

Abstract
Understanding how transcription factors (TFs) regulate mammalian gene expression in space and time is a central topic in biology. To activate a gene, a TF has first to diffuse in the available space of the nucleus until it reaches a target DNA sequence or protein (target site). This eventually results in the recruitment of the whole transcriptional machinery. All these processes take place in the mammalian nucleoplasm, a highly organized and dynamic environment, in which some complexes transiently assemble and break apart, whereas others appear more stable. This diversity of dynamic behaviors arises from the number of biomolecules that make up the nucleoplasm and their pairwise interactions. Indeed, interactions energies that span several orders of magnitude, from covalent bounds to transient and dynamic interactions, can shape nuclear landscapes. Thus, the nuclear environment determines how frequently and how fast a TF contacts its target site, and it indirectly regulates gene expression. How exactly transient interactions are involved in the regulation of TF diffusion is unclear, but are reflected by live cell imaging techniques, including single-particle tracking (SPT). Overall, the macroscopic result of these microscopic interactions is almost always anomalous diffusion, a phenomenon widely studied and modeled. Here, we review the connections between the anomalous diffusion of a TF observed by SPT and the microscopic organization of the nucleus, including recently described topologically associated domains and dynamic phase-separated compartments. We propose that anomalous diffusion found in SPT data result from weak and transient interactions with dynamic nuclear substructures, and that SPT data analysis would benefit from a better description of such structures.
Keywords: molecular interactions, nuclear transport, protein motion, transcription factor
Introduction
Mammalian gene expression and its regulation take place in the nucleus, a highly complex and subcompartmented organelle. Interactions strengths between nuclear constituents span several orders of magnitude, from covalent bounds to ‘strong’ noncovalent interactions. These interactions lead to the formation of macromolecular structures, either stable (Figure 1, left; for instance double-stranded DNA or biochemically purifiable macromolecular complexes such as the ones involved in gene expression) or transient but specific, leading to preferential associations of classes of proteins (Figure 1, right).
Figure 1. Diversity of weak interactions and example techniques to study them.

Biological interactions cover a wide spectrum in terms of complex stability (a) and number of molecules involved (b). This spectrum spans from stable protein complexes that can be purified and further imaged by techniques such as X-ray diffraction (left) to very labile, transient interactions that can involve thousands of proteins in vivo, whereas none of the interactions can be captured by traditional biochemistry (right). As one goes from one end of the spectrum to the other end, distinct sets of techniques (c) and types of representations (d) are needed. For instance, as the valency of interactions increases from a few strongly interacting partners to many weakly interacting partners, new graphical representations are needed, since traditional schematics representing macromolecular complexes whose stoichiometry is known as the juxtaposition of monomers (center) become difficult to read when depicting one protein weakly interacting with dozen of partners (right). In that case, matrices of pairwise interactions between proteins A-I might be more relevant. SAXS: Small-Angle X-Rays Scattering, NMR: Nuclear Magnetic Resonance.
The regulation of transcription is of utmost interest as it is central not only to developmental biology, but also to cancer biology, drug screening, etc. To express a mRNA, a macromolecular complex constituted of several subunits and dozens of proteins, the preinitiation complex, is first to assemble at the promoter of a gene in a time- and space-specific manner [1]. This complex is able to robustly integrate transient signals such as the ones mediated by proteins’ binding to cis-regulatory sequences such as enhancers [2].
Glossary
anomalous diffusion a phenomenon occurring when a molecule explores over time a volume lower than predicted by Brownian diffusion. It is usually characterized by a sublinear growth of the mean squared displacement (MSD) as a function of time (MSD(t) ∼ tα with α < 1), with α the anomalous diffusion exponent.
compact exploration universal mode of diffusion in which the exploration of the diffusing molecule is local and distance-dependent and a given site is explored repeatedly over time, in a highly recurrent manner. Within the compact exploration mode, several strengths of compaction can be distinguished.
facilitated diffusion biophysical phenomena allowing a molecule to find its target faster than predicted by traditional, 3D free diffusion. This includes diffusion on a surface of reduced dimensionality such as DNA.
fractal kinetics type of kinetic reactions happening within a reactor that is not well-stirred. This notably includes most reactions happening on a surface of reduced dimensionality. Fractal kinetics are characterized by a progressive segregation between reagents and products. The kinetic rate k of the reaction is time-dependent.
fractals structure exhibiting (statistical) self-similarity, that the (statistical properties of the) structure re main similar at various zoom levels. Fractals can be described by their (potentially non-integer) fractal dimension df . The fractal dimension of an object describes its space-filling property. Many types of biological objects exhibit fractal properties, such as the branching pattern of the lung, or the hierarchical folding of DNA in the nucleus.
free diffusion (also termed Brownian diffusion) chracterizes the motion of a particle in a fluid arising from thermal agitation only. Seen as a “null” model in SPT.
non-compact exploration universal mode of diffusion in which the exploration is global, and every site on the structure has a constant probability to be explored (distance independence).
phase separation a state of matter in which part of the soluble protein fraction segregates into a liquid or liquid-like droplet.
surface of reduced dimensionality an object whose fractal dimension df < 3, meaning that it exhibits some properties similar from the ones of 1D or 2D structures.
weak interactions (in this review) interactions that are usually too short-lived to be captured by traditional biochemistry techniques, that typically involve one or several wash step, during which proteins interacting specifically but transiently get diluted and washed out.
More mechanistically, the assembly of such a complex can be characterized by a set of chemical reactions describing the progressive recruitment of factors and subunits. A kinetic rate can be associated to each of these reactions. Traditional biochemistry and in vitro experiments have been the methods of choice to investigate such complex processes. Most biochemical techniques involve purification steps allowing to reveal strong, noncovalent interactions such as the ones occurring in a stably assembled complex [3,4] (Figure 1, left). Then, further quantification of stoichiometry and affinity constants became possible, progressively building a network of interacting proteins, usually represented as a graph with nodes linked with arrows.
Within this framework, the understanding of gene expression regulation reduces to elucidating how external factors [including transcription factors (TFs)] affect the kinetic constants . Although it can be assumed that kinetic rates are characterized only by the nature and concentration of enzyme, substrate, and cofactors, it was shown in 1906 by von Smoluchowski [5] that the kinetic rate of a well-mixed, diffusion-limited reaction can be decomposed as . Thus, the kinetic rate is a function of both the cross-section of interaction (reflecting the chemical properties of the partners and usually studied by biochemical approaches) and the diffusion constant of the species.
Since is determined by the local environment, this finding is striking in the context of gene expression regulation: now the kinetics of one reaction depend on the whole nuclear structure. More specifically, any factor that affects diffusion in any specific or nonspecific way will ultimately influence reaction rates. Indeed, interactions resulting in facilitated diffusion on a substructure (such as a TF on DNA, [6–8]) or (transient) segregation inside a membrane-less compartment in a phase-separated manner [9,10] can all be seen under the unifying framework of diffusion on a surface of reduced dimensionality. Diffusion on surfaces of reduced dimensionality yields kinetics that are qualitatively different from in free, 3D diffusion and leads to potentially dramatically increased reaction rates [11].
Anomalous diffusion, a phenomenon occurring when a molecule explores a volume lower than predicted by diffusion, affects all proteins inside a cell. Numerous physical models can describe anomalous diffusion [12], and several have been applied to the motion of nuclear proteins. However, many of them only provide a phenomenological description of diffusion, rather than mechanistic insights, and radically distinct models can often fit the available data equally well.
In light of these considerations, it is worthwhile to examine recent discoveries describing either stable subnuclear compartments or their more transient, weak-interactions-induced counterparts to highlight their influence on the diffusion of factors through dimensionality reduction. This includes TADs (topologically associated domains), lamina-associated domains, nucleoli, noncoding RNAs, transcription factories, phase-separated domains, etc. They constitute substructures with a high valency amenable to weak interactions that can qualitatively influence diffusion and target search.
Here, we first review anomalous diffusion models applied to protein motion and relate them to a potential physical generative model. Then, we emphasize recent advances in the characterization of regions of reduced dimensionality in mammalian nuclei, both aspecific through volume exclusion and specific through transient, weak-but-specific interactions. Finally, we propose that these weak interactions shape TF dynamics, and that single-particle tracking (SPT) analysis would greatly benefit from a better understanding of the pairwise interaction map between nuclear proteins.
Most anomalous diffusion models reflect underlying networks of weak interactions
The technique of choice to investigate protein motion in the nucleus of live cells is light microscopy of fluorescently tagged proteins. Different imaging and modeling modalities have provided significant insights, including fluorescence recovery after photobleaching (FRAP), fluorescence correlation microscopy (FCS) or SPT. In this review, we focus on SPT, because it directly provides access to the dynamics of single diffusing molecules. In a SPT experiment, a small subset of the proteins of interest are imaged and tracked over short times at the resolution of a few tens of nanometers, allowing to resolve isolated single molecules even within clusters of high densities.
In solution, the diffusion coefficient of a protein is inversely proportional to the hydrodynamic radius of the protein () and the viscosity of the medium (η) through the Stokes–Einstein relationship , where reflects thermal agitation, with the Boltzmann constant and the absolute temperature. This description, however, is too simplistic in the complex cellular environment. Indeed, with the exception of inert tracers of small molecular mass [13,14], it is well acknowledged that (a) macromolecules in a cell diffuse much slower than in a medium of comparable viscosity, (b) that complexes of high molecular mass can diffuse faster than small proteins, and (c) that most molecules exhibit anomalous diffusion.
Thus, the diffusion of TFs cannot be described by simple friction/viscosity relationships, and their behavior, perhaps unsurprisingly, has to be seen from the angle of transient interactions with a dense matrix of interactants. In the context of this review, we define transient (or ‘weak’) interactions as interactions that are usually too short-lived to be captured by traditional biochemistry techniques that typically involve one or several wash steps, during which proteins interacting specifically but transiently get diluted and washed out.
Furthermore, diffusion of many factors does not follow free, Brownian diffusion (Figure 2a,c). Such diffusion is termed anomalous (more specifically, subdiffusive), meaning that the space explored over time by one factor is lower than expected by free diffusion (reviewed in [12,15]). Anomalous diffusion is usually characterized by a sublinear growth of the mean squared displacement (MSD) as a function of time (that is, MSD with ; Figure 2a) and α is called the anomalous diffusion exponent. This anomalous diffusion exponent does not fully characterizes the type of diffusion and additional metrics, such as the distribution of translocation angles (Figure 2b), provide valuable information about the dynamics. Anomalous diffusion of proteins has been fitted with success by phenomenological anomalous diffusion models, and include fractional Brownian motion (Figure 2d) [16–19], continuous-time random walks (CTRW; Figure 2e) [20,21] and diffusion in fractal media (Figure 2f) [22–24]. Although useful as phenomenological descriptions, these models are often agnostic regarding the underlying reality of the process. In any case, the explanation of diffusion has to rely on physics and chemistry of the nucleus.
Figure 2. Models of anomalous diffusion and plausible underlying physical structures.

(a and b) Characterizations of anomalous diffusion. (a) Sub-linear mean-square displacement plotted as a function of time characterizes subdiffusion, and reflects how a diffusing particle explores space, the degree of anomalous diffusion is characterized by the exponent α, the lower the α the more subdiffusive the process. (b) Free diffusion is characterized by isotropic distribution of angles — subpanel (i) — whereas an anisotropic distribution indicates anomalous diffusion — subpanels (ii) and (iii). (c) 3D free diffusion (), as usually encountered in a homogenous media (). (d–f) Several types of heterogenous media can yield anomalous diffusion, including (d) diffusion within a viscoelastic polymer, in which a protein “bounces against” an elastic structure, a process traditionally described by fractional Brownian motion (fBm) and (e) free diffusion interspersed by long binding times — red stars, a process called CTRW, and (f) diffusion within a fractal media, that is a space obstructed by obstacles of all sizes.
From a physical perspective, proteins can adsorb and diffuse on nuclear substructures. When this happens, the exploration properties of the protein are universally given by two parameters: first, the dimension of the random walk (deduced from the anomalous diffusion exponent: , i.e. the scaling of the MSD: ), and second, the dimension of the space available to diffuse . can be integer ( for instance for sliding on DNA without jumps, for a factor freely diffusing on the surface of a subcompartment), or noninteger, a feature that characterizes self-similar structures, that is, fractals (Figure 2f). For instance, highly obstructed media or dense environments where the available volume is reduced to small pores are often accurately described by their noninteger fractal dimension . Such a case can occur in dense subcompartments such as phase-separated domains. For the sake of this review, we will denote structures of as structures of reduced dimensionality.
Depending on and , the motion of the protein falls into two universal categories, termed compact and noncompact [25–27]. In a compact exploration (), exploration is local and distance-dependent and a given site is explored repeatedly over time, in a highly recurrent manner. Conversely, in a noncompact exploration (), the exploration is global, and every site on the structure has a constant probability to be explored (distance independence); the exploration is nonrecurrent (transient). For instance, a particle freely diffusing has a of 2. When diffusion takes place in a 3D space () the particle tends not to revisit sites, adopting a noncompact exploration (indeed, ). Conversely, a particle in free, Brownian diffusion () constrained to diffuse in 1D (; hypothetically along a DNA fiber) will repeatedly sample the same sites (compact exploration, ). Consequently, target search times are decreased and reaction rates are increased in the compact case.
Structures of reduced dimensionality, including fractals, emerge naturally from various processes, including diffusion-limited aggregation and hierarchical assembly of macromolecular scaffolds, such as the multi-scale organization of chromatin. The goal of the next sections is to highlight a few structures of reduced dimensionality in the nucleus and how they influence kinetics of TFs.
Steric hindrance in the nucleus
Far from constituting a homogeneous medium, the nucleus is a highly organized and subcompartmentalized organelle. The main organizing structure, chromatin, constitutes ∼10–30% of the nuclear volume [28,29] and probably accounts for a significant part of the diffusion slowdown [30]. Since every molecule has to slalom around a dense and heterogeneous chromatin environment, diffusion is impaired. Note that, however, similar diffusion coefficients are usually observed in the cytoplasm and the nucleoplasm [31], suggesting that protein crowding can also account for diffusion slowdown ([32,33] and [23] for a discussion).
Over the past years, organizing principles of chromatin have emerged: at large scale, the genome is segregated in chromosome territories and regions of heterochromatin/euchromatin, lamina-associated domains at the periphery and nucleoli lying more at the center. At higher magnification, chromatin is organized in areas of preferential interactions such as A/B compartments and TADs that reflect the functional organization of chromatin [34].
Overall, although highly heterogeneous, chromatin in the mammalian nucleus is well described by a self-similar, fractal structure that occupies a nonzero volume. This was initially postulated [35], and later evidenced by spectroscopic [36,37], genomic [38,39] and imaging techniques [18,23,40–42].
As a consequence, factors diffusing in the available volume are constrained by this structure [43], possibly experiencing diffusion in a medium of reduced dimensionality, as evidenced by numerous reports [23,44]. In a model where only volume exclusion happens, proteins of the same size and shape should have the same diffusion coefficient. Thus, the embedding structure of the nucleus only sets a lower bound on the level of anomalous diffusion that can be observed.
Several lines of argument point to the fact that although steric hindrance induces a significant decrease in the diffusion coefficient (), its effect on anomalous diffusion (α) is mild for proteins of weight <150 kDa (that is, about the weight of a histone octamer wrapped with 150 bp DNA). Indeed, FRAP experiments performed with protein or nonprotein tracers of increasing molecular mass suggest that low molecular mass tracers diffuse almost freely in the nucleus, allowing to infer a viscosity close to the one of water [13]. At molecular mass >150 kDa, anomalous diffusion becomes more prominent [14,23,31], sometimes leading to particles occasionally being trapped in the chromatin mesh. This effect is consistent with the relatively limited volume occupied by chromatin [28,29] that can trap big complexes but not smaller proteins.
In conclusion, although volume exclusion by chromatin and other nuclear constituents is real, it affects all proteins of the same size in a similar manner. In contrast, a protein weakly but specifically interacting with such a structure (for instance, TFs sliding/hopping on DNA [6,45–48]) will immediately show a much higher level of anomalous diffusion. Furthermore, even without considering a fractal structure, simple dimensionality reduction to 1D or 2D can yield nontraditional kinetics termed fractal kinetics [11,49,50]. Fractal kinetics are characterized by a time-dependent reaction rate: for a reaction, the reaction rate can be expressed as , with that depends on the (fractal) dimension of the space. Fractal kinetics arise when the reaction cannot be assumed to be well stirred, which is the case of diffusion on surfaces of reduced dimensionality. For instance, fractal kinetics in 2D could occur by weak interaction with the nuclear lamina, Figure 2b. All in all, weak and transient interactions shape the nuclear landscape and can give rise to emergent structures and properties, as exemplified in the next section.
Weak interactions in the nucleus
Unlike inert tracers whose diffusion is only determined by volume exclusion, proteins have both a relevant shape and charge pattern that determine their interaction landscape and thus their diffusive properties. These noncovalent interactions are obviously crucial to form biochemically stable complexes such as the transcription preinitiation complex or the spliceosome (Figure 1, left), but also to form dynamic emergent structures of reduced dimensionality upon which TFs can transiently adsorb and diffuse (Figure 1, right). Under this model, proteins do not form stable complexes anymore, but rather have a high number of weakly interacting partners. Indeed, simulation studies have shown that phase separation yielding structures of reduced dimensionality can occur under very minimal hypotheses, such as weak overall protein-protein attraction [51,52]. In this context, the traditional representation of protein-protein interaction networks as graphs and arrows becomes less informative as the protein network gets fully connected, and can be replaced by representations such as pairwise interaction matrices (Figure 1d, right) [53]. Furthermore, the list of proteins exhibiting phase separation (that is, a state of matter in which part of the soluble protein fraction segregates into a liquid or liquid-like droplet) in vitro or in vivo is quickly growing, supporting the vision that the emergence of structures of reduced dimensionality is closer to a general organizing principle than an anecdotal biophysical phenomenon, and some of them were linked with transcriptional regulation [9,54–57].
First, transient structures of reduced dimensionality, including protein aggregates, phase separated domains or subnuclear compartments require at least one multivalent partner that can nucleate the aggregation. As such, many abundant constituents of the mammalian nucleus have been shown to nucleate a structure of reduced dimensionality. These structures include the formation of nucleoli in which diffusion is highly constrained, phase-separation of heterochromatin protein 1 domains (HP1; [9,10]) and highly active chromatin domains, whose center has reduced accessibility to diffusing proteins, restricting most diffusing proteins to the surface of the domain [57]. The constituents include low complexity protein domains that constitute the majority of the mammalian proteome [58,59], especially TFs [60], repeated DNA [61] or RNA sequences [62–64] or small amphiphilic molecules [65,66]. Once nucleated, these structures can have a highly tortuous, potentially fractal architecture, and can serve as a scaffold that can weakly trap other proteins, leading to anomalous diffusion.
Second, the partners have to exhibit compatible interactions: it is chemically unlikely that both highly charged and hydrophobic proteins will coexist in the same structure without the help of additional compounds acting as counterions [67], setting the basis of a ‘grammar of interactions’ [68], that is being progressively deciphered [56,66,69–72].
Third, structures of reduced dimensionality emerging from weak interactions exhibit the following properties: (1) they usually exist as an extremely dynamic equilibrium rather than a stable structure [10,64,73], and can thus be at the same time prevalent in the nucleus and hard to purify by traditional biochemistry that preferentially capture stable interactions. (2) Moreover, they emerge from a dynamic mesh of pairwise chemical interactions. They can show a high level of specificity, and several structures of reduced dimensionality can coexist in the same nucleus without intermixing [55,57,61,67,74]. Furthermore, the number and spatial relationships of such structures is only limited by the combinatorics of chemical interactions. (3) Finally, these structures can be regulated by the well-studied post-translational machinery of eukaryotic cells. For instance, phosphorylation of one of the proteins involved in such structure can trigger the timely disassembly of the whole structure and free all the factors interacting with it [9,55,75–77]. All those factors will then exhibit a dramatically different dynamics and target search properties, potentially switching from a compact exploration mode to a noncompact one. As such, a specific (and potentially functional) group of factors can be regulated at once by modulation of the post-translational modifications of one scaffolding protein [61,63].
The characterization of structures of reduced dimensionality emerging from weak interactions is still in its infancy, but appears more and more strongly as a clear organizing principle of mammalian nuclei. These structures create the matrix upon which fast-diffusing factors can specifically and transiently bind, diffuse and unbind, thus dynamically shaping the ‘diffusion landscape’ of the whole transcriptional machinery.
Even though live imaging approaches specifically characterize the behavior of one single factor, they are blind to all these substructures. Indeed, SPT reflects the dynamics of proteins transiently interacting with those structures of reduced dimensionality and one TF potentially visits several of them in the span of a few tens of milliseconds [78]. Such complex behavior therefore appears macroscopically as various kinds of anomalous diffusion.
Perspectives: seeing beyond the dots
In a complex mammalian nucleus, the diffusion of a TF is ruled by transient interactions with underlying structures of reduced dimensionality. From a more general perspective, the question arises of how gene expression regulation processes relate to the multiplicity of structures of reduced dimensionality?
Proteins often harbor several domains (Figure 3), holding the potential to interact successively and repeatedly with multiple classes of structures of reduced dimensionality. Thus, depending on its interaction domains, a TF will ‘see’ a different landscape and will interact with some structures, whereas other factors will either be excluded or cross them without any additional interactions than limited steric hindrance. In this respect, the nucleus can be described as a ‘multiverse’, in which some factors coexist in the same physical space but exhibit radically distinct dynamics and interactions (Figure 3).
Figure 3. Weak interactions of individual protein domains shape TF dynamics.
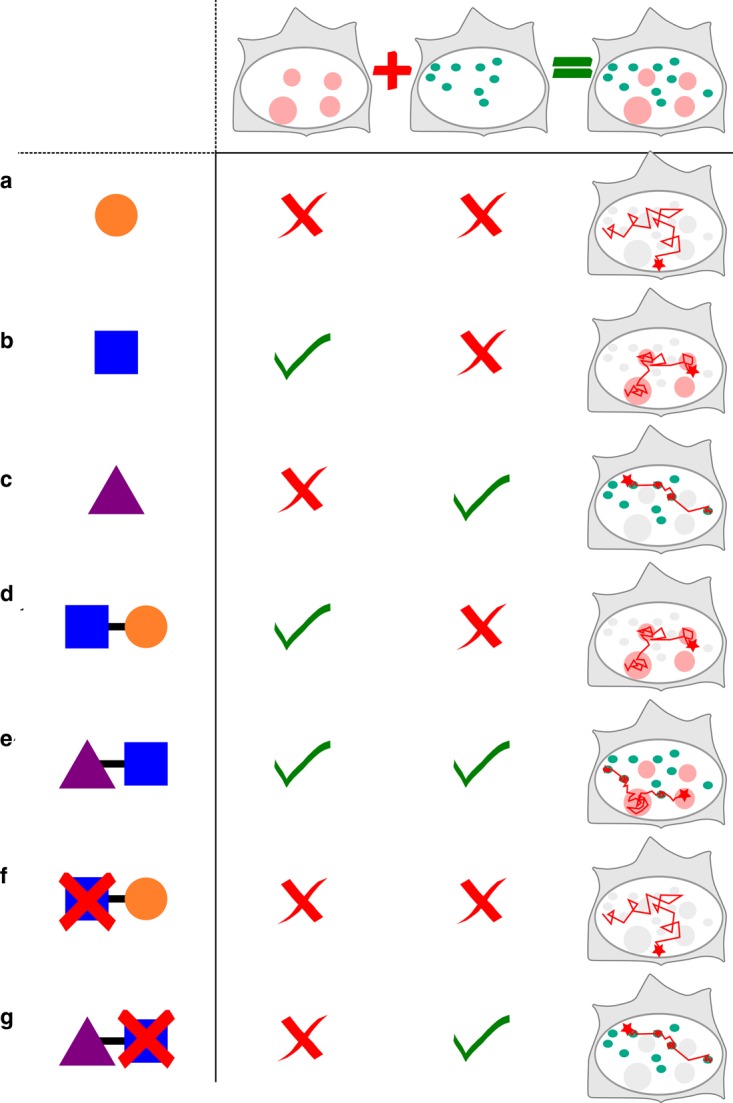
(a)–(c) Individual protein domains have specific transient interactions: the round domain (a) does not interact with any particular structure (represented by the two columns of the table), (b) the square domain interacts with a given pink structure (first column) and (c) the triangle domain interacts with the green structures (second column). This results in domain-specific dynamics (third column). (d and e) When protein domains are combined within a protein or TF, the observed SPT is a mixture between the interactions of each single domain. (f and g) When individual domains are mutated, the protein loses some transient specific interactions, and its dynamics can dramatically change (compare panels e and g).
Furthermore, structures of reduced dimensionality have been proved to be functionally relevant. For example, the dynamic and regulated switching of a TF between structures of reduced dimensionality determines its dynamics (Figure 3a–e) and function. It has been shown that TF exhibit radically different dynamics before/after a post-translational modification [79,80], or an artificial deletion of a domain (Figure 3f–g), [8,81–85]. In that case, the observed diffusion will be arising from the remaining interactions from the other interaction domains, or ultimately from simple volume exclusion [86].
Although theoretical and experimental support for the importance of weak interactions as an architectural principle of the nucleus and gene expression regulation is being actively investigated, several questions remain unaddressed:
First, how many distinct types of structures of reduced dimensionality exist? Since the numbers of types of low-complexity domains are likely to be limited, one can expect that a limited number of such structures actually coexist at a given time in a nucleus [71]. This implies that the SPT dynamics of TFs will fall in a limited number of categories, which in turn is determined by their combinatorial interactions with one or several of these structures. To take into consideration such processes paves the way for a higher-order understanding of gene expression regulation and key transitions occurring, for instance, during mitosis or development.
Second, can we determine the pairwise interaction matrix between low-complexity protein domains, which would allow us to derive predictive dynamics of a given TF modification? Ideally, such matrix will encompass all known low-complexity domains, but also abundant multivalent RNAs and DNA sequences, and each element of this matrix will reflect the affinity between two domains under physiological conditions (Figure 1d).
Third, how much detail is required to describe these structures of reduced dimensionality? Is the pairwise interaction between protein domains a good approximation of the properties of the nucleus? Conversely, one can imagine substructures of reduced dimensionality arising from interactions more complex than simple pairwise-interactions. Indeed, it is widely known that cooperativity plays a role in the assembly of many more or less stable macromolecular structures [87], including some phase-separated domains [67].
Fourth, how do key biological transitions such as differentiation intertwine with these structures of reduced dimensionality? In a similar way, as pluripotency or cell-type specific TF networks have been identified, can pluripotency or cell-type specific structures of reduced dimensionality be evidenced, integrating the expression levels of TFs and providing a framework to better understand such key processes?
To answer those questions, our understanding of nuclear processes need to be drastically expanded. Hitherto, a dynamic picture of spatially segregated factors, together with their interaction matrix, is currently missing. Promising tools to access those parameters include quantitative FRET [88], in cell NMR [89–91], low-photons SPT [92], tracking FCS [93], spatially resolved FCS [94] and computational methods [72,95].
Conclusion
Although the so far identified key players in gene expression regulation are biochemically stable complexes that can be purified using traditional methods, increasing evidence suggest that higher-order, weaker-interaction structures, acting as structures of reduced dimensionality, play a central role in transcriptional regulation. They do so by providing a remarkably versatile way of specifically and timely regulating TF target search dynamics and thus gene expression. All in all, it appears that the functional properties of the nucleus are shaped by not only stable macromolecular complexes but also transient structures (arising from a continuum of weak interactions that might seem spurious). In this context, the saying from Heraclitus makes probably more sense than ever: ‘The fairest order in the world is alike a heap of random sweepings’.
Acknowledgments
We apologize to our colleagues whose work could not be cited due to limited space. We thank Elena Rensen and Anders S. Hansen for their critical feedback on the manuscript. Work in the Darzacq laboratory is supported by National Institutes of Health (UO1-EB021236) and California Institute for Regenerative Medicine (LA1-08013). We are very thankful to the Sci-Hub database, without which the writing of this review would have been impossible. Photographs of Figure 1 are courtesy of Jeff Dahl, Vossman and Shasha Chong, respectively, from left to right.
Abbreviations
- CTRW
continuous-time random walks
- FCS
fluorescence correlation microscopy
- FRAP
fluorescence recovery after photobleaching
- MSD
mean squared displacement
- SPT
single-particle tracking
- TADs
topologically associated domains
- TF
transcription factor
Competing Interests
The Author declare that there are no competing interests associated with this manuscript.
References
- 1.Sainsbury S., Bernecky C. and Cramer P. (2015) Structural basis of transcription initiation by RNA polymerase II. Nat. Rev. Mol. Cell Biol. 16, 129–143 10.1038/nrm3952 [DOI] [PubMed] [Google Scholar]
- 2.Spitz F. (2016) Gene regulation at a distance: From remote enhancers to 3d regulatory ensembles. Semin. Cell Dev. Biol. 57, 57–67 10.1016/j.semcdb.2016.06.017 [DOI] [PubMed] [Google Scholar]
- 3.Louder R.K., He Y., López-Blanco J.R., Fang J., Chacón P. and Nogales E. (2016) Structure of promoter-bound TFIID and model of human pre-initiation complex assembly. Nature 531, 604–609 10.1038/nature17394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Murakami K., Tsai K.-L., Kalisman N., Bushnell D.A., Asturias F.J. and Kornberg R.D. (2015) Structure of an RNA polymerase II preinitiation complex. Proc. Natl Acad. Sci. U.S.A. 112, 13543–13548 10.1073/pnas.1518255112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.von Smoluchowski M. (1906) Zur kinetischen theorie der brownschen molekularbewegung und der suspensionen. Ann. Phys. 326, 756–780 10.1002/andp.19063261405 [DOI] [Google Scholar]
- 6.Hammar P., Leroy P., Mahmutovic A., Marklund E.G., Berg O.G. and Elf J. (2012) The lac repressor displays facilitated diffusion in living cells. Science 336, 1595–1598 [DOI] [PubMed] [Google Scholar]
- 7.Normanno D., Boudarène L., Dugast-Darzacq C., Chen J., Richter C., Proux F. et al. (2015) Probing the target search of DNA-binding proteins in mammalian cells using TetR as model searcher. Nat. Commun. 6, 7357 10.1038/ncomms8357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Hansen A.S., Pustova I., Cattoglio C., Tjian R. and Darzacq X. (2017) CTCF and cohesin regulate chromatin loop stability with distinct dynamics. eLife 6, e25776 10.7554/eLife.25776 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Larson A.G., Elnatan D., Keenen M.M., Trnka M.J., Johnston J.B., Burlingame A.L. et al. (2017) Liquid droplet formation by HP1α suggests a role for phase separation in heterochromatin. Nature 547, 236–240 10.1038/nature22822 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Strom A.R., Emelyanov A.V., Mir M., Fyodorov D.V., Darzacq X. and Karpen G.H. (2017) Phase separation drives heterochromatin domain formation. Nature 547, 241–245 10.1038/nature22989 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kopelman R. (1988) Fractal reaction kinetics. Science 241, 1620–1626 ISSN 0036-8075, 1095-9203 10.1126/science.241.4873.1620 [DOI] [PubMed] [Google Scholar]
- 12.Metzler R., Jeon J.-H., Cherstvy A.G. and Barkai E. (2014) Anomalous diffusion models and their properties: non-stationarity, non-ergodicity, and ageing at the centenary of single particle tracking. Phys. Chem. Chem. Phys. 16, 24128–24164 10.1039/C4CP03465A [DOI] [PubMed] [Google Scholar]
- 13.Seksek O., Biwersi J. and Verkman A.S. (1997) Translational diffusion of macromolecule-sized solutes in cytoplasm and nucleus. J. Cell Biol. 138, 131–142 PMID: [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Weiss M., Elsner M., Kartberg F. and Nilsson T. (2004) Anomalous subdiffusion is a measure for cytoplasmic crowding in living cells. Biophys. J. 87, 3518–3524 10.1529/biophysj.104.044263 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Höfling F. and Franosch T. (2013) Anomalous transport in the crowded world of biological cells. Rep. Prog. Phys. 76, 046602 10.1088/0034-4885/76/4/046602 [DOI] [PubMed] [Google Scholar]
- 16.Tejedor V., Bénichou O., Voituriez R., Jungmann R., Simmel F., Selhuber-Unkel C. et al. (2010) Quantitative analysis of single particle trajectories: mean maximal excursion method. Biophys. J. 98, 1364–1372 10.1016/j.bpj.2009.12.4282 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Guigas G. and Weiss M. (2008) Sampling the cell with anomalous diffusion–the discovery of slowness. Biophys. J. 94, 90–94 10.1529/biophysj.107.117044 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Shinkai S., Nozaki T., Maeshima K. and Togashi Y. (2016) Dynamic nucleosome movement provides structural information of topological chromatin domains in living human cells. PLoS Comput. Biol. 12, e1005136 10.1371/journal.pcbi.1005136 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ghosh R.P., Franklin M.J., Draper W.E., Shi Q. and Liphardt J.T. (2017) A fluorogenic array tag for temporally unlimited single molecule tracking. bioRxiv 10.1101/159004 [DOI] [PubMed] [Google Scholar]
- 20.Weissman H., Weiss G.H. and Havlin S. (1989) Transport properties of the continuous-time random walk with a long-tailed waiting-time density. J. Stat. Phys. 57, 301–317 [Google Scholar]
- 21.Saxton M.J. (2007) A biological interpretation of transient anomalous subdiffusion. I. Qualitative model. Biophys. J. 92, 1178–1191 10.1529/biophysj.106.092619 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Ben-Avraham D. and Havlin S. (2000) Diffusion and Reactions in Fractals and Disordered Systems, Cambridge University Press; ISBN 978-0-521-62278-3 [Google Scholar]
- 23.Bancaud A., Huet S., Daigle N., Mozziconacci J., Beaudouin J. and Ellenberg J. (2009) Molecular crowding affects diffusion and binding of nuclear proteins in heterochromatin and reveals the fractal organization of chromatin. EMBO J. 28, 3785–3798 10.1038/emboj.2009.340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Izeddin I., Récamier V., Bosanac L., Cissé I.I., Boudarene L., Dugast-Darzacq C. et al. (2014) Single-molecule tracking in live cells reveals distinct target-search strategies of transcription factors in the nucleus. eLife 3, e02230 10.7554/eLife.02230 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.de Gennes P.G. (1982) Kinetics of diffusion-controlled processes in dense polymer systems. I. Nonentangled regimes. J. Chem. Phys. 76, 3316–3321 10.1063/1.443328 [DOI] [Google Scholar]
- 26.Condamin S., Bénichou O., Tejedor V., Voituriez R. and Klafter J. (2007) First-passage times in complex scale-invariant media. Nature 450, 77–80 10.1038/nature06201 [DOI] [PubMed] [Google Scholar]
- 27.Bénichou O., Chevalier C., Klafter J., Meyer B. and Voituriez R. (2010) Geometry-controlled kinetics. Nat. Chem. 2, 472–477 10.1038/nchem.622 [DOI] [PubMed] [Google Scholar]
- 28.Milo R. and Phillips R. (2016) Cell Biology by the Numbers. Garland Science, Taylor & Francis Group; ISBN 978-0-8153-4537-4 [Google Scholar]
- 29.Ou Horng D., Phan S., Deerinck T.J., Thor A., Ellisman M.H. and O’Shea C.C. (2017) ChromEMT: visualizing 3d chromatin structure and compaction in interphase and mitotic cells. Science 357, eaag0025 10.1126/science.aag0025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Matsuda H., Putzel G.G., Backman V. and Szleifer I. (2014) Macromolecular crowding as a regulator of gene transcription. Biophys. J. 106, 1801–1810 10.1016/j.bpj.2014.02.019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guigas G., Kalla C. and Weiss M. (2007) The degree of macromolecular crowding in the cytoplasm and nucleoplasm of mammalian cells is conserved. FEBS Lett. 581, 5094–5098 10.1016/j.febslet.2007.09.054 [DOI] [PubMed] [Google Scholar]
- 32.Ando T. and Skolnick J. (2010) Crowding and hydrodynamic interactions likely dominate in vivo macromolecular motion. Proc. Natl Acad. Sci. U.S.A. 107, 18457–18462 10.1073/pnas.1011354107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.McGuffee S.R. and Elcock A.H. (2010) Diffusion, crowding & protein stability in a dynamic molecular model of the bacterial cytoplasm. PLoS Comput. Biol. 6, e1000694 10.1371/journal.pcbi.1000694 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pueschel R., Coraggio F. and Meister P. (2016) From single genes to entire genomes: the search for a function of nuclear organization. Development 143, 910–923 10.1242/dev.129007 [DOI] [PubMed] [Google Scholar]
- 35.Grosberg A., Rabin Y., Havlin S. and Neer A. (1993) Crumpled globule model of the three-dimensional structure of DNA. Europhys. Lett. 23, 373–378 10.1209/0295-5075/23/5/012 [DOI] [Google Scholar]
- 36.Lebedev D.V., Filatov M.V., Kuklin A.I., Islamov A.Kh., Kentzinger E., Pantina R. et al. (2005) Fractal nature of chromatin organization in interphase chicken erythrocyte nuclei: DNA structure exhibits biphasic fractal properties. FEBS Lett. 579, 1465–1468 10.1016/j.febslet.2005.01.052 [DOI] [PubMed] [Google Scholar]
- 37.Lebedev D.V., Filatov M.V., Kuklin A.I., Islamov A.Kh., Stellbrink J., Pantina R.A. et al. (2008) Structural hierarchy of chromatin in chicken erythrocyte nuclei based on small-angle neutron scattering: fractal nature of the large-scale chromatin organization. Crystallogr. Rep. 53, 110–115 10.1134/S1063774508010136 [DOI] [Google Scholar]
- 38.Lieberman-Aiden E., van Berkum N.L., Williams L., Imakaev M., Ragoczy T., Telling A. et al. (2009) Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 326, 289–293 10.1126/science.1181369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Grosberg A.Yu. (2012) How two meters of DNA fit into a cell nucleus: polymer models with topological constraints and experimental data. Poly. Sci. Ser. C 54, 1–10 10.1134/S1811238212070028 [DOI] [Google Scholar]
- 40.Récamier V., Izeddin I., Bosanac L., Dahan M., Proux F. and Darzacq X. (2014) Single cell correlation fractal dimension of chromatin. Nucleus 5, 75–84 10.4161/nucl.28227 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shinkai S., Nozaki T., Maeshima K. and Togashi Y. (2017) Bridging the dynamics and organization of chromatin domains by mathematical modeling. Nucleus 8, 353–359 10.1080/19491034.2017.1313937 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang S., Su J.-H., Beliveau B.J., Bintu B., Moffitt J.R., Wu C.-T. et al. (2016) Spatial organization of chromatin domains and compartments in single chromosomes. Science 353, 598–602 10.1126/science.aaf8084 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Goulian M. and Simon S.M. (2000) Tracking single proteins within cells. Biophys. J. 79, 2188–2198 10.1016/S0006-3495(00)76467-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hansen A.S., Woringer M., Grimm J.B., Lavis L.D., Tjian R. and Darzacq. X. (2018) Robust model-based analysis of single-particle tracking experiments with spot-on. eLife 7, e33125 10.7554/eLife.33125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Wunderlich Z. and Mirny L.A. (2008) Spatial effects on the speed and reliability of protein–DNA search. Nucleic Acids Res. 36, 3570–3578 10.1093/nar/gkn173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Coppey M., Bénichou O., Voituriez R. and Moreau M. (2004) Kinetics of target site localization of a protein on DNA: a stochastic approach. Biophys. J. 87, 1640–1649 10.1529/biophysj.104.045773 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Iwahara J., Zweckstetter M. and Clore G.M. (2006) NMR structural and kinetic characterization of a homeodomain diffusing and hopping on nonspecific DNA. Proc. Natl Acad. Sci. U.S.A. 103, 15062–15067 10.1073/pnas.0605868103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Doucleff M. and Clore G.M. (2008) Global jumping and domain-specific intersegment transfer between DNA cognate sites of the multidomain transcription factor Oct-1. Proc. Natl Acad. Sci. U.S.A. 105, 13871–13876 10.1073/pnas.0805050105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kopelman R. (1986) Rate processes on fractals: theory, simulations, and experiments. J. Stat. Phys. 42, 185–200 10.1007/BF01010846 [DOI] [Google Scholar]
- 50.Berry H. (2002) Monte carlo simulations of enzyme reactions in two dimensions: fractal kinetics and spatial segregation. Biophys. J. 83, 1891–1901 10.1016/S0006-3495(02)73953-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dino O. and Yizhak R. (2016) Effect of non-specific interactions on formation and stability of specific complexes. J. Chem. Phys. 144, 205104 10.1063/1.4952981 [DOI] [PubMed] [Google Scholar]
- 52.Shagolsem L.S. and Rabin Y. (2016) Particle dynamics in fluids with random interactions. J. Chem. Phys. 144, 194504 10.1063/1.4949546 [DOI] [PubMed] [Google Scholar]
- 53.Bergeron-Sandoval L.-P., Safaee N. and Michnick S.W. (2018) Mechanisms and consequences of macromolecular phase separation. Cell 165, 1067–1079 10.1016/j.cell.2016.05.026 [DOI] [PubMed] [Google Scholar]
- 54.Tsai A., Muthusamy A.K., Alves M.R.P., Lavis L.D., Singer R.H., Stern D.L. et al. (2017) Nuclear microenvironments modulate transcription from low-affinity enhancers. eLife 6, e28975 10.7554/eLife.28975 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kwon I., Kato M., Xiang S., Wu L., Theodoropoulos P., Mirzaei H. et al. (2013) Phosphorylation-regulated binding of RNA polymerase II to fibrous polymers of low-complexity domains. Cell 155, 1049–1060 10.1016/j.cell.2013.10.033 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sherry K.P., Das R.K., Pappu R.V. and Barrick D. (2017) Control of transcriptional activity by design of charge patterning in the intrinsically disordered RAM region of the notch receptor. Proc. Natl Acad. Sci. U.S.A. 114, E9243–E9252 10.1073/pnas.1706083114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Chong S., Dugast-Darzacq C., Liu Z., Dong P., Dailey G., Banala S. et al. (2017) Imaging dynamic and selective low-complexity domain interactions that control gene transcription. Science, eaar2555 10.1126/science.aar2555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Shammas S.L. (2017) Mechanistic roles of protein disorder within transcription. Curr. Opin. Struct. Biol. 42, 155–161 10.1016/j.sbi.2017.02.003 [DOI] [PubMed] [Google Scholar]
- 59.Kato M., Han T.W., Xie S., Shi K., Du X., Wu L.C. et al. (2012) Cell-free formation of RNA granules: Low complexity sequence domains form dynamic fibers within hydrogels. Cell 149, 753–767 10.1016/j.cell.2012.04.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Liu J., Perumal N.B., Oldfield C.J., Su E.W., Uversky V.N. and Dunker A.K. (2006) Intrinsic disorder in transcription factors. Biochemistry 45, 6873–6888 10.1021/bi0602718 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Nott T.J., Petsalaki E., Farber P., Jervis D., Fussner E., Plochowietz A. et al. (2015) Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles. Mol. Cell 57, 936–947 10.1016/j.molcel.2015.01.013 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Jain A. and Vale R.D. (2017) RNA phase transitions in repeat expansion disorders. Nature 546, 243–247 10.1038/nature22386 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Li P., Banjade S., Cheng H.-C., Kim S., Chen B., Guo L. et al. (2012) Phase transitions in the assembly of multivalent signalling proteins. Nature 483, 336–340 10.1038/nature10879 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Molliex A., Temirov J., Lee J., Coughlin M., Kanagaraj A.P., Kim H.J. et al. (2015) Phase separation by low complexity domains promotes stress granule assembly and drives pathological fibrillization. Cell 163, 123–133 10.1016/j.cell.2015.09.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Patel A., Malinovska L., Saha S., Wang J., Alberti S., Krishnan Y. et al. (2017) ATP as a biological hydrotrope. Science 356, 753–756 10.1126/science.aaf6846 [DOI] [PubMed] [Google Scholar]
- 66.Patel A., Lee H.O., Jawerth L., Maharana S., Jahnel M., Hein M.Y. et al. (2015) A liquid-to-solid phase transition of the ALS protein FUS accelerated by disease mutation. Cell 162, 1066–1077 10.1016/j.cell.2015.07.047 [DOI] [PubMed] [Google Scholar]
- 67.Pak C.W., Kosno M., Holehouse A.S., Padrick S.B., Mittal A., Ali R. et al. (2016) Sequence determinants of intracellular phase separation by complex coacervation of a disordered protein. Mol. Cell 63, 72–85 10.1016/j.molcel.2016.05.042 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Gimona M. (2006) Protein linguistics — a grammar for modular protein assembly? Nat. Rev. Mol. Cell Biol. 7, 68–73 10.1038/nrm1785 [DOI] [PubMed] [Google Scholar]
- 69.Brady J.P., Farber P.J., Sekhar A., Lin Y.-H., Huang R., Bah A. et al. (2017) Structural and hydrodynamic properties of an intrinsically disordered region of a germ cell-specific protein on phase separation. Proc. Natl Acad. Sci. U.S.A. 114, E8194–E8203 10.1073/pnas.1706197114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Reichheld S.E., Muiznieks L.D., Keeley F.W. and Sharpe S. (2017) Direct observation of structure and dynamics during phase separation of an elastomeric protein. Proc. Natl Acad. Sci. U.S.A. 114, E4408–E4415 10.1073/pnas.1701877114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Das R.K., Ruff K.M. and Pappu R.V. (2015) Relating sequence encoded information to form and function of intrinsically disordered proteins. Curr. Opin. Struct. Biol. 32, 102–112 10.1016/j.sbi.2015.03.008 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Quiroz F.G. and Chilkoti A. (2015) Sequence heuristics to encode phase behaviour in intrinsically disordered protein polymers. Nat. Mater. 14, 1164–1171 10.1038/nmat4418 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wei M.-T., Elbaum-Garfinkle S., Holehouse A.S., Chen C.C.-H., Feric M., Arnold C.B. et al. (2017) Phase behaviour of disordered proteins underlying low density and high permeability of liquid organelles. Nat. Chem. 9, 1118–1125 10.1038/nchem.2803 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Shav-Tal Y., Blechman J., Darzacq X., Montagna C., Dye B.T., Patton J.G. et al. (2005) Dynamic sorting of nuclear components into distinct nucleolar caps during transcriptional inhibition. Mol. Biol. Cell. 16, 2395–2413 10.1091/mbc.E04-11-0992 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Cisse I.I., Izeddin I., Causse S.Z., Boudarene L., Senecal A., Muresan L. et al. (2013) Real-time dynamics of RNA polymerase II clustering in live human cells. Science 341, 664–667 10.1126/science.1239053 [DOI] [PubMed] [Google Scholar]
- 76.Wang J.T., Smith J., Chen B.-C., Schmidt H., Rasoloson D., Paix A. et al. (2014) Regulation of RNA granule dynamics by phosphorylation of serine-rich, intrinsically disordered proteins in c. elegans. eLife 3, e04591 10.7554/eLife.04591 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cho W.-K., Jayanth N., Mullen S., Tan T.H., Jung Y.J. and Cissé I.I. (2016) Super-resolution imaging of fluorescently labeled, endogenous RNA polymerase II in living cells with CRISPR/Cas9-mediated gene editing. Sci. Rep. 6, 35949 10.1038/srep35949 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Ghosh S.K., Cherstvy A.G. and Metzler. R. (2015) Non-universal tracer diffusion in crowded media of non-inert obstacles. Phys. Chem. Chem. Phys. 17, 1847–1858, 10.1039/C4CP03599B [DOI] [PubMed] [Google Scholar]
- 79.Loffreda A., Jacchetti E., Antunes S., Rainone P., Daniele T., Morisaki T. et al. (2017) Live-cell p53 single-molecule binding is modulated by c-terminal acetylation and correlates with transcriptional activity. Nat. Commun. 8, 313 10.1038/s41467-017-00398-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Speil J., Baumgart E., Siebrasse J.-P., Veith R., Vinkemeier U. and Kubitscheck U. (2011) Activated STAT1 transcription factors conduct distinct saltatory movements in the cell nucleus. Biophys. J. 101, 2592–2600 10.1016/j.bpj.2011.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Elf J., Li G.-W. and Xie X.S. (2007) Probing transcription factor dynamics at the single-molecule level in a living cell. Science 316, 1191–1194 10.1126/science.1141967 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Mazza D., Abernathy A., Golob N., Morisaki T. and McNally J.G. (2012) A benchmark for chromatin binding measurements in live cells. Nucleic Acids Res. 40, e119–e119 10.1093/nar/gks701 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Sekiya T., Muthurajan U.M., Luger K., Tulin A.V. and Zaret K.S. (2009) Nucleosome-binding affinity as a primary determinant of the nuclear mobility of the pioneer transcription factor FoxA. Genes Dev. 23, 804–809 10.1101/gad.1775509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Clauß K., Popp A.P., Schulze L., Hettich J., Reisser M., Escoter T.L. et al. (2017) DNA residence time is a regulatory factor of transcription repression. Nucleic Acids Res. 45, 11121–11130 10.1093/nar/gkx728 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Zhen C.Y., Tatavosian R., Huynh T.N., Duc H.N., Das R., Kokotovic M. et al. (2016) Live-cell single-molecule tracking reveals co-recognition of H3K27me3 and DNA targets polycomb Cbx7-PRC1 to chromatin. eLife 5, e17667 10.7554/eLife.17667 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Isaacson S.A., McQueen D.M. and Peskin C.S. (2011) The influence of volume exclusion by chromatin on the time required to find specific DNA binding sites by diffusion. Proc. Natl Acad. Sci. U.S.A. 108, 3815–3820 10.1073/pnas.1018821108 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Chronis C., Fiziev P., Papp B., Butz S., Bonora G., Sabri S. et al. (2017) Cooperative binding of transcription factors orchestrates reprogramming. Cell 168, 442–459.e20 10.1016/j.cell.2016.12.016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Sukenik S., Ren P. and Gruebele M. (2017) Weak protein-protein interactions in live cells are quantified by cell-volume modulation. Proc. Natl Acad. Sci. U.S.A. 114, 6776–6781 10.1073/pnas.1700818114 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Maldonado A.Y., Burz D.S. and Shekhtman A. (2011) In-cell NMR spectroscopy. Prog. Nucl. Magn. Reson. Spectrosc. 59, 197–212 10.1016/j.pnmrs.2010.11.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Freedberg D.I. and Selenko P. (2014) Live cell NMR. Annu. Rev. Biophys. 43, 171–192 10.1146/annurev-biophys-051013-023136 [DOI] [PubMed] [Google Scholar]
- 91.Theillet F.-X., Binolfi A., Bekei B., Martorana A., Rose H.M., Stuiver M. et al. (2016) Structural disorder of monomeric α-synuclein persists in mammalian cells. Nature 530, 45–50 10.1038/nature16531 [DOI] [PubMed] [Google Scholar]
- 92.Balzarotti F., Eilers Y., Gwosch K.C., Gynnå A.H., Westphal V., Stefani F.D. et al. (2017) Nanometer resolution imaging and tracking of fluorescent molecules with minimal photon fluxes. Science 355, 606–612 10.1126/science.aak9913 [DOI] [PubMed] [Google Scholar]
- 93.Limouse C., Bell J.C., Fuller C.J., Straigh A.F. and Mabuchi H. (2017) Intramolecular dynamics of single molecules in free diffusion. bioRxiv, 120311 10.1101/120311 [DOI] [Google Scholar]
- 94.Singh A.P., Galland R., Finch-Edmondson M.L., Grenci G., Sibarita J.-B., Studer V. et al. (2017) 3d protein dynamics in the cell nucleus. Biophys. J. 112, 133–142 10.1016/j.bpj.2016.11.3196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Harmon T.S., Crabtree M.D., Shammas S.L., Posey A.E., Clarke J. and Pappu R.V. (2016) GADIS: algorithm for designing sequences to achieve target secondary structure profiles of intrinsically disordered proteins. Protein Eng. Des. Sel. 29, 339–346 10.1093/protein/gzw034 [DOI] [PMC free article] [PubMed] [Google Scholar]