

Abstract
Many animals, including the fruit fly, Drosophila melanogaster, are capable of discriminating minute differences in environmental temperature, which enables them to seek out their preferred thermal landscape. To define the temperature preferences of larvae over a defined linear range, we developed an assay using a temperature gradient. To establish a single-directional gradient, two aluminum blocks are connected to independent water baths, each of which controls the temperature of individual blocks. The two blocks set the lower and upper limits of the gradient. The temperature gradient is established by placing an agarose-coated aluminum plate over the two water-controlled blocks so that the plate spans the distance between them. The ends of the aluminum plate that is set on the top of the water blocks defines the minimum and maximum temperatures, and the regions in-between the two blocks form a linear temperature gradient. The gradient assay can be applied to larvae of different ages and can be used to identify mutants that exhibit phenotypes, such as those with mutations affecting genes encoding TRP channels and opsins, which are required for temperature discrimination.
Keywords: Neuroscience, Issue 136, Drosophila, larvae, thermotaxis, temperature gradient, behavior, somatosensation
Introduction
Thermotaxis is employed by mobile animals to select an environment with the most favorable conditions1,2,3. If the climate is excessively hot or cold, this behavior is vital for the survival. In addition, many animals are sensitive to very small differences in temperature in the comfortable range and seek out surroundings with an ideal temperature. This is of particular importance for poikilothermic organisms such as fruit flies, which equilibrate their body temperature with the environment. Assays to monitor larval thermotaxis have been instrumental in identifying and clarifying the roles of molecular sensors such as Drosophila Transient Receptor Potential (TRP) channels4,5,6, rhodopsins7,8, and ionotropic receptor receptors (IRs)9, which endow these animals with temperature sensitivities over different temperature ranges.
A two-way choice test provides one approach to study thermal preferences in larvae6,7. The assay entails establishing two distinct temperature zones and allows the animals to select one side over the other. The results from two-way choice tests can be robust, especially if the temperature differences between the two options are large. In addition, since each assay involves tabulating only two groups, the data can be expressed as a simple preference index. The ease and simplicity of two-way choice assays are also amenable to genetic screens. However, a major limitation is that many experiments are required to establish the preferred temperature of the wild-type or mutant animals.
A gradient assay offers the opportunity to establish the preferred temperature in a single assay8. Moreover, unlike the two-way choice test, it permits the evaluation of the distribution of a group of animals, when confronted with a continuous range of temperatures. One gradient assay uses a Petri dish and single animals and is well-suited for characterizing the detailed behavior of individual animals10. However, since Petri dishes are round, the sizes of the temperature zones vary and are progressively smaller depending on the distance from the center. Therefore, this setup is not ideal for monitoring the temperature selections of populations of animals.
A continuous thermal gradient apparatus that is well-suited to assess the temperature preferences of groups of larvae employs a rectangular arena and is described here. The apparatus is simple to construct and assemble. In addition, the gradient is linear, and is flexible, as it can be used to assess thermotaxis over large temperature ranges from 10 °C to 42 °C. The assay is rapid and straightforward to perform and yields reproducible data. In addition to reporting the favored temperature of larvae, it reveals the preferences of the population of animals over an entire linear range in a single experiment. Due to these advantages, it is an excellent choice for identifying genes required for thermotaxis.
Protocol
1. Equipment Fabrication and Assembling Apparatus for Gradient Assays
- Fabricate the aluminum assay plates for the single-directional gradient assay.
- Trim and grind each aluminum assay plate (Figure 1A) out of a single piece of aluminum using a band saw and sharp vertical mill with the following dimensions: the outer size is 140 x 100 x 9 mm and the inner size is 130 x 90 x 8 mm (Figure 1B). Anodize the inside of each assay plate with black paint to make it easier to visualize the larvae and prevent rust. NOTE: Fabrication of the aluminum assay plates for the single-directional gradient is done by a machine shop. The anodization is performed by a commercial vendor.
- Mark the upper and lower rims of each assay plate with 13 demarcations, separated by 10 mm using a permanent marker. Place the first and last demarcations 5 mm from the inner edge of the assay plates (Figure 1A, B).
To fabricate the aluminum assay plate for the bidirectional gradient, cut out an aluminum plate with the following dimensions using sheet metal shears: 250 x 220 x 2 mm. Anodize the assay plates with black paint. NOTE: Fabrication of the aluminum assay plates for the bidirectional gradients is performed by a machine shop. The anodization is done by a commercial vendor.
- Fabricate the aluminum blocks. NOTE: Two blocks are needed for the single-directional gradient and three blocks are needed for the bidirectional gradient.
- Drill the water channel inside the block (7 mm in diameter) using a vertical mill and thread grooves in the open ends on one side (Figure 1D). Close the grooved holes with bolts and thread seal tape to form a U-shaped water channel (Figure 1C, D). NOTE: The fabrication of the aluminum assay blocks is performed by a machine shop.
- Fix the connector in the aluminum block (Figure 1D) with screw threads and thread seal tape at one end. Fit the other end with multiple barbs into silicone tubing (1/4" ID x 3/8" OD x 1/16" wall).
Obtain two refrigerated/heated circulating baths.
- Assemble temperature-controlled blocks for the gradient assay system.
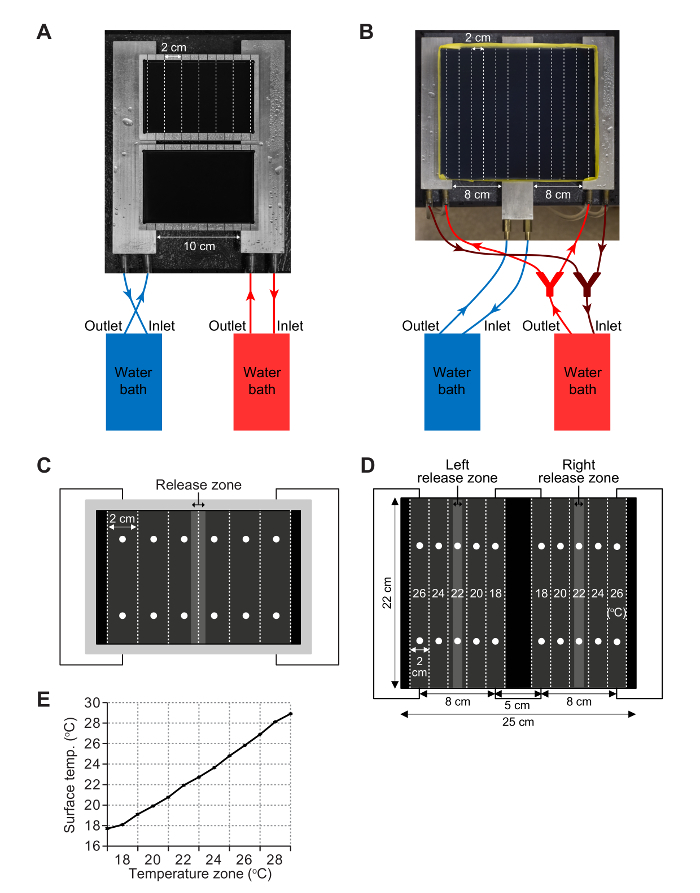
- For the single-directional gradient, connect each of the two aluminum blocks to a separate water bath so that the temperature of each block is controlled by an independent water circulating system (Figure 2A). Use silicone tubing (1/4" ID x 3/8" OD x 1/16" wall) to connect the outlet of the water bath to the connector on the inner side of the aluminum block (Figure 2A). Also, use tubing to connect the outer side connector on the aluminum block to the inlet of the water bath.
- For the bidirectional gradient, place two of the three blocks under the left and right sides of the plate and connect them to the same water bath with the tubing (Figure 2B). Connect the third aluminum block to a second water bath and place below the center of the plate (Figure 2B). NOTE: This apparatus and assay are only necessary if the larvae accumulate in the zone at one edge or the other of the single-directional gradient. If so, it is important to assess whether or not there is an edge effect. To test this possibility, establish a bidirectional gradient in which the edge zone temperature preferred by the larvae using the single-directional gradient (e.g. 18 °C) is the temperature in the middle of the plate of the bidirectional gradient (Figure 3E, F).

2. Larval Synchronization
- Nourish flies to be used for egg laying.
- Mix yeast granules and distilled water with a pestle to make yeast paste. Thoroughly grind and stir until the consistency of the paste is similar to peanut butter. Avoid excessively dry paste, which may flake off when transferring flies to fresh vials, or wet paste, which could trap the flies.
- Using a pestle, add the yeast paste close to the inner walls of standard Drosophila vials, just above the surface of the food.
- Count 12-35 females and up to half as many males (but no more than 10) on a CO2 pad, and add the females and males to each yeast paste-containing vial.
- To keep the fly food with the yeast-paste moist, while the flies feed, combine these vials in a tray that holds up to 100 vials with 20 open vials containing distilled water only. Place the tray in a clear plastic bag and seal.
- Incubate the tray in a 25 °C incubator for 48 h so that the females are nourished adequately, thereby enabling them to lay large numbers of eggs.
- Set up vials for egg laying.
- Transfer the nourished flies into new food vials for egg-collecting by tapping them over. Do not use CO2, which may cause the flies to lay fewer eggs over the following short time window. NOTE: Standard Drosophila food vials are used without yeast paste for egg collection.
- Allow the flies to lay eggs for 3 h at 25 °C so that all eggs are collected over a relatively narrow time window. NOTE: This will enable the larvae to be synchronized at the same developmental stage.
- Place the vials containing the eggs in the tray containing the water vials and tighten the bag. Incubate at 25 °C under 12 h light:12 h dark cycles.
- Age the larvae for a given number of hours after egg laying (AEL) depending on the larval stage desired. NOTE: Table 1 is an estimation of the relationship between the hours AEL and the larval stage, which will vary depending on the fly stock, food used, rearing temperature etc. The precise relationship between AEL and developmental stage needs to be verified by each investigator using physical criteria, such as the morphology of the mouth hooks and spiracles11.
3. Temperature Gradient Setup
- Prepare single-directional gradient
- To create a humid ambient environment during the assays, place the two aluminum blocks connected to two water baths on wet paper towels, separated by 10 cm (Figure 2A).
- Turn on the two water baths ~2 h prior to initiating the assay to allow enough time for the temperature of the aluminum blocks to equilibrate. NOTE: It is recommended to perform a mock experiment to determine the temperature of each water bath that is needed to achieve the desired linear temperature gradient. Some typical temperature pairs to set the water baths are listed in Table 2. However, note that the surface temperatures are affected by ambient temperature. The length of tubing also affects the temperatures as there is cooling in the tubes. The length of the tubing in our setup is 1.5 m.
- Microwave 100 mL of 1% agarose at the high-power setting in a 500 mL round wide-mouth bottle and pour 25 mL/assay plate on a level benchtop. Prepare two assay plates, which can be placed on the aluminum blocks at the same time (Figure 2A).
- After the agarose has solidified (10-20 min), gently rub each agarose surface with a standard kitchen sponge or a melamine sponge to make the agarose surface slightly coarse, so that when spraying water on the agarose gel, a smooth thin water membrane will be created, and no water droplets will form.
- Fully submerge the plates in a container with distilled water until the assay is ready to be performed to prevent the plates from getting desiccated.
- Prepare a bidirectional gradient (optional)
- To create a humid ambient environment during the assays, place three aluminum blocks 8 cm apart (Figure 2B) on wet paper towels.
- Turn on the two water baths ~2 h prior to initiating the assay to allow enough time for the temperature of the aluminum blocks to equilibrate. NOTE: It is recommended to perform a mock experiment to determine the temperature of each water bath for the bidirectional gradient. Some typical temperature pairs to set the water baths are listed in Table 3. However, note that the surface temperatures are affected by ambient temperature. The length of tubing also affects the temperatures as there is cooling in the tubes. The length of the tubing in our setup is 1.5 m.
- To prevent the agarose from spilling out of the plate, wrap the edges of the aluminum plate with labeling tape to form a 10 mm-high wall (Figure 2B).
- Using the high-power setting, microwave 200 mL of 1% agarose in a 500 mL round wide-mouth bottle and pour 120 mL on an assay plate.
- After the agarose has solidified (~30 min), gently rub each agarose surface with a standard kitchen sponge or a melamine sponge to make the agarose surface slightly coarse, so that when spraying water on the agarose gel, a smooth thin water membrane is created and no water droplets form.
- Fully submerge the assay plates in a container with distilled water until the assay is ready to be performed to prevent them from drying out.
- Set up single-directional gradient
- Prepare reagents and items on a bench next to the gradient assay apparatus (see the Table of Materials).
- To promote efficient temperature transfer, fill any gaps between the aluminum blocks and the assay plates by spraying water at the interface between the blocks and plate.
- Using gloves, remove the assay plates from the water container. If water invades between the agarose gel and the plate and causes bumps to form on the surface, remove the water with a P1000 micropipette.
- Place the assay plates on the aluminum blocks so that the demarcations that are 2 cm from either edge exactly match the edges of the aluminum blocks (Figure 2A, C).
- Spray water onto the surface of the plate (a thin water membrane covering the agarose surface is sufficient) to prevent the agarose gel from drying out. Make sure that the water membrane is continuous and free of water droplets since larvae can get trapped in water droplets.
- Cover the gradient system with a cardboard box to reduce water evaporation and help stabilize the temperature of the gel surface. Wait for 5-10 min to allow the temperature to equilibrate.
- Check the surface temperature at 12 points on the plate (Figure 2C). Take two measurements within each zone to establish whether or not there is variability within a zone. Make sure the temperature at both spots is within ± 0.2 °C of the desired temperature. NOTE: Variability within a zone usually occurs because the exact distances of the two spots to the edge of the aluminum blocks are not identical. Adjust the position of the plate on the aluminum blocks to make sure that the demarcations that are 2 cm from either edge exactly match the edges of the aluminum blocks.
- If the measured temperature gradient deviates from the desired gradient, increase or decrease the water bath temperature setting(s) and recheck the surface temperature after the temperature of the water bath(s) stabilize(s).
- Cover the gradient system with a cardboard box until starting the assay.
- Set up bidirectional gradient (optional)
- Prepare reagents and items on a bench next to the gradient assay apparatus (see the Table of Materials).
- Spray water on the surfaces of the three aluminum blocks to promote efficient temperature transfer from the aluminum blocks to the assay plates by filling any gaps between the surfaces.
- Remove the assay plates carefully from the water container and place on the aluminum blocks. If water invades between the agarose gel and the plate, which might cause bumps to form on the surface, remove the water with a P1000 micropipette.
- Place the assay plate on the aluminum blocks so that the midlines of the first and last zones from either edge exactly match the edges of the two side aluminum blocks (Figure 2B, D).
- Spray water onto the surface of the plate (a thin water membrane covering the agarose surface is sufficient) to prevent the agarose gel from drying out. Make sure that the water membrane is continuous and free of water droplets, since larvae can get trapped in water droplets.
- Cover the gradient system with a cardboard box to reduce water evaporation and help stabilize the temperature of the gel surface. Wait 5-10 min to allow the temperature to equilibrate.
- Check the surface temperature at two points along the midline of each of the 10 zones (Figure 2D). Take two measurements within each zone to establish whether or not there is variability within a zone. Make sure the temperature at both spots is within ± 0.2 °C of the desired temperature. NOTE: Variability within a zone usually occurs because the exact distances of the two spots to the edge of the aluminum blocks are not identical. Adjust the position of the plate on the aluminum blocks to make sure that the midlines of the first and last zones nearest each edge exactly match the edges of the two side aluminum blocks.
- If the measured temperature gradient deviates from the desired gradient, adjust the water bath temperature setting(s) and recheck the surface temperature after the temperature of the water bath(s) stabilize(s).
- Cover the blocks with a cardboard box until the start of the assay.
4. Larval Collection and Washing
- Isolate clean larvae (option 1)
- Add ~40 mL 18% sucrose solution to a 50 mL test tube. Scoop all the larvae from the food vial(s) with a scoopula and transfer to the 18% sucrose solution.
- Mix thoroughly but gently using a scoopula to separate larvae from the food debris. Wait for 30-60 s until the larvae float to the top layer of the tube. Pour the top layer containing the larvae (~10 mL) to another 50 mL tube and fill the tube with fresh 18% sucrose solution. Wait for 30-60 s until the larvae float to the top layer again. NOTE: Large food particles sometimes transfer together with larvae and may affect larval thermotaxis if they are not removed. If large food particles remain in the top layer, repeat steps 4.1.1-4.1.2, or manually remove them using a scoopula.
- Transfer the top layer of larvae (~10 mL) to two other 50 mL tubes and fill the two tubes with distilled water to reduce the sucrose concentration, so that larvae quickly sink in water. Wait for 30-60 s until the larvae sink to the bottom. Perform this and the following steps as quickly as possible to prevent the larvae from drowning.
- Discard as much of the water as possible by gently tilting the tubes. Combine larvae into one tube by pouring out the larvae with the remaining water and fill the tube with distilled water.
- Wait for 30-60 s until all larvae sink down and remove as much water as possible by gently tilting the tubes. Repeat this washing step 2-4 times to entirely remove sucrose and all visible food.
- Discard as much of the water as possible and transfer the larvae to an empty 35 mm Petri dish by decanting. Spray the tube with water and use a small paint brush to help transfer the larvae.
- Remove excess water from the Petri dish using a P1000 micropipette. Leave ~0.5 mL of water to prevent dehydration of the larvae.
- Place the lid on the dish to prevent the larvae from escaping. Turn the lid upside down to reduce the ability of larvae to escape through the small gap between the lid and dish. Allow the larvae to recover for 10-20 min.
- Isolate clean larvae (option 2) NOTE: This alternative method for cleaning larvae mitigates the possibility of suffocating the larvae during the washing procedure. To use this method, proceed with the following steps after performing the above steps 4.1.1-4.1.2.
- Place a cell strainer (300 µm, retains larvae 72 h AEL or older) on top of a 50 mL tube.
- After confirming that there is no adult bodies or food debris floating on the surface of the sucrose solution, pour the top layer containing the larvae through the strainer to trap all the larvae on the mesh screen. Wash the larvae thoroughly with distilled water until the 50 mL tube is filled.
- Remove the cell strainer with the larvae and dispose of the used water from the tube. Place the strainer containing the larvae on top of the empty tube and wash the larvae again with distilled water until the 50 mL tube is filled.
- Turn the strainer upside down over an empty 35 mm Petri dish, and spray water from the top of the cell strainer to transfer the larvae to the Petri dish. Transfer the remaining larvae using a small paint brush.
- Continue to step 4.1.7 above before proceeding to step 5.
5. Assay and Calculation
Remove the cardboard box and check the gel surface temperature immediately before transferring the larvae to the plate. Minimize the time that the cardboard box is open to prevent disrupting the temperature equilibration. If the surface is dry, spray a small amount of water on the surface.
Distribute 150 ± 50 larvae near the center of each plate (between zones 3 and 4 out of the 6 zones) for the single-directional gradient (release zone; Figure 2C). NOTE: For the bidirectional gradient, distribute 200-400 larvae along the middle zone of each half (Figure 2D).
Place a microplate lid over each assay plate to prevent the larvae from crawling out. Cover the setup with a cardboard box to prevent light exposure, which might affect larval choice on the agarose gel. NOTE: For the bidirectional gradient, it should not be necessary to cover the plate with a lid, because the plate is larger, and the preferred temperature of the larvae is in the middle zone. Consequently, few larvae accumulate at the edges and have an opportunity to crawl out.
Allow the assay to proceed for 10-30 min for the single-directional gradient, and for 15-35 min for the bidirectional gradient, depending on the age of the larvae (Table 4).
Remove the cardboard box and the microplate lid. Photograph the plates from above using a digital camera. Take two photographs of each assay plate so that the investigator can choose the one with better contrast and brightness for analysis.
Remove all of the larvae from the assay plates and anywhere outside the plates by aspiration.
Clean the assay plates, the tubes, and the cell strainer thoroughly with distilled water. Reuse the assay plates prepared that day unless the surface of the agarose is damaged.
- Calculate the percentage distribution of larvae in each zone.
- Open the photographic image of the assay results using any software that allows adding markings to the image. To indicate the assay zones, draw vertical lines every 2 cm based on the demarcations on the assay plate.
- Count the number of larvae in each zone and record the numbers. Do not count larvae in regions 0.5 cm from any of the walls. The gel is thicker near the walls, and the surface temperatures are not linear in these regions.
- For the single-directional gradient, calculate the percentage distribution in each zone as follows: (number of larvae in a given 2-cm zone)/ (total number of larvae in 6 zones) x 100.
- For the bidirectional gradient, calculate the percentage distribution in each zone as follows: (number of larvae in a given 2-cm zone)/ (total number of larvae in 5 zones on each side of the gradient) x 100.
Representative Results
To establish an 18 °C-28 °C single-directional gradient, we set the temperatures of two water baths to 16.8 °C and 31 °C. We obtain the temperatures at 13 points by measuring the temperature at 26 positions within the upper and lower portions of all 6 zones, the border lines between the zones, and at the extreme ends of the agarose gel surface (Figure 2C, 2E). The temperature distribution along the gradient was nearly linear (Y = 0.9672*X + 16.19, R2 = 0.9961) (Figure 2E).
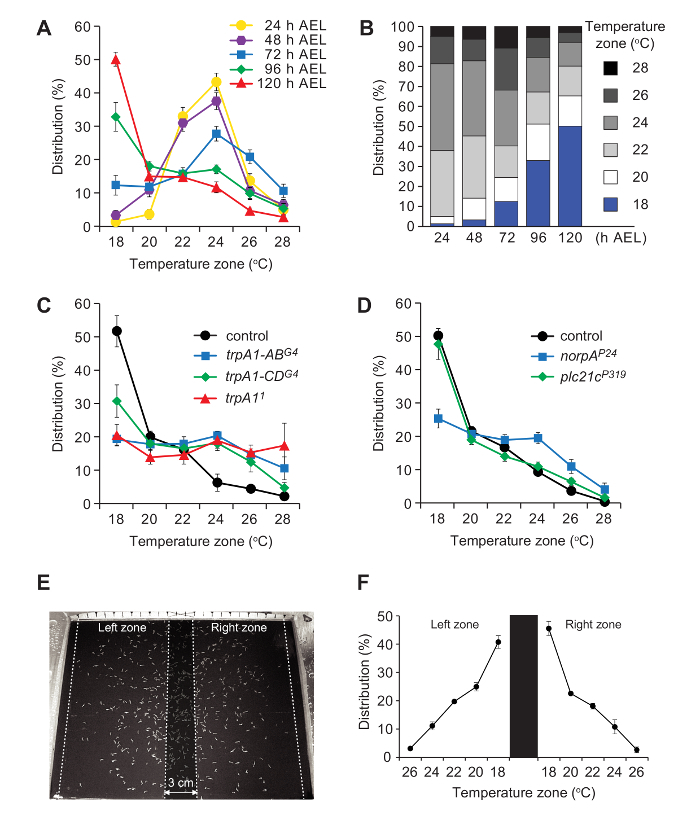
We assayed the temperature preferences of control (w1118) larvae at various ages. 1st (24 ± 1.5 h AEL), 2nd (48 ± 1.5 h AEL) and early-3rd larvae (72 ± 1.5 h) showed peaks in the 24 °C zone (Figure 3A, B). The temperature preferences changed during 3rd instar larval development. The largest percentage of mid-3rd instar (96 ± 1.5 h AEL) accumulated in the 18 °C zone (Figure 3A, B), and this bias increased with age. Among late-3rd instar larvae (pre-climbing; 120 ± 1.5 h AEL), ~50% clustered in the 18 °C zone, and the selection of this temperature was ~4-fold higher than the adjacent 20 °C zone (18 °C zone, 50.2%; 20 °C zone, 15.1%; Figure 3A, B). The proclivity to accumulate in the 18 °C zone in w1118 was not due to an edge effect since the late-3rd instar larvae still accumulated in the 18 °C zone, using a bidirectional temperature gradient (Figure 3E, F).
We also tested late-3rd instar larvae (120 ± 1.5 h AEL) with mutations in genes required for discriminating temperature differences in the comfortable range. These include trpA1, which is required for normal temperature selection in the 18 °C-24 °C range5,7,8. Larvae with a null mutation in trpA1 (trpA11) distribute equally over the entire 18 °C—28 °C gradient (Figure 3C). Larvae missing just the A and B isoforms (trpA1-ABG4), or the C and D isoforms (trpA1-CDG4) also show severe impairments (Figure 3C). Flies encode two isoforms of phospholipase Cβ (PLC21C and NORPA), and mutations affecting norpA (norpAP24) but not plc21c (plc21cP319) also disrupt accumulation in the 18 °C range (Figure 3D).
Figure 1. Apparatus for performing the single-directional temperature gradient assay. (A) An aluminum test plate used for assaying larval thermotaxis behavior. The 13 black lines at the top and bottom demarcate 12 zones (10 mm each). The bottom of the plate is anodized with black paint so that it is easier to visualize the larvae. (B) Dimensions of the aluminum plate (indicated in mm). The outer size of the aluminum plate is 140 x 100 x 9 mm. The inner size of the aluminum plate is 130 x 90 x 8 mm. The demarcations are separated by 10 mm. The first and last demarcation is 5 mm from the edges of the inner area of the plate. (C) Top and side views of one of the aluminum blocks used to control the temperature of the gradient. The block has two connectors used to attach to silicon tubes, which connect to a water bath. (D) Dimensions of an aluminum block. The outer size of the aluminum block is 255 x 50 x 14 mm. The diameter of the inner water path is 7 mm. Two 30 mm connectors on the left connect with silicon tubing that extends to the water bath. Please click here to view a larger version of this figure.
Figure 2. Single and bidirectional gradient assay setups. (A) Single-directional gradient setup with two aluminum assay plates on two aluminum blocks. The temperatures of the aluminum blocks are controlled by circulating water from two water baths. (B) The arrangement of the three aluminum blocks, water baths and an aluminum plate (250 x 220 mm) for a bidirectional gradient. The left and right blocks are connected to the same water bath and the middle block is connected to the other water bath. The aluminum assay plate is wrapped with tape to form a 10 mm wall to contain the 1% agarose. (C) Positions to check temperatures (indicated by dots) and to release larvae on the plate. Before initiating an experiment, check the temperature at two points within each zone to confirm that the desired linear temperature gradient is established. Larvae are released within the indicated area near the midline. The larvae are counted within each of the 2-cm zones. (D) Positions to check temperatures (indicated by dots) and the release zones for the larvae on a bidirectional gradient. An equal number of larvae are released along the midline of each half of the bidirectional gradient. The numbers of larvae are counted in each of the 10 (2 cm) zones. One typical set of temperatures (18 °C-26 °C) on the agarose surface is indicated. (E) Temperatures measured along the border lines and midlines of each zone in a sample single-directional gradient. Data represent mean temperatures ± SD. n=8 assays (150 ± 50 larvae/assay). Parts of this figure are reproduced from Sokabe et al.8 with slight modifications. Please click here to view a larger version of this figure.
Figure 3. Representative results using single-directional and bidirectional gradient assays. (A, B) Mean percentages of larvae in 6 zones on the single-directional gradient. Data include 3rd instar larvae at the indicated hours after egg laying (AEL). n=6-7. The error bars in A represent ±SEM. (C) Thermal distributions of the pre-climbing late-3rd instar larvae of control (w1118) and trpA1 mutants on the single-directional gradient. n=3-4. The error bars represent ±SEM. (D) Thermal distributions of the late-3rd instar larvae of control (w1118) and PLCβ mutants on the single-directional gradient. n=4-6. The error bars represent ±SEM. (E) Representative distribution of pre-climbing, late-3rd instar larvae (w1118) on the bidirectional gradient. The left and right assay zones are indicated by dotted lines and separated by the no-count zone in the center (shaded region). (F) Percentage of pre-climbing late-3rd instar larvae (w1118) in each zone along the thermal gradient. Larvae were placed in the left and the right release zones. The assay zones are separated by a 3-cm no-count zone in the center and the distributions are calculated independently. The error bars represent SEMs. n=3 assays (200-400 larvae/assay). Parts of this figure are reproduced from Sokabe et al.8 with slight modifications. Please click here to view a larger version of this figure.
| Hours AEL | Larval stage |
| 24 | 1st instar |
| 48 | 2nd instar |
| 72 | Early-3rd instar |
| 96 | Mid-3rd instar |
| 120 | Late-3rd instar, just before climbing stage |
Table 1. The relationship between the hours after egg laying (AEL) and larval stages.
| Temperature gradient on agarose plate (slope) | Temperatures of water baths | Temperatures of aluminum blocks |
| 10.0-25.0°C (1.5°C/cm) | ~6.5-7°C/~28.5°C | ~8.5°C/~26.8°C |
| 18.0-28.0°C (1°C/cm) | ~16.8°C/~31.0°C | ~17.8°C/~29.7°C |
| 14.0-34.0°C (2°C/cm) | ~10.0°C/~40.0°C | ~11.8°C/~36.8°C |
| 12.5-42.0°C (2.95°C/cm) | ~7.0°C/~55.0°C | ~9.4°C/~49.4°C |
Table 2. Typical temperature gradients and the corresponding temperatures of the water baths and aluminum blocks for single directional gradients.
| Temperature gradient on agarose plate | Temperatures of water baths | Temperatures of aluminum blocks |
| 22-10-22°C (1.5°C/cm) | ~5.0°C /~25.0°C | ~7.5°C/~24.0°C |
| 26-18-26°C (1°C/cm) | ~15.8°C /~30.6°C | ~16.9°C/~28.4°C |
| 30-14-30°C (2°C/cm) | ~8.5°C /~36.4°C | ~10.9°C/~32.8°C |
| 36-12.5-36°C (2.95°C/cm) | ~5.0°C /~47.2°C | ~7.9°C/~40.9°C |
Table 3. Typical temperature gradients and the corresponding temperatures of the water baths and aluminum blocks for bidirectional gradients.
| Larval age (AEL) | Assay time (single directional) | Assay time (bidirectional) |
| 24 h | 30 min | 35 min |
| 48 h | 22 min | 27 min |
| 72 h | 16 min | 21 min |
| 96 h | 13 min | 18 min |
| 120 h | 10 min | 15 min |
Table 4. Different larval ages (AEL) and the corresponding assay times.
Discussion
To ensure the success of this protocol, it is important to take steps to obtain adequate numbers of larvae to perform the experiments. These include pre-feeding the flies in yeast paste-containing vials for 2-3 d to improve egg laying. The vials need to be placed in a tray containing water vials and enclosed in a clear plastic bag, which maintains the moisture of the food and promotes effective feeding by the larvae while permitting exposure to normal light-dark cycles. However, the yeast paste should not be so soft that the flies become trapped. The number of females per vial depends on the genotype. In the case of w1118, it is usually adequate to allow ~12 females and ~6 males to lay eggs for 3 h. Two vials typically provide enough larvae (100-200 larvae) to place on a plate for the single-directional assay. If the fly stock lays fewer eggs than wild-type or the proportion of larvae that hatch is reduced, add additional females (up to 30-35/vial) and males.
There are many causes of unintended variability when performing these assays. The developmental stage affects the temperature preference of the larvae. Therefore, it is imperative to carefully use larvae of a defined stage. To do so, collect larvae over a narrow time frame (such as 3 h). Since the rearing conditions (temperature, humidity, light-dark cycle and type of food) and some mutations affect the rate of development, do not rely strictly on the time after egg laying to analyze comparably aged larvae. It is also important to examine physical traits, such as the size of the mouth hooks and spiracles7 to determine whether larvae of different genotypes are at the same developmental stage. The times for 1st, 2nd and 3rd instar larvae to develop (Table 1) are based on using corn meal-based food and incubation at 25 °C under 12 h light:12 h dark cycles. Larvae grow slower on molasses-based food. Proper hydration and food freshness also affects the growth of larvae. The amount of water in the food should leave the food neither too dry to peel back from the vial nor too loose due to excessive water. Ideally, the ~5 mm layer nearest the food surface is conditioned by the growing larvae and moves easily when the vial is tilted or tapped. This condition can be achieved using freshly made food with 50-100 larvae.
It is essential to establish a stable temperature gradient before initiating the assay. Therefore, turn on the water bath 1-2 h prior to initiating the assay, as the water baths produce heat and might change the ambient room temperature, which in turn have the potential to affect the gradient. After 1-2 h, the ambient temperature of most air-conditioned rooms equilibrates. However, this must be determined in each environment. In addition, when generating a gradient with a large temperature range (>3.0 °C/cm), it can be difficult to obtain a stable gradient. A tiny movement of the assay plate more significantly changes the temperature gradient when the range is large (e.g. >3.0 °C/cm). We found that a 1-2 °C/cm gradient produces the most stable gradients.
There are several additional considerations that need to be controlled to limit variability in the thermotaxis assays. Washing the larvae is critical, as the presence of food particles or sucrose on the larvae can impact the assay. The washing steps must be performed thoroughly but quickly, because limiting their oxygen supply excessively while they are submerged in water may affect their health. Therefore, we recommend using the cell strainer (option 2) to clean the larvae. A strainer with a pore size of 300 µm works well for larvae at the early-3rd instar stage (72 h AEL) or older. In addition, it is important to perform experiments at the same time of day, such as from Zeitgeber time (ZT) ZT4 to ZT8 (ZT=0 is when lights are turned on), to limit variability due to impacts of circadian rhythms on temperature selection. The level of moisture on the agarose plate can also cause variability in the results. While the surface of the plates needs to be moist, water droplets could trap the larvae, and therefore need to be avoided.
Limiting variability in the assays will make it possible to obtain robust results without performing large numbers of independent experiments. Typically, 3-8 independent experiments are sufficient to obtain a reliable outcome. Proper tabulation of the results is also critical for the success of the thermotaxis assays. Do not count larvae distributed in the regions within 0.5 cm from the aluminum walls, since the temperatures are not linear in these regions. Some larvae will be positioned at the border between two zones, at the time the assay is concluded. If more than 50% of the body length resides in one zone, then include the larva in the tabulation for that zone. If a larva is precisely 50% in each of two zones, then count the animal as 0.5 larvae in each zone.
While the gradient assay allows the larvae to discriminate a range of temperature differences, there are limitations to the lower and upper temperatures that can be effectively tested. It is not feasible to test temperatures <10 °C since the mobility of the larvae decreases greatly at lower temperatures. Although gradients with the upper temperature reaching 42 °C can be established, almost no control larvae stay in any zone >28 °C. Therefore, continuous gradient assays cannot be used to discriminate temperature preferences between various zones >28 °C. However, gradient assays at these elevated temperatures could potentially be used to characterize mutants if they have highly shifted warm temperature preferences.
If a mutant has a locomotor defect, it may be necessary to conduct additional experiments to make sure that the apparent temperature preference is not inaccurate due to a movement impairment. To remedy this potential problem, it may be necessary to establish longer assay times to allow the animals additional time to move to their desired zones. The bidirectional gradient assay can also be employed to test whether the animals select the same preferred zone, regardless as to where they are placed initially on the plate.
In conclusion, the gradient assays described here have advantages over other thermotaxis assays. While two-way choice assays are useful, as they can yield robust results, especially when the temperature differences between the two zones are large, they are not effective in revealing the ideal temperature preferred by the larvae in a single experiment. To do so requires performing many two-way choice combinations to determine the most preferred temperature5,6,7. In contrast, the gradient assay provides the animal an opportunity to select their preferred temperature zone in an environment with a single continuous temperature landscape. Thus, it is unnecessary to devise a large combination of two-way choices to determine the preferred thermal environment. Studying thermotaxis using a round Petri dish has been employed to assess the effects of temperature on the moving dynamics of a larva10. However, it is less useful in discriminating between preferences between different temperatures, since each zone is a different size. Finally, due to the simplicity of the assay, and its utility in discriminating small temperature preferences in a single assay, it can be employed to screen dozens of candidate genes that are potentially involved in larval temperature preference that might otherwise be overlooked using other assays
Disclosures
The authors have nothing to disclose.
Acknowledgments
C.M. is supported by funding from the NEI (EY008117, EY010852), NIDCD (DC007864, DC016278) and the NIAID (1DP1AI124453).
References
- Fowler MA, Montell C. Drosophila TRP channels and animal behavior. Life Sci. 2013;92:394–403. doi: 10.1016/j.lfs.2012.07.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palkar R, Lippoldt EK, McKemy DD. The molecular and cellular basis of thermosensation in mammals. Curr Opin Neurobiol. 2015;34:14–19. doi: 10.1016/j.conb.2015.01.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vriens J, Nilius B, Voets T. Peripheral thermosensation in mammals. Nat Rev Neurosci. 2014;15(9):573–589. doi: 10.1038/nrn3784. [DOI] [PubMed] [Google Scholar]
- Rosenzweig M, et al. The Drosophila ortholog of vertebrate TRPA1 regulates thermotaxis. Genes Dev. 2005;19:419–424. doi: 10.1101/gad.1278205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwon Y, Shim HS, Wang X, Montell C. Control of thermotactic behavior via coupling of a TRP channel to a phospholipase C signaling cascade. Nat Neurosci. 2008;11:871–873. doi: 10.1038/nn.2170. [DOI] [PubMed] [Google Scholar]
- Kwon Y, Shen WL, Shim HS, Montell C. Fine thermotactic discrimination between the optimal and slightly cooler temperatures via a TRPV channel in chordotonal neurons. J Neurosci. 2010;30(31):10465–10471. doi: 10.1523/JNEUROSCI.1631-10.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shen WL, et al. Function of rhodopsin in temperature discrimination in Drosophila. Science. 2011;331(6022):1333–1336. doi: 10.1126/science.1198904. [DOI] [PubMed] [Google Scholar]
- Sokabe T, Chen HS, Luo J, Montell C. A switch in thermal preference in Drosophila larvae depends on multiple rhodopsins. Cell Rep. 2016;17:336–344. doi: 10.1016/j.celrep.2016.09.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ni L, et al. The Ionotropic Receptors IR21a and IR25a mediate cool sensing in Drosophila. Elife. 2016;5:13254. doi: 10.7554/eLife.13254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo L, et al. Navigational decision making in Drosophila thermotaxis. J Neurosci. 2010;30(12):4261–4272. doi: 10.1523/JNEUROSCI.4090-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner M, Golic KG, Hawley RS. Drosophila: a laboratory handbook. Cold Spring Harbor Laboratory Press; 2005. [Google Scholar]