

Abstract
The incidence of complex metabolic diseases has increased as a result of a widespread transition towards lifestyles of increased caloric intake and lowered activity levels. These multifactorial diseases arise from a combination of genetic, environmental, and behavioral factors. One such complex disease is Metabolic Syndrome (MetS), which is a cluster of metabolic disorders, including hypertension, hyperglycemia, and abdominal obesity. Exercise and dietary intervention are the primary treatments recommended by doctors to mitigate obesity and its subsequent metabolic diseases. Exercise intervention, in particular aerobic interval training, stimulates favorable changes in the common risk factors for Type 2 Diabetes Mellitus (T2DM), Cardiovascular Disease (CVD), and other conditions. With the influx of evidence describing the therapeutic effect exercise has on metabolic health, establishing a system that models exercise in a controlled setting provides a valuable tool for assessing the effects of exercise in an experimental context. Drosophila melanogaster is a great tool for investigating the physiological and molecular changes that result from exercise intervention. The flies have short lifespans and similar mechanisms of metabolizing nutrients when compared to humans. To induce exercise in Drosophila, we developed a machine called the TreadWheel, which utilizes the fly's innate, negative geotaxis tendency to gently induce climbing. This enables researchers to perform experiments on large cohorts of genetically diverse flies to better understand the genotype-by-environment interactions underlying the effects of exercise on metabolic health.
Keywords: Genetics, Issue 136, Exercise, Drosophila, aerobic interval training (AIT), Metabolic Syndrome, obesity, triglyceride storage, negative geotaxis, genotype-by-environment interaction
Introduction
Childhood and adult obesity are growing epidemics in cultures that consume high caloric diets and remain idle for extended periods of time, which can lead to severe long-term consequences, including insulin resistance, chronic inflammation, and osteoarthritis1,2,3,4. The prevalence of these disorders continues to rise due to the growing imbalance in caloric intake and expenditure attributed to elevated consumption of fats and sugars and a primarily sedentary lifestyle5. Correspondingly, this energy imbalance has led to an increase in cases of Type 2 Diabetes Mellitus (T2DM) and Cardiovascular Disease (CVD)5. Individuals are at a higher risk of developing both disorders if they have been diagnosed with the disorder Metabolic Syndrome (MetS), whose symptoms include abdominal obesity and dyslipidemia2. MetS is shaped by the complex interactions between genotype and various environmental factors, such as diet and exercise6. Thus, to gain a full understanding of the underlying mechanisms of this complex disease, all of these factors should be considered.
When it comes to combating MetS, doctors first recommend undergoing lifestyle changes that include eating a healthy, well-balanced diet and physical activity2,7,8. Since effective medications are limited and gastric bypass surgery is costly and requires lifelong medical monitoring, pharmacological and surgical intervention is only recommended for severe cases and only in combination with these lifestyle changes3,7,8. While lifestyle interventions, such as exercise and dieting, can produce and maintain long-term weight loss goals, whether or not these corrective measures can fully ameliorate the negative effects associated with MetS needs further study7,8.
Mouse models have been used to examine the effects of exercise on metabolic diseases for years; however, the introduction of exercise to fly research on MetS is a relatively recent endeavor9,10,11,12. Flies provide the perfect vehicle to study exercise in a controlled laboratory setting, since they are easily manipulated, have a short lifespan, are inexpensive to maintain, and energy-related metabolic pathways are highly conserved between Drosophila and humans13. D. melanogaster genomes are well characterized, and there is a wealth of genetics tools available for use in Drosophila that can provide insight into various genotypes and genotype-by-environment interactions that could modulate the effect of exercise on organismal health14.
Current methods of exercising Drosophila invoke the fly's innate, negative geotaxis tendency, the behavioral instinct to climb upwards, to stimulate climbing in adults within their enclosures11,12,15. The Power Tower, one method to stimulate exercise in flies, systematically raises the fly enclosures vertically and then drops them back to the bench surface, effectively knocking the flies to the bottom of vial, thus inducing their instinctual negative geotaxis12,16. Experiments conducted using this machine showed that exercise is a powerful protective factor against many age-related diseases, including CVD and T2DM, and promotes healthy aging12,17,18. Specifically, they demonstrated that exercise can reduce the onset of age-related mobility decline in flies and improve multiple age-related symptoms, such as cardiac performance and stress response17,18. However, control flies never placed on the tower showed higher climbing scores than those experiencing the machine, suggesting the force of the repetitive drops may be injuring the flies and affecting mobility12. This suggests that an alternative method of inducing exercise that is less forceful and avoids causing physical trauma would be a useful, complementary method to the Power Tower protocol16.
To gently induce exercise in Drosophila, we developed an exercise machine called the TreadWheel (Figure 1). The TreadWheel's (henceforth abbreviated as TW) rotational motion triggers the flies' innate, negative geotaxis tendency by constantly redefining the gravitational top of the vials, which, in turn, stimulates climbing in the flies. Unlike other methods, the rotational motion of the TW is inherently gentle, which minimizes the number of additional stressors that may arise and impact the results.Thus, the machine provides the means to induce exercise in large numbers of flies without inducing stress, which will enable researchers to study the effects of exercise on metabolic health (Figure 2), aging, sleep, and many other topics11.
Our method follows an inverse pyramidal, interval training protocol, which largely incorporates aspects of aerobic interval training (AIT) with some endurance exercise training. The standard AIT regime is altered in this protocol to gradually increase the duration of each interval over a five-day period to promote endurance. AIT has been particularly useful in preventing MetS relative to other intervention methods and was more effective in reversing Metabolic Syndrome's common risk factors than continuous moderate exercise19,20. However, a disadvantage of the TW relative to the Power Tower is that flies more quickly habituate to the rotating motion, thus variation in innate tendency to habituate among the flies can complicate the interpretation of the benefits of exercise.11 An elegant solution to this limitation is described by Watanabe and Riddle15 and their complementary article21.
Protocol
1. TreadWheel Setup and Operation
NOTE: See Supplemental Figure 1 for schematics on TW exercise machine construction. The parts required are listed in the Table of Materials. The phrases "TW", "TreadWheel", "exercise machine", and "machine" are used interchangeably throughout the protocol.
- Calibrate motor using power switch such that a full rotational period is 15 s (4 rpm). Calibrate machine rotation speed before placing fly vials into secured clamps attached to rotating arms.
- Adjust speed using a standard stopwatch and an immobile object temporarily attached to the machine, such as a small paint brush taped to the motor cover with the brush just touching one of the rotating brackets. Time the number of rotations in one minute using the immobile object as a reference point and adjust speed to achieve a 15 s rotational period (4 rpm).
2. Fly Collection and Maintenance
NOTE: All flies are maintained in a 25 °C incubator with 50% humidity and a 12 h light/dark cycle between benchtop manipulations described below. Flies are fed a standard cornmeal-molasses lab diet unless otherwise noted.
- Prepare laying chambers for each genotype of interest using apple-juice agar plates, a drop of yeast paste per plate, and 6 oz. plastic bottles to collect eggs that will hatch into larvae.
- To prepare apple-juice agar plates, fill 35 mm x 10 mm Petri dishes ¾-full with 3% agar dissolved into store-bought apple juice. Store solidified plates in a refrigerator until use.
- In a 50 mL conical centrifuge tube, combine store-bought active dry yeast with distilled water at a ratio of 2 mL of water for each gram of yeast. Mix yeast with water to create a paste with the consistency of ketchup using a glass stirring rod. Adjust yeast and water proportions as needed to achieve the desired consistency.
- Store yeast paste in a refrigerator between uses. Keep the tube cap loose to allow de-gassing.
- Use a sharp needle or knife to punch small holes into the 6 oz. square-bottom bottles (polypropylene) as ventilation holes for the laying chambers.
- Dab a small spot of yeast paste (~3 mm in diameter) onto each apple-juice agar plate.
Place adult flies of the desired genotype into the 6 oz. bottle and cap bottle with the apple-juice agar plate. Secure the plate to bottle with rubber bands. Invert, and place in an incubator to allow adults to lay eggs.
Change apple-juice agar plates on a laying chamber twice a day (morning and evening). Leave used plates in the incubator for another 12–24 h to allow the first instar larvae to hatch.
Collect first instar larvae using a thin paintbrush or collection pick (flexible thin needle attached to a handle). Place 50 first instar larvae of a specific genotype on a food vial of the desired experimental food type to develop and complete metamorphosis while housed in the incubator (Figure 2A).
- Collect adults for exercise treatment (Figure 2A).
- Upon pupation, collect pupae with a small, damp paintbrush and place in empty vials prior to eclosion to prevent adult flies from being exposed to the larval diet.
- Transfer adult flies to food vials of standard lab diet as they eclose. One to five days after eclosion, separate flies based on sex.
Designate half of the collected adults each to the experimental and control groups. Maintain flies in vials of standard lab diet at a concentration of 50 flies per vial.
Transfer adults to fresh food vials every two days to prevent them from getting stuck during exercise. Add several grains of supplemental live yeast to the new food immediately before introducing the flies. NOTE: While it is possible to maintain flies without a live yeast supplement, we have found that the adult flies perform best with supplementation. Live yeast supplementation is a variable that could be modified to suit the researcher's experimental goals.
3. Exercise Protocol
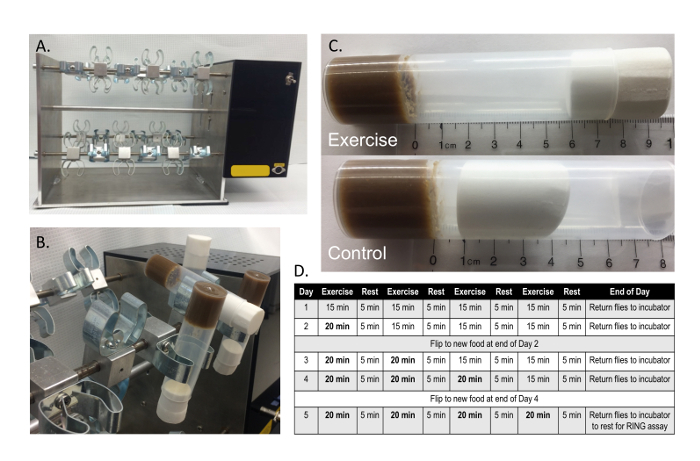
On the first day of exercise (Day 1), push vial plugs to leave 1 cm of space in control vials and 6 cm of space between the food and the plug in exercise vials (Figure 1C). NOTE: The control vials will have limited space for flies to move while on the exercise machine and thus flies will engage in much less movement than in exercise vials.
Place vials in the clamps, and then give the flies 10 min to acclimate on the machine (Figure 1B). NOTE: An alternative control is to maintain the unexercised flies on the benchtop adjacent to the exercise machine with 6 cm of space (in the vial) to move normally. Similar results of exercise relative to both the "1 cm on TreadWheel" and the "6 cm on benchtop" controls have been found11.
- Treat both control and exercise flies to rotation on the machine at constant room temperature for 5 consecutive days per week with alternating bouts of rotation and rest (Figure 1D). NOTE: A constant temperature is needed to avoid the confounding effects of the temperature on activity levels. For temperature manipulation studies, place the TW inside an incubator to modify temperature treatments during exercise training.
- Run the machine for four 15 min bouts of exercise on Day 1. Alternate these bouts with 5 min rest periods.
- Each of the following days, add 5 min of exercise to one of the preexisting exercise bouts. For example, on Day 2, make the first exercise bout 20 min long, but maintain the rest of the bouts at 15 min long.
- On Day 3, exercise flies for 20 min during the first two bouts and 15 min for the following two bouts.
- On Day 4, use 20 min exercise bouts for all except the final bout, which should only be 15 min long.
- On Day 5, keep all exercise bouts 20 min long. NOTE: During rest periods, the vials containing the flies should remain securely clamped on the TW. The orientation of the vials during these periods is not relevant because the gravitational force experienced by all flies will be coming from the same direction.
After completing the exercise treatment for a given day, return the food vial plugs to their normal positions and return flies to their incubators until the next day.
After completion of the full exercise regime, anesthetize flies using standard fly CO2 anesthesia methods. Then transfer flies to new food vials or microcentrifuge tubes for assessment of phenotypic characteristics of interest. NOTE: Example phenotypes include climbing ability (food vial), triglyceride storage (microcentrifuge tube), or gene expression (microcentrifuge tube). The protocol can be paused here with samples stored appropriately depending on the assessments to be performed. Assays involving living flies will need to be conducted promptly in accordance with the specific goals of the phenotypic assay.
4. Climbing Assessment
- After one day of rest following completion of the exercise protocol, test general fly climbing performance using a rapid iterative negative geotaxis (RING)-like negative geotaxis climbing assay22.
- Place groups of 10 flies in empty vials using CO2 anesthesia (or tapping directly from food vials if already sorted) and seal with paraffin film to prevent the flies from escaping. Give the flies at least 10 min to acclimate to the new vial. To prevent vials from tipping over, tape a ~5 cm diameter cardstock circle to the bottom of vial to provide extra surface area. NOTE: Vial plugs should not be used because they impede part of the vial used to determine height climbed.
- Place vials 20 cm in front of a 1 cm x 1 cm grid in full view of a mounted, stationary camera with a built-in timer (a smart phone with a timer camera application works well).
- Tap vials gently on the counter three times to knock all flies down to bottom of the vial. After tapping the vial down the third time, immediately start the 4 s camera timer to capture an image to determine the height climbed by each fly. NOTE: Tapping should be done at a consistent intensity and by the same researcher in each exercise-control contrast to minimize variation in the flies' responses due to tapping.
- Repeat step 4.1.3 twice, with at least 1 min rest between trials, to produce a total of three trials per vial.
- To preserve the flies for other analyses, immediately after the climbing assay has been completed, flash freeze the flies by immersion in liquid nitrogen.
- Use a shallow, open-topped dewar flask filled to a depth of ~5 cm with liquid nitrogen.
- Transfer flies to small microcentrifuge tubes using CO2 anesthesia and then place tubes in the dewar flask filled with liquid nitrogen. Alternatively, if future analyses require avoidance of CO2, snap freeze the flies directly in their climbing vials by gently tapping flies to the bottom of their vial while the bottom is submerged in liquid nitrogen, and then transfer the frozen flies to microcentrifuge tubes using forceps. NOTE: Liquid nitrogen is cryogenic and should only be used with proper protective equipment and room ventilation.
- Process images using the ImageJ23 multi-point selection tool.
- Open up an image for processing within ImageJ.
- Set a 1 cm scale based on the background grid paper to calculate the distance climbed by each individual fly. Use the "Line Tool" from the tool bar to trace one side of a 1 cm2 square from the background grid paper. Click on the "Analyze" tab and select "Set Scale". Set the "Known Distance" as '1.00' and "Unit of Length" as 'cm', make sure "Global" is unchecked, and click on "OK".
- Select the "Multi-point Selection" icon from the tool bar and zoom in on the bottom of the vial in the image. Set the first point as the very bottom of the vial by clicking on the lowest part of the vial.
- Click on the center of each fly in the vial to mark it as a data point. Take note that there are a total of 11 points per vial, one marking the bottom of the vial and one for each fly.
- Click on the "Analyze" tab and select "Measure" to generate a table of measured values. Save table in as a csv file.
- Open the csv file in a spreadsheet program and calculate the exact distance climbed by each fly by subtracting the y-values of points 2-11 from point 1 in the measured values table.
- Repeat steps 4.3.1 through 4.3.6 for each image.
5. Triglyceride Storage Assay
NOTE: The samples, standards, glycerol standard solution, and triglycerol working solution used throughout the assay should be kept on ice for the duration of the protocol and should be stored in the refrigerator when not in use.
- Prepare standards using the Glycerol Standard solution.
- Make the Blank (B) by adding 1,000 µL of homogenization buffer to a microcentrifuge tube labeled B.
- Make Standard 1 (S1), 2.5 mg/mL per well, by adding 1,000 µL of the glycerol standard solution to a microcentrifuge tube labeled S1.
- Make Standard 2 (S2), 1.25 mg/mL per well, by adding 500 µL of the glycerol standard solution to 500 µL of diH2O in a microcentrifuge tube labeled S2.
- Make Standard 3 (S3), 0.625 mg/mL per well, by adding 250 µL of the glycerol standard solution to 750 µL of diH2O in a microcentrifuge tube labeled S3.
- Make Standard 4 (S4), 0.3125 mg/mL per well, by adding 125 µL of the glycerol standard solution to 875 µL of diH20 in a microcentrifuge tube labeled S4.
- Prepare triglycerol working solution from the Serum Triglyceride Determination Kit.
- Add 40 mL of diH2O to the free glycerol reagent from the kit and mix by inverting bottle.
- Add 10 mL of diH2O to the triglyceride solution from kit and mix by inverting bottle.
- Combine glycerol reagent and triglyceride solution and mix by inverting to prepare the triglycerol working solution. NOTE: The triglycerol working solution is good for 60 days and can complete four 96-well plate assays per kit using the protocol below. The volumes used are proportionally scaled down from the official Serum Triglyceride Determination kit for use in the 96-well plate format.
- Verify Blank and Standards by carefully pipetting 5 µL of each standard into a 96-well flat bottomed, clear microplate in triplicate (or quadruplicate for the blank).
- Add exactly 125 µL of Triglycerol Working Solution to each well containing a standard. Allow solutions to react for 30 min.
- Place a 96-well plate in a spectrophotometer and read the absorbance at 540 nm. Use the absorbance measurements to create a standard curve and verify the accuracy of standards by evaluating the R2 value. If the R2 for the standards is less than 0.98, re-prepare the standards more carefully.
- Begin Day 1 of the assay for triglyceride storage14 using microcentrifuge tubes containing 10 frozen flies each. NOTE: A repeat or multi-channel pipette should never be used in this step because the amounts of each reagent used in the assay must be very accurate and consistent to get reproducible results. In our experience, repeat and multi-channel pipettes do not have this degree of accuracy and consistency. Each aliquot from a single channel pipette can be visually confirmed by the experimenter for quality control and any single channel pipette-specific biases will be experienced by all samples to the same degree.
- Prepare a 200 mL stock of Homogenization Buffer by combining 0.272 g of KH2PO4, 400 µL of 0.5 M EDTA, and 199.6 mL of diH2O in a glass bottle.
- Add exactly 100 µL of homogenization buffer to each of the 40 microcentrifuge tubes containing 10 adults frozen after the climbing assay. Then, centrifuge samples for 30 s at 18,000 g.
- Grind flies using a motorized grinder and pestle (or another form of tissue homogenization) to prepare a milky solution. Make sure a fresh pestle is used for each sample. Then, centrifuge samples for 2 min at 18,000 g.
- Pipette only the top 75 µL of supernatant fluid from each tube into new microcentrifuge tubes. Ensure that no fly bits from the pellet are transferred over. Place new tubes in the refrigerator overnight.
- Begin Day 2 of the triglyceride protocol by removing Day 1 samples from the refrigerator. NOTE: Step 5.5 can be performed the same day as 5.4 but we have found the signal for triglyceride concentration to be more robust after a night in the refrigerator. However, the time between Steps 5.4 and 5.5 should not exceed 36 h.
- (Optional) Vortex the first tube with supernatant fluid for 10 s and pipette 5 µL into a clean microcentrifuge tube. Add 95 µL of 0.15 M NaCl into the new microcentrifuge tube. Store these subsamples in the -20 °C freezer for a later assay of protein if desired. NOTE: These samples of supernatant with the added NaCl solution can be used to determine protein content using the researcher's preferred assay. We use the Bradford Method24. The protein content is one way to standardize the measure of triglyceride content, but the researcher should also be cautious in interpreting these ratios since exercise and diet can also affect protein storage.
- For the original supernatant vials, vortex the first tube with supernatant fluid for 10 s, then pipette 5 µL into two separate wells of a 96-well microplate to produce technical replicates. Be sure to note which wells are used for each sample.Repeat this step for the 39 other tubes.
- Pipette exactly 5 µL of the blank into four wells of the microplate, and for the remaining wells, pipette exactly 5 µL of each standard in triplicate.
- Add exactly 125 µL triglycerol working solution to each well, and allow solution to react for 30 min.
- Place the 96-well plate in a spectrophotometer and read the absorbance at 540 nm.
- Convert absorbance values into concentration (mg/mL) using the standard curve calculated from the standards.
Representative Results
We are especially interested in identifying the factors that contribute to the overall metabolic health of an individual. It was previously found that genotype-by-diet interactions contribute substantially to population level variation in metabolic traits14. This means that each genotype responds to environmental differences in a unique and complex manner. To extend our work on genotype-by-environment effects to include physical exercise, we developed the TreadWheel, which is capable to exposing large numbers of genotypes to aerobic interval training (AIT) in a high-throughput manner.
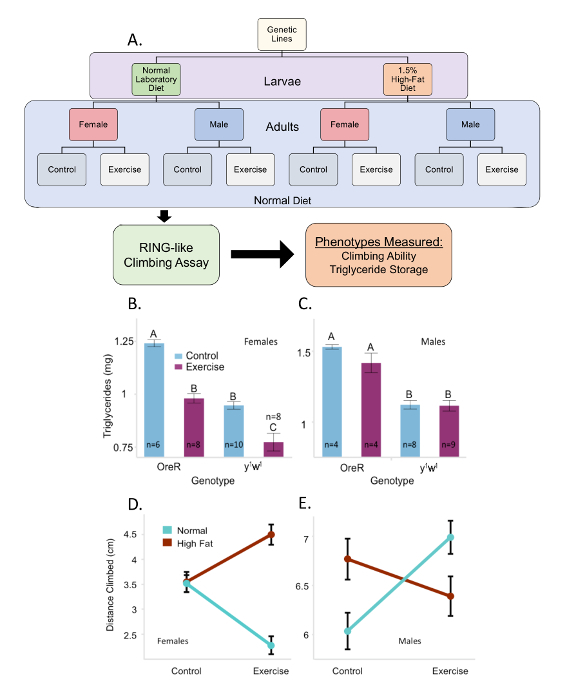
To establish whether exercise on the TW influenced metabolic traits, we measured triglyceride storage in Oregon-R (OreR) and y1w1 flies, common wildtype flies (Figure 2B–C), and normalized those values against the flies' protein concentration, as originally reported in Mendez et al.11. We analyzed the data by multivariate analysis of variance (MANOVA) accounting for genotype, sex, exercise treatment (and their interactions), and experimental block effects such as time replicates and food vial and found that there was a significant genotype-by-exercise interaction (p = 0.0017) affecting triglyceride storage. There was a significant sexual dimorphic effect between males and females, with males storing more triglycerides than females (p < 0.0001). We saw that in females, exercised flies had significantly lower triglyceride levels than their unexercised counterparts (Figure 2B, p <0.0001). In the males, while the decrease in triglyceride storage observed in Oregon-R exercised flies (as compared to controls) was not statistically significant, a significant difference in triglyceride storage was observed between the two separate lines (Figure 2C, p <0.0001). Note that while standardizing triglyceride concentration against the protein concentration provides insight on overall fly body composition ratios, comparison of direct triglyceride or protein concentration among different groups of flies can also provide specific information on the effect of exercise on these phenotypes individually.
Since variables like adult exercise, sex, and genotype are shown to affect triglyceride storage, it was expected that these factors also impact other phenotypes and interact with diet. We raised larvae from a representative wildtype Drosophila Genetic Reference Panel line (DGRP 153)25 on either a high fat or normal diet and induced exercise in adult flies for one week (Figure 2A). Subsequently, we performed a RING-like negative geotaxis assay to measure climbing ability. The climbing assay used differed from a standard RING assay; instead of a RING apparatus, vials with paraffin film covering the openings were used to house flies during the assay. Other aspects of the original RING assay, such as time between tapping the flies to the bottom of the vial and taking the photo, were retained22.
All treatments were repeated three separate times with a minimum of 59 individual flies per time replicate and treatment. Data was analyzed by MANOVA accounting for diet, genotype, sex, exercise treatment (and their interactions), as well as experimental block effects of time replicate, assay vial, and vial assay replicate. We found that the exercised females climbed significantly higher (p <0.005) when reared on the high fat diet than any of the other female treatments (Figure 2D). For the males, it was seen that exercise only improved climbing when males were raised on the normal diet, and those raised on the high fat diet showed no significant change (Figure 2E). We also found significant sexually dimorphic effects (p <0.0001) in climbing with males climbing higher than females. The surprising result of a decrease in climbing performance following exercise for females from the DGRP 153 line (Figure 2D) for females consuming a normal diet (p <0.0001) is an example of how this type of exercise may not be a uniformly positive intervention for all genotypes and could be contingent on other environmental factors. In Mendez et al.11, females from four other genetic lines tested for their climbing performance after having been raised on a normal diet all showed enhanced climbing ability with exercise training. This suggests that the response observed in DGRP 153 is genotype-specific and not a general property of the TW exercise treatment. The variability in response across sex, diet, and exercise treatment groups indicates that there are significant sex-by-diet-by-exercise interactions that affects the climbing ability of the line (p <0.0001).
Taken together, the results indicate that the impact of exercise on an adult fly's metabolic health can be a function of its sex, genotype, and larval diet. The phenotypic variation observed in response to genotype, environment variables (e.g., diet and exercise), and sex has also been observed in other studies11,12,14,15. Thus, the TW and Drosophila can be a powerful strategy to elucidate the genetic and environmental factors shaping metabolic health.
Figure 1: The TreadWheel exercise machine. (A) The machine holds 48 vials and has an adjustable speed feature. Here experiments were performed at 4 rpm. (B) Individual food vials containing the experimental flies were snapped into brackets attached to a rotating axle. (C) The distance between vial plug and food was 6 cm for exercise vials and 1 cm for control vials. The vials were then placed on the machine for exercise. (D) The 5 day inverse pyramid exercise regime was used to exercise adult flies. Each day, an additional five minutes was added to one of the exercise bouts to ramp up exercise intensity incrementally, which models endurance, interval training. This figure has been modified from Mendez et al.11. Please click here to view a larger version of this figure.
Figure 2: Visual methods and representative results. (A) Methods for testing larval diet and adult exercise interaction effects. Larvae were raised on high fat or normal laboratory diet and switched to normal food upon eclosion. Adult flies were separated by sex, placed into experimental groups and exercised for five consecutive days. Afterwards, a RING-like negative geotaxis climbing assay was conducted, and flies were frozen for triglyceride measurements. Representative triglyceride data are shown for two genetic lines, Oregon R and y1w1 for (B) females and (C) males. All flies were reared on a normal diet and exercised in adulthood. Levels with different letters are significantly different (p <0.05) using a post hoc Student's t-test. This figure contains a subset of data reported in Mendez et al.11. Representative climbing data are shown for line DGRP 153 for (D) females and (E) males. Each point represents the averaged climbing performance of 232 or more individuals across three independent time points. Error bars indicate one standard error. Please click here to view a larger version of this figure.
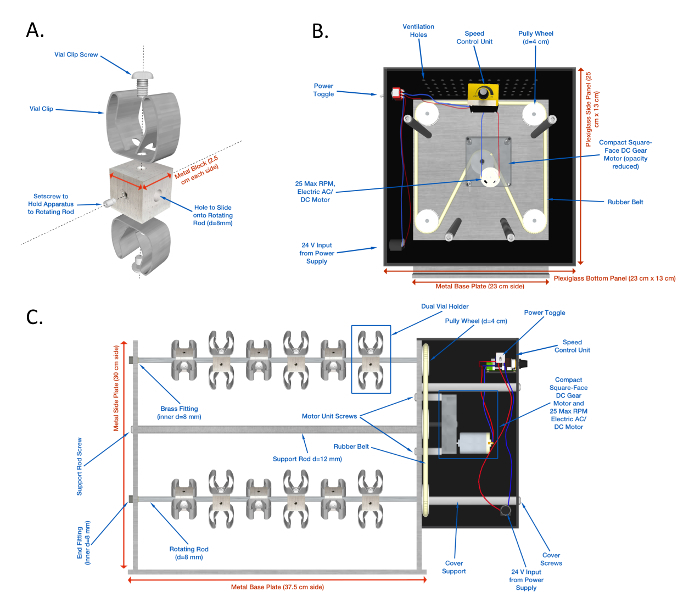 Supplemental Figure 1: A detailed schematic of the TreadWheel. (A) Diagram of the dual vial clamp structure used to attach vials to the machine. (B) Interior view of the machine's rotational drive system. (C) Parallel projection of the machine's front face. Please click here to view a larger version of this figure.
Supplemental Figure 1: A detailed schematic of the TreadWheel. (A) Diagram of the dual vial clamp structure used to attach vials to the machine. (B) Interior view of the machine's rotational drive system. (C) Parallel projection of the machine's front face. Please click here to view a larger version of this figure.
Supplemental Code File: TreadWheel.skp Please click here to download this file.
Discussion
The exercise protocol detailed here has been shown to successfully stimulate gentle exercise in Drosophila and can be used to simulate endurance exercise in a controlled laboratory setting11. It should be noted that when initially developing the TreadWheel concept, we considered commercial products that might be modified to perform similarly (e.g., a laboratory rotisserie). However, we ultimately rejected this approach, choosing a custom-built design because the commercial equipment did not have a sufficiently low rotational speed (4 rpm) and lacked sufficient vial capacity for high-throughput sample generation.
The TW protocol itself can be adjusted to encompass a wide range of research topics. For example, adjusting the frequency and duration of the flies' exercise regimes can alter the intensities of the work-outs they receive. By extending the protocol longer than one week or to other age groups, it would be possible to study the effects of exercise on aging and various age-related diseases. We recommend that if an extended exercise approach is implemented, then at least one rest day per week should be added to allow for recovery. Performance and improvement was shown to be greater in regimes that included a recovery day than those regimes that used consecutive daily exercise12. Also, we would caution against extending exercise bout lengths past 30 min since habituation to the rotational motion can occur after long periods of continuous exercise11. We have also observed genetic variations in flies' motivation to maintain their activity levels on the TW11. Thus, if longer bouts are desired, consider monitoring the flies' activity levels, as presented by Watanabe and Riddle15,21 describing their modification of the TW concept.Other factors that could be varied include temperature and sleep-wake patterns, which are shown to also affect fly movements26,27.
There is a plethora of available analyses during and after protocol completion to address the effects of exercise at the physiological, behavioral, and molecular level, as well as the potential to unravel epigenetic effects of exercise. Additional metabolic assays (e.g., glucose, glycogen, and protein), cardiac performance, and inflammatory response measures can be used to further explore the effect exercise has on fly physiology and body composition12,14,24,28. Various behavioral changes associated with exercise induction, such as changes in feeding behavior, locomotor activity, and sleep, can also be measured using tools like the CAFE assay29 or activity monitoring devices12,15,30. Changes in gene expression and cellular respiration attributed to exercise can also be quantified using methods like qRT-PCR11 and respirometry31. Finally, Drosophila have valuable genetic resources available, such as the Drosophila Genetics Reference Panel 2 and the Drosophila Synthetic Population Resource that provide researchers the platform to perform quantitative genetic studies25,32. These tools enable mapping experiments, such as Genome-Wide Association Studies and Quantitative Trait Loci mapping, to identify candidate loci associated with exercise and diet.
Studies completed using the TreadWheel demonstrated that, on average, exercise decreases body weight, total triglyceride storage, and glycogen, while increasing protein content and climbing performance11. Additionally, there were variable responses to exercise across sexes and genotypes for body weight, as well as triglycerides, protein, glycogen, glucose, and activity levels11,15. While the variability of response to exercise and diet across genotypes and sex may be challenging to interpret and, at times, counterintuitive, it reflects the sources of real biological variation observed in natural populations. As we strive to understand the diversity of underlying factors contributing to the high rates of MetS, tools that facilitate disentangling the relative role of various contributing factors by testing these factors in model organisms will be critical to our ability to develop personalized prevention and treatment interventions. To fully assess the effectiveness of exercise, each of these factors and how they interact must be considered when conducting experiments and formulating conclusions.
The TW, like most other fly exercise machines, is limited in its ability to quantify fly motion. Recently, Watanabe and Riddle developed the Rotating Exercise Quantification System (REQS), a TW backbone modified with an activity monitoring unit (LAM25H) from the Drosophila Monitoring System series15,21. Like the TW, this system uses rotary motion to gently induce exercise while using the LAM25H arm to track and quantify fly activity, but it holds a smaller number of vials (32 vials) than the TW15. For high-throughput exercise studies, if activity quantification is not necessary or desired, the TW allows for a greater sample number. The TW design could also be modified from its current form to accommodate a greater number of vials. This system, along with other existing exercise methods, helped to establish that exercise is not only achievable in Drosophila but can also be studied to determine the effect of physical activity on a variety of physiological and molecular responses11,12,15. Thus, the TW, as a proven method of inducing gentle exercise in flies, can be used to probe a large variety of biological questions.
Disclosures
The authors have nothing to disclose.
Acknowledgments
We would like to thank Julie Jarnigan, Meredith Owens, Rachel Hill, Brandon Moye, Laura Mafla, Olivia Fish, and the rest of the Reed Lab for their help with fly husbandry and image processing. Sean Mendez built the original TreadWheel with assistance from the UAB machine shop. Funding for this study was provided by NIH-R01 GM 098856 to LKR and the Undergraduate Creativity and Research Academy at the University of Alabama to KEL and BJW.
References
- Yanovski JA. Pediatric obesity. An introduction. Appetite. 2015;93:3–12. doi: 10.1016/j.appet.2015.03.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alberti KGMM, Zimmet P, Shaw J. Metabolic syndrome-a new world-wide definition. A Consensus Statement from the International Diabetes Federation. Diabetic Medicine. 2006;23:469–480. doi: 10.1111/j.1464-5491.2006.01858.x. [DOI] [PubMed] [Google Scholar]
- American Diabetes Association. 6. Obesity Management for the Treatment of Type 2 Diabetes. Diabetes Care. 2016;39:47–51. doi: 10.2337/dc16-S009. Suppl 1. [DOI] [PubMed] [Google Scholar]
- Bliddal H, Leeds AR, Christensen R. Osteoarthritis, obesity and weight loss: evidence, hypotheses and horizons - a scoping review. Obesity Reviews. 2014;15:578–586. doi: 10.1111/obr.12173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bray GA. The epidemic of obesity and changes in food intake: the Fluoride Hypothesis. Physiol. Behav. 2004;82:115–121. doi: 10.1016/j.physbeh.2004.04.033. [DOI] [PubMed] [Google Scholar]
- O'Rahilly S, Farooqi IS. Genetics of obesity. Philosophical Transactions of the Royal Society B: Biological Sciences. 2006;361:1095–1105. doi: 10.1098/rstb.2006.1850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kenkre J, Tan T, Bloom S. Treating the obese diabetic. Expert Rev Clin Pharmacol. 2013;6:171–183. doi: 10.1586/ecp.13.5. [DOI] [PubMed] [Google Scholar]
- Fiuza-Luces C, Garatachea N, Berger NA, Lucia A. Exercise is the Real Polypill. Physiology. 2013;28:330–358. doi: 10.1152/physiol.00019.2013. [DOI] [PubMed] [Google Scholar]
- Matos A, et al. Acute exercise reverses TRB3 expression in the skeletal muscle and ameliorates whole body insulin sensitivity in diabetic mice. Acta Physiologica. 2010;198:61–69. doi: 10.1111/j.1748-1716.2009.02031.x. [DOI] [PubMed] [Google Scholar]
- Mercken EM, Carboneau BA, Krzysik-Walker SM, de Cabo R. Of mice and men: The benefits of caloric restriction, exercise, and mimetics. Ageing Research Reviews. 2012;11:390–398. doi: 10.1016/j.arr.2011.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendez S, et al. The TreadWheel: A Novel Apparatus to Measure Genetic Variation in Response to Gently Induced Exercise for Drosophila. PLoS ONE. 2016;11:0164706. doi: 10.1371/journal.pone.0164706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piazza N, Gosangi B, Devilla S, Arking R, Wessells R. Exercise-Training in Young Drosophila melanogaster Reduces Age-Related Decline in Mobility and Cardiac Performance. PLoS ONE. 2009;4:5886–5911. doi: 10.1371/journal.pone.0005886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham P, Pick L. Drosophila as a Model for Diabetes and Diseases of Insulin Resistance. Fly Models of Human Diseases. 2017;121:397–419. doi: 10.1016/bs.ctdb.2016.07.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reed LK, et al. Genotype-by-Diet Interactions Drive Metabolic Phenotype Variation in Drosophila melanogaster. Genetics. 2010;185:1009–1019. doi: 10.1534/genetics.109.113571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watanabe LP, Riddle NC. Characterization of the Rotating Exercise Quantification System (REQS), a novel Drosophila exercise quantification apparatus. PLoS ONE. 2017;12:0185090. doi: 10.1371/journal.pone.0185090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tinkerhess MJ, Ginzberg S, Piazza N, Wessells RJ. Endurance training protocol and longitudinal performance assays for Drosophila melanogaster. J Vis Exp. 2012. [DOI] [PMC free article] [PubMed]
- Tinkerhess MJ, et al. The Drosophila PGC-1α Homolog spargel Modulates the Physiological Effects of Endurance Exercise. PLoS ONE. 2012;7:31633. doi: 10.1371/journal.pone.0031633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sujkowski A, Bazzell B, Carpenter K, Arking R, Wessells RJ. Endurance exercise and selective breeding for longevity extend Drosophila healthspan by overlapping mechanisms. Aging. 2015;7:535–552. doi: 10.18632/aging.100789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tjønna AE, et al. Aerobic Interval Training Versus Continuous Moderate Exercise as a Treatment for the Metabolic Syndrome: A Pilot Study. Circulation. 2008;118:346–354. doi: 10.1161/CIRCULATIONAHA.108.772822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ciolac EG, et al. Effects of high-intensity aerobic interval training vs. moderate exercise on hemodynamic, metabolic and neuro-humoral abnormalities of young normotensive women at high familial risk for hypertension. Hypertension Research 2010 33:8. 2010;33:836–843. doi: 10.1038/hr.2010.72. [DOI] [PubMed] [Google Scholar]
- Watanabe LP, Riddle NC. Measuring exercise levels in Drosophila melanogaster using the Rotating Exercise Quantification System (REQS) J Vis Exp. 2018. [DOI] [PMC free article] [PubMed]
- Gargano J, Martin I, Bhandari P, Grotewiel M. Rapid iterative negative geotaxis (RING): a new method for assessing age-related locomotor decline in Drosophila. Experimental Gerontology. 2005;40:386–395. doi: 10.1016/j.exger.2005.02.005. [DOI] [PubMed] [Google Scholar]
- Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nature Methods 2012 9:7. 2012;9:671–675. doi: 10.1038/nmeth.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammond JBW, Kruger NJ. New Protein Techniques. 1988;3:25–32. [Google Scholar]
- Mackay TFC, et al. The Drosophila melanogaster Genetic Reference Panel. Nature. 2012;482:173–178. doi: 10.1038/nature10811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miquel J, Lundgren PR, Bensch KG, Atlan H. Effects of temperature on the life span, vitality and fine structure of Drosophila melanogaster. Mechanisms of Ageing and Development. 1975;5:347–370. doi: 10.1016/0047-6374(76)90034-8. [DOI] [PubMed] [Google Scholar]
- Berlandi J, et al. Swing Boat: Inducing and Recording Locomotor Activity in a Drosophila melanogaster Model of Alzheimer's Disease. Front Behav Neurosci. 2017;11:159. doi: 10.3389/fnbeh.2017.00159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leng SX, et al. ELISA and Multiplex Technologies for Cytokine Measurement in Inflammation and Aging Research. The Journals of Gerontology Series A: Biological Sciences and Medical Sciences. 2008;63:879–884. doi: 10.1093/gerona/63.8.879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ja WW, et al. Prandiology of Drosophila and the CAFE assay. Proceedings of the National Academy of Sciences. 2007;104:8253–8256. doi: 10.1073/pnas.0702726104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu JC, Low KH, Pike DH, Yildirim E, Edery I. Assaying locomotor activity to study circadian rhythms and sleep parameters in Drosophila. J Vis Exp. 2010. [DOI] [PMC free article] [PubMed]
- MÖlich AB, FÖrster TD, Lighton JRB. Hyperthermic Overdrive: Oxygen Delivery does Not Limit Thermal Tolerance in Drosophila melanogaster. Journal of Insect Science. 2012;12:1–7. doi: 10.1673/031.012.10901. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Long AD, Macdonald SJ, King EG. Dissecting complex traits using the Drosophila Synthetic Population Resource. Trends in Genetics. 2014;30:488–495. doi: 10.1016/j.tig.2014.07.009. [DOI] [PMC free article] [PubMed] [Google Scholar]