

Abstract
Immunofluorescence is an effective method that helps to identify different cell types on tissue sections. In order to study the desired cell population, antibodies for specific cell markers are applied on tissue sections. In adult skeletal muscle, satellite cells (SCs) are stem cells that contribute to muscle repair and regeneration. Therefore, it is important to visualize and trace the satellite cell population under different physiological conditions. In resting skeletal muscle, SCs reside between the basal lamina and myofiber plasma membrane. A commonly used marker for identifying SCs on the myofibers or in cell culture is the paired box protein Pax7. In this article, an optimized Pax7 immunofluorescence protocol on skeletal muscle sections is presented that minimizes non-specific staining and background. Another antibody that recognizes a protein (laminin) of the basal lamina was also added to help identify SCs. Similar protocols can also be used to perform double or triple labeling with Pax7 and antibodies for additional proteins of interest.
Keywords: Developmental Biology, Issue 134, Skeletal muscle, satellite cells, Pax7, immunofluorescence, stem cell, basal lamina
Introduction
Skeletal muscle is composed of multinucleated muscle cells, called myotubes, organized in myofibers, which generate force and movements through contraction. Most skeletal muscles, except for some craniofacial muscles, are derived from a temporary embryonic structure called a somite1. Myogenic precursor cells delaminate from the epithelial somite to become myoblasts. Myoblasts further differentiate into myocytes that fuse to become myotubes to form multi-nucleated myofibers. The above process is called myogenesis and is characterized by temporally-regulated control of gene expression. Myogenic precursors express Pax3 and Pax7, whereas myoblasts express MyoD and/or Myf5 and myocytes express myogenin and myosins2,3. Muscle growth is a process in which myofibers become larger by incorporating more myonuclei into existing fibers (hyperplasia) and by an increase in muscle fiber size (hypertrophy)4. During muscle growth, there is a sustainable source of myogenic cells that have stem cell properties in that they can differentiate and self-renew. These cells are termed satellite cells based on their physical location between the sarcolemma (cell membrane of the myofiber) and the basal lamina5. SCs vigorously contribute to muscle growth in the juvenile stage (the first 2-3 weeks of postnatal mice), but become quiescent in resting adult muscle6. Remarkably, they can be re-activated in response to muscle injures and differentiate into new muscle cells to repair the damaged muscle7.
The stem cell properties make the study of SCs relevant for both basic muscle biology and therapies of muscle diseases8. As a result, it has been an area of intense investigation in the past decades. A tremendous progress has been made in dissecting the genetics and epigenetics of SCs9,10. Techniques involved in isolating and identifying SCs in situ were developed and optimized along the way11. Immunofluorescent staining allows the identification of SCs through the use of specific antibodies, including that for Pax7. However, the scarcity and small size of the SCs combined with a strong auto-fluorescence of adult skeletal muscle tissue render the visualization challenging. Here, we describe an immunofluorescent staining protocol optimized for mouse muscle tissue for Pax7 and based on an existing method for zebrafish muscle12. In addition, a Laminin antibody labeled with a distinct fluorophore is employed to identify the basal lamina under which the SCs are located. This protocol consistently allows the visualization of Pax7-positive SCs and myogenic precursors under all tested physiological conditions and developmental stages.
Protocol
In this protocol, the anterior hind limb muscles of adult mice (2-6 months), tibialis anterior (TA) and extensor digitorum longus (EDL), were employed as an example to perform the immunofluorescence staining on their SCs. All the steps handling mice and muscle tissue dissections have been approved by the Animal Care and Use Committee (ACUC) of NIAMS/NIH.
1. Dissect the TA/EDL Muscle from the Mouse Hind Limb
Euthanize the mouse in a CO2 filled euthanasia chamber under the guidelines of the ACUC of the NIH. Perform cervical dislocation if needed.
Put the mouse face-up on a dissection pad (supine). Spray 70% ethanol onto its belly skin. Cut a 1-2 cm opening horizontally on the center of the mouse belly skin, then deskin the mouse by pulling the opening toward the mouse feet. Expose one side of the mouse hind limb.
Use two sharp forceps: use one to hold the tendons that attach the TA/EDL to the ankle, and use the other one to break and shear the fascia covering the TA/EDL. Once the fascia is peeled off, use the sharp tip of the forceps to detach the TA/EDL from the tibia bone by running the tip underneath the TA/EDL muscle.
Cut off the ankle tendons with scissors while still holding them with the forceps. Trim the muscle with scissors along the longitudinal line of the TA/EDL from the ankle to the knee while lifting the TA/EDL from the ankle side with the forceps. Cut the knee-side tendons to detach the whole TA/EDL from the tibia.
2. Cryo-embedding of the TA/EDL Muscle
Prepare a liquid nitrogen bath in a container, e.g., a liquid nitrogen Dewar flask. Pour methylbutane into a small plastic beaker (about 20 mL) to half-full. Then put the beaker in the liquid nitrogen bath to freeze it. After a few minutes, check that the methylbutane is frozen. NOTE: The liquid nitrogen can be replaced by a bucket of dry ice, but the freezing time is slower.
Lay the dissected TA/EDL onto the flat surface of a small plastic syringe plunger. Fully cover the TA/EDL with a few drops of Optimal Cutting Temperature (O.C.T.) compound (the freezing medium).
Take the methylbutane beaker out of the liquid nitrogen bath and thaw the surface with a warm object (e.g., the handle of scissors). Quickly dip the O.C.T. covered TA/EDL on the plunger into the melted methylbutane to snap-freeze the tissue.
Detach the embedded tissue from the plunger with forceps, and put it into a 1.5 mL microcentrifuge tube. Place the tube into liquid nitrogen to store it while handling more samples. NOTE: The embedded tissue can be stored at -80 °C for long term (2 years) or -20 °C for short term (6 months).
3. Cryostat Sectioning
Add a drop of O.C.T. compound onto the center of a specimen holder of a cryostat. Wait until the O.C.T. compound is almost solidified. Quickly stick the embedded TA/EDL to the O.C.T. compound, perpendicularly with the ankle-side down and the knee-side up.
Mount the specimen holder onto a cryostat, cut the tissue at 10 µm interval, and collect the cryo-sections on frost coated slides with about 4-8 sections on one slide.
Dry the sections at room temperature for at least 3 h to overnight. NOTE: Although drying the sections in the freezer for overnight or longer is acceptable, it will inevitably increase the autofluorescence. For best results, dry the sections for 0.5 h, then proceed with the fixation and staining steps immediately.
Collect the slides in a slide box and use them immediately, or store the slides at -80 °C or -20 °C for later use.
4. Preparation of Reagents for Immunofluorescence Staining
- Prepare buffers:
- Prepare 2 L of 1x phosphate buffered saline (PBS) from 10x PBS.
- Prepare 40 mL of 4% paraformaldehyde (PFA) in 1x PBS from 16% PFA.
- Prepare 2 L of PBST: 1x PBS with 0.1% Triton-100.
- Prepare 50 mL of PBST-B: PBST with 2% bovine serum albumin (BSA).
- Prepare 10 mL of PBST-B-G: PBST with 2% BSA and 5% Normal Goat Serum.
- Prepare 1 mL of Blocking solution. Dilute AffiniPure Fab Fragment Goat Anti-Mouse IgG (H+L) (1:10) in PBST-B.
- Prepare 200 mL of 1x citrate buffer. NOTE: The volumes of solutions above are sufficient for at least 5 slides. Volumes should be adjusted according to the number of slides and the size of the slide containers.
- Prepare antibody stains:
- Primary Antibodies (working concentration): Prepare 1.2 mL of primary antibody mix by diluting Pax7 monoclonal mouse antibody (IgG1) (1-5 µg/mL) + Laminin polyclonal rabbit antibody (0.5-2 µg/mL) + MF20 monoclonal mouse antibody (IgG2b) (0.5-2.5 µg/mL) in PBST-B-G. NOTE: MF20 monoclonal mouse antibody (IgG2b) is optional.
- Secondary Antibodies (working concentration): Prepare 1.2 mL of secondary antibody mix by diluting Goat anti-Mouse IgG1 cross-adsorbed secondary antibody, Alexa Fluor 488 (0.5-2 µg/mL) + Goat anti-Rabbit IgG (H+L) highly cross-adsorbed secondary antibody, Alexa Fluor Plus 555 (0.5-2 µg/mL) + Goat anti-Mouse IgG2b cross-adsorbed secondary antibody, Alexa Fluor 647 (0.5-2 µg/mL) in PBST-B-G. NOTE: Alexa Fluor 647 is optional.
5. Immunofluorescence Staining Steps
- Rehydrate and fix the cryo-sections with PFA.
- Remove a few of the TA/EDL cryo-section slides from the slide box. Warm the slides in a slide-holding tray at room temperature.
- Use a liquid blocker pen (PAP pen) to draw an area that includes all the cryo-sections on the slide. This circle will prevent the solutions from flowing off the slide.
- Take the tray to a laboratory fume hood. Add 300-500 µL of 4% PFA onto the slide within the circled area to fix the sections at room temperature for 10 min. Wash out the PFA with 1x PBS at least 3x 5 min in a slide container. CAUTION: PFA is toxic and a hazardous waste; it should be operated with care and disposed into a hazardous waste container.
- Antigen retrieval
- Fill another slide container with 200 mL of citrate buffer.
- Transfer the slides into the container. Transfer the container with the slides to a pressure cooker to cook the slides in high pressure mode for 10 min.
- Cool down the container with draining water for 10 min. Transfer all slides to a new container with PBST (200 mL), rinse the slides with PBST for at least 3x 5 min.
- Blocking
- Add water-soaked kimwipes to the tray holding the slides to create a moist environment to avoid drying of the slides.
- Move the slides back to the tray, and add 200 µL of blocking solution onto each slide. Cover the tray with a cap and let the blocking develop for 30 min. NOTE: Pax7 antibody is a mouse monoclonal antibody, so there is unspecific binding of mouse IgG in mouse muscle tissues that creates high background. Using the AffiniPure Fab Fragment Goat Anti-Mouse IgG (H+L) blocks the mouse-on-mouse unspecific binding.
- Apply primary antibodies.
- Rinse the slides with PBST once in the slide container. Then move the slides back to the staining tray.
- Apply 200 µL of primary antibody mix on each slide, cover the tray with a cap, and let the reaction develop at room temperature for 1 h or at 4 °C for overnight. NOTE: The overnight reaction at 4 °C gives the best result, although developing the reaction at room temperature gives satisfactory results. The myosin MF20 antibody can be added in the mix to perform a triple (Pax7, Laminin, MF20) labeling. MF20 stains differentiated muscle fibers. This step also serves as another blocking step with PBST-B-G to reduce potential unspecific binding from secondary antibodies (goat antibodies) in the following step.
- Apply secondary antibodies.
- Wash out the primary antibodies with PBST at room temperature at least 3x 5 min.
- Add 200 µL of secondary antibody mix to each slide, and allow the reaction to develop at room temperature for at least 1 h. NOTE: For Pax7+Laminin, use Goat Anti-Mouse IgG1 Alexa 488 and Goat Anti-Rabbit Alexa 555. For Pax7+Laminin+MF20, add Goat Anti-Mouse IgG2b Alexa 647 in the mix.
- Wash out the secondary antibodies with PBST at least 3x 5min.
- DAPI (4',6-diamidino-2-phenylindole) counter-stain and mounting
- Dilute DAPI (1:20,000) in PBST in the slide container, e.g., 10 µL of DAPI in 200 mL of PBST.
- Stain the slides in the DAPI container for 3 min.
- Wash out the DAPI with PBST for at least 5x 5 min.
- Mount the slides with mounting medium. Cover the slides with coverslips. CAUTION: DAPI is toxic and hazardous; it should be handled with care and disposed in the hazardous waste bottle.
6. Fluorescent Microscopy
NOTE: The immunofluorescent staining protocol reported above will allow SCs visualization with either a wide-field fluorescent or confocal microscope. It may be initially challenging to identify SCs. Here are a few recommended steps that may be of help.
Use the DAPI channel and low magnification objectives to visualize the sections on the slides.
Switch the magnification from low to high. To observe SCs, at least a 20X objective is required).
Use the dual filter cube for 488 (FITC)/555 (Rhodamine) to observe both Pax7 and Laminin signals at the same time to identify SCs. Under this setting, the signal of Pax7 staining has a better contrast to the background noise.
Quickly alternate the channel from FITC to DAPI to correctly identify SCs. All SCs should be Pax7/DAPI-positive.
Representative Results
Following the above steps, SCs can be successfully visualized in adult resting muscle sections under a fluorescent microscope (Figure 1). Although the adult muscle tissue has strong auto-fluorescence in certain type of fibers, the bright Alexa series dyes can overcome the background noise and the signal stands out (Figure 1A, B; arrow heads). The two-photon confocal microscopy captures a relatively cleaner image (Figure 1B) than the wide-field fluorescent microscope (Figure 1A). The same protocol also worked well on muscle tissues of juvenile mice (Figure 2A), mouse embryos (Figure 2B), and injured adult muscle tissues (Figure 3). In juvenile and embryonic muscle tissues, there are more activated SCs (Figure 2A, arrows) or Pax7-positive myogenic precursors (Figure 2B, arrows) that have relatively larger nuclei; therefore, it is relatively easier to visualize SCs of younger mice than of adult mice (Figure 2 versus Figure 1). In the injured muscle tissue, there also are more activated Pax7-positive SCs (Figure 3).
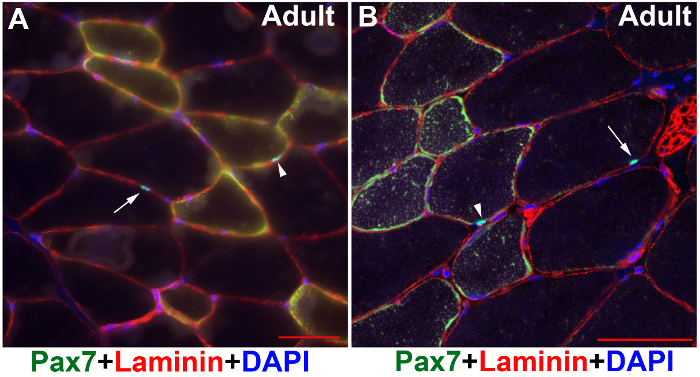
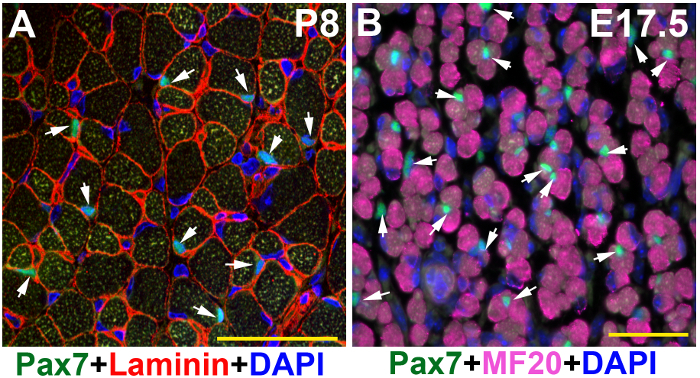
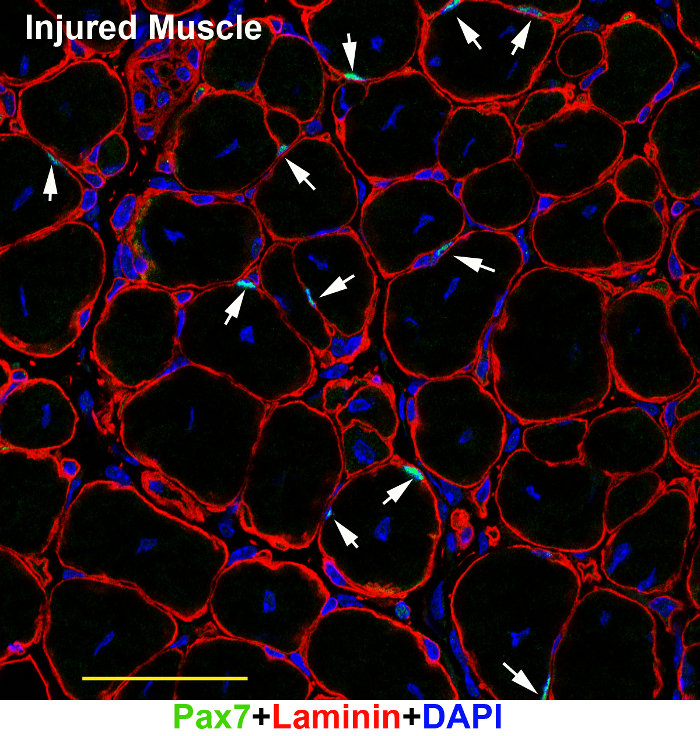
Figure 1: Images of Pax7 and Laminin immunofluorescence on mouse adult resting muscle tissues. TA/EDL muscle sections of adult mice were immunostained with Pax7 (green) and Laminin (red) antibodies, and counter-stained with DAPI (blue). (A) Image captured under a wide-field fluorescent microscope. (B) Image captured under a confocal microscope. Scale bars: 50 µm. Arrows: SCs from fibers with low autofluorescence. Arrowheads: SCs from fibers with high autofluorescence. Please click here to view a larger version of this figure.
Figure 2: Images of Pax7 and Laminin or MF20 immunofluorescence on muscle tissues of younger mice. TA/EDL muscle sections of younger mice were immunostained with Pax7 (green) and Laminin (red) or MF20 (magenta) antibodies, and counter-stained with DAPI (blue). (A) Postnatal day 8 (P8) muscle section captured under a confocal microscope. (B) Embryonic day 17.5 (E17.5) muscle section captured under a wide-field fluorescent microscope. Scale bars = 50 µm. Arrows: SCs in P8 muscle section (A); representative Pax7+ myogenic precursors in E17.5 muscle section, while there are more in the image (B). These two images were modified from raw data that also generated images in a published paper13 (sFigure 3D) and (sFigure 1B), respectively. Please click here to view a larger version of this figure.
Figure 3: A representative image of Pax7 and Laminin immunofluorescence on injured adult muscle tissues. TA/EDL muscle sections of an injured adult muscle tissues (7 days after injury) were immunostained with Pax7 (green) and Laminin (red) antibodies, and counter-stained with DAPI (blue). The image was captured under a confocal microscope. Scale bar = 50 µm. Arrows: SCs in the section. Please click here to view a larger version of this figure.
Discussion
The above protocol was based on a method of Pax7/MF20 staining on zebrafish skeletal muscle12. The solutions used and blocking steps are identical or similar. The antibodies used are identical. The adjusted steps were based on the features of mouse muscle tissue and SCs. First, Laminin antibody was added in the mix to help visualize and confirm the position of SCs. It was particularly helpful to count the number of SCs under the microscope when using the dual filter cube of 488/555; this greatly enhanced the SC-derived Pax7 signal to stand out from the noisy autofluorescent background. Second, since the Pax7 antibody used is a mouse antibody, we added a mouse-on-mouse blocking step (step 5.3) to reduce the background resulting from the unspecific binding of the mouse IgG. Third, the antigen retrieval step (step 5.2) was critical for staining with Pax7 antibody on PFA fixed tissues.
The same protocol was used on muscle tissues of younger mice (pups and embryos) (Figure 2). In fact, it was relatively easier to visualize SCs in pups than in adult mice by the same protocol. It might be helpful to start with staining SCs on pups to gain experience before proceeding with adult muscle tissue. A negative control without primary antibodies (step 5.4) is also necessary in every experiment. A similar protocol was successfully used on paraffin sections with extra paraffin processing steps. However, the paraffin sections tend to have a higher autofluorescence background, which masked signals from the SCs that have relatively weaker Pax7 expression. If possible, avoid using paraffin sections. The same protocol was also employed on injured muscle tissues that have numerous SCs, and gave similar results to those obtained with the pup's muscle tissue (Figure 3).
Muscle tissue autofluorescence represents the major obstacle in obtaining clean immunofluorescence results14. Reducing the drying time of cryo-sections can decrease autofluorescence and using freshly prepared cryo-sections is highly recommended. Similar to other protocols15, the mouse-on-mouse blocking step is required to further reduce the background.
While both wide-field fluorescent microscope and confocal microscope were effective for SCs visualization through eye pieces, the use of confocal microscope is recommended to capture high-quality images.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We thank the NIAMS Light Imaging Section for providing the microscopes and technical help. The MF20 and Pax7 antibodies were obtained from the Developmental Studies Hybridoma Bank developed under the auspices of the NICHD and maintained by the Department of Biological Sciences, The University of Iowa, Iowa City. This work was supported by the Intramural Research Program of NIAMS of the National Institutes of Health.
References
- Buckingham M. Gene regulatory networks and cell lineages that underlie the formation of skeletal muscle. Proc Natl Acad Sci U S A. 2017;114(23):5830–5837. doi: 10.1073/pnas.1610605114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckingham M, Relaix F. PAX3 and PAX7 as upstream regulators of myogenesis. Semin Cell Dev Biol. 2015;44:115–125. doi: 10.1016/j.semcdb.2015.09.017. [DOI] [PubMed] [Google Scholar]
- Relaix F, Buckingham M. From insect eye to vertebrate muscle: redeployment of a regulatory network. Genes Dev. 1999;13(24):3171–3178. doi: 10.1101/gad.13.24.3171. [DOI] [PubMed] [Google Scholar]
- Chang NC, Chevalier FP, Rudnicki MA. Satellite Cells in Muscular Dystrophy - Lost in Polarity. Trends Mol Med. 2016;22(6):479–496. doi: 10.1016/j.molmed.2016.04.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mauro A. Satellite cell of skeletal muscle fibers. J Biophys Biochem Cytol. 1961;9:493–495. doi: 10.1083/jcb.9.2.493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuang S, Rudnicki MA. The emerging biology of satellite cells and their therapeutic potential. Trends Mol Med. 2008;14(2):82–91. doi: 10.1016/j.molmed.2007.12.004. [DOI] [PubMed] [Google Scholar]
- Wang YX, Rudnicki MA. Satellite cells, the engines of muscle repair. Nat Rev Mol Cell Biol. 2011;13(2):127–133. doi: 10.1038/nrm3265. [DOI] [PubMed] [Google Scholar]
- Rinaldi F, Perlingeiro RC. Stem cells for skeletal muscle regeneration: therapeutic potential and roadblocks. Transl Res. 2014;163(4):409–417. doi: 10.1016/j.trsl.2013.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Grand F, Rudnicki MA. Skeletal muscle satellite cells and adult myogenesis. Curr Opin Cell Biol. 2007;19(6):628–633. doi: 10.1016/j.ceb.2007.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giordani L, Puri PL. Epigenetic control of skeletal muscle regeneration: Integrating genetic determinants and environmental changes. FEBS J. 2013;280(17):4014–4025. doi: 10.1111/febs.12383. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu L, Cheung TH, Charville GW, Rando TA. Isolation of skeletal muscle stem cells by fluorescence-activated cell sorting. Nat Protoc. 2015;10(10):1612–1624. doi: 10.1038/nprot.2015.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Feng X, Adiarte EG, Devoto SH. Hedgehog acts directly on the zebrafish dermomyotome to promote myogenic differentiation. Dev Biol. 2006;300(2):736–746. doi: 10.1016/j.ydbio.2006.08.056. [DOI] [PubMed] [Google Scholar]
- Juan AH, et al. Polycomb EZH2 controls self-renewal and safeguards the transcriptional identity of skeletal muscle stem cells. Genes Dev. 2011;25(8):789–794. doi: 10.1101/gad.2027911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson KA, Snyder DS, Goodell MA. Skeletal muscle fiber-specific green autofluorescence: potential for stem cell engraftment artifacts. Stem Cells. 2004;22(2):180–187. doi: 10.1634/stemcells.22-2-180. [DOI] [PubMed] [Google Scholar]
- Lepper C, Conway SJ, Fan CM. Adult satellite cells and embryonic muscle progenitors have distinct genetic requirements. Nature. 2009;460(7255):627–631. doi: 10.1038/nature08209. [DOI] [PMC free article] [PubMed] [Google Scholar]