

Abstract
Alveolar macrophages are terminally differentiated, lung-resident macrophages of prenatal origin. Alveolar macrophages are unique in their long life and their important role in lung development and function, as well as their lung-localized responses to infection and inflammation. To date, no unified method for identification, isolation, and handling of alveolar macrophages from humans and mice exists. Such a method is needed for studies on these important innate immune cells in various experimental settings. The method described here, which can be easily adopted by any laboratory, is a simplified approach to harvesting alveolar macrophages from bronchoalveolar lavage fluid or from lung tissue and maintaining them in vitro. Because alveolar macrophages primarily occur as adherent cells in the alveoli, the focus of this method is on dislodging them prior to harvest and identification. The lung is a highly vascularized organ, and various cell types of myeloid and lymphoid origin inhabit, interact, and are influenced by the lung microenvironment. By using the set of surface markers described here, researchers can easily and unambiguously distinguish alveolar macrophages from other leukocytes, and purify them for downstream applications. The culture method developed herein supports both human and mouse alveolar macrophages for in vitro growth, and is compatible with cellular and molecular studies.
Keywords: Immunology and Infection, Issue 134, Lung, Tissue resident macrophage, Bronchoalveolar lavage, Embryonic macrophage, Macrophage culture, Lung homeostasis
Introduction
The lung microenvironment is a uniquely complex ecosystem with an elaborate air conduit and vasculature. The inhaled air travels through the trachea and through numerous branches of bronchi and bronchioles before reaching the alveoli, where the blood-air gas exchange occurs. Due to direct interaction with the atmosphere, the respiratory surface requires protection from the potentially harmful effects of airborne particles and pollutants. A number of physical, chemical, and immunologic barriers protect the lungs. Notably, deployment of phagocytes at the respiratory surface serves an important first-line defense system. Alveolar macrophages (AMs) are one type of lung-resident phagocytes, and they make up the vast majority of the lung macrophage pool. As their name suggests, AMs are primarily localized to the alveolar lumen and occur as sessile cells that constantly sample the ambient atmosphere and communicate with the alveolar epithelium1. In steady state lungs, more than 95% of the phagocytes in the alveolar space are AMs2, whose composition may alter due to inflammation, infection, or chronic exposure to pollutants.
AMs participate in a wide range of functions that may be local to the lungs and/or of systemic importance. For example, AMs are essential in the development and the optimal functioning of lungs; immune surveillance; and clearance of cellular debris, invading pathogens, and inhaled particles3,4,5,6,7. Targeted depletion of AMs is known to impair clearance of respiratory viruses and bacteria4,8. Besides their role as phagocytes and a first-line defenders of pulmonary homeostasis, AMs are known to function as antigen-presenting cells in eliciting T cell immunity9, potentiating the efficacy of intranasal vaccine10 and influencing lung-restricted autoimmunity after lung transplantation11,12. Deficiency in AM function has been linked to pulmonary alveolar proteinosis (PAP), a condition resulting from a genetic mutation, malignancy or infection that impairs clearance of pulmonary surfactants13,14. Transplantation of AMs is now being explored as a therapeutic approach for treatment of PAP 15,16.
AMs are known to originate during embryogenesis and to persist in the lungs throughout life without being replaced by circulating leukocytes2,17. Although, AM turnover is undetectable in homeostatic lungs, varying levels of AM turnover have been reported in certain clinical conditions including infection by the influenza virus4, myeloablative irradiation18, exposure to endotoxin19, and old age20. AMs are believed to self-renew via a low-grade proliferation17,21, but some recent studies claim that monocytes can give rise to a population of intravascular lung macrophages22,23 under experimental conditions, but functionality of these newly converted pulmonary macrophages have yet to be defined in lung diseases. Furthermore, understanding the threshold of stimulus in the context of AM activation is a potentially interesting area, as the lung attempts to preserve an equilibrium between the inflammatory signals and the immunoregulatory machinery.
The physiologic or pathologic alterations that lead to loss of immune regulation are important to evaluate in various clinical settings (e.g., respiratory infections, inflammatory lung disease and fibrotic lung disease). Nevertheless, AMs are increasingly recognized as indicators or even determinants of pulmonary health11,24. Currently, there are no unified protocols available for harvesting, characterizing, and/or maintaining AMs from humans and preclinical murine models. Lack of a consensus on AM precursors and phenotypes, and absence of a detailed methodology had been the major roadblock in deciphering role(s) of AM in pulmonary health and disease. The following protocol offers a definitive identification, isolation, and in vitro culture strategy that will greatly advance the understanding of AM behavior and facilitate AM-targeted diagnostic and therapeutic studies.
Protocol
All methods described here have been approved by the Institutional Animal Care and Use Committee (IACUC) and the Institutional Review Board (IRB) at St Joseph's Hospital and Medical Center.
1. Isolation of AMs from Murine Bronchoalveolar Lavage (BAL) Fluid
Anaesthetize an eight-week-old C57BL/6 mouse with ketamine (87.5 mg/kg body weight) and xylazine (12.5 mg/kg body weight) cocktail via an intraperitoneal injection. Proceed when mouse attains surgical anesthesia with loss of reflexes and relaxation of muscles.
Place the mouse on a dissection surface with the ventral side facing up. Apply ophthalmic vet ointment on eyes to prevent dehydration and wipe the entire ventral surface of mouse with sterile alcohol prep pads saturated with 70% isopropanol to disinfect.
Mount the mouse with all four legs restrained.
Gently open the abdominal cavity with micro dissection tools. Care must be taken to open as much area as possible without damaging any visceral organs or creating overhanging tissue.
Gently open the thoracic cavity by slowly incising the rib cage through the mediastinum. Ideally, the rib cage can be excised from side to side. Extreme care must be taken not to injure the pleura of lungs while opening the thoracic cavity.
Locate the inferior vena cava and make an incision to bleed the mouse. Use absorbent cotton pads to soak up blood.
Inject 10 mL ice-cold phosphate buffered saline (PBS) into the right ventricle (using 10 mL syringe and 25G needle) to flush circulating blood cells. Absorb the flow of blood with absorbent cotton pads. Once completely perfused, the lungs will appear blanched and are then ready for lavage.
Gently cut open the skin and muscle in the neck to expose the airway. Carefully excise the adjoining muscles, cartilages, and fat tissues without damaging the trachea.
Make a small incision (<2 mm) on the trachea posterior to the larynx. This incision should be just enough to insert a catheter into while the trachea is still attached to the larynx. Insert a 1-inch 22G catheter without a needle into the trachea towards the lungs. Secure the catheter with a silk braided suture (4-0; non-absorbable) with a square knot. NOTE: The catheter needs to be trimmed based on the mouse size. Extremely short or long catheters tend to leak and/or tear the airway during lavage. Catheter length should be such that when fully inserted the tip remains well within the trachea and not reach the primary bronchi.
Attach a 1-mL syringe filled with ice-cold BAL buffer (Ca2+ and Mg2+ free PBS + 1 mM Ethylenediaminetetraacetic acid, EDTA) to the catheter and slowly instill the buffer into the lungs. This will inflate the lungs. Keep the syringe attached to the catheter for five seconds and then aspirate the lavage fluid by gently pulling the piston. This will deflate the lungs. Collect the fluid in a 15 mL conical tube placed on ice.
Repeat Step 1.10 for nine more times and pool the lavage fluid. Do not exceed 1 mL buffer per flush, as that may exceed the lung capacity and lead to its rupture.
Euthanize the mouse with CO2 overdose by IACUC approved protocol.
Centrifuge the 10 mL BAL fluid at 250 x g at 4 °C for 10 minutes. The cell pellet contains BAL cells. Caution: The pellet may be very small, so exercise caution when aspirating the supernatant.
Resuspend the BAL cells in 100 μL of flow/sorting buffer (PBS + 2% Heat inactivated fetal bovine serum (FBS) + 1 mM EDTA + 25 mM N-2-Hydroxyethylpiperazine-N'-2-ethanesulphonic acid (HEPES)).
Block the nonspecific antibody binding to Fc receptor by adding mouse Fc block (clone 2.4G2, 10 μg/mL) and incubating for 20 minutes on ice.
Perform antibody staining along with fluorescence minus one (FMO) and single-stain controls11,25. Evaluate the BAL cells by a cell analyzer (see Table of Materials) followed by single-cell flow cytometry analysis by appropriate analysis software (see Table of Materials).
Isolate AMs by a fluorescence-activated cell sorter (see Table of Materials) directly into 1 mL mouse AM culture medium when AMs are intended for in vitro culture or in vivo cell transfer. Alternatively, sort AMs into 1 mL Lysis buffer (see Table of Materials) supplemented with 10 μL/mL β-Mercaptoethanol for RNA extraction.
2. Isolation of AMs from Mouse Lung, Single-cell Suspension
Anaesthetize an eight-week-old C57BL/6 mouse with ketamine (87.5 mg/kg body weight) and xylazine (12.5 mg/kg body weight) cocktail via an intraperitoneal injection. Proceed when mouse attains surgical anesthesia with loss of reflexes and relaxation of muscles.
Place the mouse on a dissection surface with the ventral side facing up. Apply ophthalmic vet ointment on eyes to prevent dehydration and wipe the entire ventral surface of mouse with sterile alcohol pads saturated with 70% isopropanol to disinfect.
Mount the mouse with all four legs restrained.
Gently open the abdominal cavity with micro dissection tools. Care must be taken to open as much area as possible without damaging any visceral organs or creating overhanging tissue.
Gently open the thoracic cavity by slowly incising the rib cage through the mediastinum. Ideally, the rib cage can be excised from side to side. Extreme care must be taken not to injure the pleura of lungs while opening the thoracic cavity.
Locate the inferior vena cava and make an incision to bleed the mouse. Use absorbent cotton pads to soak up blood.
Inject 10 mL ice-cold PBS into the right ventricle (using 10 mL syringe and 25G needle) to flush circulating blood cells. Once completely perfused, the lungs will appear blanched and are ready for harvest.
Dissect out the lungs by severing the trachea, blood vessels and ligaments into a 15 mL conical tube with 5 mL of cold Dulbecco's Modified Eagle's medium (DMEM). Maintain it on ice until the next step.
Euthanize the mouse with CO2 overdose by IACUC approved protocol.
Transfer the perfused lungs into a sterile and pyrogen-free 60 mm culture dish with 3 mL DMEM. With help of dissecting forceps tease out the airways and other hard non-lung tissue material, if present. Mince the lung tissue with a scalpel to <1 mm size.
Add 300 µg/mL Liberase TL and 5 U/mL DNase I, mix by pipetting and incubate at 37° C in an incubator for 25 minutes. Gently mix once by pipetting after 10 minutes.
Pass the dissociated lungs through a 100 µm cell strainer installed on a 50 mL conical tube. Use the back of a plunger from 1 mL syringe to mash up cell clumps on the filter. Wash the filter with 20 mL wash buffer (PBS + 2% Heat inactivated FBS + 2 mM EDTA).
Centrifuge at 500 x g at 4 °C for 5 minutes. Discard the supernatant and resuspend the cell pellet in 20 mL wash buffer.
Repeat steps 2.12 and 2.13 twice, changing the filter and tube each time. Pre-soaking the filter in wash buffer for more than five minutes increases the AM yield. Prepare single-cell suspension by adding 100 μL flow/sorting buffer to the cell pellet.
Block the nonspecific antibody binding to the Fc receptor by adding mouse Fc block (clone 2.4G2, 10 μg/mL) and incubating for 20 minutes on ice.
Perform antibody staining along with FMO and single-stain controls11,25. Evaluate the lung cells by a cell analyzer (see Table of Materials) followed by single-cell flow cytometry analysis by analysis software.
Sort AMs by a cell sorter (see Table of Materials) directly into 1 mL mouse AM culture medium when AMs are intended for in vitro culture or in vivo cell transfer. Alternatively, sort AMs into 1 mL Lysis buffer supplemented with 10 μL/mL β-Mercaptoethanol for RNA extraction.
3. Isolation of AMs from Human BAL Fluid
Arrange transfer of the human BAL fluid from the clinic to the laboratory at 4 °C.
Centrifuge the BAL fluid (10 - 50 mL) at 250 x g at 4 °C for 10 minutes. The cell pellet contains BAL cells11.
Wash the pellet with 20 mL wash buffer (PBS + 2% Heat inactivated FBS + 2 mM EDTA) and centrifuge at 250 x g at 4 °C for 10 minutes.
Resuspend the BAL cells are in 100 μL of flow/sorting buffer (PBS + 2% Heat inactivated FBS + 1 mM EDTA + 25 mM HEPES).
Block the nonspecific antibody binding to the Fc receptor by adding human Fc block (clone Fc1.3070, 25 μg/mL) and incubating for 20 minutes on ice.
Perform antibody staining along with FMO and single-stain controls11,24. Evaluate the BAL cells by a cell analyzer (see Table of Materials) followed by single-cell flow cytometry analysis by analysis software.
Sort AMs by a cell sorter (see Table of Materials) directly into 1 mL human AM culture medium or 1 mL Lysis buffer supplemented with 10 μL/mL β-Mercaptoethanol.
4. In vitro Culture of AMs
Following the cell sorting, harvest the cells by centrifuging at 250 x g at 4 °C for 10 min.
Resuspend human AMs in 1 mL human AM culture medium [Roswell Park Memorial Institute medium (RPMI) 1640 supplemented with 10% FBS, 20% L-929 culture supernatant (as a source of macrophage colony stimulating factor (M-CSF), 100 U/mL final concentration), 1 mM sodium pyruvate, 10 mM N-2-hydroxyethylpiperazine-N'-2-ethanesulphonic acid (HEPES), and 1x penicillin/streptomycin (optional)]. Similarly, resuspend mouse AMs in 1 mL mouse AM culture medium [DMEM supplemented with 10% FBS, 20% L-929 culture supernatant (as a source of M-CSF, 100 U/mL final concentration), 1 mM Sodium Pyruvate, 10 mM HEPES, and 1x penicillin/streptomycin (optional)].
Enumerate the trypan blue-excluding viable cells by a cell counter and adjust the viable cell concentration at 1 x 106/mL. NOTE: Ideally, up to 5 x 105 AMs can be isolated from BAL fluid collected from an 8 - 10 week old C57BL/6 male mouse. The AM yield from the mouse lung digest is variable and may be slightly less than that of the mouse BAL fluid. AM yield from human BAL fluid depends on the volume of the BAL fluid, the total volume of saline used in the lavage, and patient's clinical condition.
Based on the yield and experimental necessity, seed the cells in chamber slides, culture dishes or culture flasks. Ideally, seed the AMs at 1 x 105/mL with 10 mL medium in a T25 tissue culture flask.
Incubate the cells in a 37 °C humidified incubator with 5% CO2 atmosphere. NOTE: AMs will grow as adherent cells and are extremely slow-growing (doubling time >7 days).
At 24 hours post-seeding, AMs should be adherent and will not be detached by a gentle change of culture medium. Remove the medium by aspiration from one end and replenish with fresh medium pre-warmed to 37 °C. NOTE: The cells can now be used to assess AM physiology or the effects of stimuli. AMs grown in chamber slides can be conveniently stained with appropriate antibodies and are ideal for microscopic visualization. For flow cytometric evaluation, AMs are harvested by incubation with non-enzymatic cell dissociating solution.
Aspirate the culture medium and add dissociating solution enough to cover the culture surface (approximately 2 mL per T25 flask) and incubate at 37 °C for 5 - 10 minutes. No scraping will be necessary, and it is not recommended to scrape the cells mechanically. Gently tap the sides, add 1 - 2 mL flow/sorting buffer and gently pipette up and down to detach the majority of adherent cells.
For RNA studies, lyse the cells directly on the culture dish by adding Lysis buffer.
Representative Results
The flow cytometric approach to identify mouse AMs is shown in Figure 1. This includes analysis of a minimum set of surface markers necessary in distinguishing AMs from other lung-resident or lung-infiltrating phagocytes. Differential analysis is required to positively identify AMs from interstitial macrophages, dendritic cells, neutrophils, monocytes, and monocyte-derived pulmonary macrophages that occur in lungs. The following scheme of surface markers can be employed to readily distinguish these cell types from each other where AMs are CD45+, CD11c+, Siglec-F+, CD64+, I-Ab+/-, F4/80-, CD11b-; Interstitial macrophages are CD45+, CD11b+, I-Ab+, CD64+, CD24-; CD11b+ Dendritic cells are CD45+, CD11b+, CD11c+, I-Ab+, CD24+, CD64-; CD103+ Dendritic cells are CD45+, CD11c+, CD24+, CD103+, I-Ab+, CD11b-; Plasmacytoid Dendritic cells are CD45+, CD317+, Siglec-H+, Ly-6C+, I-Ab-, Neutrophils are CD45+, Ly-6G+, CD11b+, CD11c-, I-Ab-, Ly-6C-, and Monocytes are CD45+, CD64+/-, Ly-6C+/-, I-Ab-. The AM populations highlighted in red (CD11chi and Siglec-Fhi) and teal (CD11chi, Siglec-Fint), respectively, represent conventional AMs (major population) and monocyte-derived pulmonary macrophages (minor population) that occur in the alveolar space.Flow sorting of CD45+, CD11chi, Siglec-Fhi cells yields purified mouse AMs. Figure 2 illustrates cell surface markers necessary for a precise identification and isolation of human AMs. The human AMs are positively identified for being CD45+, CD11b+, HLA-DR+, CD163+, CD169+, and CD206+ BAL cells, and can be flow sorted for downstream applications.
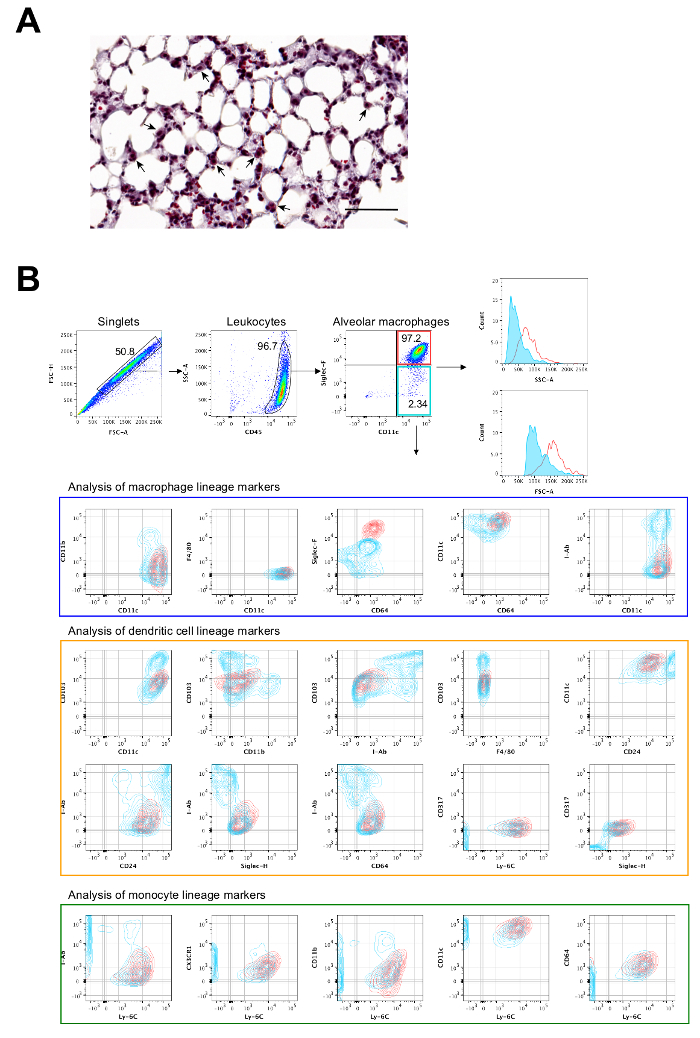
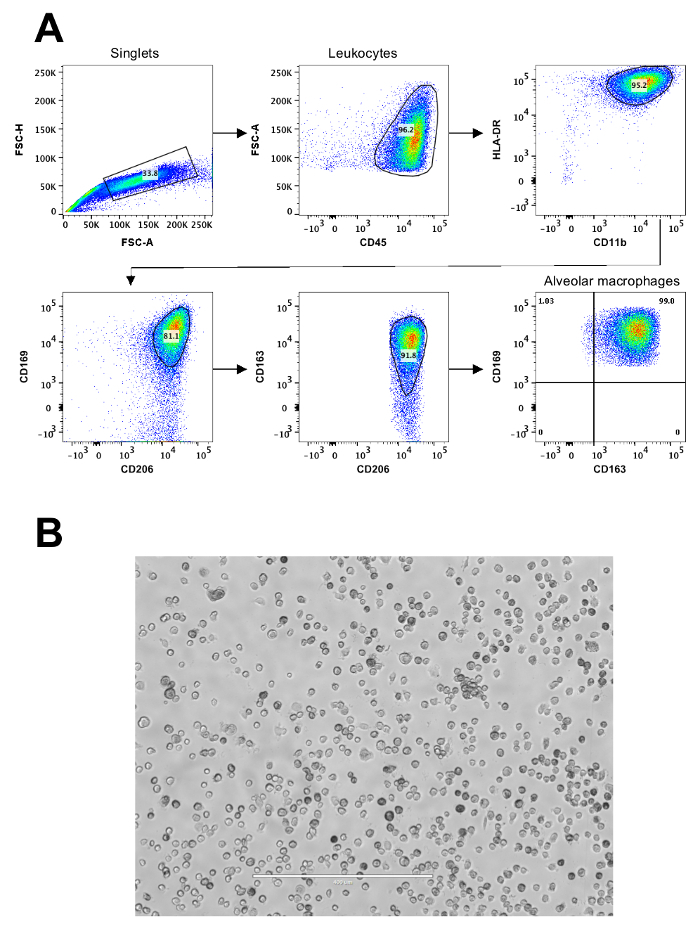
Figure 1: Representative flow cytometric analysis of mouse AMs. (A) Anatomic location of AMs in a mouse lung tissue section (hematoxylin and eosin). AMs are indicated with arrows and the scale bar is 60 μm. (B) Representative flow cytometry gating strategy to identify mouse AMs in BAL cells. Samples were evaluated by a cell analyzer and data were down sampled to 500 events for comparison by the analysis software. Red (CD11chi and Siglec-Fhi) and teal (CD11chi, Siglec-Fint), respectively, are conventional AMs (major population) and monocyte-derived pulmonary macrophages (minor population). Please click here to view a larger version of this figure.
Figure 2: Representative flow cytometric analysis of human AMs.(A) Gating strategy to identify human AMs from BAL fluid from a human lung transplant recipient. (B) Bright field micrograph of human AMs at seven days of culture. The scale bar is 400 μm. Please click here to view a larger version of this figure.
Discussion
AMs are long-living lung-resident macrophages that populate the lungs beginning at birth and enduring over the entire life span26. Their roles in pulmonary physiology7 and pathology12 and their potential to predict of pulmonary autoimmunity24 have been recognized. Because AMs have a long-term presence in lungs11,27 and because they are involved in activation and progression of immune responses11,24, their role(s) in other chronic inflammatory and fibrotic lung diseases needs to be evaluated. Historically, AMs had a confused identity and were often misidentified. With recently adopted phenotypic markers25,28, it is now feasible to distinguish human and mouse AMs from other pulmonary phagocytes (e.g., interstitial macrophages, pulmonary dendritic cells, and monocyte derived pulmonary macrophages). The methods described here have been optimized for isolation and for in vitro culture of mouse and human AMs. Although these protocols can be broadly adopted for AMs from other species, appropriate phenotypic determination should be conducted to establish a set of "minimally required" surface markers for thorough characterization.
For isolation of murine AMs by lavage, inclusion of chelating agent (i.e., EDTA) and pre-chilling the BAL buffer were found to be important in the current study. Because AMs are adherent to the alveolar wall, use of PBS alone was ineffective in dislodging the cells and produced a significantly lower yield. Securing installation of the catheter to the trachea is another critical step. Care should be taken to ensure a leak-proof attachment that minimizes loss of BAL fluid. Extra caution is needed while lavaging a younger mouse (<6 weeks old), as the structural delicacy of the trachea may result in tearing or leakage during lavage. Installing two consecutive non-overlapping sutures often ensures a secure attachment of the catheter and prevents leaks. Although non-enzymatic dissociation (i.e., with a chelator supplemented buffer) was only performed for AM isolation in this study, it is possible that enzymatic dissociation with collagenase during lung lavage may produce a better AM yield.
The yield of human AM from BAL fluid is variable. AMs constitute nearly 10% of all BAL cells collected from human lung transplant recipients at Norton Thoracic Institute during routine post-lung transplant follow-up, and the total AM yield is dependent on several variables. Such variables include patient's clinical condition, the volume of saline used in lavage, and the volume of BAL fluid processed for AM isolation. Pre-lavage procedures (e.g., needle biopsies) that result in microhemorrhage alter the cellular composition of BAL cells; nevertheless, implementation of phenotypic markers allows for an unambiguous detection and characterization of the human AM pool (Figure 2).
Use of L-929 conditioning medium (a source of M-CSF) was necessary for the in vitro maintenance of AMs. It is our belief that supplementation of purified M-CSF would be sufficient to sustain the AMs in culture; however, an optimized concentration needs to be worked out for both human and mouse AMs. The cultured cells can used for studies on endocytosis, response to inflammation, antigen presentation, maturation/activation status, or any other assay that necessitates maintenance of live cells for a time-course analysis11,24. In the course of this study, it was not attempted to maintain AMs as a continuously cultured line and only short-term terminal studies (<30 days since harvest) were performed. Therefore, an obvious limitation to this method is lack of a long-term culture data, particularly on stability of the cells (i.e., whether AMs can grow as a finite or continuous line, the response of AMs to cryoprotectants and cryopreservation, and phenotypic and/or genotypic drift) after ongoing culture. AMs are long living cells in vivo, and some degree of variation is expected between the cells isolated from healthy lungs and from those undergoing respiratory infection or inflammatory/autoimmune responses.
Overall, the method presented here documents a simplified approach to isolating, characterizing, and maintaining human and mouse AMs in a cell culture system. Although this protocol can be further optimized based on individual experimental needs, it may serve as a beginner's protocol to dissect the functional relevance of AMs in respiratory diseases and pulmonary medicine.
Disclosures
The authors have no conflicts of interest to declare.
Acknowledgments
We thank Clare Prendergast for assistance with editing the manuscript. DKN is supported by a research grant (#2095) from the Flinn Foundation and TM is supported by grants from National Institutes of Health (R01HL056643 and R01HL092514). DKN developed the methods, designed the study and wrote the manuscript; OM assisted with animal studies and clinical sample procurement; SB assisted with Flow cytometric analysis and cell sorting; TM supervised the studies and reviewed the manuscript.
References
- Westphalen K, et al. Sessile alveolar macrophages communicate with alveolar epithelium to modulate immunity. Nature. 2014;506(7489):503–506. doi: 10.1038/nature12902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guilliams M, et al. Alveolar macrophages develop from fetal monocytes that differentiate into long-lived cells in the first week of life via GM-CSF. J Exp Med. 2013;210(10):1977–1992. doi: 10.1084/jem.20131199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cardani A, Boulton A, Kim TS, Braciale TJ. Alveolar macrophages prevent lethal influenza pneumonia by inhibiting infection of type-1 alveolar epithelial cells. PLoS Pathog. 2017;13(1):e1006140. doi: 10.1371/journal.ppat.1006140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ghoneim HE, Thomas PG, McCullers JA. Depletion of alveolar macrophages during influenza infection facilitates bacterial superinfections. J Immunol. 2013;191(3):1250–1259. doi: 10.4049/jimmunol.1300014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacLean JA, et al. Sequestration of inhaled particulate antigens by lung phagocytes. A mechanism for the effective inhibition of pulmonary cell-mediated immunity. Am J Pathol. 1996;148(2):657–666. [PMC free article] [PubMed] [Google Scholar]
- Nakamura T, et al. Depletion of alveolar macrophages by clodronate-liposomes aggravates ischemia-reperfusion injury of the lung. J Heart Lung Transplant. 2005;24(1):38–45. doi: 10.1016/j.healun.2003.10.007. [DOI] [PubMed] [Google Scholar]
- Schneider C, et al. Alveolar macrophages are essential for protection from respiratory failure and associated morbidity following influenza virus infection. PLoS Pathog. 2014;10(4):e1004053. doi: 10.1371/journal.ppat.1004053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pribul PK, et al. Alveolar macrophages are a major determinant of early responses to viral lung infection but do not influence subsequent disease development. J Virol. 2008;82(9):4441–4448. doi: 10.1128/JVI.02541-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macdonald DC, et al. Harnessing alveolar macrophages for sustained mucosal T-cell recall confers long-term protection to mice against lethal influenza challenge without clinical disease. Mucosal Immunol. 2014;7(1):89–100. doi: 10.1038/mi.2013.27. [DOI] [PubMed] [Google Scholar]
- Benoit A, Huang Y, Proctor J, Rowden G, Anderson R. Effects of alveolar macrophage depletion on liposomal vaccine protection against respiratory syncytial virus (RSV) Clin Exp Immunol. 2006;145(1):147–154. doi: 10.1111/j.1365-2249.2006.03114.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nayak DK, et al. Long-term persistence of donor alveolar macrophages in human lung transplant recipients that influences donor specific immune responses. Am J Transplant. 2016;16(8):2300–2311. doi: 10.1111/ajt.13819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sekine Y, et al. Role of passenger leukocytes in allograft rejection: effect of depletion of donor alveolar macrophages on the local production of TNF-alpha, T helper 1/T helper 2 cytokines, IgG subclasses, and pathology in a rat model of lung transplantation. J Immunol. 1997;159(8):4084–4093. [PubMed] [Google Scholar]
- Borie R, et al. Pulmonary alveolar proteinosis. Eur Respir Rev. 2011;20(120):98–107. doi: 10.1183/09059180.00001311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Greenhill SR, Kotton DN. Pulmonary alveolar proteinosis: a bench-to-bedside story of granulocyte-macrophage colony-stimulating factor dysfunction. Chest. 2009;136(2):571–577. doi: 10.1378/chest.08-2943. [DOI] [PubMed] [Google Scholar]
- Happle C, et al. Pulmonary transplantation of macrophage progenitors as effective and long-lasting therapy for hereditary pulmonary alveolar proteinosis. Sci Transl Med. 2014;6(250):250ra113. doi: 10.1126/scitranslmed.3009750. [DOI] [PubMed] [Google Scholar]
- Suzuki T, et al. Pulmonary macrophage transplantation therapy. Nature. 2014;514(7523):450–454. doi: 10.1038/nature13807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimoto D, et al. Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity. 2013;38(4):792–804. doi: 10.1016/j.immuni.2013.04.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy J, Summer R, Wilson AA, Kotton DN, Fine A. The prolonged life-span of alveolar macrophages. Am J Respir Cell Mol Biol. 2008;38(4):380–385. doi: 10.1165/rcmb.2007-0224RC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maus UA, et al. Resident alveolar macrophages are replaced by recruited monocytes in response to endotoxin-induced lung inflammation. Am J Respir Cell Mol Biol. 2006;35(2):227–235. doi: 10.1165/rcmb.2005-0241OC. [DOI] [PubMed] [Google Scholar]
- Perdiguero GE, et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature. 2015;518(7540):547–551. doi: 10.1038/nature13989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bitterman PB, Saltzman LE, Adelberg S, Ferrans VJ, Crystal RG. Alveolar macrophage replication. One mechanism for the expansion of the mononuclear phagocyte population in the chronically inflamed lung. J Clin Invest. 1984;74(2):460–469. doi: 10.1172/JCI111443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misharin AV, et al. Monocyte-derived alveolar macrophages drive lung fibrosis and persist in the lung over the life span. J Exp Med. 2017. [DOI] [PMC free article] [PubMed]
- Zheng Z, et al. Donor pulmonary intravascular nonclassical monocytes recruit recipient neutrophils and mediate primary lung allograft dysfunction. Sci Transl Med. 2017;9(394) doi: 10.1126/scitranslmed.aal4508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nayak DK, et al. Zbtb7a induction in alveolar macrophages is implicated in anti-HLA-mediated lung allograft rejection. Sci Transl Med. 2017;9(398) doi: 10.1126/scitranslmed.aal1243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misharin AV, Morales-Nebreda L, Mutlu GM, Budinger GR, Perlman H. Flow cytometric analysis of macrophages and dendritic cell subsets in the mouse lung. Am J Respir Cell Mol Biol. 2013;49(4):503–510. doi: 10.1165/rcmb.2013-0086MA. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kopf M, Schneider C, Nobs SP. The development and function of lung-resident macrophages and dendritic cells. Nat Immunol. 2015;16(1):36–44. doi: 10.1038/ni.3052. [DOI] [PubMed] [Google Scholar]
- Eguiluz-Gracia I, et al. Long-term persistence of human donor alveolar macrophages in lung transplant recipients. Thorax. 2016;71(11):1006–1011. doi: 10.1136/thoraxjnl-2016-208292. [DOI] [PubMed] [Google Scholar]
- Yu YA, et al. Flow cytometric analysis of myeloid cells in human blood, bronchoalveolar lavage, and lung tissues. Am J Respir Cell Mol Biol. 2015. [DOI] [PMC free article] [PubMed]