

Abstract
Heteropteran insects such as assassin bugs (Reduviidae) and giant water bugs (Belostomatidae) descended from a common predaceous and venomous ancestor, and the majority of extant heteropterans retain this trophic strategy. Some heteropterans have transitioned to feeding on vertebrate blood (such as the kissing bugs, Triatominae; and bed bugs, Cimicidae) while others have reverted to feeding on plants (most Pentatomomorpha). However, with the exception of saliva used by kissing bugs to facilitate blood-feeding, little is known about heteropteran venoms compared to the venoms of spiders, scorpions and snakes.
One obstacle to the characterization of heteropteran venom toxins is the structure and function of the venom/labial glands, which are both morphologically complex and perform multiple biological roles (defense, prey capture, and extra-oral digestion). In this article, we describe three methods we have successfully used to collect heteropteran venoms. First, we present electrostimulation as a convenient way to collect venom that is often lethal when injected into prey animals, and which obviates contamination by glandular tissue. Second, we show that gentle harassment of animals is sufficient to produce venom extrusion from the proboscis and/or venom spitting in some groups of heteropterans. Third, we describe methods to harvest venom toxins by dissection of anaesthetized animals to obtain the venom glands. This method is complementary to other methods, as it may allow harvesting of toxins from taxa in which electrostimulation and harassment are ineffective. These protocols will enable researchers to harvest toxins from heteropteran insects for structure-function characterization and possible applications in medicine and agriculture.
Keywords: Environmental Sciences, Issue 134, Heteroptera, true bug, Reduviidae, Belostomatidae, venom, toxin, saliva, electrostimulation, harassment, venom gland, labial gland, salivary gland
Introduction
Heteropteran venoms are potently bioactive substances1. For example, the venom/saliva secretions of blood-feeding Heteroptera such as kissing bugs (Triatominae) and bed bugs (Cimicidae) facilitates feeding by disrupting hemostasis2. Toxins in these venoms target multiple pathways including coagulation, platelet aggregation and vasoconstriction, as well as the pain and itch pathways. Venoms from most other heteropteran species are adapted to facilitate predation rather than blood-feeding. Their venoms cause paralysis, death and tissue liquefaction when injected into invertebrates3,4. When injected into vertebrates, their venom may also have drastic effects. For example, injection of venom from the assassin bug Holotrichius innesi into vertebrates causes pain, muscle paralysis and hemorrhage; mice envenomated by this bug die quickly due to respiratory paralysis5.
Transcriptomic and proteomic studies have revealed the protein composition of some heteropteran venoms. Venoms of predaceous species are rich in proteases, other enzymes, and peptides and proteins of unknown structure and function6,7,8. Kissing bug venom is rich in the triabin protein family, whose members profoundly affect coagulation, platelet aggregation, and vasoconstriction2,9. However, it is not known which toxins underlie most bioactivities of venom. For example, venom of the kissing bug Triatoma infestans has been reported to be analgesic and inhibit sodium channels10, but the components responsible remain to be elucidated. Likewise, it is not known what component(s) of assassin bug venom cause paralysis or pain. A prerequisite for identifying the toxins responsible for particular venom bioactivities, and for characterizing the structure and function of novel venom toxins, is obtaining venom.
Venom has been obtained from heteropterans by electrostimulation5,6,7,8,11,12,13, provocation of defensive responses4,8, mechanically squeezing the thorax12,14,15,16, dissecting out venom glands8,17,18,19,20,21,22, and application of agonists of the muscarinic acetylcholine receptor23. Judging the potential advantages and disadvantages of any method is complicated by the morphology of heteropteran venom glands, which consist of a main gland with two separate lumens, the anterior main gland (AMG) and posterior main gland (PMG), as well as an associated accessory gland (AG). These different gland compartments produce different protein secretions, which may be specialized for different biological functions including prey capture, defense and extra-oral digestion8,17. In peiratine and ectrichodiine assassin bugs, the AMG has been associated with prey capture and the PMG with extra-oral digestion17. However, in the harpactorine bug Pristhesancus plagipennis the PMG is specialized for prey capture and digestion whereas the AMG is hypothesized to secrete defensive venom8. The AG has been described as having little secretory function in assassin bugs8 or as a major site of protease storage in giant water bugs23. Clearly, further work is required to clarify the function of each gland compartment among various heteropteran subgroups, and to determine the function of most venom toxins. In this report we describe protocols for harvesting venom toxins from heteropterans toward this goal.
Protocol
This protocol complies with The University of Queensland's policy set out in Responsible Care and Use of Animals in Teaching and Research (PPL 4.20.11) as well as the National Health and Medical Research Council's Australian code for the care and use of animals for scientific purposes (8th Edition 2013).
Caution: Take care not to be envenomated when handling assassin bugs. Take care to protect the eyes when handling species that spit venom defensively. Take care throughout not to injure the experimental animals. This includes monitoring of pressure on restraints such as rubber bands and ensuring that the proboscis is not broken.
NOTE: Optionally, anaesthetize animals by exposure to CO2 for 0.5-2 min or cooling to 4-10 °C prior to venom harvesting in Aims 1-3 to facilitate safe transfer and restraint. Anaesthetization is not strictly required but may facilitate safe restraint of agile or strong specimens. However, animals must be awake to allow venom harvesting. Keep downstream applications in mind when deciding whether or not to add protease inhibitors.
1. Harvesting Venom Toxins by Electrostimulation
Obtain live specimens from which to harvest toxins.
- Use pre-prepared plastic tweezers with positive and negative electrodes mounted on either tip. Connect electrified tweezers to an electrostimulator or a source of constant voltage that allows adjustment of the voltage.
- For small (~10 mm) and large (~25 mm) assassin bugs, use peak voltages of 15 and 25 V respectively.
- For larger heteropterans such as giant water bugs, use up to 40 V.
Restrain live bugs by strapping them to a platform, using a rubber band over the thorax.
- Place the tip of the proboscis in a suitable collecting tip. For assassin bugs, use a P200 pipette tip. For giant water bugs, cut the extremity off a P200 tip to increase the size of the aperture.
- Gently lift the proboscis with a closed pair of clean tweezers and push the open aperture of the collection tip over the end of the proboscis.
- If desired, uptake ~5 µL ultrapure water prior to placing the proboscis into the collecting tip. This reduces losses of venom remaining inside of the tip, though the harvested venom will be diluted.
Apply electrostimulation. Dip the stimulating electrodes in conductive gel, such as 2.5 M NaCl/50% glycerol. Apply the electrodes to the thorax. For belostomatids, apply the two electrodes to the dorsal posterior surface of the head.
Store venom to prevent autodegradation. After venom is extruded, quickly transfer it to a tube at -20 °C or -60 °C, or a tube containing protease inhibitor cocktail.
Repeat steps 1.5 and 1.6 until sufficient venom is acquired or no further venom is forthcoming.
2. Harvesting of Venom Toxins by Harassment
Prepare animals for venom harvesting and place the tip of the proboscis in a collection vestibule, as described in subsections 1.1 and 1.3-1.4.
If venom is extruded spontaneously, go to step 2.3. If not, harass the animals by gently touching it on the legs, abdomen and antennae with tweezers until venom is produced.
Quickly transfer venom to a tube at -20 °C or -60 °C, or a tube containing protease inhibitor cocktail, if desired.
3. Harvesting of Venom Toxins by Harassment from Venom "Spitting" Species
Anaesthetize, or partly anaesthetize, the insect before removing it from its enclosure to prevent any premature defensive spitting.
Provoke venom spitting behavior. Contain and reposition the insect using the deep lid of a standard 90 x 16 mm Petri dish. Hold the lid slightly posterior and 1-4 cm above the insect to prevent flight. Most insects will spit several times, often in quick succession. Ensure all venom is collected on the underside of the dish.
Collect the venom on the underside of the Petri dish by rinsing with 10 µL of ultrapure water. Quickly transfer it to a tube at -20 °C or -60 °C, or a tube containing protease inhibitor cocktail.
4. Harvesting Venom Toxins by Gland Dissection
Sacrifice animals. Heavily anaesthetize or kill animals using >5 min exposure to CO2. Pipe pure CO2 directly into the air-holes of the animal's housing enclosure.
Pin insect to dissection tray. For assassin bugs, dissect through the ventral surface (4.3). For giant water bugs, dissect through the dorsal surface (4.4).
- Ventral dissection
- Insert three pins into the posterior abdomen to hold the insect down without puncturing the venom glands.
- Cut a short midline incision in the ventral surface of the abdomen using a miniature scalpel. Use miniature scissors to extend the midline incision anteriorly to the head, taking care to cut the exoskeleton only and not damage internal structures.
- To expose the internal structures, make multiple lateral cuts extending from the midline incision to the side of the insect. Then, pin back each flap of ventral exoskeleton to reveal internal structures.
- For large assassin bugs, make four lateral incisions, in the mid-abdomen, anterior abdomen, between first and second legs, and in front of the first leg.
- Dorsal dissection
- Remove the wings near the base. Insert three pins into the posterior abdomen to hold the insect down without puncturing the venom glands.
- Cut a midline incision from the head to the abdomen using miniature scissors and scalpel, taking care to cut the exoskeleton only and not damage internal structures.
- Force apart the two halves of the insect. Place several pins laterally along the length of the insect to leave the internal cavity exposed.
- Remove the flight muscles using tweezers.
Flood the dissection tray. Add PBS until the bug is submerged to allow internal structures float up and be more easily visualized.
- Using tweezers and micro-scissors, carefully remove connective and nervous tissue and trachea. The venom glands appear as elongated, translucent structures extending along each side of the alimentary canal.
- Identify the main gland by its characteristic appearance, with anterior and posterior lobes and two ducts meeting at the hilus.
- If desired, identify the accessory gland by tracing the duct from the hilus. Free the main gland by cutting the two ducts emanating from the hilus.
- Harvest the desired gland lumens. Transfer the gland to a microcentrifuge on ice containing 30 µL of PBS or PBS plus protease inhibitor cocktail. Lance the glands with a clean sharp pin.
- Vortex for 10 s and centrifuge (1 min, 5,000 × g, 4 °C) to empty the gland lumens. Remove the glandular tissue using tweezers.
Clarify the toxin extract. Centrifuge (5 min, 17,000 × g, 4°C) to remove any solid particles, retaining the supernatant and discarding the pellet. Store at -20°C or -60 °C to prevent autoproteolytic degradation.
Representative Results
Some heteropteran species, such as the harpactorine P. plagipennis and the reduviine Platymeris rhadamanthus, reliably yield large quantities (5-20 µL) of venom in response to electrostimulation (Table 1). In general, most peiratine, reduviine, and harpactorine bugs yield venom in response to this method. Among stenopodaine bugs, electrostimulation elicited venom from Oncocephalus sp. but not Thodelmus sp. The holoptiline and emesine bugs sampled did not yield significant venom (e.g. enough for analysis by mass spectrometry) in response to electrostimulation. Electrostimulation can also be used to harvest venom from belostomatid bugs and predatory stinkbugs. However, electrostimulation of water scorpions (Nepidae) induced release of contents of the cephalic glands only, rather than venom from the proboscis. Failure to harvest venom by electrostimulation in some species is most likely due to the morphological complexity of the venom glands and the physiological mechanisms controlling the release of venom8.
In addition to releasing venom due to electrostimulation, the reduviids P. plagipennis, Havinthus rufovarius, P. rhadamanthus and the belostomatid Lethocerus distinctifemur, will spontaneously eject venom from the proboscis during handling. Such venom ejection is frequently accompanied by defensive displays. P. rhadamanthus also spits venom defensively4, a behavior that occurs in snakes24 and spiders25 but of which we are not aware of in any other reduviid species.
SDS-PAGE and proteomics experiments demonstrate that the venoms harvested by electrostimulation and harassment are protein-rich6,7,8. Proteins account for a large proportion of the material present, though it is also likely that venoms contain inorganic ions and other substances. Assassin bug venom obtained by electrostimulation and harassment typically contains over one hundred peptides and proteins (Figure 1, Figure 2). Belostomatid venom has previously been reported to be rich in lysophospholipids13. Infrared absorbance spectra of venom from the belostomatine water bug Diplonychus eques are consistent with a content of both proteins and lysophospholipids. For the lethocerine L. distinctifemur, evidence was found only for protein and not lysophospholipids6.

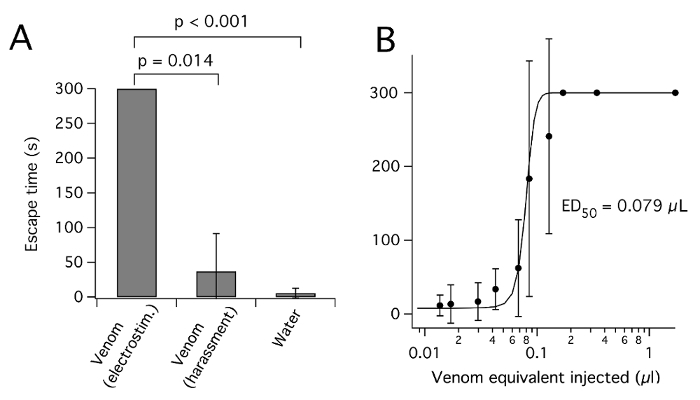
As reported for spider venoms26, venom harvested from heteropteran insects is likely to vary in concentration and composition, depending on the insect used and the method by which it is harvested. UV spectroscopy of diluted venom samples suggests absorbance values (A280) of 50-250 (10 mm path length) for undiluted venom, consistent with a high protein concentration of ~50-250 mg/mL7,12,19. Prey deprivation has been reported to cause successive increase in venom concentration and paralytic potential3 as well as successive decreases in pH27. However, prolonged starvation will result in loss of condition and death. As well as concentration, the method by which venom is harvested from heteropterans may affect its composition. The toxin composition of venom from the assassin bug P. plagipennis differed markedly depending on whether it was harvested by electrostimulation or harassment8. In the case of P. plagipennis, this was shown to be due to electrostimulation yielding the contents of the PMG, whereas harassment yielded the contents of the AMG. Venom obtained by electrostimulation, but not harassment, potently paralyzed prey insects (Figure 3). However, it is unclear to what extent this result can be generalized to other Reduviidae or other Heteroptera.
Harvesting venom directly by dissecting venom glands allows the control mechanisms of the venom glands to be circumvented, at the cost of contamination with glandular tissue (non-venom) proteins. Regardless, extracts obtained from dissected material can be used for bioactivity/toxicity assays. For example, extracts of the PMG, AMG and AG of P. plagipennis, prepared using the above protocol, were analyzed using liquid chromatography/tandem mass spectrometry8. This process identified a total of 182, 114, and 71 proteins in total, of which 45, 51, and 12 were classified as putative venom proteins based on amino acid sequence characteristics, with the remaining proteins classified as putative housekeeping proteins. Injection of extracts of the PMG, but not AMG or AG, into insects resulted in paralysis and death8.
| Infraorder | Family | Subfamily | Binomial name | Common name | Electrostimulation | Harassment | Dissection |
| Cimicomorpha | Reduviidae | Harpactorinae | Pristhesancus plagipennis | Common Brisbane assassin bug | √ | √ | √ |
| Havinthus rufovarius | Red tiger assassin bug | √ | √ | √ | |||
| Scipinia arenacea | Red spiny assassin bug | √ | nd | √ | |||
| Gminatus spp. | Large orange assassin bug | √ | nd | √ | |||
| Trachylestes aspericollis | Small red assassin bug | √ | nd | nd | |||
| Reduviinae | Platymeris spp. | Giant African assassin bug | √ | √ | √ | ||
| Psytalla horrida | Spiny assassin bug | √ | nd | √ | |||
| Peiratinae | Ectomocoris spp. | Orange ground assassin bug | √ | nd | √ | ||
| Peirates spp. | Black assassin bug | √ | nd | nd | |||
| Stenopodainae | Oncocephalus spp. | - | √ | nd | √ | ||
| Thodelmus spp. | - | x | nd | √ | |||
| Holoptilinae | Ptilocnemus lemur | Feather-legged bug | x | x | nd | ||
| Emesinae | Stenolemus spp. | Thread-legged bug | x | x | x | ||
| Pentatomomorpha | Pentatomidae | Asopinae | Amyotea hamata | Yellow predatory stink bug | √ | nd | nd |
| Nepomorpha | Nepidae | Ranatrinae | Ranatra dispar | Water scorpion | x, cg | x | √ |
| Belostomatidae | Belostomatinae | Diplonychus eques | Water bug | √ | nd | nd | |
| Belostomatidae | Lethocerinae | Lethocerus sp. | Giant water bug | √ | √ | √ | |
| tick, successful; cross, unsuccessful; nd, not determined; cg, cephalic gland discharge only |
Table 1: Taxon specificity of methods used to harvest venom from heteropterans.
Figure 1: Proteins detected by LC-MS/MS analysis of 2D SDS-PAGE spots and HPLC fractions of venom collected from P. plagipennis by electrostimulation (Protocol 1), showing abundant proteases, CUB-domain proteins and heteropteran Venom Family 1 proteins. (A) 2D SDS-PAGE gel of crude P. plagipennis venom, showing protein families identified by LC-MS/MS of gel spots. (B) HPLC chromatogram from fractionation of P. plagipennis venom, showing protein families identified by LC-MS/MS analysis of collected fractions. Reproduced with permission7. Please click here to view a larger version of this figure.
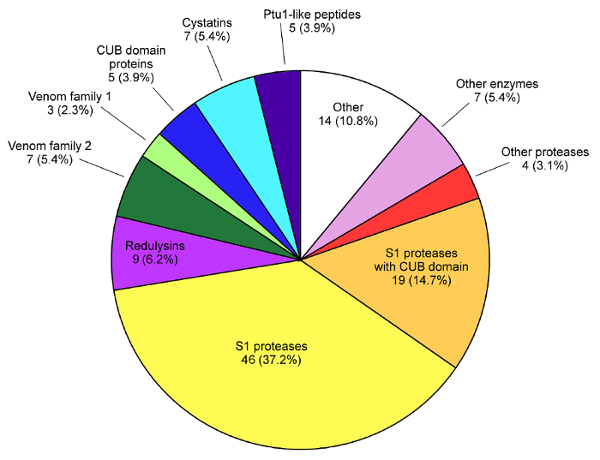
Figure 2: Proportion of sequences belonging to each major protein class in the venom ofP. plagipennis. Reproduced with permission7. Please click here to view a larger version of this figure.
Figure 3:P. plagipennis venom obtained by electrostimulation, but not harassment, paralyzes insects. (A) Effect of injecting venom obtained by electrostimulation or harassment, or water, on cricket escape. For each venom condition, 0.17 µL venom equivalent was injected into the abdomen and the time to escape an upturned petri-dish lid (in s, up to 300 s, mean ± SD) was scored. (B) Dose-response curve for inhibition of escape success by venom obtained from P. plagipennis by electrostimulation. Reproduced with permission8. Please click here to view a larger version of this figure.
Discussion
The most critical step in harvesting assassin bug venom is selecting the appropriate method depending on the purposes of the study. Each of the three methods presented for harvesting heteropteran venoms has advantages and disadvantages depending on downstream applications.
Inducing bugs to expel venom from the proboscis (Protocols 1-3) avoids contamination of venom by glandular tissues. In addition, these methods are non-lethal and can be repeated many times over the course of a bug's life. Electrostimulation usually provides the greatest quantities of venom, and yields venom with potent toxicity to prey insects according to several studies5,8. Provoking a defensive response is another way to elicit venom from the proboscis, and one that may yield venom of different protein content to electrostimulation8. However, electrostimulation and provocation do not work for many species, and without parallel investigations of the secretory output of the venom glands it is unclear which gland lumen (or which combination of gland lumens) is being harvested.
Harvesting venom by dissection (Protocol 4) is in many ways complementary. Dissection represents a direct way to access stored venom, and each compartment of the venom gland can be harvested separately or pooled (i.e., the possibility that the 'wrong' venom has been harvested is obviated). However, the method is lethal and additionally causes slight contamination of venom by tissue components. Many Heteroptera are too small (or too elongated in the case of Emesinae, the thread-legged bugs) to allow venom harvesting by dissection. If dissection is used to extract proteins from the individual gland compartments separately, it is critical to separate the lobes quickly and extract their contents separately to avoid cross-contamination.
The methods presented here will need to be modified depending on the particular species studied. For collection of venom by electrostimulation, the main aspects to optimize are how the bug is restrained. For example, most reduviids are able to extend their proboscis over a wide range of movement. These species can be simply restrained right-way-up on a platform using a rubber band, and the proboscis everted manually. For species with less flexible proboscis, such as belostomatids, it is instead necessary to restrain insects in an upside-down position and lower a collection receptacle at the correct angle using a retort or mechanical arm. The magnitude and pattern of electricity applied must also be optimized, and in this case, it is better to start low and slowly increase the applied voltage to avoid lethality.
If the aim of a study is to attain a detailed understanding of how a particular species produces and uses venom, an in-depth investigation combining multiple harvesting methods, as well as technologies such as mass spectrometry and RNA-Seq experiments, may be required. If the aim is to use heteropteran venoms as libraries of biological molecules to be screened for some desired biological activity, then a panel of venom samples obtained by electrostimulation, harassment, and/or dissection may be suitable. However, we note that the normal biological role of the venom harvested is likely to determine what bioactivities are present. For example, venom used for predation is more likely to contain insecticidal compounds, whereas venom used for defense is more likely to contain algogenic (pain-causing) agents.
We have not included venom harvest by application of the muscarinic acetylcholine receptor agonist pilocarpine in this protocol. Future experiments are required to determine the characteristics of pilocarpine-induced venom expulsion compared to the above methods.
In this article, we have presented methods that will allow researchers to obtain venoms from heteropteran insects. Successful venom collection will allow further investigations into the production, composition, function and evolution of venom in Heteroptera. In addition, some heteropteran toxins may find utility as eco-friendly insecticides, lead molecules to develop human therapeutics, or as scientific tools to investigate biological systems.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We acknowledge financial support from the Australian Research Council (Grants DP130103813 and LP140100832 to G.F.K., DECRA Fellowship DE160101142 to EABU), the Australian National Health & Medical Research Council (Principal Research Fellowship APP1044414 to G.F.K.), and The University of Queensland (Postdoctoral Fellowship to A.A.W.).
References
- Walker AA, Weirauch C, Fry BG, King GF. Venoms of heteropteran insects: A treasure trove of diverse pharmacological toolkits. Toxins. 2016;8(2):43. doi: 10.3390/toxins8020043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribeiro JMC, Assumpção TC, Francischetti IMB. An insight into the sialomes of bloodsucking Heteroptera. Psyche (Stuttg) 2012;2012:1–16. [Google Scholar]
- Ambrose DP, Maran SPM. Quantification protein content and paralytic potential of saliva of fed and prey deprived reduviid Acanthaspis pedestris Stål (Heteroptera: Reduviidae: Reduviinae) Indian Journal of Environmental Science. 1999;3(1):11–16. [Google Scholar]
- Edwards JS. The action and compostion of the saliva of an assassin bug Platymeris rhadamanthus Gaerst. (Hemiptera, Reduviidae) Journal of Experimental Biology. 1961;38:61–77. [Google Scholar]
- Zerachia T, Bergmann F, Shulov A. In: Animal and Plant Toxins. Kaiser E, editor. Goldman: 1973. pp. 143–146. [Google Scholar]
- Walker AA, Hernández-Vargas MJ, Corzo G, Fry BG, King GF. Giant fish-killing water bug reveals ancient and dynamic venom evolution in Heteroptera. Cellular and Molecular Life Sciences. 2018. [DOI] [PMC free article] [PubMed]
- Walker AA, et al. Giant fish-killing water bug reveals ancient and dynamic venom evolution in Heteroptera. Cell. Mol. Life Sci. 2018. [DOI] [PMC free article] [PubMed]
- Walker AA, et al. The assassin bug Pristhesancus plagipennis produces two distinct venoms in separate gland lumens. Nature Communications. 2018;9(1):755. doi: 10.1038/s41467-018-03091-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernández-Vargas MJ, Santibáñez-López CE, Corzo G. An insight into the triabin protein family of American hematophagous reduviids: Functional, structural and phylogenetic analysis. Toxins. 2016;8(2):44. doi: 10.3390/toxins8020044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dan A, Pereira MH, Pesquero JL, Diotaiuti L, Beirao PS. Action of the saliva of Triatoma infestans (Heteroptera: Reduviidae) on sodium channels. Journal of Medical Entomology. 1999;36(6):875–879. doi: 10.1093/jmedent/36.6.875. [DOI] [PubMed] [Google Scholar]
- Corzo G, Adachi-Akahane S, Nagao T, Kusui Y, Nakajima T. Novel peptides from assassin bugs (Hemiptera: Reduviidae): isolation, chemical and biological characterization. FEBS Letters. 2001;499(3):256–261. doi: 10.1016/s0014-5793(01)02558-3. [DOI] [PubMed] [Google Scholar]
- Sahayaraj K, Kumar SM, Anandh GP. Evaluation of milking and electric shocks for venom collection from hunter reduviids. Entomon. 2006;31(1):65–68. [Google Scholar]
- Silva-Cardoso L, et al. Paralytic activity of lysophosphatidylcholine from saliva of the waterbug Belostoma anurum. Journal of Experimental Biology. 2010;213(19):3305–3310. doi: 10.1242/jeb.041954. [DOI] [PubMed] [Google Scholar]
- Noeske-Jungblut C, et al. Triabin, a highly potent exosite inhibitor of Thrombin. Journal of Biological Chemistry. 1995;270(48):28629–28634. doi: 10.1074/jbc.270.48.28629. [DOI] [PubMed] [Google Scholar]
- Noeske-Jungblut C, et al. An inhibitor of collagen-induced platelet aggregation from the saliva of Triatoma pallidipennis. Journal of Biological Chemistry. 1994;269(7):5050–5053. [PubMed] [Google Scholar]
- Sahayaraj K, Borgio JF, Muthukumar S, Anandh GP. Antibacterial activity of Rhynocoris marginatus (Fab.) and Catamirus brevipennis (Servile) (Hemiptera: Reduviidae) venoms against human pathogens. Journal of Venomous Animals and Toxins Including Tropical Diseases. 2006;12(3):487–496. [Google Scholar]
- Haridass ET, Ananthakrishnan TN. Functional morphology of the salivary system in some reduviids (Insecta-Heteroptera-Reduviidae) Proceedings of the Indian Academy of Sciences. Animal Sciences. 1981;90(2):145–160. [Google Scholar]
- Maran SPM, Ambrose DP. Ignacimuth A, Sen A, Janarthanan S. Biotechnological Applications for Integrated Pest Management. Oxford Publishing; 2000. pp. 125–131. [Google Scholar]
- Maran SPM, Selvamuthu K, Rajan K, Kiruba DA, Ambrose DP. In: Insect Pest Management, A Current Scenario. Ambrose DP, editor. Entomology Research Unit; 2011. pp. 346–361. [Google Scholar]
- Pereira MH, et al. Anticoagulant activity of Triatoma infestans and Panstrongylus megistus saliva (Hemiptera/Triatominae) Acta Tropica. 1996;61:255–261. doi: 10.1016/0001-706x(96)00007-1. [DOI] [PubMed] [Google Scholar]
- Ribeiro JM, Marinotti O, Gonzales R. A salivary vasodilator in the blood-sucking bug, Rhodnius prolixus. British Journal of Pharmacology. 1990;101(4):932–936. doi: 10.1111/j.1476-5381.1990.tb14183.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ribeiro JM, Schneider M, Guimarães JA. Purification and characterization of prolixin-S (nitrophorin 2), the salivary anticoagulant of the blood-sucking bug Rhodnius prolixus. Biochem Journal. 1995;308(1):243–249. doi: 10.1042/bj3080243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swart CC, Deaton LE, Felgenhauer BE. The salivary gland and salivary enzymes of the giant waterbugs (Heteroptera; Belostomatidae) Comparative Biochemistry and Physiology A Molecular & Integrative Physiology. 2006;145(1):114–122. doi: 10.1016/j.cbpa.2006.05.010. [DOI] [PubMed] [Google Scholar]
- Rasmussen S, Young B, Krimm H. On the 'spitting' behaviour in cobras (Serpentes: Elapidae) Journal of Zoology. 1995;237(1):27–35. [Google Scholar]
- Fink LS. Venom spitting by the green lynx spider, Peucetia viridans (Araneae, Oxyopidae) Journal of Arachnology. 1984;12:372–373. [Google Scholar]
- Herzig V. Ontogenesis, gender, molting influence the venom yield in the spider Coremiocnemis tropix (Araneae, Theraphosidae) Journal of Venomous Research. 2010;1:76–83. [PMC free article] [PubMed] [Google Scholar]
- Sahayaraj K, Subramanium M, Rivers D. Biochemical and electrophoretic analyses of saliva from the predatory reduviid species Rhynocoris marginatus (Fab) Acta Biochimica Polonica. 2013;60(1):91–97. [PubMed] [Google Scholar]