal-like stromal cells derived from human foreskin permits long term propagation and differentiation of human embryonic stem cells. Journal of Cellular Biochemistry. 2011;112(5):1353–1363. doi: 10.1002/jcb.23052. [DOI] [PubMed] [Google Scholar]
- 34.Lee J. B., Song J. M., Lee J. E., et al. Available human feeder cells for the maintenance of human embryonic stem cells. Reproduction. 2004;128(6):727–735. doi: 10.1530/rep.1.00415. [DOI] [PubMed] [Google Scholar]
- 35.Zhang K., Cai Z., Li Y., et al. Utilization of human amniotic mesenchymal cells as feeder layers to sustain propagation of human embryonic stem cells in the undifferentiated state. Cellular Reprogramming. 2011;13(4):281–288. doi: 10.1089/cell.2010.0103. [DOI] [PubMed] [Google Scholar]
- 36.Xi J., Wang Y., Zhang P., et al. Human fetal liver stromal cells that overexpress bFGF support growth and maintenance of human embryonic stem cells. PLoS One. 2010;5(12, article e14457) doi: 10.1371/journal.pone.0014457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gledhill K., Gardner A., Jahoda C. A. B. Isolation and establishment of hair follicle dermal papilla cell cultures. Methods in Molecular Biology. 2013;989:285–292. doi: 10.1007/978-1-62703-330-5_22. [DOI] [PubMed] [Google Scholar]
- 38.Reynolds A. J., Oliver R. F., Jahoda C. A. Dermal cell populations show variable competence in epidermal cell support: stimulatory effects of hair papilla cells. Journal of Cell Science. 1991;98(1):75–83. doi: 10.1242/jcs.98.1.75. [DOI] [PubMed] [Google Scholar]
- 39.Eiges R., Schuldiner M., Drukker M., Yanuka O., Itskovitz-Eldor J., Benvenisty N. Establishment of human embryonic stem cell-transfected clones carrying a marker for undifferentiated cells. Current Biology. 2001;11(7):514–518. doi: 10.1016/S0960-9822(01)00144-0. [DOI] [PubMed] [Google Scholar]
- 40.Bain G., Kitchens D., Yao M., Huettner J. E., Gottlieb D. I. Embryonic stem cells express neuronal properties in vitro. Developmental Biology. 1995;168(2):342–357. doi: 10.1006/dbio.1995.1085. [DOI] [PubMed] [Google Scholar]
- 41.Dani C., Smith A. G., Dessolin S., et al. Differentiation of embryonic stem cells into adipocytes in vitro. Journal of Cell Science. 1997;110(11):1279–1285. doi: 10.1242/jcs.110.11.1279. [DOI] [PubMed] [Google Scholar]
- 42.Dexter T. M., Moore M. A., Sheridan A. P. Maintenance of hemopoietic stem cells and production of differentiated progeny in allogeneic and semiallogeneic bone marrow chimeras in vitro. The Journal of Experimental Medicine. 1977;145(6):1612–1616. doi: 10.1084/jem.145.6.1612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Collins L. S., Dorshkind K. A stromal cell line from myeloid long-term bone marrow cultures can support myelopoiesis and B lymphopoiesis. The Journal of Immunology. 1987;138(4):1082–1087. [PubMed] [Google Scholar]
- 44.Pragnell I. B., Wright E. G., Lorimore S. A., et al. The effect of stem cell proliferation regulators demonstrated with an in vitro assay. Blood. 1988;72(1):196–201. [PubMed] [Google Scholar]
- 45.Ying Q. L., Nichols J., Chambers I., Smith A. BMP induction of Id proteins suppresses differentiation and sustains embryonic stem cell self-renewal in collaboration with STAT3. Cell. 2003;115(3):281–292. doi: 10.1016/S0092-8674(03)00847-X. [DOI] [PubMed] [Google Scholar]
- 46.Hardy M. H. The secret life of the hair follicle. Trends in Genetics. 1992;8(2):55–61. doi: 10.1016/0168-9525(92)90350-D. [DOI] [PubMed] [Google Scholar]
- 47.Reynolds A. J., Jahoda C. A. Hair matrix germinative epidermal cells confer follicle-inducing capabilities on dermal sheath and high passage papilla cells. Development. 1996;122(10):3085–3094. doi: 10.1242/dev.122.10.3085. [DOI] [PubMed] [Google Scholar]
- 48.Coraux C., Hilmi C., Rouleau M., et al. Reconstituted skin from murine embryonic stem cells. Current Biology. 2003;13(10):849–853. doi: 10.1016/S0960-9822(03)00296-3. [DOI] [PubMed] [Google Scholar]
- 49.Mitsui K., Tokuzawa Y., Itoh H., et al. The homeoprotein Nanog is required for maintenance of pluripotency in mouse epiblast and ES cells. Cell. 2003;113(5):631–642. doi: 10.1016/S0092-8674(03)00393-3. [DOI] [PubMed] [Google Scholar]
- 50.Pan G. J., Chang Z. Y., Scholer H. R., Pei D. Stem cell pluripotency and transcription factor Oct4. Cell Research. 2002;12(5-6):321–329. doi: 10.1038/sj.cr.7290134. [DOI] [PubMed] [Google Scholar]
- 51.Darr H., Mayshar Y., Benvenisty N. Overexpression of NANOG in human ES cells enables feeder-free growth while inducing primitive ectoderm features. Development. 2006;133(6):1193–1201. doi: 10.1242/dev.02286. [DOI] [PubMed] [Google Scholar]
- 52.Draper J. S., Pigott C., Thomson J. A., Andrews P. W. Surface antigens of human embryonic stem cells: changes upon differentiation in culture. Journal of Anatomy. 2002;200(3):249–258. doi: 10.1046/j.1469-7580.2002.00030.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chin A. C. P., Fong W. J., Goh L.-T., Philp R., Oh S. K. W., Choo A. B. H. Identification of proteins from feeder conditioned medium that support human embryonic stem cells. Journal of Biotechnology. 2007;130(3):320–328. doi: 10.1016/j.jbiotec.2007.04.013. [DOI] [PubMed] [Google Scholar]
- 54.Smith A. G., Heath J. K., Donaldson D. D., et al. Inhibition of pluripotential embryonic stem cell differentiation by purified polypeptides. Nature. 1988;336(6200):688–690. doi: 10.1038/336688a0. [DOI] [PubMed] [Google Scholar]
- 55.Conover J. C., Ip N. Y., Poueymirou W. T., et al. Ciliary neurotrophic factor maintains the pluripotentiality of embryonic stem cells. Development. 1993;119(3):559–565. doi: 10.1242/dev.119.3.559. [DOI] [PubMed] [Google Scholar]
- 56.Rose T. M., Weiford D. M., Gunderson N. L., Bruce A. G. Oncostatin M (OSM) inhibits the differentiation of pluripotent embryonic stem cells in vitro. Cytokine. 1994;6(1):48–54. doi: 10.1016/1043-4666(94)90007-8. [DOI] [PubMed] [Google Scholar]
- 57.Pennica D., Shaw K. J., Swanson T. A., et al. Cardiotrophin-1. Biological activities and binding to the leukemia inhibitory factor receptor/gp130 signaling complex. Journal of Biological Chemistry. 1995;270(18):10915–10922. doi: 10.1074/jbc.270.18.10915. [DOI] [PubMed] [Google Scholar]
- 58.Matsuda T., Nakamura T., Nakao K., et al. STAT3 activation is sufficient to maintain an undifferentiated state of mouse embryonic stem cells. The EMBO Journal. 1999;18(15):4261–4269. doi: 10.1093/emboj/18.15.4261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Malik N., Kallestad J. C., Gunderson N. L., et al. Molecular cloning, sequence analysis, and functional expression of a novel growth regulator, oncostatin M. Molecular and Cellular Biology. 1989;9(7):2847–2853. doi: 10.1128/MCB.9.7.2847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Aubert J., Dessolin S., Belmonte N., et al. Leukemia inhibitory factor and its receptor promote adipocyte differentiation via the mitogen-activated protein kinase cascade. Journal of Biological Chemistry. 1999;274(35):24965–24972. doi: 10.1074/jbc.274.35.24965. [DOI] [PubMed] [Google Scholar]
- 61.Gearing D. P., Gough N. M., King J. A., et al. Molecular cloning and expression of cDNA encoding a murine myeloid leukaemia inhibitory factor (LIF) The EMBO Journal. 1987;6(13):3995–4002. doi: 10.1002/j.1460-2075.1987.tb02742.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Yu M., Kissling S., Freyschmidt-Paul P., Hoffmann R., Shapiro J., McElwee K. J. Interleukin-6 cytokine family member oncostatin M is a hair-follicle-expressed factor with hair growth inhibitory properties. Experimental Dermatology. 2008;17(1):12–19. doi: 10.1111/j.1600-0625.2007.00643.x. [DOI] [PubMed] [Google Scholar]
- 63.Kwack M. H., Ahn J. S., Kim M. K., Kim J. C., Sung Y. K. Dihydrotestosterone-inducible IL-6 inhibits elongation of human hair shafts by suppressing matrix cell proliferation and promotes regression of hair follicles in mice. Journal of Investigative Dermatology. 2012;132(1):43–49. doi: 10.1038/jid.2011.274. [DOI] [PubMed] [Google Scholar]
- 64.Sano S., Kira M., Takagi S., Yoshikawa K., Takeda J., Itami S. Two distinct signaling pathways in hair cycle induction: Stat3-dependent and -independent pathways. Proceedings of the National Academy of Sciences of the United States of America. 2000;97(25):13824–13829. doi: 10.1073/pnas.240303097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Legrand J. M. D., Roy E., Ellis J. J., Francois M., Brooks A. J., Khosrotehrani K. STAT5 activation in the dermal papilla is important for hair follicle growth phase induction. Journal of Investigative Dermatology. 2016;136(9):1781–1791. doi: 10.1016/j.jid.2016.04.014. [DOI] [PubMed] [Google Scholar]
- 66.Harel S., Higgins C. A., Cerise J. E., et al. Pharmacologic inhibition of JAK-STAT signaling promotes hair growth. Science Advances. 2015;1(9, article e1500973) doi: 10.1126/sciadv.1500973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Johnson L. F., deSerres S., Herzog S. R., Peterson H. D., Meyer A. A. Antigenic cross-reactivity between media supplements for cultured keratinocyte grafts. Journal of Burn Care & Rehabilitation. 1991;12(4):306–312. doi: 10.1097/00004630-199107000-00005. [DOI] [PubMed] [Google Scholar]
- 68.Meyer A. A., Manktelow A., Johnson M., deSerres S., Herzog S., Peterson H. D. Antibody response to xenogeneic proteins in burned patients receiving cultured keratinocyte grafts. The Journal of Trauma: Injury, Infection, and Critical Care. 1988;28(7):1054–1059. doi: 10.1097/00005373-198807000-00023. [DOI] [PubMed] [Google Scholar]
- 69.Ellerstrom C., Strehl R., Moya K., et al. Derivation of a xeno-free human embryonic stem cell line. Stem Cells. 2006;24(10):2170–2176. doi: 10.1634/stemcells.2006-0130. [DOI] [PubMed] [Google Scholar]
- 70.Tsai S. Y., Bouwman B. A., Ang Y. S., et al. Single transcription factor reprogramming of hair follicle dermal papilla cells to induced pluripotent stem cells. Stem Cells. 2011;29(6):964–971. doi: 10.1002/stem.649. [DOI] [PubMed] [Google Scholar]
- 71.Higgins C. A., Itoh M., Inoue K., Richardson G. D., Jahoda C. A. B., Christiano A. M. Reprogramming of human hair follicle dermal papilla cells into induced pluripotent stem cells. Journal of Investigative Dermatology. 2012;132(6):1725–1727. doi: 10.1038/jid.2012.12. [DOI] [PubMed] [Google Scholar]
- 72.de Oliveira V. C. C., dos Santos D. S., Vairo L., et al. Hair follicle-derived mesenchymal cells support undifferentiated growth of embryonic stem cells. Experimental and Therapeutic Medicine. 2017;13(5):1779–1788. doi: 10.3892/etm.2017.4195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Sugii S., Kida Y., Kawamura T., et al. Human and mouse adipose-derived cells support feeder-independent induction of pluripotent stem cells. Proceedings of the National Academy of Sciences of the United States of America. 2010;107(8):3558–3563. doi: 10.1073/pnas.0910172106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Nakao N., Nakayama T., Yahata T., et al. Adipose tissue-derived mesenchymal stem cells facilitate hematopoiesis in vitro and in vivo . The American Journal of Pathology. 2010;177(2):547–554. doi: 10.2353/ajpath.2010.091042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Zhao K., Liu Q. The clinical application of mesenchymal stromal cells in hematopoietic stem cell transplantation. Journal of Hematology & Oncology. 2016;9(1):p. 46. doi: 10.1186/s13045-016-0276-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary material consists of a schematic illustrating the method and processes used for haematopoietic support assays and RT-PCR showing similarities in marker expression between hair follicle dermal cells and bone marrow cells. In particular, both express stromal cell-derived factor 1 (SDF-1) which has an important role in haematopoiesis. Supplementary Figure S1: schematic showing the strategy used to compare the support of hair follicle dermal cells and bone marrow stromal cells for blood cells. Supplementary Figure S2: RT-PCR demonstrates that important markers of the haematopoietic environment including, thrombospondin-1, V-CAM 1, and SDF-1 are expressed in bone marrow stromal cells (BM), S-17 an immortalized stromal cell culture line, and hair follicle dermal papilla (DP) and dermal sheath (DS) cells (two strains shown).