

Abstract
Determinations of the NH3 compensation point for the understory plant of semi-natural woodlands Luzula sylvatica (Huds.) Gaud. were carried out by measurements of gas exchange and by calculation from the NH4+ concentration and pH of extracts of the foliar apoplast. Compensation points determined by gas exchange measurements were among the lowest yet reported (0.51–1.10 μg NH3 m−3) and those calculated from apoplast extracts were lower than any yet reported (0.017–0.54 μg NH3 m−3). Those determined by gas exchange were consistently found to be between 2 and 30 times higher than those determined from apoplast extracts. Consideration of possible causes of this discrepancy, which is not confined to this investigation, showed that all likely errors would result in an increase in the discrepancy, or were insufficient to account for observed differences. It is suggested that spatial variability of pH and NH4+ concentration within the foliar apoplast represents the most promising line for further investigation. It is also shown that the foliar apoplast of L. sylvatica is sufficiently buffered to eliminate the need for correction of H+ concentration for dilution during extraction, but that it is necessary to correct the NH4+ concentration of apoplast extracts for dilution.
After N2 and N2O, NH3 is the most abundant nitrogen species in the atmosphere (Raven et al., 1992) with NH4+ being the dominant cation in precipitation in Europe and North America for the last four decades (Flechard, 1998). There is increasing evidence to suggest that where the atmosphere becomes artificially enriched with NH3 there may be detrimental effects on sensitive (low nitrogen) ecosystems due to eutrophication. The best documented instances of eutrophication due to nitrogen deposition have been the replacement of ericaceous species, such as Calluna vulgaris and Erica tetralix, by grass species such as Molinia caerulea and Deschampsia flexuosa (e.g. Heil and Diemont, 1983; Roelofs, 1986; Leith et al., 1999). Furthermore, although NH3 rapidly protonates to NH4+ in solution, metabolism of NH4+ by plants and microbes can lead to the acidification of poorly buffered ecosystems (van Breemen et al., 1982, 1983). Raven (1988) calculated that the uptake and assimilation of NH3 by the aerial parts of plants leads to the generation of 0.22 H+ N−1, whereas uptake and assimilation of NH4+ leads to the generation of 1.22 H+ N−1. Nitrification of NH4+ by soil microbiota leads to the generation of 2 H+ N−1 (van Breemen et al., 1983), whereas there is a net generation of 1 H+ N−1 if deposition is as NH3.
It was demonstrated as early as 1850 that plants can take up and utilize gaseous NH3 (Ville, 1850; cited in Sutton et al., 1993). It has since become accepted that a stomatal NH3 compensation point exists that determines, at a given atmospheric NH3 concentration, whether NH3 will be taken up by, or released from stomata (e.g. Farquhar et al., 1980; Lemon and Van Houtte, 1980). The NH3 compensation point is thought to be determined by the pH and the NH4+ concentration of gas exchange sites in the foliar apoplast (e.g. Farquhar et al., 1980; Husted and Schjoerring, 1995). As a consequence, there has recently been interest in measuring the pH and NH4+ concentration of the foliar apoplast as a simpler method of determining the NH3 compensation point than carrying out accurate measurements of gas exchange with foliage, which are considerably more time consuming. Apoplast pH has been measured by a variety of methods including the use of pH sensitive microelectrodes (Bowling and Edwards, 1984; Okazaki et al., 1995; Hanstein and Felle, 1999), equilibration with solutions (e.g. Raven and Farquhar, 1989; Pearson et al., 1998), and pH-sensitive fluorescent dyes (Pfanz and Deitz, 1987; Mühling et al., 1995). The use of pH-sensitive dyes, although not allowing the calculation of NH3 compensation points, has provided data that suggest spatial heterogeneity of pH within the foliar apoplast.
Hanstein and Felle (1999) attempted to measure apoplast NH4+ concentration in intact leaves using a microelectrode inserted through a stoma, but encountered problems of sensitivity and interference from K+. Extraction of the foliar apoplast by pressure vessel (Jachetta et al., 1986; Hartung et al., 1988) or by infiltration of leaves and extraction by centrifugation (e.g. Husted and Schjoerring, 1995; Hanstein et al., 1999) allows pH and NH4+ concentration of extracted liquids to be measured. Only the infiltration and centrifugation extraction method has been used for the prediction of NH3 compensation points to date.
It is important for estimation of the magnitude and direction of NH3 fluxes that the NH3 compensation point be determined accurately. This investigation made a comparison between the determination of NH3 compensation points for a widespread plant of semi-natural habitats (Luzula sylvatica [Huds.] Gaud.) by calculation from the pH and NH4+ concentration of apoplast extracts and by direct measurement of gas exchange. For the first time an evaluation of possible causes of discrepancies between compensation points determined by the two methods (e.g. Mattsson et al., 1997; Mattsson and Schjoerring, 1999) was made. Evaluation included the important issue of apoplast buffering, which has been assumed to be zero in previous determinations of NH3 compensation points.
RESULTS AND DISCUSSION
Cytoplasmic Contamination
Comparison of the malate dehydrogenase activity of total leaf homogenates and apoplast extracts showed a mean cytoplasmic contamination of apoplast extracts of 0.14% with a maximum value of 0.86% when the homogenate of lowest activity was compared with the apoplast extract of highest activity.
Influence of Indigo Carmine on the pH and NH4+ Analysis of Extracts
No significant difference (P ≤ 0.05) was found when the pH of apoplast extracts collected after infiltration with deionized water was compared by t test (assuming unequal variance, Microsoft Excel) with those collected after infiltration with 50 μm indigo carmine.
The presence of indigo carmine in standard NH4+ solutions had no systematic effect on the analysis of the NH4+ concentration (data not shown).
Osmotica
The five methods for estimating the osmolality of the foliar symplast gave widely varying estimates (Table I). It was considered that freezing caused the greatest cell disruption, although there was no statistical difference between boiling and freezing determinations (t test, P ≤ 0.05).
Table I.
Summary of osmolality determinations for L. sylvatica by different techniques
| Extraction Technique | Osmolality |
|---|---|
| mosmol kg−1 | |
| Grinding frozen tissue before elution in deionized water | 453 ± 6.3 (n = 12) |
| Grinding frozen tissue before elution in deionized water with added Triton X-100 | 435 ± 2.8 (n = 23) |
| Squeezing fresh leaves in a vise | 72 ± 1.8 (n = 6) |
| Freezing leaves in syringe prior to squeezing out liquid | 558 ± 4.5 (n = 13) |
| Boiling leaves in syringe prior to squeezing out liquid | 548 ± 2.3 (n = 18) |
Values are ±se, with no. of replicates in parentheses.
Estimates of the intercellular air space volume (Table II) were not significantly different when compared by single factor analysis of variance (P ≤ 0.05, Microsoft Excel). The estimate derived via infiltration with silicone fluid was the lowest. This may have been influenced by the difficulty of achieving complete infiltration of L. sylvatica leaves with the silicone fluid.
Table II.
Estimates of the intercellular gas space and NH4+ concentration of apoplast extracts of leaves of L. sylvatica, determined by infiltration with different liquids
| Liquid Used for Infiltration | Estimate of Mean Volume of Intercellular Gas Spaces | NH4+ Concentration of Apoplast Extract |
|---|---|---|
| mL g−1 fresh wt | mm | |
| Deionized water | 0.490 ± 0.004 (n = 10) | n/ma |
| Silicone fluid | 0.463 ± 0.004 (n = 10) | n/m |
| 91 g L−1d-Mannitol | 0.479 ± 0.002 (n = 10) | 0.040 ± 0.006 (n = 10) |
| 50 g L−1d-Mannitol | n/m | 0.046 ± 0.008 (n = 8) |
All values ± se with number of samples shown in parentheses.
n/m, Not measured.
The NH4+ concentration of apoplast extracts was not significantly different (t test, P ≤ 0.05) between those infiltrated with 50 g L−1 mannitol and those infiltrated with 91 g L−1 mannitol, although the non-significant difference between the means (Table II) was in the direction that would be expected if either the 50 g L−1 solution caused a flux of water from apoplast to symplast, or the 91 g L−1 solution caused a flux of water from the symplast to the apoplast.
As mentioned earlier, the uncertainty in the methods for symplastic osmolality determination and the lack of statistically significant differences in other tests made it preferable to use low osmolality solutions for infiltration.
Comparison of NH3 Compensation Points Determined by Gas Exchange and Bioassay
Values for the NH3 compensation point as determined by gas exchange measurements were among the lowest yet reported. Values of the NH3 compensation point calculated from the pH and NH4+ concentration of apoplast extracts were the lowest yet reported (Table III). Other reported controlled gas exchange compensation point measurements (25°C) range from 0.05 μg m−3 (1 μg NH3 m−3 is approximately 1.5 nmol NH3 mol−1 air at 1 atm and 25°C) for Arrhenatherum elatius (Hanstein et al., 1999), which is exceptionally low as compared with other gas exchange determinations, to >26 μg m−3 for wheat (Morgan and Parton, 1989). Reported compensation points calculated from apoplast extracts range from 0.04 μg m−3 for low-nitrogen maize (Loubet et al., 1999) to 36.2 μg m−3 for high-nitrogen barley (Mattsson et al., 1998). In general, non-crop plants have been found to have lower NH3 compensation points than crop plants (Morgan and Parton, 1989; Husted and Schjoerring, 1996; Husted et al., 1997; e.g. Hanstein et al., 1999; Mattsson and Schjoerring, 1999), although compensation point values (25°C) such as 0.3 for μg m−3 low-nitrogen oilseed rape (Husted and Schjoerring, 1996; Schjoerring et al., 1998a), 5.9 μg m−3 for Calluna vulgaris (Schjoerring et al., 1998b), and 0.03 μg m−3 for maize (Loubet et al., 1999) are notable exceptions. Higher NH3 compensation points in crop plants may reflect their adaptation to situations where nitrogen availability is high.
Table III.
Comparison of values of the NH3 compensation point and Γ as determined by gas-exchange measurements and apoplast extraction
| Nitrogen Treatment | Apoplast
Extraction
|
Gas Exchange
|
||||
|---|---|---|---|---|---|---|
| n | NH4+/H+ (Γ) | Calculated compensation point at 25°C | n | NH4+/H+ (Γ) | Compensation point at 25°C | |
| μg m−3 | ||||||
| kg N ha−1 y−1 as NH4Cl or NH4NO3 | ||||||
| 2 NH4Cl | 11 | 3.5 | 0.02 ± 0.01 | 2 | 105.5 | 0.7 ± 0.4 |
| 20 NH4Cl | 26 | 9.7 | 0.07 ± 0.01 | 3 | 85.3 | 0.6 ± 0.1 |
| 50 NH4Cl | 16 | 25.4 | 0.18 ± 0.06 | 2 | 96.9 | 0.7 ± 0.1 |
| 100 NH4Cl | 12 | 3.2 | 0.022 ± 0.004 | 3 | 94.0 | 0.6 ± 0.2 |
| 200 NH4Cl | 32 | 78.4 | 0.5 ± 0.1 | 6 | 159.0 | 1.1 ± 0.1 |
| 2 NH4NO3 | 8 | 2.5 | 0.017 ± 0.005 | 1 | 73.7 | 0.51 |
| 20 NH4NO3 | 5 | 24.0 | 0.17 ± 0.04 | 1 | 92.5 | 0.64 |
| 50 NH4NO3 | 18 | 4.9 | 0.03 ± 0.02 | 1 | 85.3 | 0.59 |
| 100 NH4NO3 | 10 | 14.5 | 0.10 ± 0.05 | 1 | 82.4 | 0.57 |
| 200 NH4NO3 | 26 | 20.8 | 0.14 ± 0.03 | 2 | 135.9 | 0.94 ± 0.09 |
| Overall mean | 18.7 | 0.129 | 101.1 | 0.70 | ||
Where appropriate, errors are ses.
A marked discrepancy was found between values of the NH3 compensation point and Γ determined by gas exchange measurements and those determined from apoplast extracts (Table III; Fig. 1). NH3 compensation points determined via gas exchange were between 2- and 30-fold higher than those calculated from apoplast extracts, with a mean difference of 5-fold. Such a discrepancy between compensation point values determined by the two methods can also be seen in the data of Mattsson et al. (1997) and Mattsson and Schjoerring (1999), with discrepancies ranging from 1.5- to 7-fold. In a similar manner, such a difference can be inferred from the data of Hermann et al. (1999), which shows sustained emission fluxes of NH3, measured by micrometeorological methods, during periods when the calculated NH3 compensation point was approximately 8-fold lower than the atmospheric NH3 concentration. Sutton et al. (1999) drew attention to apoplast extract-based (Loubet et al., 1999) and micrometeorological determinations (Milford et al., 1999) of the NH3 compensation point for a fertilized grassland in Southern Scotland that again show lower NH3 compensation points calculated from bioassays than would be predicted from gaseous flux measurements. Husted and Schjoerring (1995, 1996) reported good agreement between compensation points determined by the two methods, whereas Hanstein et al. (1999) found compensation points calculated from apoplast bioassays to be 3- to 8-fold larger than those determined from gas exchange measurements. However, Hanstein et al. (1999) suggested that the discrepancy was caused by coelution of amines with NH4+ in ion chromatograph measurements leading to an overestimation of apoplast NH4+ concentration.
Figure 1.

Comparison of values of Γ (NH4+/H+) determined by gas exchange and by apoplast extraction. AC, Ammonium chloride; AN, ammonium nitrate.
Given that a discrepancy has been found between the two methods of determination in all investigations other than those of Husted and Schjoerring (1995, 1996) on oilseed rape, it appears likely that a flaw exists in the methodology or the interpretation of results. Two basic differences exist in the procedure used by Husted and Schjoerring (1995, 1996) for the calculation of NH3 compensation points and that used here, (a) the correction for dilution of H+ during the extraction procedure, and (b) the correction of the NH4+ dissociation constant (Kd) for the ionic strength of the apoplast. However, in both cases, utilization of the methods of Husted and Schjoerring (1995, 1996) leads to lower estimates of the NH3 compensation point. Furthermore, the change in pH during the dilution of apoplast extracts from L. sylvatica suggested that the apoplast was sufficiently buffered to eliminate the need for correction of H+ concentration for dilution up to a factor of at least 2.5-fold (Fig. 2). The data of Nielsen and Schjoerring (1998) suggests that the pH of the apoplast of oilseed rape is similarly regulated and/or buffered.
Figure 2.

Change in measured H+ concentration of an example apoplast extract with dilution (solid line) compared with the calculated H+ concentration if the extract were unbuffered (broken line).
Correction of Kd for the ionic strength of the apoplast gives estimates of the NH3 compensation point and Γ approximately 0.9 of those presented here assuming an apoplast ionic strength of 20 mm calculated on the basis of measured ion concentrations (Table IV) and a predicted negative charge of 4.9 mm. Given that the unquantified negative charge was “added” as a monovalent species, the ionic strength of the apoplast may in reality be greater than 20 mm, and the calculated NH3 compensation points lower than 0.9 of the values presented here.
Table IV.
Concentrations of inorganic ions and ion-forming organic species in the apoplast of L. sylvatica
| Species | Concentration |
|---|---|
| mm | |
| Malate | 0.287 ± 0.044 (n = 30) |
| Citrate | 0.009 ± 0.001 (n = 25) |
| Lactate | 0.077 ± 0.009 (n = 29) |
| Phosphate | 0.002 ± 0.0002 (n = 12) |
| Asn | 0.194 ± 0.013 (n = 30) |
| Na+ | 2.1 ± 0.4 (n = 24) |
| K+ | 5.2 ± 0.4 (n = 24) |
| Mg2+ | 1.4 ± 0.2 (n = 24) |
| Ca2+ | 2.5 ± 0.4 (n = 24) |
| NH4+ | 0.029 ± 0.019 (n = 129) |
| SO42− | 0.28 ± 0.02 (n = 24) |
| Cl− | 9 ± 1 (n = 24) |
Values are ±se with number of samples in parentheses.
Infiltration of leaves with solutions of differing pH also suggested that the apoplast was subject to almost complete homeostasis with respect to H+ concentration within the range of perturbations carried out here (Fig. 3). The change of the H+ concentration of solutions after infiltration was almost 1:1 correlated with the departure of the infiltrated solution from the apoplast H+ determined by infiltration with unadjusted indigo carmine solution (see gradient of measured data regression line, Fig. 3). Approximate homeostasis of the apoplast with respect to pH is likely to be due to apoplast buffering and transplasmalemma fluxes. Nielsen and Schjoerring (1998), in similar infiltrations of oilseed rape leaves, found the pH of the apoplast to be regulated. They considered the regulation only in terms of transplasmalemma fluxes of H+ rather than buffering, although the data presented here (Fig. 2) clearly show that buffering occurs in the apoplast of L. sylvatica. This is consistent with the reports of apoplast buffering made by Böttger et al. (1980), Pfanz and Dietz (1987), Ikoma and Okamoto (1988), Peters and Felle (1991), and Hanstein and Felle (1999).
Figure 3.

Comparison of the departure of the H+ concentration of the infiltrated solution from that of the apoplast, with the difference of H+ concentration between the infiltrated solution and the extract ([H+]infiltration solution − [H+]extract) (solid line). The H+ concentration of the apoplast was assumed to be that of extractions performed with unadjusted indigo carmine solution. Values are means of 10 values ± se. The broken line shows the theoretical values based on simple mixing of the apoplast with the infiltration solution.
Buffering would also be expected from the presence in the foliar apoplast of organic acids undergoing dissociation or protonation at physiological values of apoplast pH e.g. citrate, and malate (Table IV; Speer and Kaiser, 1991; Lohaus et al., 1995; Mühling and Sattelmacher, 1995; Gabriel and Kesselmeier, 1999).
Homeostasis of the Apoplast with Respect to NH4+ Concentration
Infiltration of leaves with solutions of differing NH4+ concentration, although suggesting that NH4+ is transported from the foliar apoplast to the symplast, did not show complete homeostasis of the apoplast with respect to NH4+ concentration in L. sylvatica within the approximate 4 min between infiltration and centrifugation (Fig. 4). However, Nielsen and Schjoerring (1998) reported that the NH4+ concentration of the apoplast of oilseed rape was upwardly regulated to a value of 0.8 mm within 1.5 min after infiltration with an NH4+-free solution. Such a rapid upward regulation was not taking place in leaves of L. sylvatica. Figure 4 shows that infiltration with a solution approximately equal to the NH4+ concentration of the apoplast extract collected after infiltration with NH4+-free indigo carmine solution caused an increase in the NH4+ concentration of the resulting extract. It therefore appears that, despite the evidence for uptake of NH4+ from the apoplast, a correction for the dilution of NH4+ in the apoplast of L. sylvatica during the extraction procedure is necessary when calculating NH3 compensation points.
Figure 4.

Variation in the NH4+ concentration of extracts with the NH4+ concentration of the solution infiltrated into leaves. The broken line shows the theoretical NH4+ concentration of extracts if simple mixing was the only process involved. For this the concentration of the extract after infiltration with deionized water and correction for dilution was used as the apoplast NH4+ concentration. Values are means of 10 values ± se Two regression lines are shown to allow for possible curvature in the plot. The linear regression is significant at P < 0.001. The curvilinear regression is a second order polynomial.
Errors in the Determination of NH3 Compensation Points by Gas-Exchange Measurements
Gas-exchange measurements, being the more empirical method of compensation point determination, should theoretically give the most reliable estimates. No obvious sources of error existed in these measurements other than possible NH3 exchange between the plant growing medium and the atmosphere. In control experiments with artificial plants measurable exchange of NH3 did not appear to be taking place with the growing medium of experimental plants.
Cytoplasmic Contamination of Extracts
Cytoplasmic contamination of extracts is not a likely explanation for the observed discrepancy in the determinations of the NH3 compensation points since such contamination would be likely to increase the pH and the NH4+ concentration of apoplast extracts. Thus, calculated NH3 compensation points would probably be overestimated.
Use of Osmotica in Infiltration Solutions
Assuming that infiltration of leaves with low osmolality solutions caused a flux of water from the apoplast to the symplast, it would be likely that indigo carmine concentrations would be higher than in the absence of such fluxes. Correction for dilution would therefore underestimate the proportion of the extract that was apoplastic in origin, and increase the factor by which NH4+ concentrations were multiplied during correction for dilution. Again this would lead to an overestimation of the apoplast NH4+ concentration and the NH3 compensation point.
Loss of Indigo Carmine within the Leaf
If indigo carmine were adsorbed by surfaces within the leaf, there would be a tendency to overestimate the proportion of extracts that were of apoplastic origin. This could lead to an underestimate of the factor by which extract NH4+ concentrations were multiplied to correct for dilution during extraction, and thus underestimation of the NH3 compensation point. The use of indigo carmine in infiltrations to determine apoplast liquid volumes has precedent in the literature (Cosgrove and Cleland, 1983; Husted and Schjoerring, 1995, 1996; Husted et al., 1997; Mattsson et al., 1997; Mattsson et al., 1998; Nielsen and Schjoerring, 1998; Hanstein et al., 1999; Mattsson and Schjoerring, 1999). Furthermore, results have been validated by Husted and Schjoerring (1995) who compared results from indigo carmine infiltrations with [14C]sorbitol infiltrations and found good agreement between the two approaches. They did, however, find 2% of the indigo carmine in a 6-μm solution thereof adsorbed to isolated cell walls of oilseed rape. Such an adsorption would be insufficient to account for the discrepancy between NH3 compensation points determined by gas exchange and those calculated from apoplast extractions. Cosgrove and Cleland (1983) also found good agreement between apoplast liquid volume determined by indigo carmine and [14C]mannitol in stems of pea and soybean, but suspected that some indigo carmine might be lost during infiltrations of cucumber. It should be noted that Cosgrove and Cleland (1983) reported averages of only two determinations with no indication of variation and that there is, as they point out, no guarantee that the [14C]mannitol determination was not responsible for the observed discrepancy.
It is possible that there may have been significant loss of indigo carmine during the infiltration and extraction process, although assuming apoplast pH was correct, the magnitude of the indigo carmine loss would have needed to be great to account for such large discrepancies in compensation points determined by the two methods used here. For instance, in a hypothetical extraction a 50-μm solution of indigo carmine was infiltrated into leaves and resulted in an extract indigo carmine concentration of 15 μm. If the concentration of indigo carmine in the extract without any loss should have been 25 μm, the correct calculated compensation point would only be 1.4 times higher than that calculated from the extract where dye loss had occurred. Therefore, although such loss of indigo carmine during the extraction process would lead to an underestimation of calculated NH3 compensation points, it does not seem possible that it could have made more than a contribution to observed discrepancies.
Error in the Determination of Apoplast pH
Small errors in the determination of apoplast pH could potentially lead to much larger errors in calculated NH3 compensation points due to the logarithmic relationship between pH and H+ concentration. However, it is clear that a large margin of error in the measurement of apoplast pH (e.g. 0.3–1.5 pH units) would be necessary to account for the magnitude of the discrepancies between the two methods of compensation point determination.
Rust Infection of Experimental Plants
Rust infection of experimental plants may have had an influence on the magnitude of the NH3 compensation point. However, documented effects of rust infection suggest that the NH3 compensation points of infected plants would be higher than those of uninfected plants. Tetlow and Farrar (1993) reported an alkalization of the apoplast of rust-infected barley plants as compared with controls, and Fukuda (1996) noted a similar extracellular alkalization from cultured cells in response to a fungal elicitor. Fungal infection has been linked to an increase in phenylalanine ammonia-lyase activity (Habereder et al., 1989; Hahlbrock and Scheel, 1989; Walter, 1989), which is one of the principal NH3-generating enzymes in plants (Raven et al., 1992). Higher NH4+ generation rates are consistent with reports of increased foliar NH4+ concentrations (Farkas and Király, 1961) and increased volatilization of NH3 (Jenkyn and Finney, 1984), which also suggest an increase in the NH3 compensation point in response to biotrophic infection.
Within Leaf Spatial Variability of Apoplast pH and NH4+ Concentration
Apoplast extractions have a number of sources of error. The principal ones have been dealt with above and appear to be quantitatively too small or liable to result in calculated NH3 compensation points becoming lower. However, it is not possible to ever be completely sure what the origin of the “apoplast” extract really is. The extraction technique has been widely used with a range of validation techniques (Cosgrove and Cleland, 1983; Husted and Schjoerring, 1995) and available data indicate that extracted liquid is apoplastic in origin. A major consideration in the application of this technique to the calculation of NH3 compensation points is whether or not extracted liquid is representative of liquid in the apoplast at leaf gas exchange sites. Hoffmann and Kosegarten (1995) found spatial differences in the apoplast of leaf cells of sunflower to be up to 1 pH unit. Bowling and Edwards (1984) report differences in apoplast pH of Tradescantia virginiana between guard cell and epidermal cell of 1.4 pH units in the dark, although in the light, gradients were much smaller. However, even these large reported pH gradients are insufficient to account for the largest discrepancies found between NH3 compensation points determined by the two different approaches (i.e. a ratio of 30). A difference in pH of 0.7 unit is, however, sufficient to account for the mean 5-fold discrepancy. However, the pH of bulk foliar apoplast extracts is unlikely to represent one extreme of spatial apoplast pH variability, but rather some intermediate value.
It is possible that there may be gradients of pH radially through the cell wall due to solute transport and volatilization. The existence of such gradients in plant cell walls is yet to be demonstrated, but such a possibility merits further investigation if a suitable measurement technique could be developed.
NH4+ concentration in the foliar apoplast is unlikely to be spatially uniform, but no investigation of its variation has been carried out to date. Indeed it is doubtful whether a suitable technique for its investigation is currently available. However, reports and speculation on the evaporative concentration of solutes at the end of the transpiration stream such as Canny (1995), would suggest that the apoplast at transpiration sites could be enriched in NH4+ relative to the bulk of the foliar apoplast. Furthermore, such an accumulation of solutes may not be uniform over the leaf but is thought to be greater at leaf margins (Wilson et al., 1991). Consequently extracts of the foliar apoplast would be likely to underestimate the NH4+ concentration at transpiration sites, and thus the NH3 compensation point.
CONCLUSION
NH3 compensation points for L. sylvatica are low compared with values in the literature when determined by either gas exchange measurements or the pH and NH4+ concentration of apoplast extracts. This is consistent with other measurements that suggest that wild species generally have lower NH3 compensation points than crop plants. The marked discrepancy between compensation point values determined by the two discussed methods is not readily explained, since most conceivable errors are liable to result in overestimates of compensation points, or are of insufficient magnitude. In the absence of other data, spatial variability of pH and/or NH4+ concentration within the foliar apoplast appears the most promising area for further investigation.
MATERIALS AND METHODS
Experimental Plants
Wild-growing Luzula sylvatica (Huds.) Gaud. plants were collected from Carlops (Scottish Borders, UK; UK Ordnance Survey grid reference NT179567) and potted singly in a 4:1 vermiculite:perlite mix. Plants were grown in a greenhouse at a minimum temperature of 10°C under ambient light supplemented with mercury vapor lamps during winter months to increase the photoperiod to 16 h day−1. Artificial light intensity was approximately 400 μmol photons m−2 s−1 at canopy height. Plants received Long-Ashton nutrient solution (Hewitt and Smith, 1974) concentrations of 0.214, 2.14, 5.35, 10.7, or 21.4 mm N as NH4Cl or NH4NO3 at a rate of 20 mL pot−1 week−1 for approximately 8 months prior to use in experiments. This rate of nitrogen addition was equivalent to 2, 20, 50, 100, and 200 kg N ha−1 y−1 based on a growing medium surface area of 156 cm2.
During growth, plants were found to be infected with the rust Puccinia luzulae Libert., which is widespread in wild-growing L. sylvatica.
Typical leaf areas ranged from 300 to 1,250 cm2 pot−1 depending upon nitrogen treatment.
Gas-Exchange Measurements
Measurements of the exchange of NH3, water vapor, and carbon dioxide with plants were carried out in a purpose-built dynamic cuvette facility (Fig. 5) contained within a controlled temperature room.
Figure 5.

Schematic diagram of gas-exchange measurement system.
Experimental plants were placed inside a 0.216-m3 Perspex cuvette. Pots were placed in polythene bags secured tightly around the base of the foliage with cable ties. Light was supplied with a 250 W discharge lamp at a rate of approximately 400 μmol photons m−2 s−1 at canopy height after passing through a glass water bath to reduce heating effects. A control experiment was performed, replacing experimental plants with artificial Chlorophytum sp. (Philip Display, Edinburgh) of similar morphology to quantify any gaseous exchange between the growth medium and the cuvette atmosphere. Experiments using artificial plants or an empty cuvette also showed there to be no line loss of ammonia once stable conditions had been reached.
During experiments plants were fed and watered as necessary with a syringe through a 1.6-mm outside diameter Teflon tube sealed inside the plastic bags covering pots.
Air entering (30 L min−1) and leaving the cuvette was continually sampled for NH3 content with an AMANDA NH3 analysis system (Wyers et al., 1993; ECN, Petten, The Netherlands) and was sampled for water vapor and carbon dioxide content on a 10-min cycle. Water vapor and carbon dioxide were measured with a temperature and humidity probe (Rotronic Instruments, Horley, Surrey, UK) and an infrared gas analyzer (type 255 mk3, ADC, Hoddesdon, Hertsfordshire, UK), respectively. Air temperature and humidity within the cuvette were continuously monitored using a probe (Vaisala, Cambridge, UK) and leaf temperature was measured using six type-T thermocouples attached to the underside of leaves with paper clips in such a way as to hold the junction against the leaf surface.
Prior to entering the cuvette, the air stream was dried by passing through copper coils submerged in a water bath at 4°C with condensed liquid being captured in a flask. Adjustment of humidity was carried out by bubbling a portion of the air stream through deionized water. Relative humidity was maintained at 50% ± 10%. NOx was removed by passing air through Purafil and activated charcoal, and NH3 was removed by passing air through oxalic acid suspended on rockwool. Particles were captured on a depth filter (Pall Biomedical, Portsmouth, Hampshire, UK).
Generation of NH3 was achieved by introducing, with a peristaltic pump, small volumes of dilute NH3 solution into a heated flask through which the air stream passed.
Determination of NH3 Compensation Points
NH3 compensation points were determined from the linear regression equation of NH3 flux against ambient NH3 concentration where the ambient NH3 concentration was considered to be that of air entering the cuvette. Figure 6 shows a typical plot.
Figure 6.

Variation of NH3 flux to and from plant foliage with concentration of ambient NH3. Positive fluxes represent emission from foliage and negative fluxes represent deposition to foliage. Values are means of 11 values from the most stable portions of AMANDA data. Error bars are ses of 11 values. Plants used in this case received 200 kg N ha−1 y−1 as ammonium nitrate, but the plot is typical of those used to determine compensation points for all treatments.
Apoplast Extraction
To prepare apoplast extracts, leaves were detached from plants, washed in deionized water, dried with tissues (Kimberly Clark, Aylesford, Kent, UK), and infiltrated with 50 μm indigo carmine solution (4°C) under alternate vacuum and pressure in a 50-mL syringe. Leaves were again dried and rolled perpendicular to the long axis before they were placed in a 15-mL syringe-type serial pipettor tip barrel (Boeringer-Mannheim, Lewes, East Sussex, UK). Pipettor tip and leaves were then placed in a 50-mL centrifuge tube with a 1.5-mL microcentrifuge tube at the bottom to collect liquid expelled from the leaf. The assembly was centrifuged at 2,000g for 4 min at 4°C.
NH4+ and pH Measurement
The pH of extracts collected during centrifugation was measured with an InLab 423 semi-micro electrode (Mettler Toledo, Udorf, Switzerland) inserted in the microcentrifuge tube. Calibration was only with low ionic strength standards (Russell pH, Auchtermuchty, Fife, UK). NH4+ concentration was assessed in an AMFIA NH4+ flow injection analyzer (ECN). Extracts of less than 100 μL were diluted to the minimum analyzable volume of 100 μL with deionized water prior to NH4+ analysis.
Cytoplasmic Contamination
Cytoplasmic contamination of apoplast extracts was assessed by comparison of malate dehydrogenase activity (EC 1.1.1.37) in apoplast extracts and leaf homogenates. Apoplast extracts for assay were prepared as above excepting that a buffer (0.1 m N-tris[hydroxymethyl]methyl-2-aminoethanesulphonic acid, 2 mm dithiothreitol, and 0.2 mm EDTA) was substituted for indigo carmine solution and leaves were weighed before and after infiltration. Leaf homogenates were prepared by grinding tissue in liquid nitrogen before diluting a weighed portion with the aforementioned buffer. Malate dehydrogenase activity was assessed by adding 30 μL of extract to 1 mL of a solution composed of 0.05 m Tris, 0.1 mm NADH, and 0.4 mm oxalacetate, and measuring absorbance decrease at 340 nm (20°C) in a Shimadzu UV-160A spectrophotometer (Shimadzu, Kyoto).
Assessment of Apoplast Dilution
Dilution of the apoplast liquid during the extraction procedure was assessed by measuring the concentration of indigo carmine in extracts spectrophotometrically at 610 nm. Since dilution was somewhat variable, indigo carmine was utilized in all extracts (excepting those used to assess cytoplasmic contamination) to eliminate errors due to the use of a mean dilution correction, as has been the case in previous studies (e.g. Husted and Schjoerring, 1995). Furthermore, the assessment of apoplast dilution in each sample eliminates interpretation problems resulting from the incomplete infiltration of leaves.
The influence of indigo carmine on pH analysis was tested by twice comparing 10 extracts from leaves infiltrated with deionized water and 10 of those infiltrated with 50 μm indigo carmine.
Standard NH4+ solutions with 0 to 200 μm indigo carmine were analyzed to test for any influence of indigo carmine on NH4+ analysis.
Osmotica
An estimate of the osmolality of the symplast of L. sylvatica was made by using several different methods to produce extracts for analysis in an osmometer (model 5100C vapor pressure osmometer, Wescor, Logan, UT). (a) Leaves were frozen in liquid nitrogen and ground in a pestle and mortar. A weighed portion of the ground leaves was allowed to equilibrate with deionized water overnight in a sealed microcentrifuge tube at 4°C. Following equilibration the sample was centrifuged at 10,000g for 6 min before measuring the osmolality of the supernatant. (b) The above technique was modified by equilibrating ground tissue in deionized water with 1 drop of Triton X-100 50 mL−1 to further disrupt cell membranes. Correction was made for the influence of the Triton X-100 on the osmolality of the extract. (c) A tight roll of leaves was squeezed in a vise and the osmolality of the expelled liquid measured. (d) A roll of leaves was placed in a 5-mL syringe that was sealed with a blocked needle hub. The syringe was placed in liquid nitrogen and liquid was expelled with the syringe plunger after thawing. (d) A variation of the above technique was carried out in which cells were disrupted by boiling rather than freezing. Sealed syringes were immersed in boiling water for 3 min and cooled before expelling liquid from leaves with the syringe plunger.
As a consequence of the widely varying estimates of osmolality resulting from the above methods (see “Results”), osmotica were not used in this study during infiltration. It was considered that because the osmolality of the symplast could not be accurately measured, the effect of infiltrating with osmotica was less predictable than that of infiltrating a well-hydrated leaf with a low osmolality solution, relying on cell turgor to prevent significant transplasmalemma water fluxes.
To test the potential for water to move from the apoplast to the symplast during infiltration with low osmolality solutions, a comparison was made between determinations of the foliar intercellular air space volume when infiltrating with different solutions. Leaves were weighed before and after infiltration with deionized water, high-viscosity silicone fluid (5 cs, Dow-Corning, Poole, Dorset, UK), which theoretically cannot cross the plasmalemma (Cosgrove and Cleland, 1983), and 91 g L−1 d-mannitol solution that was theoretically isoosmotic (see “Results”) with the symplast. Account was taken of the density of the infiltrated solutions.
A further comparison was made of the NH4+ concentration of apoplast extracts after infiltration with either 50 or 91 g L−1 d-mannitol solution.
Apoplast Buffering and pH Regulation
To assess the necessity for correction of apoplast hydrogen ion concentration for dilution during the extraction procedure, dilutions with 50 μm indigo carmine were carried out on apoplast extracts while monitoring pH changes.
Further investigation of apoplast pH regulation was carried out by infiltrating leaves from plants receiving 200 kg N ha−1 year−1 as NH4Cl with 50 μm indigo carmine solution adjusted to pH 3, 4, 7, or 10 with HCl or NaOH. The pH of resulting extracts was compared with the theoretical values based on simple mixing of infiltrated solution and apoplast liquid. To calculate the expected pH of extracts based on simple mixing, it was assumed that the extracts obtained after infiltration with unadjusted indigo carmine solution (pH 7) gave a reasonable estimate of apoplast pH, i.e. that apoplast buffering was sufficient to prevent significant perturbation of apoplast pH during infiltration with a neutral, unbuffered solution.
Regulation of Apoplast NH4+ Concentration
As for the investigation of apoplast pH regulation, leaves from plants receiving the 21.4 mm N NH4Cl nutrient solution were infiltrated with 50 μm indigo carmine solution adjusted to 0, 0.2, 0.4, 0.6, 0.8, 1.0, 1.5, and 2.0 mg NH4+ L−1 with (NH4)2SO4. Again the NH4+ concentrations of extracts were compared with those predicted from simple mixing, assuming that the extracts obtained following infiltration with unadjusted indigo carmine solution, and correction for dilution, gave a reasonable estimate of apoplast NH4+ concentration.
Calculation of NH3 Compensation Points
NH3 compensation points were calculated from apoplast pH and NH4+ concentration according to the equilibria applied by Sutton et al. (1994) using the simplified numerical fit of Nemitz (1998):
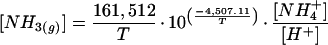 |
1 |
where [NH3(g)] is the NH3 compensation point in M, and T is temperature in Kelvin.
The temperature-independent analog for the NH3 compensation point, Γ (the ratio of apoplast NH4+ concentration to apoplast H+ concentration; Sutton et al.,1996) was also used as a method of comparison.
Ionic Composition of the Apoplast
Apoplast extracts prepared as above were analyzed for inorganic ion content on an Alltech Odyssey high performance ion chromatography system (Alltech, Deerfield, IL) with either an Alltech Universal Cation column (3 mm methane sulfonic acid eluent) or an Alltech Universal anion column (A-15; 0.86 mm Na2CO3/0.34 mm NaHCO3 with 5% [w/v] methanol, eluent). Na+ in extracts due to the presence of indigo carmine was subtracted from measured values.
Similar extracts were prepared for organic ion analysis. Extracts were dried in a Heto CT 110 centrifugal drier (Heto-Holten A/S, Allerod, Denmark), dissolved in pyridine, and derivatized in N-tert-butyldimethylsilyl-N-methyltrifluoroacetamide at 70°C for 30 min. Derivatized samples were then analyzed in a VG Organic MD800 gas chromatograph-mass spectrometer (Fisons, Manchester, UK). Quantification of chemical species was carried out by comparison with standards.
A simple computer model based on measured ionic concentrations and organic ion dissociation constants was used to predict the concentration of unquantified negative charge needed to predict a physiologically realistic apoplast pH and the dissociation state of organic ions and NH4+ (Hill, 1999). In the model, changes in pH with increasing dilution were modeled from the concentrations of pH-active species and their dissociation constants at 20°C or 25°C (Weast, 1987; Budavari, 1989) and concentration of inorganic ions. The calculations were derived on the principle of charge balance and the function was solved numerically by iteration using the Newton-Raphson approximation (Burden and Faires, 1993). Concentrations of charged forms of dissociating species were calculated from their dissociation constants, total concentrations, and H+ concentrations. The function was of the form:
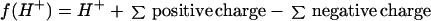 |
2 |
Addition of unquantified negative charge was probably necessary due to a greater proportion of positive ions being identified and quantified than negative ions.
ACKNOWLEDGMENTS
Thanks to Dr. John Beardall for inorganic ion analysis, Robert Hardy for allowing experimental plants to be gathered from his land, Frank Harvey and Colin McBeath for gathering experimental plants, Anna-Laure Corriol for indigo carmine tests, Dr. Richard Parsons and Ewan Kier for assistance with GCMS analysis, and Professor Graham Farquhar for suggestions on the accumulation of NH4+ at transpiration termini.
Footnotes
This work was supported by a Cooperative Awards in Sciences of the Environment Studentship (no. GT4/95/64/T) from the Natural Environment Research Council and funds from the UK Department of Environment Transport and Regions (UMBRELLA project).
LITERATURE CITED
- Böttger M, Soll H-J, Gasché A. Modification of the external pH by maize coleoptiles and velamen radicum of Vanilla planifolia ANDR. Z Pflanzenphysiol. 1980;99:89–93. [Google Scholar]
- Bowling DJF, Edwards A. pH gradients in the stomatal complex of Tradescantia virginiana. J Exp Bot. 1984;35:1641–1645. [Google Scholar]
- Budavari S, editor. The Merck Index. Rathway, NJ: Merck; 1989. [Google Scholar]
- Burden RL, Faires DJ. Numerical Analysis. Boston, MA: PWS Publishing; 1993. pp. 56–69. [Google Scholar]
- Canny MJ. Apoplastic water and solute movement: new rules for an old space. Annu Rev Plant Physiol Plant Mol Biol. 1995;46:215–236. [Google Scholar]
- Cosgrove DJ, Cleland RE. Solutes in the free space of growing stem tissues. Plant Physiol. 1983;72:326–331. doi: 10.1104/pp.72.2.326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farkas GL, Király Z. Amide metabolism in wheat leaves infected with stem rust. Physiol Plant. 1961;14:344–353. [Google Scholar]
- Farquhar GD, Firth PM, Wetselaar R, Weir B. On the gaseous exchange of ammonia between leaves and the environment: determination of the ammonia compensation point. Plant Physiol. 1980;66:710–714. doi: 10.1104/pp.66.4.710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Flechard CR. Turbulent exchange of ammonia above vegetation. PhD thesis. Nottingham, UK: University of Nottingham; 1998. [Google Scholar]
- Fukuda Y. Coordinated activation of chitinase genes and extracellular alkalinization in suspension-cultured tobacco cells. Biosci Biotechnol Biochem. 1996;60:2005–2010. [Google Scholar]
- Gabriel R, Kesselmeier J. Apoplastic solute concentrations of organic acids and mineral nutrients in the leaves of several Fagaceae. Plant Cell Physiol. 1999;40:604–612. [Google Scholar]
- Habereder H, Schröder G, Ebel J. Rapid induction of phenylalanine ammonia-lyase and chalcone synthase mRNAs during fungus infection of soybean (Glycine max L.) roots or elicitor treatment of soybean cell cultures at the onset of phytoalexin synthesis. Planta. 1989;177:58–65. doi: 10.1007/BF00392154. [DOI] [PubMed] [Google Scholar]
- Hahlbrock K, Scheel D. Physiology and molecular biology of phenylpropanoid metabolism. Annu Rev Plant Physiol Plant Mol Biol. 1989;40:347–369. [Google Scholar]
- Hanstein S, Felle HH. The influence of atmospheric NH3 on the apoplastic pH of green leaves: a non-invasive approach with pH-sensitive microelectrodes. New Phytol. 1999;143:333–338. [Google Scholar]
- Hanstein S, Mattsson M, Jaeger H-J, Schjoerring JK. Uptake and utilisation of atmospheric ammonia in three native Poaceae species: leaf conductances, composition of apoplastic solution and interactions with root nitrogen supply. New Phytol. 1999;141:71–83. [Google Scholar]
- Hartung W, Radin J, Hendrix DL. Abscisic acid movement into the apoplastic solution of water-stressed cotton leaves: role of apoplastic pH. Plant Physiol. 1988;86:908–913. doi: 10.1104/pp.86.3.908. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heil GW, Diemont WH. Raised nutrient levels change heathlands into grasslands. Vegetation. 1983;53:113–120. [Google Scholar]
- Hermann B, Blatter A, Jones S, Hinderling-Schwob J, Fuhrer J, Neftel A, Feller U, Hartwig UA. Influence of symbiotic nitrogen fixation on NH3 exchange between a grass/clover crop and the atmosphere at two levels of N application: an experimental approach. Program and Abstracts, 10th. KVL, Copenhagen: Nitrogen Workshop; 1999. , II.42. [Google Scholar]
- Hewitt EJ, Smith TA. Plant Mineral Nutrition. London: English Universities Press; 1974. [Google Scholar]
- Hill PW. Physiological aspects of the exchange of gaseous ammonia between Luzula sylvatica (Huds.) Gaud. and the atmosphere. PhD thesis. Dundee, UK: University of Dundee; 1999. [Google Scholar]
- Hoffmann B, Kosegarten H. FITC-dextran for measuring apoplast pH and apoplastic pH gradients between various cell types in sunflower leaves. Physiol Plant. 1995;95:327–335. [Google Scholar]
- Husted S, Mattsson M, Schjoerring JK. Ammonia compensation points in two cultivars of Hordeum vulgare L. during vegetative and generative growth. Plant Cell Environ. 1997;19:1299–1306. [Google Scholar]
- Husted S, Schjoerring JK. Apoplastic pH and ammonium concentration in leaves of Brassica napus L. Plant Physiol. 1995;109:1453–1460. doi: 10.1104/pp.109.4.1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Husted S, Schjoerring JK. Ammonia flux between oilseed rape plants and the atmosphere in response to changes in leaf temperature, light intensity, and air humidity. Plant Physiol. 1996;112:67–74. doi: 10.1104/pp.112.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikoma S, Okamoto H. The quantitative and chronological relationship between IAA-induced H+-pump activation and elongation growth studied by means of xylem perfusion. Plant Cell Physiol. 1988;29:261–267. [Google Scholar]
- Jachetta JJ, Appleby AP, Boersma L. Use of the pressure vessel to measure concentrations of solutes in apoplastic and membrane-filtered symplastic sap in sunflower leaves. Plant Physiol. 1986;82:995–999. doi: 10.1104/pp.82.4.995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jenkyn JF, Finney ME. Experiments to examine the significance of ammonia evolution from barley seedlings infected with the powdery mildew fungus, Erysiphe graminis f.sp. hordei. J Agric Sci Camb. 1984;102:679–685. [Google Scholar]
- Leith ID, Hicks WK, Fowler D, Woodin SJ. Differential responses of UK upland plants to nitrogen deposition. New Phytol. 1999;141:277–289. doi: 10.1046/j.1469-8137.1999.00333.x. [DOI] [PubMed] [Google Scholar]
- Lemon E, Van Houtte R. Ammonia exchange at the land surface. Agron J. 1980;72:876–883. [Google Scholar]
- Lohaus G, Winter H, Reins B, Heldt HW. Further studies of the phloem loading process in leaves of barley and spinach: the comparison of metabolite concentrations in the apoplastic compartment with those in the cytosolic compartment and in the sieve tubes. Bot Acta. 1995;108:270–275. [Google Scholar]
- Loubet B, Hill PW, Sutton MA, Tang YS, Cellier P. Apoplastic and bulk ammonium and pH in leaves of Lolium perenne L. and Zea mays L.: effects of management and fertilisation. Report. Paris: Institut National de la Recherche Agronomique Thiveral Grignon; 1999. /Institute of Terrestrial Ecology, Edinburgh Research Station, Edinburgh. [Google Scholar]
- Mattsson M, Häusler RE, Leegood RC, Lea PJ, Schjoerring JK. Leaf-atmosphere NH3 exchange in barley mutants with reduced activities of glutamine synthetase. Plant Physiol. 1997;114:1307–1312. doi: 10.1104/pp.114.4.1307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mattsson M, Husted S, Schjoerring JK. Influence of nitrogen nutrition and metabolism on ammonia volatilisation in plants. Nutr Cycling Agroecosyst. 1998;32:491–498. [Google Scholar]
- Mattsson M, Schjoerring JK. Controlled environment measurements of ammonia exchange in two grass species. Program and abstracts, 10th. KVL, Copenhagen: Nitrogen Workshop; 1999. , II.46. [Google Scholar]
- Milford C, Sutton MA, Theobald MR, Nemitz E. Fluxes of ammonia with an intensively managed cut grassland in Southern Scotland. Program and Abstracts, 10th. KVL, Copenhagen: Nitrogen Workshop; 1999. , II.47. [Google Scholar]
- Morgan JA, Parton WJ. Characteristics of ammonia volatilisation from spring wheat. Crop Sci. 1989;29:726–731. [Google Scholar]
- Mühling KH, Pleith C, Hansen U-P, Sattelmacher B. Apoplastic pH of intact leaves of Vicia faba as influenced by light. J Exp Bot. 1995;46:377–382. [Google Scholar]
- Mühling KH, Sattelmacher B. Apoplastic ion concentration of intact leaves of field bean (Vicia faba) as influenced by ammonium and nitrate nutrition. J Plant Physiol. 1995;147:81–86. [Google Scholar]
- Nemitz EG. Surface-atmosphere exchange of ammonia and chemically interacting species. PhD thesis. Manchester, UK: University of Manchester Institute of Science and Technology; 1998. [Google Scholar]
- Nielsen KH, Schjoerring JK. Regulation of apoplastic NH4+ concentration in leaves of oilseed rape. Plant Physiol. 1998;118:1361–1368. doi: 10.1104/pp.118.4.1361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Okazaki Y, Nishizaki Y, Iwazaki N. Effects of a pulse of blue light on the extracellular pH in the pulvinus of Phaseolus vulgaris L.: measurements with a double-barreled pH-sensitive electrode. Plant Cell Physiol. 1995;36:1131–1134. [Google Scholar]
- Pearson J, Clough ECM, Woodall J, Havill DC, Zhang Z-H. Ammonia emissions to the atmosphere from leaves of wild plants and Hordeum vulgare treated with methionine sulphoximine. New Phytol. 1998;138:37–48. [Google Scholar]
- Peters WS, Felle H. Control of apoplast pH in corn coleoptile segments: I. The endogenous regulation of cell wall pH. J Plant Physiol. 1991;137:655–661. [Google Scholar]
- Pfanz H, Deitz K-J. A fluorescence method for the determination of the apoplastic proton concentration in intact leaf tissues. J Plant Physiol. 1987;129:41–48. [Google Scholar]
- Raven JA. Acquisition of nitrogen by the shoots of land plants: its occurrence and implications for acid-base regulation. New Phytol. 1988;109:1–20. [Google Scholar]
- Raven JA, Farquhar GD. Leaf apoplast pH estimation in Phaseolus vulgaris. In: Dainty J, De Michelis MI, Marré E, Rasi-Caldogno F, editors. Plant Membrane Transport: The Current Position. Amsterdam: Elsevier; 1989. pp. 607–610. [Google Scholar]
- Raven JA, Wollenweber B, Handley LL. Ammonia and ammonium fluxes between photolithotrophs and the environment in relation to the global nitrogen cycle. New Phytol. 1992;121:5–18. [Google Scholar]
- Roelofs JGM. The effect of airborne sulphur and nitrogen deposition on aquatic and terrestrial heathland vegetation. Experientia. 1986;42:372–377. [Google Scholar]
- Schjoerring JK, Husted S, Mattsson M. Physiological parameters controlling plant-atmosphere ammonia exchange. Atmos Environ. 1998a;32:491–498. [Google Scholar]
- Schjoerring JK, Husted S, Poulsen MM. Soil-plant-atmosphere ammonia exchange associated with Calluna vulgaris and Deschampsia flexuosa. Atmos Environ. 1998b;32:507–512. [Google Scholar]
- Speer M, Kaiser WM. Ion relations of symplastic and apoplastic space in leaves from Spinacia oleracea L. and Pisum sativum L. under salinity. Plant Physiol. 1991;97:990–997. doi: 10.1104/pp.97.3.990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sutton MA, Asman WAH, Schjoerring JK. Dry deposition of reduced nitrogen. Tellus. 1994;46B:255–273. [Google Scholar]
- Sutton MA, Milford C, Nemitz E, Theobald MR, Hill PW, Fowler D, Schjoerring JK, Mattsson ME, Nielsen KH, Husted S, Erisman JW, Otjes R, Hensen A, Cellier P, Loubet B, David M, Generemont S, Neftel A, Blatter A, Hermann B, Jones SK, Horvath L, Sitkey J, Raso J, Huszar E, Mantanzas K, Koukoura Z, Gallagher M, Williams P, Riedo M. Institute of Terrestrial Ecology, Edinburgh Research Station, Edinburgh. 1999. First year science overview report GRAMINAE: grassland ammonia interactions across europe: experimental strategy and results from 1998–1999. Report. [Google Scholar]
- Sutton MA, Nemitz EG, Fowler D, Wyers GP, Otjes RP, San José R, Moreno J, Schjoerring JK, Husted S, Meixner FX, Ammann C, Neftel A, Gut A. Exchange of atmospheric ammonia with European ecosystems (EXAMINE): final report to the European Commission. Vol. 1. Edinburgh: Institute of Terrestrial Ecology; 1996. [Google Scholar]
- Sutton MA, Pitcairn CER, Fowler D. The exchange of ammonia between the atmosphere and plant communities. Adv Ecol Res. 1993;24:301–393. [Google Scholar]
- Tetlow IJ, Farrar JF. Apoplastic sugar concentration and pH in barley leaves infected with brown rust. J Exp Bot. 1993;44:929–936. [Google Scholar]
- van Breemen N, Burrough PA, Velthorst EJ, van Dobben HF, de Wit T, Ridder TB, Reijuders HFR. Soil acidification from atmospheric ammonium sulphate in forest canopy throughfall. Nature. 1982;299:548–550. [Google Scholar]
- van Breemen N, Mulder J, Driscoll CT. Acidification and alkalinisation of soils. Plant Soil. 1983;75:283–308. [Google Scholar]
- Walter MH. The induction of phenylpropanoid biosynthetic enzymes by ultraviolet light or fungal elicitor in cultured parsley cells is overriden by a heat-shock treatment. Planta. 1989;177:1–8. doi: 10.1007/BF00392148. [DOI] [PubMed] [Google Scholar]
- Weast RC, editor. CRC Handbook of Chemistry and Physics. Boca Raton, FL: CRC Press; 1987. [Google Scholar]
- Wilson TP, Canny MJ, McCully ME. Leaf teeth, transpiration and the retrieval of apoplastic solutes in balsam poplar. Physiol Plant. 1991;83:225–232. [Google Scholar]
- Wyers GP, Otjes RP, Slanina J. A continuous-flow denuder for the measurement of ambient concentrations and surface-exchange fluxes of ammonia. Atmos Environ. 1993;27A:2085–2090. [Google Scholar]