

Abstract
The intestinal epithelial barrier separates the host from the microbiota that is normally tolerated or ignored. The breach of this barrier results in the entrance of bacteria or bacteria-derived products into the host, accessing the host circulation and inner organs leading to the uncontrolled inflammation as observed in patients with inflammatory bowel disease (IBD), that are characterized by an increased intestinal epithelial permeability.
To mimic the entrance of bacterial-derived compounds into the host, an endotoxemia model has been adopted in which lipopolysaccharide (LPS), a component of the outer cell wall of Gram-negative bacteria, were injected into mice. In this study, a sublethal dose of LPS was intraperitoneally injected and the mice were subsequently monitored for 8 h using a disease score. Furthermore, the expression levels of the inflammatory cytokines Il6, Il1b, and Tnfa were analyzed in the spleen, liver and colon by qPCR at different time points post LPS injection. This model could be useful for the studies involving investigation of immune responses after the invasion of microorganisms or bacterial-derived products caused by a barrier breach of body surfaces.
Keywords: Immunology and Infection, Issue 135, Lipopolysaccharide, endotoxemia, colitis, inflammatory bowel disease, barrier breach, cytokines
Introduction
The human intestine is colonized with a large consortium of microorganisms that forms the microbiota, who has developed a mutually beneficial relationship with the host during the evolution. In this relationship, the host provides a secure niche for the microbiota, whereas the microbiota provides vitamins, nutrient digestion and protection from pathogens to the host, where the microbiota resides1. When this beneficial relationship between the host and the microbiota is disturbed, diseases can develop, such as inflammatory bowel disease (IBD). IBD is a multifactorial chronic inflammatory disease of the intestine caused by genetic and environmental factors that occur in two major forms, Crohn's disease (CD) and ulcerative colitis (UC). Despite similarities between the two IBD forms, they are characterized by certain differences in the location and nature of inflammatory modifications. CD is a relapsing transmural inflammatory disorder that can potentially extend to any part of the gastrointestinal tract, while UC is non-transmural and is restricted to the colon. Furthermore, mutations in Nucleotide-binding oligomerization domain-containing protein 2 (NOD2), a pattern recognition receptor (PRR) that recognizes muramyl dipeptide (MDP), a component of the cell wall of most Gram-positive and - negative bacteria, is associated with CD2. Furthermore, Escherichia coli (E. coli), Listeria and Streptococci and their products were all found within macrophages in CD patients that have entered the host after a barrier breach3. When bacteria or their products enter the host during the development of CD, the immune system develops a response leading to the production of circulating anti-bacterial antibodies4. Maybe, the most convincing evidence for the role of the microbiota in the pathogenesis of IBD stems from mouse models. When animals are treated with antibiotics, or when mice are kept in germ-free (GF) conditions, the severity of the disease is reduced in most colitis models, such as in IL-10-/- mice that do not develop colitis in GF facilities5,6. Furthermore, colitis also disturbs the composition of the microbiota, which is characterized by an imbalanced composition and reduced richness called dysbiosis7. The consequence of IBD can be an increased intestinal permeability that can lead to the entrance of microbes and microbial-derived products into the host.
In animals, the application of Dextran Sodium Sulfate (DSS) induces an intestinal epithelial breach leading to an increased permeability of the epithelial barrier8. Portal LPS concentrations are elevated in animals with DSS colitis9. Interestingly, animals lacking the C type lectin receptor specific intracellular adhesion molecule-3 grabbing nonintegrin homolog-related 1 (SIGN-R1) are protected from DSS colitis and LPS-induced endotoxemia10. To further disseminate into the host, bacteria or bacteria derived products have to pass the vascular barrier11, the peritoneal cavity, in which the small and large intestine is located, the mesenteric lymph nodes and/or the liver12. To reduce the complexity of this system, a defined bacterial-derived compound was used. LPS, which causes endotoxemia after intraperitoneal (i.p.) or intravenous (i.v.) injection13 was injected into mice, to study the expression of the interleukins Il6 and Ilb and the cytokine Tnfa in response to LPS.
LPS is a pathogen-associated molecular pattern (PAMP) expressed as a cell-wall component of Gram-negative bacteria, that consists of lipid A (the main PAMP in the structure of LPS), a core oligosaccharide and an O side chain14. Toll-like receptor 4 (TLR4) expressed by dendritic cells, macrophages, and epithelial cells recognizes LPS15, that requires co-receptors for appropriate binding. The acute phase protein LPS-binding protein (LBP) binds LPS to form a complex that transfers LPS to the cluster of differentiation 14 (CD14), a glycosylphosphatidylinositol-anchored membrane protein. CD14 further shuttles LPS to Lymphocyte antigen 96 or also known as MD-2, which is associated with the extracellular domain of TLR4. The binding of LPS to MD-2 facilitates the dimerization of TLR4/MD-2 to induce conformational changes to recruit intracellular adaptor molecules to activate the downstream signaling pathway14, which includes the myeloid differentiation primary response gene 88 (MyD88)-dependent pathway and the TIR domain-containing adaptor-inducing interferon-β (TRIF)-dependent pathway16. The recognition of LPS by TLR4 then activates the NF-κB pathway and induces the expression of proinflammatory cytokines, such as TNFα, IL-6, and IL-1β17.
In particular, when LPS is injected into animals, the concentration of LPS given to the animals, the genetic background of the animal and the diet has to be considered. High concentrations of LPS leads to a septic shock, characterized by hypotension and multiple organ failures, and finally to death18. Mice are less sensitive to LPS compared to humans, where LPS concentrations between 2-4 ng/kg body weight (BW) are able to induce a cytokine storm19. For mice, the lethal dose (LD50), which induces death in half of the mice ranges from 10-25 mg/kg BW20 depending on the mouse strain used. For the commonly used mouse strains, C57Bl/6 and BALB/c, the lethal dose 50% (LD50) is 10 mg/kg BW. In contrast, the strains C3H/HeJ and C57BL/10ScCr are protected from LPS induced endotoxemia, which is due to mutations in Tlr421. Consequently, Tlr4-deficient mice are hyporesponsive to injections with LPS22. Other genetically modified mouse lines, such as PARP1-/- mice23 are resistant to LPS-induced toxic shock.
The mouse model described here uses a sublethal dose of LPS administered systemically to mimic the consequences of LPS dissemination after a barrier breach of the body`s surfaces. The chosen LPS concentration (2 mg/kg BW) did not induce mortality in C56Bl/6 mice, but the induced release of pro-inflammatory cytokines.
Protocol
Mice were bred and kept under specific pathogen-free (SPF) conditions in the animal facility of Department of Biomedicine, University of Basel (Basel, Switzerland). All mouse experiments were performed in accordance with the Swiss Federal and Cantonal regulations (animal protocol number 2816 [Canton of Basel-Stadt]).
1. Preparation of LPS Solution
Open the stock of LPS purified from Escherichia coli 0111:B4 under sterile conditions and reconstitute it in water to the concentration of 5 mg/mL.
Dilute the LPS stock with the sterile phosphate buffered saline (PBS) to the working concentration of 0.2 µg/µL.
2. Intraperitoneal Injection of LPS to Mice
Aspirate the LPS working solution in a 0.5 mL syringe with a 30 G needle in a laminar air flow.
Weigh female C57Bl/6 mice (six to 10 weeks of age) and calculate the amount of LPS that will be injected: 10 µL (2 µg)/g body weight. NOTE: For example, if a mouse weighs 20 g, then 20 g x 10 µL = 200 µL LPS working solution needs to be injected.
Handle the mouse gently but firmly and restrain the animal in one hand. Ensure that the animal is able to breathe normally but not to twist and turn during the restraint.
Tilt the mouse nose slightly towards the floor to expose the abdomen for injection. Locate the midline of the abdomen and the injection place in the low abdomen left or right from the midline. Inject i.p. PBS instead of LPS into control mice.
With the free hand, inject the appropriate volume (the volume calculated in step 2.2) of the LPS working solution. Make sure that the needle has entered the peritoneal cavity as otherwise, mis-injections into the abdominal muscle layer may occur. Still, do not go too deep with the needle into the peritoneal cavity to avoid injuring inner organs located in that area.
Return the animal carefully to the cage with up to 5 mice per cage.
3. Monitor the Mice
Monitor the animals at the time of injection and every 2 h after the injection for 8 h for the occurrence and severity of endotoxemia. Therefore, use a score sheet (Table 1) that has been adapted from a previously published manuscript 24.
- Score the LPS injected mice for the following parameters listed in Table 1
- Check for the appearance of the fur which is normally smooth. If the fur appears ruffled, spiky or puffy fur which indicates discomfort and/or pain, score them as 1, 2, or 3.
- Check for the activity of the mouse. NOTE: Mice normally move freely around the whole cage eating, drinking, climbing, but suppressed or restricted activity could be a sign of pain.
- Check for the level of consciousness. NOTE: Mice are very curious, and they normally move around the cage investigating the surrounding. Lack of or reduced movement can suggest pain and discomfort.
- Check the eyes. NOTE: Fully open eyes are expected in healthy mice, but partially or fully closed eyes, possibly with secretion, can be an indication of pain.
- Check for the respiration rate. NOTE: Healthy mice usually have a normal and rapid respiration rate, a reduced respiration rate could be an indication of discomfort).
- Check for the respiration quality. NOTE: Labored breathing with or without gasping is a sign of pain.
4. Tissue Sampling for Ribonucleic Acid (RNA) Extraction and Homogenization
Prepare the collection/homogenization tubes: Add 1 mL single-step RNA isolation reagent to 2 mL tubes prefilled with 1.4 mm ceramic spheres.
At the appropriate time points (before (0 h) and 2 h, 4 h and 8 h after LPS injection) euthanize the mouse with anesthetic overdose (isoflurane) followed by exsanguination and proceed with the dissection.
Cut the skin and the muscle layer exposing the abdominal cavity with the inner organs with a pair of scissors.
Carefully dissect the spleen, liver, and colon, clean the organs from fat and keep them in PBS on ice.
Cut out a small piece (about 0.5 cm long) from each organ using scissors or scalpel. NOTE: It is important to always take the piece from approximately the same part of the organ in each mouse (same lobe of liver, proximal or distal part of the colon).
Shortly dry the tissue on a piece of paper tissue and place it in the collection/homogenization tube making sure that it is completely immersed in RNA isolation reagent.
Homogenize the tissue in a high-speed benchtop homogenizer. For soft tissues like spleen, liver, and colon use 1 cycle of homogenization at speed 6.00 for 30 s.
Incubate the samples for 5 min at room temperature.
Carefully place the tube in the liquid nitrogen to snap freeze the tissue lysate. Store the snap frozen lysate at -80 °C until the RNA isolation.
5. RNA Extraction from Tissue
NOTE: RNA isolation reagents, chloroform, and alcohols are considered as hazardous material. Perform RNA extraction under a chemical hood. Use certified RNase-free reagents and equipment to maintain an RNase-free environment.
- Phase separation
- Thaw the frozen tissue lysate on ice.
- Centrifuge the sample on 1,000 x g for 5 min at 4 °C to pellet the remaining tissue particles that might be left.
- Transfer the supernatant to a new nuclease-free 1.5 mL tube.
- Add 200 µL ice-cold chloroform per 1 mL RNA isolation Reagent and shake vigorously by hand for 10-15 s.
- Incubate for 2-3 min at room temperature.
- Centrifuge at 12, 000 x g for 15 min at 4 °C. NOTE: The sample will now contain 3 phases: lower red phenol-chloroform phase, the interphase and the upper colorless aqueous phase. The RNA is present in the upper aqueous phase.
- Collect the upper aqueous phase ( 50% of total volume) and transfer into a new nuclease-free 1.5 mL tube.
- RNA precipitation
- Add 0.5 mL ice-cold isopropanol per 1 mL RNA isolation reagent and mix gently by inverting the tube 5-6 times.
- Incubate for 10-15 min at room temperature.
- Centrifuge at 12,000 x g for 10 min at 4 °C. NOTE: RNA pellet should be visible at this stage.
- Washing of RNA pellet
- Completely remove the supernatant containing the isopropanol without disturbing the pellet.
- Wash the pellet with 1 mL ice-cold 75% ethanol per 1 mL RNA isolation reagent used for the initial lysis.
- Vortex the sample briefly.
- Centrifuge the sample at 7,500 (or 12,000) x g for 5 min at 4 °C.
- Dissolving RNA
- Completely remove the ethanol in the supernatant without disturbing the pellet.
- Leave the opened tube under the chemical hood for 3-5 min to air-dry the RNA pellet at room temperature (do not over-dry as in that case it will be difficult to dissolve RNA).
- Dissolve the RNA pellet in 20-50 µL nuclease-free water by pipetting carefully up and down.
- Incubate the sample at 55 °C for 10-15 min to further improve the solution of the RNA. NOTE: At this stage, RNA can be stored at -80 °C till further analysis.
6. Digestion of Remaining DNA
Measure the RNA concentration with a spectrophotometer by pipetting an aliquot (2 µl) into the spectrophotometer and adjust to the recommended concentration.
Start the digestion of contaminating DNA with max. 10 µg RNA in 50 µL nuclease-free water.
Add 0.1 volume of 10x DNase I Buffer and 1 µL rDNase I per sample and mix gently by pipetting up and down.
Incubate at 37 °C for 20-30 min.
Vortex DNase Inactivation Reagent and add 0.1 volume to the sample mixing well with the pipette.
Incubate for 5 min at room temperature mixing occasionally by hand.
Centrifuge at 10,000 x g for 1.5 min.
Transfer the supernatant containing DNA-free RNA into the new nuclease-free tube.
Measure the RNA concentration and quality by spectrophotometer.
7. Reverse Transcription
Use a commercially available Deoxyribonucleic acid cDNA reverse transcription (RT) kit for reverse transcription. NOTE: Here, up to 2 µg of RNA can be reversed transcribed.
Calculate the volume, which contains 2 µg RNA. Pipette 2 µg RNA in a new PCR tube.
Add nuclease-free water to the sample in the PCR tube to the final volume of 10 µL.
Prepare 2x Master Mix by mixing the following components listed in Table 2
Add 10 µL 2x Master Mix to 10 µl RNA to obtain 1x Master Mix in the total volume of 20 µL.
Close the polymerase chain reaction (PCR) tube and spin down the sample briefly.
Place the samples in a PCR Thermocycler and set the settings listed in Table 3.
Store the generated cDNA at -20 °C for further analysis.
8. Quantitative PCR (qPCR)
Perform the qPCR reaction with a commercially available kit.
Self-design and test the intron-spanning primers, before usage, with different concentrations of template cDNA. Analyze the efficacy of the primer by creating a standard curve and use the primers with high efficacy and correct melting curves. NOTE: The sequences of primers used in this experiment are listed in Table 2.
Prepare the quantitative polymerase chain reaction (qPCR) reaction in the 384-well plate with the reaction setup described in Table 4.
Keep all solutions and the plate on ice using aluminum foil to prevent the plate getting wet when the reaction mixture is prepared.
Perform all the reactions in triplicates.
Carry out the PCR reaction on a thermocycler using the setting listed in Table 5.
Calculate the average Ct value for all reactions from the triplicates. Normalize all target genes to the expression level of glycerinealdehyde-3-phosphate-dehydrogenase (Gapdh) housekeeping gene by calculating 2^(-ΔCt). NOTE: All the target genes with the average Ct >35 were considered to be under the detection limit.
Representative Results
To mimic the consequences for the host after the entrance of bacteria or bacterial-derived products that occurs after intestinal barrier breach, LPS was injected into C57Bl/6 mice in a sublethal dose (2 µg/g body weight). Every single mouse was monitored and scored for the occurrence of endotoxemia with parameters listed in score sheet that includes, the appearance of the mice, the activity of the animals, the condition of eyes, and the respiration rate and quality (Table 1). The animals showed clinical signs of disease that peaked 6 to 10 h after the injection of LPS and recovered within 24 h (Figure 1). Individual mice were sacrificed 2 h, 4 h, and 8 h after the injection of LPS. Tissue pieces were collected from spleen, liver, and colon. RNA was isolated from these organs and reverse transcribed to cDNA. The cDNA was used as a template for qPCR to determine the expression levels of Il6 (Figure 2A) Il1b (Figure 2B), TNFa (Figure 2C) and Il10 (Figure 2D) with primers used for the qPCR listed in Table 6. The expression of Il6 peaked 2h after injection in spleen and colon, and 4 h after injection in the liver. The expression of Il1b, TNFa, and Il10 peaked 4 h after injection in spleen, liver, and colon. Within 8 h after injection, the expression of Il6, Il1b, TNFa, and Il10 returned to the baseline levels.
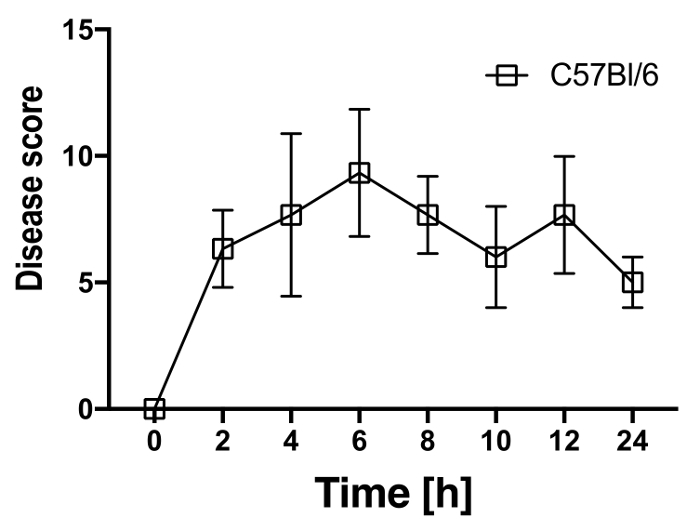
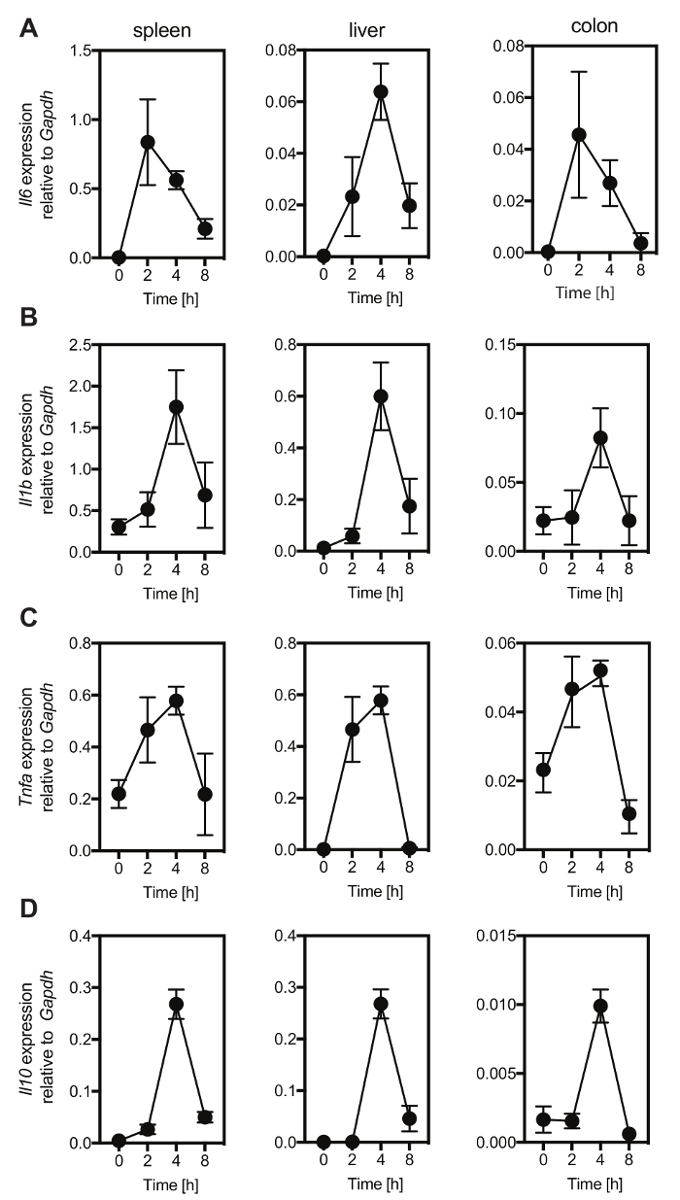
Figure 1: LPS injection (2 µg/g body weight) induced clinical signs of endotoxemia in C57Bl/6mice. After injection of 2 µg/g body weight of LPS, individual mice were monitored with the disease score presented in Table 1 that includes the appearance and the activity of the animals, opening of their eyes and their respiration rate and quality every 2 h for 12 h and 24 h after LPS injection. The disease score (y-axis) was blotted to the respective time (x-axis). Mean ± SD for 4-5 mice per each time point is presented. Please click here to view a larger version of this figure.
Figure 2: LPS injection (2 µg/g body weight) induces the expression of pro-inflammatory cytokines in C57Bl/6 mice. Mice were injected with 2 µg/g body weight of LPS and the expression levels of Il6 (A), Il1b (B), Tnfa (C) and Il10 (D) were analyzed by qPCR in spleen, liver, and colon at indicated time points after injection. Cytokine expression levels were all normalized to the expression of Gapdh. Mean ± SD for 4-5 mice per each time point is shown. Please click here to view a larger version of this figure.
Table 1: Disease score sheet to monitor C57Bl/6 mice after LPS injection. Parameters that are monitored after LPS injection. Animals were monitored every 2 h after injection for a period of 12 h, and scores were given for every individual parameter listed in the table. The final score for each animal represents the sum of all individual scores. This is adapted from reference24. Please click here to download this file.
| Amount | Reagent |
| 2 µL/sample | 10x RT Buffer |
| 0.8 µL/sample | 25x deoxynucleotide (dNTP) Mix (100 mM) |
| 1 µL/sample | RT Random Primers |
| 4.2 µL/sample | nuclease-free water |
Table 2: Reagents used to prepare the master mix for reverse transcription.
| Temperature | Time |
| 25 °C | 10 min |
| 37 °C | 120 min |
| 37 °C | 5 min |
| 4 °C | hold |
Table 3: Thermocycler settings used for reverse transcription.
| Amount | Reagent |
| 5 µL/well | 2x Master Mix |
| 0.05 µL/well | Reference Dye |
| 500 nM final/well | forward primer |
| 500 nM final/well | reverse primer |
| between 10 and 100 ng/well, we used 40 ng/well | template cDNA diluted in a Template Dilution Buffer (1/100 dilution of this buffer was made in nuclease-free water and used to dilute the template cDNA). Diluting the template in this Buffer allows the visualization of the wells where template is added as the reaction color changes from blue to green |
| nuclease-free water to final volume of 10 µL/well | nuclease-free water |
Table 4: Description of the PCR reaction.
| Temperature | Time | |
| 95 °C | 2 min | |
| 95 °C | 5 s | 40 cycles |
| 60 °C | 20 s | |
| 95 °C | 15 s | melting curve analaysis |
| 60 °C | 1 min | |
| 95 °C | 15 s |
Table 5: Thermocycler settings used for PCR.
| Primer | Sequence | Melting temperature | Product length |
| Il6-F | TCG GAG GCT TAA TTA CAC ATG TTC T | 60.3 | 94 bp |
| Il6-R | GCATCATCGTTGTTCATACAATCA | 58.2 | |
| Il1b-F | TGTGAAATGCCACCTTTTGA | 56.1 | 94 bp |
| Il1b-R | GTCAAAGGTTTGGAAGCAG | 57.2 | |
| Il10-F | ATCGATTTCTCCCCTGTGAA | 56.2 | 108 bp |
| Il10-R | TGTCAAATTCATTCATGGCCT | 56.1 | |
| Tnfa-F | CCACCACGCTCTTCTGTCTAC | 60.4 | 103 bp |
| Tnfa-R | AGGGTCTGGGCCATAGAACT | 60 | |
| Gapdh-F | CATCAAGAAGGTGGTGAAGC | 56.7 | 199 bp |
| Gapdh-R | CCTGTTGCTGTAGCCGTATT | 58 |
Table 6: Primers used in the qPCR reaction. The sequence of primers used for qPCR to detect mouse cytokines and housekeeping gene Gapdh.
Discussion
This protocol mimics immunological processes that occur after the invasion by microbial-derived products. Critical steps within the protocol are the selection of the mouse line, the hygiene status of the mice, the dose of LPS, the monitoring of the animals for the occurrence of endotoxemia, and the time point of experiment termination. Most importantly, the genetic background of the mouse strain has to be considered. Different mouse strains have different susceptibility to LPS. For example, the C3H/HeJ and C57BL/10ScCr mice are resistant to LPS induced endotoxemia21,25. Furthermore, the hygiene status of the animals may influence the course of endotoxemia in LPS. Specific-pathogen-free (SPF) animals are most commonly used. These mice are free from a defined list of pathogens, but the complete microbiota composition of these animals is not known26,27. Sterile germ-free or axenic animals (that do not harbor a microbiota) are distinct from SPF mice26. Likely, the colonization of axenic animals with one microorganism or with a consortium of microorganisms (gnotobiotic animals) may influence the expression of genes, such as IL-19, after LPS injection compared to SPF animals8. Moreover, a score sheet has been suggested that was used as a score sheet for a sepsis model induced by injecting the fecal solution into the peritoneal cavity24. This sheetcan be discussed with the local animal welfare committee and may be adapted to the local needs. In particular, the criteria for termination needs to be discussed with the local animal welfare committee. This experiment had to be stopped when a score > 12 is achieved or when the maximum score for one individual parameter is reached. In this experiment, no animal out of eight animals exceeded a disease score of 12 after injection of LPS (2 µg/g body weight). In this study, results showing the expression of the cytokines Il6, Il1b, TNFa and Il10 in the liver, spleen and colon after injection of LPS are presented. The expression of Il1b, TNFa and Il10 peaked 4 h after LPS injection in spleen, liver, and colon, whereas, Il6 expression peaked 2 h after LPS injection in spleen and colon and 4 h after LPS injection in the liver. Within 8 h the expression Il6, Il1b, TNFa and Il10 returned to baseline levels in spleen, liver and colon. Disease severity peaked between 6 h and 10 h after LPS injection when the expression of Il6, Il1b, TNFa and Il10 have returned already to the baseline levels indicating that increased expression of Il6, Il1b, TNFa and Il10 occurs rapidly after LPS injection, but that the kinetics of other genes may show a different pattern after LPS injection. The expression of these cytokines could also be analyzed on the protein level in the blood of the animals by ELISA, which can be considered in addition to qPCR.
Modifications and troubleshooting of the technique are necessary in cases when a disease score > 12 is reached. Therefore, the dose of LPS has to be reduced and better adjusted to the used mouse line and the local conditions to avoid premature termination of the experiment. The manipulation of genes by deleting a given genomic region or overexpressing a gene may influence the susceptibility of mice to LPS. In preliminary experiments, the dose of LPS may be titrated to the appropriate dose to avoid unnecessary harm and death to the animals in this experiment. Of note, contamination of LPS with other bacterial-derived products and mis-injections have to be avoided. Furthermore, the kinetics of the expression of genes of interest after LPS injection needs to be determined.
Limitations are that this technique provides information on the consequences of the dissemination of microbial products into the host after an intestinal barrier breach but will not give information on events that lead to intestinal barrier breach and to causes that trigger IBD. Of course, this model is not a colitis model as the DSS colitis model, but it can be useful in addition to colitis models to study the consequence of the entrance of microbial products into the host. Furthermore, the i.p. injection of LPS will disrupt the peritoneal cavity, in which the small and the large intestines are located. This may facilitate the translocation of intestinal-derived bacterial products from the peritoneal cavity into the host.
The significance of this model is the fact it uses a defined PAMP. In other models including the i.p. injection of entire bacteria, the i.p. injection of fecal solutions24 or the septic peritonitis models, in which peritonitis is induced by cecal ligation and puncture28, a vast array of microbes or their products induces disease. Furthermore, injection of LPS into mice is an uncomplicated approach and does not require surgery as in septic peritonitis induced by cecal ligation and puncture.
Further application of this model are studies that aim to investigate endotoxemia or LPS induced septicemia in any contexts characterized by the translocation of bacteria-derived products into the host, such as breach of the skin barrier or breached of the urogenital tract. In conclusion, this is a straightforward model that can be used in every laboratory setting. Depending on the local conditions, there might be adaptations to the local needs necessary. In particular, the score sheet has to be discussed with the local authorities before the experiment is performed. This approach was used to mimic consequences for the host when bacteria or bacterial-derived products enter the host after a barrier breach has occurred. This may be in particular relevant in studies examining the basis of IBD where the occurrence of an intestinal barrier breach is a common event in the pathology of the disease.
Disclosures
The authors have nothing to disclose.
Acknowledgments
JHN is supported by the Swiss National Foundation (SNSF 310030_146290).
References
- Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science. 2005;307:1915–1920. doi: 10.1126/science.1104816. [DOI] [PubMed] [Google Scholar]
- Abreu MT, et al. Mutations in NOD2 are associated with fibrostenosing disease in patients with Crohn's disease. Gastroenterology. 2002;123:679–688. doi: 10.1053/gast.2002.35393. [DOI] [PubMed] [Google Scholar]
- Liu Y, et al. Immunocytochemical evidence of Listeria, Escherichia coli, and Streptococcus antigens in Crohn's disease. Gastroenterology. 1995;108:1396–1404. doi: 10.1016/0016-5085(95)90687-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaffer T, et al. Anti-Saccharomyces cerevisiae mannan antibodies (ASCA) of Crohn's patients crossreact with mannan from other yeast strains, and murine ASCA IgM can be experimentally induced with Candida albicans. Inflamm Bowel Dis. 2007;13:1339–1346. doi: 10.1002/ibd.20228. [DOI] [PubMed] [Google Scholar]
- Sellon RK, et al. Resident enteric bacteria are necessary for development of spontaneous colitis and immune system activation in interleukin-10-deficient mice. Infect Immun. 1998;66:5224–5231. doi: 10.1128/iai.66.11.5224-5231.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gkouskou KK, Deligianni C, Tsatsanis C, Eliopoulos AG. The gut microbiota in mouse models of inflammatory bowel disease. Front Cell Infect Microbiol. 2014;4:28. doi: 10.3389/fcimb.2014.00028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaubeck M, et al. Dysbiotic gut microbiota causes transmissible Crohn's disease-like ileitis independent of failure in antimicrobial defence. Gut. 2016;65:225–237. doi: 10.1136/gutjnl-2015-309333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinert A, et al. The Stimulation of Macrophages with TLR Ligands Supports Increased IL-19 Expression in Inflammatory Bowel Disease Patients and in Colitis Models. J Immunol. 2017;199:2570–2584. doi: 10.4049/jimmunol.1700350. [DOI] [PubMed] [Google Scholar]
- Gabele E, et al. DSS induced colitis increases portal LPS levels and enhances hepatic inflammation and fibrogenesis in experimental NASH. J Hepatol. 2011;55:1391–1399. doi: 10.1016/j.jhep.2011.02.035. [DOI] [PubMed] [Google Scholar]
- Saunders SP, et al. C-type lectin SIGN-R1 has a role in experimental colitis and responsiveness to lipopolysaccharide. J Immunol. 2010;184:2627–2637. doi: 10.4049/jimmunol.0901970. [DOI] [PubMed] [Google Scholar]
- Spadoni I, et al. A gut-vascular barrier controls the systemic dissemination of bacteria. Science. 2015;350:830–834. doi: 10.1126/science.aad0135. [DOI] [PubMed] [Google Scholar]
- Balmer ML, et al. The liver may act as a firewall mediating mutualism between the host and its gut commensal microbiota. Sci Transl Med. 2014;6:237ra266. doi: 10.1126/scitranslmed.3008618. [DOI] [PubMed] [Google Scholar]
- Maier RV, Mathison JC, Ulevitch RJ. Interactions of bacterial lipopolysaccharides with tissue macrophages and plasma lipoproteins. Prog Clin Biol Res. 1981;62:133–155. [PubMed] [Google Scholar]
- Lu YC, Yeh WC, Ohashi PS. LPS/TLR4 signal transduction pathway. Cytokine. 2008;42:145–151. doi: 10.1016/j.cyto.2008.01.006. [DOI] [PubMed] [Google Scholar]
- Deng M, et al. Lipopolysaccharide clearance, bacterial clearance, and systemic inflammatory responses are regulated by cell type-specific functions of TLR4 during sepsis. J Immunol. 2013;190:5152–5160. doi: 10.4049/jimmunol.1300496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kagan JC, et al. TRAM couples endocytosis of Toll-like receptor 4 to the induction of interferon-beta. Nat Immunol. 2008;9:361–368. doi: 10.1038/ni1569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell. 2006;124:783–801. doi: 10.1016/j.cell.2006.02.015. [DOI] [PubMed] [Google Scholar]
- Cohen J. The immunopathogenesis of sepsis. Nature. 2002;420:885–891. doi: 10.1038/nature01326. [DOI] [PubMed] [Google Scholar]
- Suffredini AF, et al. Effects of recombinant dimeric TNF receptor on human inflammatory responses following intravenous endotoxin administration. J Immunol. 1995;155:5038–5045. [PubMed] [Google Scholar]
- Fink MP. Animal models of sepsis. Virulence. 2014;5:143–153. doi: 10.4161/viru.26083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poltorak A, et al. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 1998;282:2085–2088. doi: 10.1126/science.282.5396.2085. [DOI] [PubMed] [Google Scholar]
- Hoshino K, et al. Cutting edge: Toll-like receptor 4 (TLR4)-deficient mice are hyporesponsive to lipopolysaccharide: evidence for TLR4 as the Lps gene product. J Immunol. 1999;162:3749–3752. [PubMed] [Google Scholar]
- Corral J, et al. Role of lipopolysaccharide and cecal ligation and puncture on blood coagulation and inflammation in sensitive and resistant mice models. Am J Pathol. 2005;166:1089–1098. doi: 10.1016/S0002-9440(10)62329-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrum B, et al. A robust scoring system to evaluate sepsis severity in an animal model. BMC Res Notes. 2014;7:233. doi: 10.1186/1756-0500-7-233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brandwein SL, et al. Spontaneously colitic C3H/HeJBir mice demonstrate selective antibody reactivity to antigens of the enteric bacterial flora. J Immunol. 1997;159:44–52. [PubMed] [Google Scholar]
- Macpherson AJ, McCoy KD. Standardised animal models of host microbial mutualism. Mucosal Immunol. 2015;8:476–486. doi: 10.1038/mi.2014.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masopust D, Sivula CP, Jameson SC. Of Mice, Dirty Mice, and Men: Using Mice To Understand Human Immunology. J Immunol. 2017;199:383–388. doi: 10.4049/jimmunol.1700453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wirtz S, et al. Protection from lethal septic peritonitis by neutralizing the biological function of interleukin 27. J Exp Med. 2006;203:1875–1881. doi: 10.1084/jem.20060471. [DOI] [PMC free article] [PubMed] [Google Scholar]