

Abstract
Cocaine use disorder (CUD) follows a trajectory of repetitive self-administration during which previously neutral stimuli gain incentive value. Cue reactivity, the sensitivity to cues previously linked with the drug-taking experience, plays a prominent role in human craving during abstinence. Cue reactivity can be assessed as the attentional orientation toward drug-associated cues that is measurable as appetitive approach behavior in both preclinical and human studies. Herein describes an assessment of cue reactivity in rats trained to self-administer cocaine. Cocaine self-administration is paired with the presentation of discrete cues that act as conditioned reinforcers (i.e., house light, stimulus light, infusion pump sounds). Following a period of abstinence, lever presses in the cocaine self-administration context accompanied by the discrete cues previously paired with cocaine infusion are measured as cue reactivity. This model is useful to explore neurobiological mechanisms underlying cue reactivity processes as well as to assess pharmacotherapies to suppress cue reactivity and therefore, modify relapse vulnerability. Advantages of the model include its translational relevance, and its face and predictive validities. The primary limitation of the model is that the cue reactivity task can only be performed infrequently and must only be used in short duration (e.g., 1 hour), otherwise rats will begin to extinguish the pairing of the discrete cues with the cocaine stimulus. The model is extendable to any positively reinforcing stimulus paired with discrete cues; though particularly applicable to drugs of abuse, this model may hold future applications in fields such as obesity, where palatable food rewards can act as positively reinforcing stimuli.
Keywords: Neuroscience, Issue 136, Cocaine, Substance use disorder, Self-administration, Cue reactivity, Rats, Abstinence, Cue, Relapse, Operant conditioning, Drug, Drug-seeking, Translational
Introduction
Cocaine use disorder (CUD) follows a trajectory of repetitive self-administration during which previously neutral stimuli gain incentive value1. Cue reactivity is the sensitivity to cues previously linked with the drug-taking experience, and it plays a prominent role in human craving2,3,4,5. The risk of progression to CUD, as well as relapse during abstinence, is thought to be higher for individuals who express high sensitivity to drug-associated cues6,7. Both environmental contexts (e.g., people, buildings, music genres) and discrete drug-associated stimuli (e.g., paraphernalia) become associated with the cocaine reward; exposure to these cues can trigger changes in peripheral physiology (e.g., heart rate, skin temperature, and skin resistance), brain plasticity, and brain functional connectivity2,8,9,10. In other words, re-exposure to cocaine-associated cues activates limbic corticostriatal circuits to evoke conditioned physiological and subjective responses that drive appetitive approach (drug-seeking) behavior11,12,13,14,15.
Cue reactivity measured with functional brain imaging analyses is predictive of relapse vulnerability in subjects with CUD16. Cue reactivity measurements in rodent models serve as a surrogate measure for relapse risk and can be exploited for translational studies. Thus, a pharmacotherapy that decreases cue reactivity in rodents may be carried forward as a relapse-prevention treatment in human clinical trials. Preclinical models with the necessary translational merit and predictive validity are especially important since there are currently no FDA-approved pharmacotherapies for CUD17.
The rodent self-administration procedure is the gold standard, translational model with predictive validity for human drug-taking18 and critically important to understanding the molecular and physiological processes underlying CUD. Response-independent delivery of cocaine results in distinct behavioral, molecular, and neurochemical effects relative to response-dependent cocaine exposure; e.g., response-independent cocaine delivery evokes significantly higher mortality19. Furthermore, the neurochemical consequences of abstinence from response-dependent cocaine self-administration are distinct from those triggered by abstinence from response-independent cocaine delivery20,21. Thus, CUD models based upon response-dependent delivery of cocaine are superior translational models when assessing cue reactivity and associated mechanisms of action.
In the protocol outlined below, cocaine is delivered intravenously through an indwelling intra-jugular catheter. However, alternative methods to self-administer drug via oral and inhalation routes have been developed. Importantly, rodents control delivery of the drug, analogous humans, through operant responses. Therefore, there is high concordance between drugs self-administered by rodents and humans22. The preclinical drug self-administration procedure below employs lever pressing, reinforced by drug delivery, to motivate response rates higher than vehicle control. Drug-seeking behavior is trained by pairing originally "neutral" cues (e.g., a stimulus light or tone and the contextual environment in which cocaine self-administration occurs) with cocaine infusion; these cues become conditioned reinforcers (for review: Cunningham & Anastasio, 201423). Subsequent re-exposure to cocaine-associated cues triggers drug-seeking behavior in rodents (i.e., attempts to deliver cocaine through pressing on the previously-active lever) as well as craving and relapse in CUD subjects24,25,26,27.
Typically, preclinical rodent studies of drug-seeking behavior following cocaine self-administration utilize extinction training and/or drug reinstatement conducted within the drug-associated environment28,29,30,31,32. Presses on the previously-active lever, in the absence of drug and/or cue delivery, typically constitute the measure of reinstatement following extinction33,34,35. On the contrary, cue reactivity drug-seeking behavior is assessed following forced abstinence without prior extinction training28,36,37,38,39.
Outcome measures and experimental variables have been carefully chosen and validated to dissect different aspects of the neurobiology of drug-seeking and relapse-like behavior, and it is well-established that neuroadaptations differ between models with and without extinction training 40,41,42,43. Furthermore, from a translational perspective, rodent extinction training is not mirrored in clinical settings for CUD since drug-related cues include mood states, places, and people44; the unique combination of these cues are likely not available in a clinical environment45,46,47. Thus, the rodent model described herein acts as a better parallel to the human condition than many of the models currently available.
The following describes a validated cocaine self-administration training, forced abstinence and cue reactivity test protocol for rats. Briefly, rats are implanted with intra-jugular catheters, trained to self-administer cocaine or saline via 'active' lever press, and receipt of the cocaine or saline stimulus is paired with discrete light and sound cues which serve as conditioned reinforcers. Following 14 days of cocaine self-administration, rats are subjected to 30 days of forced abstinence and a subsequent 60-min cue reactivity test in which lever pressing is measured. The cue reactivity test is a surrogate measure for cocaine relapse vulnerability in humans.
Protocol
All animal manipulations are carried out in accordance with the Guide for the Care and Use of Laboratory Animals (2011) and with approval from the Institutional Animal Care and Use Committee.
1. Animals
Acclimate male Sprague Dawley rats approximately 8-9 weeks of age (250-260 g) for a minimum of seven days in a colony room maintained at 21-23 °C and 45-50% humidity on a 12 h light-dark cycle (lights on 6:00-18:00 h).
House rats two/cage and handle daily throughout the study. Provide food and water ad libitum in the home cage throughout all phases of the study. NOTE: Food restriction increases the saliency of the cocaine reward48, potentiates cocaine-induced locomotor activity49, and acts as a stressor for reinstatement50 and thus may also impact cue reactivity.
Randomly assign rats to saline or cocaine treatment groups.
2. Surgery
- Place rat under anesthesia using a cocktail containing 8.6 mg/kg of xylazine, 1.5 mg/kg of acepromazine, and 43 mg/kg of ketamine suspended in bacteriostatic saline. Confirm that the rat is sufficiently anesthetized by pinching its toe and monitoring motor reflexes and respiration. Apply ophthalmic ointment directly to eyes to prevent eye dryness. NOTE: If the rat is not sufficiently anesthetized it will exhibit a motor response to the toe pinch, and the rat's breathing rate may rapidly increase.
- Using aseptic technique while the rat is under anesthesia, implant an intravenous catheter into the jugular vein connected to a back mount with a cannula. After surgical implantation, allow rats to recover for 5-7 days 28,36,37,51. NOTE: The intravenous route of cocaine self-administration in humans results in rapid increases in brain cocaine levels52,53. Thus, in one of its most common preclinical variants, rats are trained to self-administer cocaine via an intra-jugular catheter.
- Maintain catheter patency with daily flushes using a solution of 0.1 mL of bacteriostatic saline containing heparin sodium (10 U/mL), streptokinase (0.67 mg/mL), and ticarcillin disodium (66.67 mg/mL). Begin catheter flushing the day after surgery and continue each day following self-administration sessions.
- Fill a syringe with the solution and connect the back mount to the syringe via polyethylene tubing to flush.
- Verify proper catheter function periodically throughout the experiment by intravenous administration of 10 mg/kg of methohexital sodium. NOTE: This is a dose sufficient to briefly anesthetize the animal only when administered intravenously.
3. Self-Administration
- Use software programming to generate 4 separate programs as described below.
- Code two programs for self-administration on a fixed ratio (FR) 1 schedule, i.e., where one active lever press results in one infusion. For program #1, code the left lever as the active lever; for program #2, code the right lever as the active lever. Construct both programs to be 180-min in length, and deliver the cocaine stimulus and discrete cue complex in the following manner:
- Have one active lever press cause simultaneous illumination of both the house light and the stimulus light above the active lever.
- After a 1-s delay, have the infusion pump deliver 0.1 mL of solution over 6-s.
- Have the stimulus light shut off after 7-s (inactivating at the same time as the infusion pump).
- Have the house light shut off after 27-s, with the last 20-s indicating a time-out period during which active lever presses have no scheduled consequences, yet these are still recorded.
- For data output, collect infusions, active lever presses, inactive lever presses, and latency to first active lever press.
- Code two more programs for self-administration on a FR5 schedule, i.e., where five lever presses result in one infusion. Again, make one program with the active lever as left, and one program with the active lever as right. Make both programs 180-min in length, and deliver the reinforcer and discrete cue complex in the following manner:
- Have five active lever presses cause simultaneous illumination of both the house light and the stimulus light above the active lever.
- Repeat Steps 3.1.1.2 through 3.1.1.5.
- Self-Administration Training
- Prepare the cocaine solution daily in sterile 0.9% NaCl to a dose of 0.75 mg/kg/0.1 mL. NOTE: Over the duration of each 180 min self-administration session, rats administer approximately 4-5 mL of cocaine solution, thus to prepare in excess, anticipate approximately 8 mL/rat daily.
- Fill the catheter syringe (10 mL) with at least 8 mL of cocaine or saline solution. After ensuring that the cocaine/saline solution is flowing, insert the syringes into the infusion pump and connect to the polyethylene tubing encased by a metal spring leash at the start of each self-administration session. Carefully adjust the syringe in each infusion pump such that the solution is evenly and completed distributed in the polyethylene tubing prior to attaching to each catheter.
- Place rats into respective standard operant conditioning chambers housed in ventilated, sound-attenuating cubicles with fans. Consistently place each rat in the same operant chamber, and keep the active lever on the same side throughout all experiments. Counterbalance assignment of rats between left and right active levers. NOTE: Each chamber is equipped with a pellet receptacle flanked by two retractable response levers, a stimulus light above each response lever, and a house light opposite the levers. Cocaine/saline syringes are connected to a 23g needle (filed to avoid poking tubing) attached to polyethylene tubing encased inside a metal spring leash which connects to each rat's catheter and is operated on a liquid swivel. Syringes are placed into infusion pumps located adjacent to the cubicle.
- Train rats to lever press for cocaine infusions (0.75 mg/kg/0.1 mL infusion) or saline infusions (0.1 mL) during daily 180-min sessions using established methods28,36,37,51. NOTE: This training is performed simply by placing the rat into the operant chamber and allowing it to lever press; pressing the active lever will result in receipt of the discrete cue complex and cocaine infusion, while pressing the inactive lever bears no scheduled consequences.
- Schedule completions on the active lever result in delivery of a cocaine or saline infusion over a 6-s period paired simultaneously with illumination of the house light and stimulus light above the active lever and activation of the infusion pump (this is the discrete cue complex paired with cocaine delivery); inactive lever presses produce no scheduled consequences. Following reinforcer (cocaine) delivery, the stimulus light as well as the infusion pump are inactivated; the house light remains on for an additional 20-s to indicate a timeout period during which lever presses have no scheduled consequences. NOTE: A description of how to set this up is found in Steps 3.1.1.1-3.1.1.4
- Upon completion of the 180-min session, remove each rat from its operant conditioning chamber and flush its catheter (Step 2.2). Carefully clean the chambers between sessions, wiping each surface with a 70% ethanol solution. NOTE: Rats that receive cocaine during the self-administration session may be difficult to remove from the chamber and are more likely to be aggressive. Use slow, careful motions to disconnect each rat from its tether.
- Train rats on a FR1 schedule of reinforcement and progress to a FR5 schedule after achieving a criterion of seven infusions/h with less than 10% variability in the total number of infusions per session for three consecutive days. Continue self-administration sessions until rats have reached a total of 14 days of self-administration training. NOTE: This is one variant of operant learning; some protocols prefer to have rats remain at FR1 throughout the entirety of the experiment. Changes in fixed ratio schedule from FR1 to FR5 do not impact the number of infusions an animal self-administers54,55. However, increasing the fixed ratio size helps to ensure stability of responding in the rat, and may increase the saliency of the reward56. Additionally, if a treatment during forced abstinence is desired, rats should be pseudo-randomly assigned to treatment groups upon achieving stability (less than 10% variability in the total number of infusions per session for a minimum of 3 consecutive days) at the FR5 schedule. Additionally, saline rats will not progress from FR1 to FR5, as the saliency of saline is not sufficient to achieve the stability required to progress from FR1 to FR5.
4. Forced Abstinence
Place rats into forced abstinence for 30 days; that is, do not provide rats with the opportunity to self-administer cocaine or saline for a duration of 30 days. Importantly, during this period do not return rats to the operant chambers (the previous context of self-administration). However, remove rats from home cages for daily handling and weighing, as a measure of general health. NOTE: The duration of forced abstinence can be altered, but it is important to choose a duration that is sufficient to avoid ceiling and floor effects. Factors to be cognizant of when choosing the duration of the forced abstinence period include rodent strain, age, self-administration session length and duration, drug, drug dose and infusion duration, and the sex of the animal.
5. Cocaine Cue Reactivity
- Code two software programs for the cue reactivity task at a FR1 schedule; one program with the left lever as the active lever, and one with the right lever as the active lever. Construct each program to collect lever pressing data for 60-min and deliver the discrete cue complex as described below.
- Repeat Steps 3.1.1.1-3.1.1.4
- For data output, collect cue presentations (previously known as infusions), previously-active lever presses, inactive lever presses, and latency to first lever press.
- Cocaine Cue Reactivity Test Session
- Assess cue reactivity on day 30 of forced abstinence in a 60-min operant test session. Stagger start times of rats to facilitate rapid decapitation and brain harvest. Importantly, for the cue reactivity test, do not place the syringe in the infusion pump so that the sound of the infusion pump serves as an auditory cue but no cocaine reinforcement is delivered. NOTE: Cue reactivity can be assessed on days other than day 30 of forced abstinence, however the fact that responsiveness to cocaine-paired cues varies over time34 should be kept in mind when choosing the date for the cue reactivity test.
- Place each rat in the self-administration chamber in which it was trained and tether to its cannula as during the daily self-administration sessions. Presses on the lever that previously delivered cocaine or saline are now only reinforced by delivery of the discrete cue complex (i.e., the stimulus light and house are illuminated, and the infusion pump is activated) on a FR1 schedule (i.e., one discrete cue complex is presented per active lever press); count the cue presentations and previously-active lever presses during one 60-min session, and also record the number of inactive lever presses. However, as with self-administration, inactive lever presses produce no scheduled consequences. NOTE: An additional control option is to record previously-active and inactive lever presses in the absence of scheduled consequences (i.e., no discrete cue complex reinforcer).
- Upon completion of the cue reactivity test session, immediately euthanize rats to capture the biochemical effects of re-exposure to the operant conditioning chamber and discrete cue complex.
6. Data Collection, Organization, and Analysis
- Break down data into three conventional categories: general health, self-administration, and cue reactivity.
- Monitor general health (e.g., animal weight, fur condition) throughout the experiment, and provide animals with veterinary care, if necessary, or remove from the study if lever pressing appears affected by health.
- Collect self-administration data during the 14 days of self-administration. Use an unpaired, two-tailed t-test to determine statistical differences between cocaine and saline self-administration groups for total number of infusions, active lever presses, inactive lever presses, and latency to first active lever press. NOTE: Self-administration data needs to be analyzed prior to forced abstinence if one desires to study the effect of a treatment during forced abstinence on cue reactivity (so that groups can have an equal average number of infusions, as well as active and inactive lever presses). If a two-by-two design is chosen (cocaine ± treatment, saline ± treatment), a two-way ANOVA with appropriate post hoc analysis should be used to analyze this data.
- For cue reactivity data to collect and analyze for the total number of cue presentations, previously-active lever presses, inactive lever presses, and latency to first lever press. Also analyze this data with an unpaired, two-tailed t-test to determine differences between animals self-administering cocaine and those self-administering saline. NOTE: Again, if a two-by-two design is chosen (cocaine ± treatment, saline ± treatment), a two-way ANOVA with appropriate post hoc analysis should be used to analyze this data.
Representative Results
Results of a cocaine self-administration and abstinence experiment followed by a cue reactivity test from a previously published study57 are shown in Figure 1. The study timeline is depicted in Figure 1A.
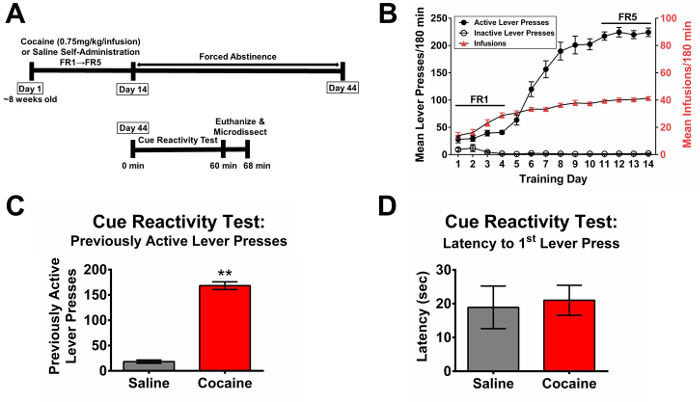
Rats individually transition from FR1 to FR5 as they meet criteria. As operant conditioning proceeds in the cocaine-administering group, rats gradually increase the number of infusions until they reach a plateau (Figure 1B). Inactive lever presses remain consistently low in both the cocaine-administering and saline-administering groups; differences in inactive lever presses suggest changes in motivation or health. Saline self-administration serves as a control in that saline is not reinforcing; thus, lever presses and infusions do not escalate from day 1 to day 14 (not pictured). If active lever presses in the cocaine-administering group decline between day 1 and day 14, the catheter has likely lost patency (also evidenced by the method described in Step 2.2.2), and this animal should be removed from the study. Animals that are statistical outliers (as calculated using a Dixon's Q Test) should also be removed from the study.
Multiple output measures are gathered from the cocaine cue reactivity test. For example, Figure 1C depicts the number of lever presses on the previously-active lever. Rats which previously self-administered cocaine should press the previously-active lever much more than rats which self-administered saline. And, if a treatment is effective at attenuating cocaine cue reactivity, there should also be a significant reduction in the number of previously-active lever presses in rats which received cocaine and the treatment compared to rats which received cocaine and the vehicle (not pictured).
Other measurements include latency to first lever press (Figure 1D), number of inactive lever presses, and number of cue presentations. Changes in latency to first lever press may be observed; an increased latency to first lever press may suggest that the animal is physically ill or that there is decreased motivation for cocaine. Appropriate controls for locomotor behavior, such as the Rotarod or Open Field tasks, should be employed to differentiate between physical illness and changes in motivation.
Although one might presume that the number of cue presentation should be reflected in the number of previously active lever presses, this is not often the case. Oftentimes previously-active lever presses are much greater than cue presentations. Rats often show perseverative previously-active lever pressing during the timeout period, ostensibly in response to a failure to receive the expected cocaine stimulus.
Figure 1. Experimental timeline and representative self-administration and cue reactivity results. This figure has been modified from Miller et al., 201657. (A) Experimental timeline as described in Protocol sections 3,4,5. (B) Average daily active lever presses, inactive lever presses, and infusions ± SEM for self-administration training; self-administration sessions were 180-min each day for 14 consecutive days. Cocaine was initially delivered on a FR1 schedule of reinforcement until meeting criterion, then a FR5 schedule was introduced. (C) Average lever presses on the previously-active lever press ± SEM for the 60-min cue reactivity test; an unpaired, two-tailed t-test shows a significant difference between cocaine and saline self-administering rats (t(30) = 14.82, **p <0.0001). (D) Average latency to first lever press ± SEM for the 60-min cue reactivity test; an unpaired, two-tailed t-test shows no significant difference between cocaine and saline self-administering rats (t(29) = 0.2758, p = 0.7847). Please click here to view a larger version of this figure.
Discussion
Exposure to drug-paired cues and physiological changes in response to these cues16 are associated with relapse,11,16 and the cocaine cue reactivity test employed above contingently presents cocaine-paired cues in the absence of drug; thus, drug-seeking behavior in the form of previously-active lever presses serves as a measure of relapse vulnerability. The cue reactivity protocol described herein is a preclinical means by which relapse-modifying pharmacotherapies can be tested, and genetic and environmental factors that contribute to relapse risk can be investigated. This is an operant task for drug-seeking behavior that aligns with many properties of human drug-taking and relapse, and many publications have optimized key drug dose and delivery parameters as well as adopted appropriate controls to establish face validity and translational predictive power28,36,37,39,51. This model has recently been published in a study by Miller et al. investigating the use of a peroxisome proliferator-activated receptor gamma (PPARγ agonist as a pharmacotherapeutic to attenuate cocaine cue reactivity57. This study showed that pioglitazone (a PPARγ agonist) decreased responsiveness to cocaine-paired cues following forced abstinence from cocaine self-administration. Since pioglitazone is FDA-approved, the Miller et al. study prompted a two-arm, double-blind, randomized, placebo-controlled pilot clinical trial to examine the effectiveness of PPARγ agonism in suppressing cocaine cue reactivity, impulsivity, and risky decision-making. The translational merit and predictive validity of the rodent model is supported by the Schmitz et al. publication58 on this pilot study in which cocaine-dependent subjects treated with pioglitazone exhibited a treatment x time interaction for reduced self-reported craving as measured by the Brief Substance Craving Scale and a Visual Analog Scale for craving58.Thus, pioglitazone shows promise as a relapse prevention treatment for CUD, consistent with the Miller et al. preclinical data.
This rat cocaine cue reactivity test during abstinence recapitulates chronic human cocaine self-administration with intermittent short (e.g., 21 hour) and long (e.g., 30 day) abstinence periods, providing the ability to create parallel datasets between humans and rodent models. However, this protocol differs from the more pervasive extinction-reinstatement model which measures time to drug-taking or -seeking following an extinction process and (typically) an abstinence period33,34,35,59. Though the extinction-reinstatement model holds predictive validity33, it also has noteworthy limitations. First, the extinction process proceeds with re-exposing saline and cocaine self-administering groups to the self-administration context in the absence of drug reinforcement60 which is rarely recapitulated in humans since drug-related cues include mood states, places, and people44—cues which cannot be easily copied in a clinical setting45,46,47. Second, during the reinstatement process, rats exhibit low levels of responding creating a floor effect that suppresses one's ability to detect effects of interventions to reduce drug-seeking behavior33—an issue that is resolved by removing extinction since extinction training decreases responding. Some may argue that the 60-min cue reactivity test session described above could serve as an extinction session. However, the fact that the protocol includes non-drug reinforcement, the test session is not repeated, and rats maintain motivation for subsequent self-administration after cue reactivity testing suggests that this test design does not induce extinction36.
An additional issue is whether the relatively short self-administration session length employed (180-min) is sufficient to reinforce subsequent self-administration. However, literature shows that this regimen consistently results in a stable pattern of rat cocaine self-administration37,39,51,57,61, providing less variability between animals which allows the experimenter to sort subjects into post-self-administration treatment groups, if desired28,39,51,57. However, it should be noted that the short duration and short session length procedure does not entirely recapitulate human self-administration, where typically binge-like patterns emerge in cocaine-dependent subjects62. Nonetheless, if continuous self-administration is allowed in rodent models, mortality rates are extremely high63.
Additional variables to consider are cocaine dose and infusion rate since these influence sensitization of the subjective reward response. Faster speeds of infusion increase sensitization to the reinforcing effects of cocaine, while slower speeds decrease it64. Since the dose-dependence curve for cocaine self-administration is an inverted U shape and dose influences the number of infusions per hour55; the protocol above uses a dose which falls near the peak of the dose-response curve in both short-term access and long-term access procedures61 so as to detect shifts in the number of infusions. Counterbalancing the levers is another important control to ensure that the rats do not show preference for one lever over the other.
The representative results of Figure 1 show data from male, Sprague Dawley rats. Historically, these studies have been restricted to male rats because of the estrous cycle's effects on acquisition of cocaine self-administration65 and extinction/reinstatement66. Since sex as a biological variable is now mandated for NIH-funded studies, it has become imperative to monitor estrous stage and test for sex differences in mixed sex studies and, as such, statistical power must be considered for all experimental groups. On another note, use of an outbred rat strain, such as Sprague Dawley, is beneficial in that it provides genetic variation more comparable to the human population—though even outbred rodent strains may not fully recapitulate the diverse genetic influences thought to underlie addictive-like behaviors in humans67. Finally, age of initial cocaine exposure is an important consideration. In rats, exposure to cocaine during adolescence results in a conditioned place preference that requires a greater number of extinction trials to extinguish68, suggesting that the adolescent rat demonstrates greater persistence of cocaine-seeking behavior. Thus, prenatal and early postnatal exposure to cocaine will likely alter cocaine cue reactivity.
Because extinction is not the goal of cue reactivity testing, one limitation is that the task cannot be assessed repeatedly or over long durations, or else the animal will extinguish the pairing of the cocaine (primary) reinforcer with the discrete cue complex (conditioned reinforcers). Thus, care must be taken when designing the experiment to ensure that this extinction does not occur. However, an advantage of this task is that the precise biochemical changes induced by exposure to cocaine-paired cues can be elucidated using this procedure. A second limitation to the task is that it only works for positive reinforcers, though this is a limitation common to many models for drugs of abuse. Critical steps in the protocol include maintenance of catheter patency (Step 2.2), acquisition of self-administration (Step 3.2), appropriate distribution of rats into treatment groups, if desired, that are statistically equivalent for infusions and number of lever presses (Step 3.2.6), and proper software programming throughout to ensure that discrete cues are presented and paired with the cocaine stimulus during self-administration training and the cue reactivity test (Steps 3.1, 5.1). As with other relapse vulnerability models, it is imperative to achieve a reasonable length of abstinence to avoid floor and ceiling effects on responding during the cue reactivity test33,34 (Step 4). The most likely complications to encounter with this procedure are 1) loss of catheter patency preventing acquisition of self-administration, 2) improper distribution of rodents into treatment groups following self-administration training and prior to cue reactivity testing causing a Type 1 or Type 2 error, and 3) insufficient abstinence to determine differences in responding between groups. These can be resolved by daily assessment of self-administration performance, regular catheter flushes, assignment to post hoc analysis groups after balancing for self-administration performance, and pilot trials determining appropriate abstinence duration for rodent strain, age, self-administration session length and duration, drug, dose of drug, drug infusion duration, and sex.
This procedure is extendable to many rodent strains, including mice69, and any model which involves pairing of a primary rewarding reinforcer with discrete cues (conditioned reinforcers). Alternative primary reinforcers include other drug classes, high fat food, and other palatable rewards69,70. Finally, translational relevance of this model has been demonstrated by taking forward a FDA-approved intervention tested in rats to CUD human subjects which resulted in reduced cocaine craving. Thus, the cocaine cue reactivity test following cocaine self-administration and abstinence holds promise as a translational rodent model.
Disclosures
The authors have no conflicts of interest to disclose.
Acknowledgments
All behavioral testing was performed in the University of Texas Medical Branch (UTMB) Rodent In Vivo Assessment (RIVA) Core, directed by Dr. Kelly Dineley and housed within the Center for Addiction Research, directed by Dr. Kathryn Cunningham. Support for this work came from the Peter F. McManus Charitable Trust, National Institute of Environmental Health Sciences Center for Environmental Toxicology at UTMB (T32ES007254), Institute for Translational Sciences at UTMB (UL1TR001439), Mitchell Center for Neurodegenerative Diseases, and Center for Addiction Research at UTMB (DA007287, DA070087, and pilot study funds).
References
- Koob GF, Volkow ND. Neurobiology of addiction: a neurocircuitry analysis. Lancet Psychiatry. 2016;3(8):760–773. doi: 10.1016/S2215-0366(16)00104-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carter BL, Tiffany ST. Meta-analysis of cue-reactivity in addiction research. Addiction. 1999;94(3):327–340. [PubMed] [Google Scholar]
- Drummond DC. Theories of drug craving, ancient and modern. Addiction. 2001;96(1):33–46. doi: 10.1046/j.1360-0443.2001.961333.x. [DOI] [PubMed] [Google Scholar]
- Mahler SV, de Wit H. Cue-reactors: individual differences in cue-induced craving after food or smoking abstinence. PLoS One. 2010;5(11) doi: 10.1371/journal.pone.0015475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Brien CP, Childress AR, Ehrman R, Robbins SJ. Conditioning factors in drug abuse: can they explain compulsion? J Psychopharmacol. 1998;12(1):15–22. doi: 10.1177/026988119801200103. [DOI] [PubMed] [Google Scholar]
- Hendershot CS, Witkiewitz K, George WH, Marlatt GA. Relapse prevention for addictive behaviors. Subst Abuse Treat Prev Policy. 2011;6:17. doi: 10.1186/1747-597X-6-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Prisciandaro JJ, Myrick H, Henderson S, McRae-Clark AL, Brady KT. Prospective associations between brain activation to cocaine and no-go cues and cocaine relapse. Drug Alcohol Depend. 2013;131(1-2):44–49. doi: 10.1016/j.drugalcdep.2013.04.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Foltin RW, Haney M. Conditioned effects of environmental stimuli paired with smoked cocaine in humans. Psychopharmacology. 2000;149(1):24–33. doi: 10.1007/s002139900340. [DOI] [PubMed] [Google Scholar]
- Wang X, et al. Nucleus Accumbens Core Mammalian Target of Rapamycin Signaling Pathway Is Critical for Cue-Induced Reinstatement of Cocaine Seeking in Rats. J Neurosci. 2010;30(38):12632–12641. doi: 10.1523/JNEUROSCI.1264-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gipson CD, Kupchik YM, Shen H, Reissner KJ, Thomas CA, Kalivas PW. Relapse Induced by Cues Predicting Cocaine Depends on Rapid, Transient Synaptic Potentiation. Neuron. 2013;77(5):867–872. doi: 10.1016/j.neuron.2013.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wallace BC. Psychological and environmental determinants of relapse in crack cocaine smokers. J Subst Abuse Treat. 1989;6(2):95–106. doi: 10.1016/0740-5472(89)90036-6. [DOI] [PubMed] [Google Scholar]
- Weiss F, et al. Compulsive Drug-Seeking Behavior and Relapse: Neuroadaptation, Stress, and Conditioning Factors. Annals of the New York Academy of Sciences. 2001;937(1):1–26. doi: 10.1111/j.1749-6632.2001.tb03556.x. [DOI] [PubMed] [Google Scholar]
- Potenza MN, et al. Neural Correlates of Stress-Induced and Cue-Induced Drug Craving: Influences of Sex and Cocaine Dependence. Am J Psych. 2012;169(4):406–414. doi: 10.1176/appi.ajp.2011.11020289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaham Y, Hope BT. The role of neuroadaptations in relapse to drug seeking. Nature Neuroscience. 2005;8(11):1437–1439. doi: 10.1038/nn1105-1437. [DOI] [PubMed] [Google Scholar]
- Neisewander JL, et al. Fos protein expression and cocaine-seeking behavior in rats after exposure to a cocaine self-administration environment. J Neurosci. 2000;20(2):798–805. doi: 10.1523/JNEUROSCI.20-02-00798.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kosten TR, et al. Cue-induced brain activity changes and relapse in cocaine-dependent patients. Neuropsychopharmacology. 2006;31(3):644–650. doi: 10.1038/sj.npp.1300851. [DOI] [PubMed] [Google Scholar]
- Skolnick P, Volkow ND. Addiction therapeutics: obstacles and opportunities. Biol Psychiatry. 2012;72(11):890–891. doi: 10.1016/j.biopsych.2012.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Connor EC, Chapman K, Butler P, Mead AN. The predictive validity of the rat self-administration model for abuse liability. Neuroscience & Biobehavioral Reviews. 2011;35(3):912–938. doi: 10.1016/j.neubiorev.2010.10.012. [DOI] [PubMed] [Google Scholar]
- Dworkin SI, Mirkis S, Smith JE. Response-dependent versus response-independent presentation of cocaine: differences in the lethal effects of the drug. Psychopharmacology (Berl) 1995;117(3):262–266. doi: 10.1007/BF02246100. [DOI] [PubMed] [Google Scholar]
- Dworkin SI, Co C, Smith JE. Rat brain neurotransmitter turnover rates altered during withdrawal from chronic cocaine administration. Brain Res. 1995;682(1-2):116–126. doi: 10.1016/0006-8993(95)00327-m. [DOI] [PubMed] [Google Scholar]
- Twining RC, Bolan M, Grigson PS. Yoked delivery of cocaine is aversive and protects against the motivation for drug in rats. Behav Neurosci. 2009;123(4):913–925. doi: 10.1037/a0016498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gardner EL. What we have learned about addiction from animal models of drug self-administration. Am J Addict. 2000;9(4):285–313. doi: 10.1080/105504900750047355. [DOI] [PubMed] [Google Scholar]
- Cunningham KA, Anastasio NC. Serotonin at the nexus of impulsivity and cue reactivity in cocaine addiction. Neuropharmacology. 76 Pt B. 2014;76 Pt B:460–478. doi: 10.1016/j.neuropharm.2013.06.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carpenter KM, Schreiber E, Church S, McDowell D. Drug Stroop performance: relationships with primary substance of use and treatment outcome in a drug-dependent outpatient sample. Addict Behav. 2006;31(1):174–181. doi: 10.1016/j.addbeh.2005.04.012. [DOI] [PubMed] [Google Scholar]
- Copersino ML, et al. Cocaine craving and attentional bias in cocaine-dependent schizophrenic patients. Psychiatry Res. 2004;128(3):209–218. doi: 10.1016/j.psychres.2004.07.006. [DOI] [PubMed] [Google Scholar]
- Field M, Munafò MR, Franken IH. A meta-analytic investigation of the relationship between attentional bias and subjective craving in substance abuse. Psychol Bull. 2009;135(4):589–607. doi: 10.1037/a0015843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robbins SJ, Ehrman RN, Childress AR, O'Brien CP. Relationships among physiological and self-report responses produced by cocaine-related cues. Addict Behav. 1997;22(2):157–167. doi: 10.1016/s0306-4603(96)00007-x. [DOI] [PubMed] [Google Scholar]
- Cunningham KA, et al. Synergism between a serotonin 5-HT2A receptor (5-HT2AR) antagonist and 5-HT2CR agonist suggests new pharmacotherapeutics for cocaine addiction. ACS Chem Neurosci. 2013;4(1):110–121. doi: 10.1021/cn300072u. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fletcher PJ, Rizos Z, Sinyard J, Tampakeras M, Higgins GA. The 5-HT2C receptor agonist Ro60-0175 reduces cocaine self-administration and reinstatement induced by the stressor yohimbine, and contextual cues. Neuropsychopharmacology. 2008;33(6):1402–1412. doi: 10.1038/sj.npp.1301509. [DOI] [PubMed] [Google Scholar]
- See RE. Neural substrates of cocaine-cue associations that trigger relapse. Eur J Pharmacol. 2005;526(1-3):140–146. doi: 10.1016/j.ejphar.2005.09.034. [DOI] [PubMed] [Google Scholar]
- Shaham Y, Shalev U, Lu L, De Wit H, Stewart J. The reinstatement model of drug relapse: history, methodology and major findings. Psychopharmacology (Berl) 2003;168(1-2):3–20. doi: 10.1007/s00213-002-1224-x. [DOI] [PubMed] [Google Scholar]
- Meil WM, See RE. Conditioned cued recovery of responding following prolonged withdrawal from self-administered cocaine in rats: an animal model of relapse. Behav Pharmacol. 1996;7(8):754–763. [PubMed] [Google Scholar]
- Fuchs RA, Tran-Nguyen LT, Specio SE, Groff RS, Neisewander JL. Predictive validity of the extinction/reinstatement model of drug craving. Psychopharmacology (Berl) 1998;135(2):151–160. doi: 10.1007/s002130050496. [DOI] [PubMed] [Google Scholar]
- Grimm JW, Hope BT, Wise RA, Shaham Y. Neuroadaptation. Incubation of cocaine craving after withdrawal. Nature. 2001;412(6843):141–142. doi: 10.1038/35084134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panlilio LV, Goldberg SR. Self-administration of drugs in animals and humans as a model and an investigative tool. Addiction. 2007;102(12):1863–1870. doi: 10.1111/j.1360-0443.2007.02011.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anastasio NC, et al. Variation within the serotonin (5-HT) 5-HT2C receptor system aligns with vulnerability to cocaine cue reactivity. Transl Psychiatry. 2014;4:e369. doi: 10.1038/tp.2013.131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anastasio NC, et al. Functional status of the serotonin 5-HT2C receptor (5-HT2CR) drives interlocked phenotypes that precipitate relapse-like behaviors in cocaine dependence. Neuropsychopharmacology. 2014;39(2):370–382. doi: 10.1038/npp.2013.199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu HS, et al. Dorsolateral caudate nucleus differentiates cocaine from natural reward-associated contextual cues. Proc Natl Acad Sci U S A. 2013;110(10):4093–4098. doi: 10.1073/pnas.1207531110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swinford-Jackson SE, Anastasio NC, Fox RG, Stutz SJ, Cunningham KA. Incubation of cocaine cue reactivity associates with neuroadaptations in the cortical serotonin (5-HT) 5-HT2C receptor (5-HT2CR) system. Neuroscience. 2016;324:50–61. doi: 10.1016/j.neuroscience.2016.02.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Ciano P, Everitt BJ. Reinstatement and spontaneous recovery of cocaine-seeking following extinction and different durations of withdrawal. Behav Pharmacol. 2002;13(5-6):397–405. doi: 10.1097/00008877-200209000-00013. [DOI] [PubMed] [Google Scholar]
- Schmidt EF, et al. Extinction training regulates tyrosine hydroxylase during withdrawal from cocaine self-administration. J Neurosci. 2001;21(7):RC137. doi: 10.1523/JNEUROSCI.21-07-j0003.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Self DW, Choi KH, Simmons D, Walker JR, Smagula CS. Extinction training regulates neuroadaptive responses to withdrawal from chronic cocaine self-administration. Learn Mem. 2004;11(5):648–657. doi: 10.1101/lm.81404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutton MA, et al. Extinction-induced upregulation in AMPA receptors reduces cocaine-seeking behaviour. Nature. 2003;421(6918):70–75. doi: 10.1038/nature01249. [DOI] [PubMed] [Google Scholar]
- Gawin FH, Ellinwood EH. Cocaine dependence. Annu Rev Med. 1989;40:149–161. doi: 10.1146/annurev.me.40.020189.001053. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Context, ambiguity, and unlearning: sources of relapse after behavioral extinction. Biological Psychiatry. 2002;52(10):976–986. doi: 10.1016/s0006-3223(02)01546-9. [DOI] [PubMed] [Google Scholar]
- Torregrossa MM, Taylor JR. Learning to forget: manipulating extinction and reconsolidation processes to treat addiction. Psychopharmacology. 2013;226(4):659–672. doi: 10.1007/s00213-012-2750-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Conklin CA, Tiffany ST. Applying extinction research and theory to cue-exposure addiction treatments. Addiction. 2002;97(2):155–167. doi: 10.1046/j.1360-0443.2002.00014.x. [DOI] [PubMed] [Google Scholar]
- Carr KD, Kim G-Y, Cabeza de Vaca S. Chronic food restriction in rats augments the central rewarding effect of cocaine and the δ 1 opioid agonist, DPDPE, but not the δ 2 agonist, deltorphin-II. Psychopharmacology. 2000;152(2):200–207. doi: 10.1007/s002130000523. [DOI] [PubMed] [Google Scholar]
- Stamp JA, Mashoodh R, van Kampen JM, Robertson HA. Food restriction enhances peak corticosterone levels, cocaine-induced locomotor activity, and ΔFosB expression in the nucleus accumbens of the rat. Brain Research. 2008;1204:94–101. doi: 10.1016/j.brainres.2008.02.019. [DOI] [PubMed] [Google Scholar]
- Shalev U, Marinelli M, Baumann MH, Piazza P-V, Shaham Y. The role of corticosterone in food deprivation-induced reinstatement of cocaine seeking in the rat. Psychopharmacology. 2003;168(1-2):170–176. doi: 10.1007/s00213-002-1200-5. [DOI] [PubMed] [Google Scholar]
- Cunningham KA, et al. Selective serotonin 5-HT(2C) receptor activation suppresses the reinforcing efficacy of cocaine and sucrose but differentially affects the incentive-salience value of cocaine- vs. sucrose-associated cues. Neuropharmacology. 2011;61(3):513–523. doi: 10.1016/j.neuropharm.2011.04.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fowler JS, et al. Mapping cocaine binding sites in human and baboon brain in vivo. Synapse. 1989;4(4):371–377. doi: 10.1002/syn.890040412. [DOI] [PubMed] [Google Scholar]
- Kufahl PR, et al. Neural responses to acute cocaine administration in the human brain detected by fMRI. Neuroimage. 2005;28(4):904–914. doi: 10.1016/j.neuroimage.2005.06.039. [DOI] [PubMed] [Google Scholar]
- Goldberg SR, Hoffmeister F, Schlichting UU, Wuttke W. A comparison of pentobarbital and cocaine self-administration in rhesus monkeys: effects of dose and fixed-ratio parameter. J Pharmacol Exp Ther. 1971;179(2):277–283. [PubMed] [Google Scholar]
- Pickens R, Thompson T. Cocaine-reinforced behavior in rats: effects of reinforcement magnitude and fixed-ratio size. J Pharmacol Exp Ther. 1968;161(1):122–129. [PubMed] [Google Scholar]
- Boren JJ. Resistance to extinction as a function of the fixed ratio. J Exp Psychol. 1961;61(4):304–308. [Google Scholar]
- Miller WR, et al. PPARγ agonism attenuates cocaine cue reactivity. Addict Biol. 2016. [DOI] [PMC free article] [PubMed]
- Schmitz JM, et al. PPAR-gamma agonist pioglitazone modifies craving intensity and brain white matter integrity in patients with primary cocaine use disorder: a double-blind randomized controlled pilot trial. Addiction. 2017. [DOI] [PMC free article] [PubMed]
- Kalivas PW, Peters J, Knackstedt L. Animal Models and Brain Circuits in Drug Addiction. Molecular Interventions. 2006;6(6):339–344. doi: 10.1124/mi.6.6.7. [DOI] [PubMed] [Google Scholar]
- Reichel CM, Bevins RA. Forced abstinence model of relapse to study pharmacological treatments of substance use disorder. Curr Drug Abuse Rev. 2009;2(2):184–194. doi: 10.2174/1874473710902020184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ahmed SH, Koob GF. Transition from moderate to excessive drug intake: change in hedonic set point. Science. 1998;282(5387):298–300. doi: 10.1126/science.282.5387.298. [DOI] [PubMed] [Google Scholar]
- Gawin FH, Ellinwood EH. Cocaine and other stimulants. Actions, abuse, and treatment. N Engl J Med. 1988;318(18):1173–1182. doi: 10.1056/NEJM198805053181806. [DOI] [PubMed] [Google Scholar]
- Bozarth MA, Wise RA. Toxicity associated with long-term intravenous heroin and cocaine self-administration in the rat. JAMA. 1985;254(1):81–83. [PubMed] [Google Scholar]
- Liu Y, Roberts DCS, Morgan D. Sensitization of the reinforcing effects of self-administered cocaine in rats: effects of dose and intravenous injection speed. European Journal of Neuroscience. 2005;22(1):195–200. doi: 10.1111/j.1460-9568.2005.04195.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jackson LR, Robinson TE, Becker JB. Sex differences and hormonal influences on acquisition of cocaine self-administration in rats. Neuropsychopharmacology. 2006;31(1):129–138. doi: 10.1038/sj.npp.1300778. [DOI] [PubMed] [Google Scholar]
- Feltenstein MW, See RE. Plasma progesterone levels and cocaine-seeking in freely cycling female rats across the estrous cycle. Drug Alcohol Depend. 2007;89(2-3):183–189. doi: 10.1016/j.drugalcdep.2006.12.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreek MJ, Nielsen DA, Butelman ER, LaForge KS. Genetic influences on impulsivity, risk taking, stress responsivity and vulnerability to drug abuse and addiction. Nature Neuroscience. 2005;8(11):1450–1457. doi: 10.1038/nn1583. [DOI] [PubMed] [Google Scholar]
- Brenhouse HC, Andersen SL. Delayed extinction and stronger reinstatement of cocaine conditioned place preference in adolescent rats, compared to adults. Behav Neurosci. 2008;122(2):460–465. doi: 10.1037/0735-7044.122.2.460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kmiotek EK, Baimel C, Gill KJ. Methods for intravenous self administration in a mouse model. J Vis Exp. 2012. p. e3739. [DOI] [PMC free article] [PubMed]
- Grimm JW, et al. Brief exposure to novel or enriched environments reduces sucrose cue-reactivity and consumption in rats after 1 or 30 days of forced abstinence from self-administration. PLoS One. 2013;8(1):e54164. doi: 10.1371/journal.pone.0054164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grimm JW, Barnes J, North K, Collins S, Weber R. A general method for evaluating incubation of sucrose craving in rats. J Vis Exp. 2011. p. e3335. [DOI] [PMC free article] [PubMed]