

Abstract
To clarify the functions and mechanism of stimulated by retinoic acid gene 8 (Stra8) in spermatogenesis, we analyzed the testes from Stra8 knockout and wild-type mice during the first wave of spermatogenesis. Comparisons showed no significant differences in morphology and number of germ cells at 11 days postpartum, while 21 differentially expressed genes (DEGs) associated with spermatogenesis were identified. We speculate that Stra8 performs many functions in different phases of spermatogenesis, such as establishment of spermatogonial stem cells, spermatogonial proliferation and self-renewal, spermatogonial differentiation and meiosis, through direct or indirect regulation of these DEGs. We therefore established a preliminary regulatory network of Stra8 during spermatogenesis. These results will provide a theoretical basis for further research on the mechanism underlying the role of Stra8 in spermatogenesis.
Keywords: differentiation, meiosis, proliferation, spermatogenesis, spermatogonia, Stra8
INTRODUCTION
In mammals, spermatogenesis is a highly ordered and precisely regulated process, ensuring that millions of spermatozoa are produced at a constant rate. Mature sperm production depends on the stable self-renewal, proliferation, and differentiation of spermatogonial stem cells (SSCs). The A1 spermatogonia divide successively into A2, A3, A4, intermediate (In), and B spermatogonia. After a round of DNA replication, B spermatogonia undergo two rounds of meiosis to form spermatocytes, which then undergo morphological changes to become haploid sperm cells. Successful spermatogenesis occurs through precise regulation of gene expression. Stimulated by retinoic acid gene 8 (Stra8) is a key molecule involved in meiosis initiation and plays an important role in spermatogenesis.1
Retinoic acid (RA), which is an active metabolite of Vitamin A, is a vital signaling molecule involved in germ cell development and correct entry into meiosis. Stra8 was identified as RA-responsive gene by Oulad-Abdelghani in 1995.2,3 RA directly regulates the expression of Stra8 via its specific nuclear hormone receptors (retinoic acid receptors [RARs] and retinoid X receptors [RXRs]). RARs form heterodimers with RXRs, which bind to the retinoic acid responsive elements (RAREs) located in the Stra8 promoter. RA-bound RAR-RXR heterodimers activate or repress RA-responsive gene expression by recruiting histone acetyltransferase or deacetylase.4,5 The lack of Stra8 expression in the embryonic testis is mainly due to the degradation of RA by cytochrome P450 26B1 (CYP26B1). However, after embryonic day 13.5 (E13.5), the expression levels of CYP26B1 gradually decrease.6 At this point, Nanos2 production is initiated, which mediates direct downregulation of Stra8 expression, preventing the induction of meiosis in the germ cells in embryonic testes and inhibiting apoptosis.4 Moreover, Creb-binding protein (CBP), a coactivator of histone acetyltransferase, mediates Stra8 transcription, while p300 represses Stra8 transcription by SUMO-1 modification.7 In addition, Sohlh1 and Sohlh2 repress Stra8 expression by binding to the E-Boxes located in the promoter.8 DMRT1 regulates meiosis by enhancing Sohlh1 expression and repressing Stra8 expression.9 To date, most studies have focused on the proteins that directly or indirectly regulate Stra8; however, the identities of proteins regulated by Stra8 as well as regulatory proteins downstream of Stra8 remain to be established. Furthermore, the functional mechanism and regulatory network of Stra8 during spermatogenesis remain to be clarified.
In this study, we performed a histological analysis of Stra8 knockout (Stra8−/− KO) testes and wild-type (WT) testes during the first wave of spermatogenesis. Although there were no obvious differences in morphology or the number of germ cells at 11 days postpartum (dpp), significant differences were detected at 12 dpp. Compared with WT testes, RNA sequencing (RNA-Seq) analysis revealed 96 differentially expressed genes (DEGs) in Stra8−/− testes, 21 of which are reported to be directly or indirectly involved in spermatogenesis. We propose a regulatory network for Stra8 as a multifunctional protein involved in the establishment and maintenance of SSCs, cell proliferation, self-renewal, differentiation, and maintenance of the undifferentiated state during spermatogenesis.
MATERIALS AND METHODS
Mice
Heterozygous B6. Cg-Stra8tm1Dcp/J mice (Stra8+/− mice) were purchased from Jackson Laboratory (Bar Harbor, ME, USA) on a C57BL/6 genetic background. All mice were maintained at the Laboratory Animal Center of Yangzhou University (Yangzhou, China) under conditions of controlled temperature (21°C–25°C), a 12 h/12 h light/dark cycle, and a normal diet. The day of birth was defined as 0 dpp. All animal experiments conducted as part of this study were approved by the Animal Ethics Committee of Yangzhou University.
DNA extraction and genotyping
Female and male heterozygous mice were housed together, and the genotypes of the offspring were identified following protocols provided by the Jackson Laboratory. All mice genotypes were identified by polymerase chain reaction (PCR) amplification of the DNA extracted from tail tissue. The tissue was incubated with 700 μl Lysis buffer I and 30 μl proteinase K (Sigma Chemical Co., St. Louis, MO, USA) for 2 h at 55°C before the DNA was obtained by phenol/chloroform extraction and ethanol precipitation.
Tail DNA was genotyped by PCR amplification using the following primers: Stra8-P1 (5’-CAACCAACCCAGTGATGATG-3’), Stra8-P2 (5’-TCAGGTCAGGCTGCTAGGAT-3’), and Stra8-P3 (5’-GATAGCTTGGCTGCAGGTC-3’). PCR conditions were as follows: 10 cycles of 94°C for 20 s, 65°C for 15 s, and 68°C for 10 s and 28 cycles of 94°C for 15 s, 60°C for 15 s, and 72°C for 10 s, respectively. Genotypes were identified from the PCR products separated by 1.2% (w/v) agarose gel electrophoresis (WT: 316 bp; mutant type [MT]: 200 bp).
Histological sample preparation and staining
Mouse testes were fixed with Bouin's solution for 4 h or 4% (w/v) paraformaldehyde for 24 h, then in chloroform overnight followed by dehydration in a gradient alcohol series. After transfer into xylene, testicular tissue was soaked in liquid paraffin for 2 h and then embedded in paraffin for further analysis.
Testes from 5–12 dpp and adult mice were collected, and sections were prepared (thickness, 4 μm) for HE staining. Types of germ cells were identified by their location, nuclear size, and chromatin characteristics.
Immunofluorescence, immunohistochemistry analysis, and cell counting
Mouse testes (5–12 dpp) were fixed in 4% (w/v) paraformaldehyde overnight for immunofluorescence analysis. Testes from Stra8−/− and WT mice (11 dpp) were used for immunohistochemistry. Tissues were dewaxed, treated by methanol and hydrogen peroxide, and heated in antigen repair solution (citric acid and trisodium citrate acid). After cooling, sections were blocked by incubation in donkey serum (Solarbio® Life Sciences, Beijing, China) at room temperature before incubation with primary antibodies (Ddx4, Nanos3, Egr4, Abcam, Cambridge, England; Asb9, Santa Cruz Biotechnology, Santa Cruz, CA, USA; Msh5, Biorbyt, Cambridge, England; Utf1, HuaAn Biotechnology Co., Ltd., Hangzhou, China) overnight at 4°C. On the 2nd day, sections were stained with fluorescein isothiocyanate (FITC)-labeled anti-rabbit/mouse IgG (Sigma Chemical Co.) at 37°C for 1 h and then restained with diamidino-phenyl-indole (DAPI, Solarbio® Life Sciences) for immunofluorescence analysis. Immunohistochemistry was then performed followed the protocol provided for the GT Vision™ III Detection System (Gene Tech, Shanghai, China). Images were obtained from a fluorescence microscope (Eclipse 70i, Nikon, Tokyo, Japan) and the number of Ddx4-positive cells was counted. Statistical difference between two groups was assessed by t-test (n ≥ 6).
RNA sequencing
WT and homozygous testes from 11 dpp mice were collected and stored in liquid nitrogen after addition of Trizol (Life Technologies, Frederick, MD, USA). Three groups of four testes from two mice were used for RNA sequencing. Total RNA was prepared according to normal protocols. The eukaryotic mRNA was purified from total RNA using Oligo(dT) magnetic beads. After purification and terminal repair, the mRNAs were then fragmented, and double-stranded cDNA was generated for use as a template in PCR amplification to generate the complete library. The library was then analyzed in an Agilent 2100 Bioanalyzer (Agilent Technologies Co., Ltd., Palo Alto, CA, USA) and ABI StepOnePlus™ Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) for quality and yield.
RNA-Seq analysis was performed by BGI Genomics Co., Ltd., Shenzhen, China. Gene expression levels were quantified using the RSEM software package, University of Tokyo, Tokyo, Japan. FPKM represents a unit of gene expression that can be used for direct comparison of the different gene expression levels of different samples. The NOISeq method was used to screen for DEGs in WT and Stra8−/− testes. Gene ontology (GO) annotation analysis of the distribution of gene functions was performed using WEGO software (Web Gene Ontology Annotation Plot, http://wego.genomics.org.cn/). Pathway enrichment analysis of the interaction between genes was performed by searching the Kyoto Encyclopedia of Genes and Genomes (KEGG) database.
RNA extraction and qRT-PCR analysis
Total RNA was extracted from WT and homozygous mice testes (11 dpp) using the one-step guanidine isothiocyanate-phenol-chloroform method. After removal of genomic DNA, reverse transcription was performed according to the manufacturer's instructions. Quantitative reverse transcription-PCR (qRT-PCR) analysis was then performed using GoTaq® qPCR Master Mix (TaKaRa, Tokyo, Japan) with the primers listed in Table 1. The thermal cycling conditions were as follows: 40 cycles of denaturation at 95°C for 15 s, annealing at 60°C for 30 s, and elongation at 72°C for 30 s. qRT-PCR results were normalized against Gapdh as a reference to determine the relative expression levels of the genes.
Table 1.
Primer sequences for reverse transcription polymerase chain reaction
Western blot analysis
Testis proteins from 11 dpp mice were extracted using RIPA buffer (Solarbio® Life Sciences). After centrifugation at 10 000 g for 15 min at 4°C, the supernatant was collected. Total proteins (50 μg) were separated by sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride membrane. After blocking in Tris-buffered saline (TBS) containing 5% (w/v) nonfat milk, the membrane was incubated with the primary detection antibodies (Stra8, Nanos3, Egr4, Pou5f1, Gapdh, Abcam; Rhox10, donated by Miles F Wilkinson Lab, University of California, Berkeley, CA, USA; Asb9, Dmc1, Santa Cruz; Msh5, Biorbyt) overnight at 4°C. The membranes were then incubated with secondary antibodies (ZSbio, Beijing, China) at room temperature for 1.5 h. Proteins were detected by electrogenerated chemiluminescence (ECL).
Statistical analysis
Statistical analysis was performed using SPSS18.0 (SPSS Inc., Chicago, IL, USA). Data were presented as mean ± standard deviation (s.d.) and analyzed by Student's t- test. All experiments were repeated at least three times. P ≤ 0.05 was considered to indicate statistical significance.
RESULTS
Morphological analysis of Stra8−/− mouse testes during the first wave of spermatogenesis
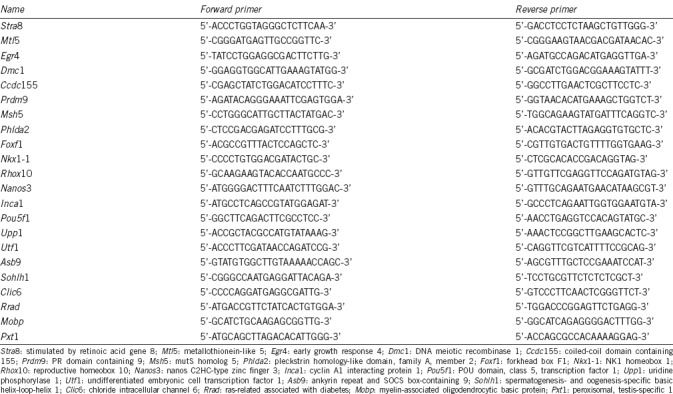
The offspring of heterozygous mice were genotyped by PCR amplification of testicular DNA fragments. Fragments of 316 bp and 200 bp were amplified from DNA extracts from heterozygous testes, while fragments of 200 bp and 316 bp were detected from Stra8−/− and WT testes, respectively (Figure 1a).
Figure 1.
Histological analysis of testis from pubescent Stra8−/− mice. (a) Genotyping of Stra8−/− mice. Marker: DM2000 Plus II DNA Marker. (b–j) Hematoxylin and eosin staining of testes sections from WT and Stra8−/− mice. (b–i) Left, sections from WT testes at 5–12 dpp; right, sections from homozygotic testes at 5–12 dpp. (j) Left, testis sections from WT adult mice; right, testis sections from homozygotic adult mice (n ≥ 6; scale bars = 50 μm). WT: wild-type mouse; Heterozygotic type: Stra8+/− mouse; homozygotic type: Stra8−/− mouse; Control: water; 1–3: genotyping of three Stra8 knockout mice. Stra8: stimulated by retinoic acid gene 8.
Spermatogenesis in Stra8−/− mice is arrested in meiotic prophase I and the testes undergo morphological changes during the first wave of spermatogenesis. Therefore, testes of Stra8+/+ and Stra8−/− mice were collected at 5–12 dpp and tissue sections were stained with HE and evaluated by optical microscopy to identify the earliest point at which morphological differences in germ cells could be detected. This information was also important for selecting RNA-Seq samples. At 5, 6, and 7 dpp, only spermatogonia and Sertoli cells were observed in WT testis (Figure 1b–1d, left). Some spermatogonia showed more intense staining of the cells that were located in the adluminal compartment of seminiferous tubules, which then migrated to the basal lamina of the tubules at 8 dpp and 9 dpp (Figure 1e and 1f, left). In addition, the prospermatogonia transitioned to type A and B spermatogonia. Subsequently, at 10 dpp and 11 dpp, the diameter of the seminiferous tubules increased and the number of type B spermatogonia increased (Figure 1g and 1h, left). Primary spermatocytes first appeared at 12 dpp (Figure 1i, left). All types of germ cells were contained in the adult mouse testis (Figure 1j, left). There were no obvious differences in testicular histomorphology between Stra8−/− and WT mice from 5 to 11 dpp (Figure 1b–1i). However, at 12 dpp, there were significantly fewer primary spermatocytes in Stra8−/− mouse testes than in WT testes, and vacuoles appeared (P < 0.05; Figure 1i). In accordance with these observations, adult Stra8−/− mice are infertile and lack meiotic and postmeiotic cells (Figure 1j).
Germ cell counts in developing testes
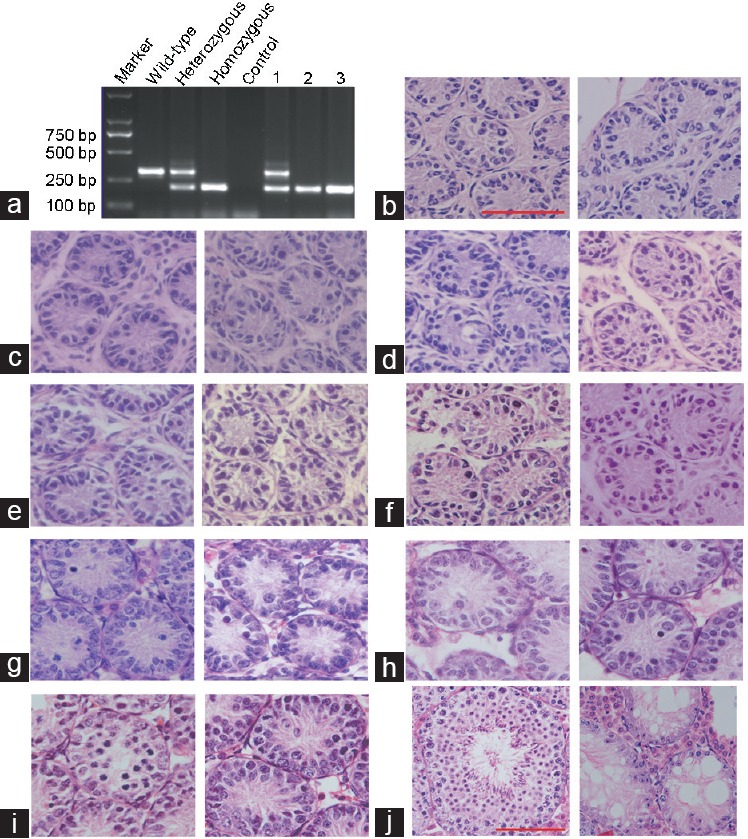
Germ cells were counted by immunofluorescence staining of the DEAD (Asp-Glu-Ala-Asp) box polypeptide 4 (Ddx4), which is a specific marker expressed in the cytoplasm of germ cells. The number and signal intensity of Ddx4-positive cells increased gradually in the developing testis (Figure 2). In accordance with the HE staining at 5–11 dpp, there was no significant difference in the number and signal intensity of Ddx4-positive cells between Stra8−/− and Stra8+/+ testes (Figure 2a–2g and 2i). However, at 12 dpp, the number of Ddx4-positive cells decreased significantly in Stra8−/− mice compared with that in the WT mice (P < 0.05, n ≥ 6; (Figure 2h and 2i). Therefore, testes were obtained from Stra8−/− and Stra8+/+ mice at 11 dpp for further RNA-Seq analysis.
Figure 2.
Germ cell counts in developing testes. (a–h) Immunofluorescence staining for Ddx4 (a specific germ cell marker) at 5–12 dpp. Left, sections from wild-type testes at 5–12 dpp; right, sections from Stra8 homozygote testes at 5–12 dpp. Immunolabelling of Ddx4 (red), and DAPI (blue). n ≥ 6; scale bars = 50 μm. (i) Average number of Ddx4-positive spermatogenic cells per tubule in cross-section (n ≥ 6, mean ± standard deviation, *P < 0.05). Stra8: stimulated by retinoic acid gene 8; DAPI: diamidino-phenyl-indole.
Identification of DEGs in Stra8−/− testes
To investigate the DEGs associated with Stra8 deficiency, we generated a RNA-Seq profile of the transcriptome of Stra8−/− testes and identified 40 downregulated transcripts and 56 upregulated transcripts in Stra8−/− testes compared with the WT (P < 0.05).
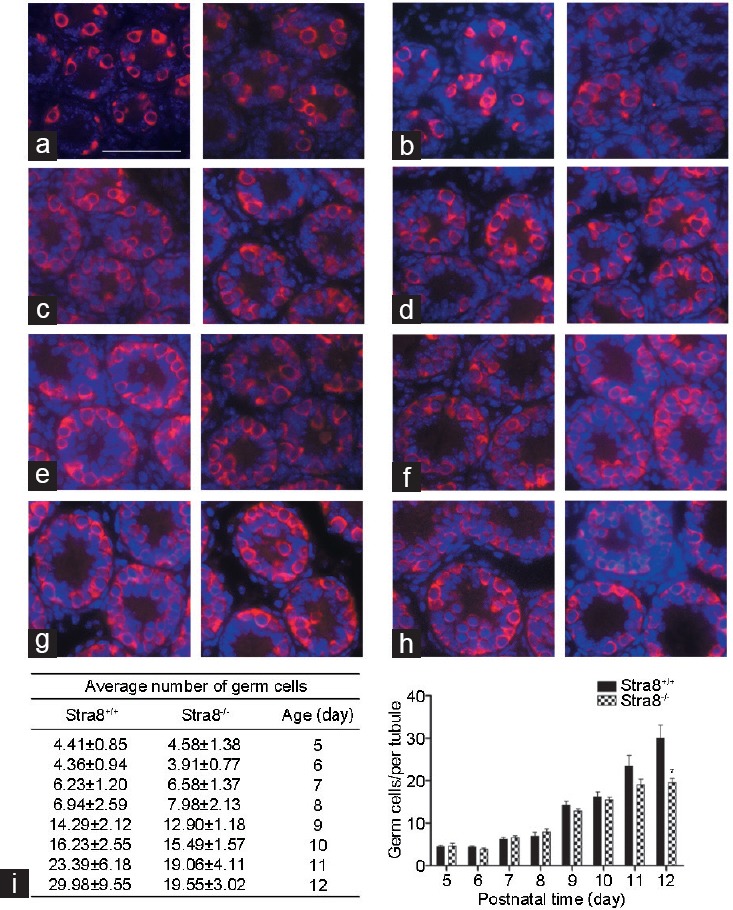
GO analysis of the distribution of gene functions revealed that the DEGs were predominantly involved in biological processes, cellular components, and molecular functions including reproduction, reproductive process, and transcription (Figure 3). Genes often interact with each other in biological functions; therefore, we performed pathway enrichment analysis of 58 RNAs encoded by the DEGs. The top 20 KEGG enrichment and term types are shown in Figure 4. These DEGs were enriched in the pathways of spliceosome, RNA transport and Toll-like receptor signaling. They also involved in the KEGG pathway terms cellular processes, environmental information processing, genetic information processing, human disease and organismal systems.
Figure 3.
Gene ontology functional classification of differentially expressed genes. All gene ontology terms are grouped into three ontologies: biological process, cellular component, and molecular function.
Figure 4.
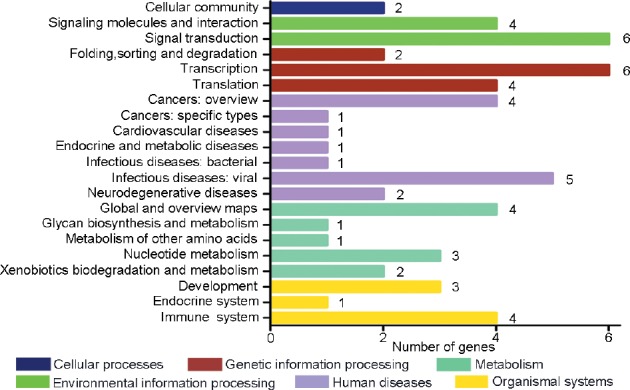
Pathway enrichment analysis of differentially expressed genes. Kyoto encyclopedia of genes and genomes classification of differentially expressed genes.
Role of DEGs in Stra8-mediated regulation of spermatogenesis
From the FPKM value and previous reports, we identified 21 DEGs directly or indirectly related to spermatogenesis. In addition to Stra8, the results of qRT-PCR analysis of the expression levels of the 21 DEGs were consistent with the results of RNA-Seq analysis. Testis expressions of metallothionein-like 5 (Mtl5), DNA meiotic recombinase 1 (Dmc1), coiled-coil domain containing 155 (Ccdc155), inhibitor of CDK, cyclin A1 interacting protein 1 (Inca1), mutS homolog 5 (Msh5), and PR domain containing 9 (Prdm9) were downregulated, while reproductive homeobox10 (Rhox10), early growth response 4 (Egr4), uridine phosphorylase 1 (Upp1), undifferentiated embryonic cell transcription factor 1 (Utf1), POU domain, class 5, transcription factor 1 (Pou5f1), nanos C2HC-type zinc finger 3 (Nanos3), ankyrin repeat and SOCS box-containing 9 (Asb9), spermatogenesis- and oogenesis-specific basic helix-loop-helix1 (Sohlh1), peroxisomal, testis-specific 1 (Pxt1), pleckstrin homology-like domain, family A, member 2 (Phlda2), forkhead box F1 (Foxf1), NK1 homeobox1 (Nkx1-1), ras-related associated with diabetes (Rrad), chloride intracellular channel 6 (Clic6), and myelin-associated oligodendrocytic basic protein (Mobp) were upregulated (Figure 5a).
Figure 5.
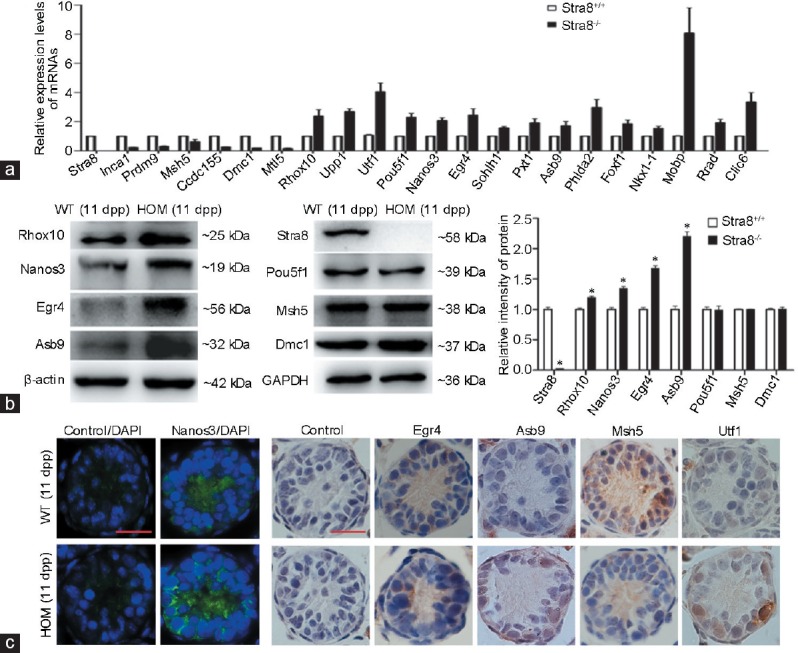
(a) Quantitative RT-PCR analysis of the DEGs involved in spermatogenesis. Data represent mean ± s.d., and values were normalized to WT mRNA levels. All 21 DEGs were statistically significant (P < 0.05). (b) Western blot analysis of proteins extracted from Stra8−/− testes at 11 dpp. Blots were probed with primary antibodies for the detection of Stra8, Rhox10, Nanos3, Egr4, Asb9, Pou5f1, Msh5 and Dmc1. The protein levels of Rhox10, Nanos3, Egr4, and Asb9 were significantly increased in HOM testes compared with WT testes (*P < 0.05). Pou5f1, Msh5, and Dmc1 protein levels were not significantly different between WT and Stra8 knockout testes. (c) Immunohistochemistry/immunofluorescence analysis of Nanos3, Egr4, Asb9, Msh5, and Utf1 expression in WT and Stra8 knockout testes; Nanos3 (green), DAPI (blue). Scale bars = 10 μm. RT-PCR: reverse transcription polymerase chain reaction; DAPI: diamidino-phenyl-indole; s.d.: standard deviation; WT: wild-type; HOM: homozygous; dpp: days postpartum; DEGs: differentially expressed genes. Stra8: stimulated by retinoic acid gene 8; Mtl5: metallothionein-like 5; Egr4: early growth response 4; Dmc1: DNA meiotic recombinase 1; Ccdc155: coiled-coil domain containing 155; Prdm9: PR domain containing 9; Msh5: mutS homolog 5; Phlda2: pleckstrin homology-like domain, family A, member 2; Foxf1: forkhead box F1; Nkx1-1: NK1 homeobox 1; Rhox10: reproductive homeobox 10; Nanos3: nanos C2HC-type zinc finger 3; Inca1: cyclin A1 interacting protein 1; Pou5f1: POU domain, class 5, transcription factor 1; Upp1: uridine phosphorylase 1; Utf1: undifferentiated embryonic cell transcription factor 1; Asb9: ankyrin repeat and SOCS box-containing 9; Sohlh1: spermatogenesis- and oogenesis-specific basic helix-loop-helix 1; Clic6: chloride intracellular channel 6; Rrad: ras-related associated with diabetes; Mobp: myelin-associated oligodendrocytic basic protein; Pxt1: peroxisomal, testis-specific 1.
Of the 21 DEGs, we analyzed the expression levels of seven corresponding proteins by western blot (Figure 5b). In accordance with the levels of DEG expression determined by RNA-Seq and qRT-PCR analyses, the protein levels of Rhox10, Nanos3, Egr4, and Asb9 were significantly increased in Stra8−/− testes compared with those in WT testes (P < 0.05). By contrast, there were no significant differences in the protein levels of Pou5f1, Msh5, and Dmc1 between WT and KO testes. We further detected the cellular distribution of Nanos3, Egr4, Asb9, and Msh5 in WT and KO testes by immunohistochemistry or immunofluorescence analysis (Figure 5c). Nanos3 protein was located in the cytoplasm of spermatogonia, which showed an obvious increase in KO testes compared with WT testes. Egr4 was weakly expressed in spermatogonia and early spermatocytes of wild-type mouse testes, but showed higher intensity in homozygous spermatocytes. Asb9 had a similar pattern and tend of expression to Egr4, but the expression of Msh5 was obviously reduced in homozygotic testes, and it mainly appeared in the nucleus of spermatocytes. Utf1, as an undifferentiated marker, was expressed in the basal spermatogonia and the expression intensity was enhanced in KO testes.
Regulatory network involving Stra8 in spermatogenesis
Different proteins often form complexes through complicated interactions to perform their biological functions. A literature review and protein–protein interaction network analysis of the 21 DEGs indicated thatStra8 plays a variety of roles in multiple stages of spermatogenesis through cooperative interactions with the genes listed. We established a preliminary regulatory network of Stra8 during spermatogenesis (Figure 6). In addition to the roles in spermatogonial differentiation and meiotic prophase I, Stra8 is implicated in the establishment of SSCs, spermatogonial proliferation, self-renewal, and physiological processes.
Figure 6.
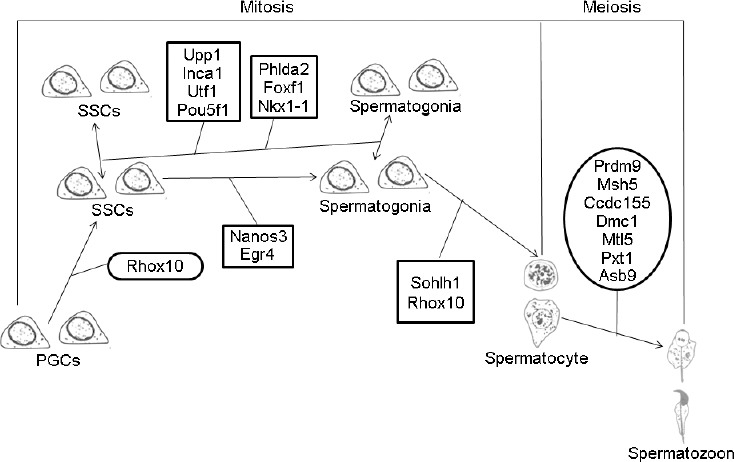
Regulatory network involving Stra8 in spermatogenesis. SSC: spermatogonial stem cell; PGC: primordial germ cell. Stra8: stimulated by retinoic acid gene 8; Mtl5: metallothionein-like 5; Egr4: early growth response 4; Dmc1: DNA meiotic recombinase 1; Ccdc155: coiled-coil domain containing 155; Prdm9: PR domain containing 9; Msh5: mutS homolog 5; Phlda2: pleckstrin homology-like domain, family A, member 2; Foxf1: forkhead box F1; Nkx1-1: NK1 homeobox 1; Rhox10: reproductive homeobox 10; Nanos3: nanos C2HC-type zinc finger 3; Inca1: cyclin A1 interacting protein 1; Pou5f1: POU domain, class 5, transcription factor 1; Upp1: uridine phosphorylase 1; Utf1: undifferentiated embryonic cell transcription factor 1; Asb9: ankyrin repeat and SOCS box-containing 9; Sohlh1: spermatogenesis- and oogenesis-specific basic helix-loop-helix 1; Pxt1: peroxisomal, testis-specific 1.
DISCUSSION
Spermatogenesis is a multifaceted process by which SSCs undergo mitotic and meiotic divisions, eventually giving rise to haploid spermatozoa. Stra8 plays a critical role in mammalian spermatogenesis, and mutations are associated with increased risk of azoospermia and oligozoospermia in the Han Chinese population.1 This phenomenon has been demonstrated in previous studies; however, the mechanism underlying the function of Stra8 in spermatogenesis is still not fully understood.
In this study, we investigated the function of Stra8 and its potential regulatory networks during spermatogenesis using male Stra8 KO mice. The choice of 5–12 dpp testes was dictated by the expression of Stra8, which started at 5 dpp, reached a peak at 10 dpp, and then began to decline4 and was, therefore, suitable for investigating the mechanism underlying the functions of Stra8. There were no apparent differences in the morphology and number of germ cells in WT and Stra8−/− testes at 11 dpp. The DEGs identified at 11 dpp were most likely due to Stra8 deficiency. According to the FPKM value and previous reports, we demonstrated that 21 of these DEGs were directly or indirectly associated with spermatogenesis. Thus, it can be speculated that Stra8 regulates spermatogenesis by coordinating with these genes.
The role of Stra8 in establishment of SSCs, cell proliferation, self-renewal, and maintenance of the undifferentiated state of spermatogonia
In mammals, primordial germ cells (PGCs) and prospermatogonia (ProSG) transform into SSCs under certain microenvironmental conditions.10 SSCs maintain long-term male fertility via a highly controlled process of self-renewal and maintenance of the undifferentiated state. Spermatogonial self-renewal is ensured by three elements: maintained survival, stable proliferation, and a continuous state of undifferentiation. These three elements are not isolated but are completely interdependent.
Testes of adult Stra8 KO mice are much smaller than those of age-matched WT mice, lacking meiotic and postmeiotic cells.11,12 However, aged testes from Stra8 KO mice progressively enlarged compared with WT testis, in which seminiferous tubules were filled with type A spermatogonia which even spilled into the interstitium due to the presence of only a few tubule structures. The absence of Stra8 gave rise to the accumulation of type A spermatogonia.1 Saba et al.6 showed that Stra8 was activated in testes containing the Cyp26b1 mutation and germ cells entered meiosis prematurely. Cyp26b1 and Stra8 double KO testes display a disruption of meiosis but increased mitotic activity. These findings suggest that Stra8 plays a role in spermatogonial proliferation and differentiation.
Rhox10 promotes the initial establishment of SSCs by suppressing the conversion of ProSG into differentiating A spermatogonia.13 Stra8 is expressed in both undifferentiated and differentiated spermatogonia. The expression of Rhox10 is translocated from the cytoplasm to the nucleus, which is similar to the pattern of Stra8 expression. In this study, we noted a marked upregulation of Rhox10 expression in Stra8 KO testes, which indicated the involvement of Stra8 in the establishment of SSCs by coordinating with Rhox10.
We also found obvious upregulation of the expression of Utf1, Pou5f1, and Upp1 in Stra8−/− testes, while Inca1 expression was downregulated. Utf1 plays a role in the prevention of embryonic stem (ES) cell differentiation. Pou5f1 plays an essential role in the maintenance of the stem cell state and induces rapid proliferation of ES cells through binding to the regulatory region of Utf1 via a variant octamer sequence together with Sox-2 leading to activation of Utf1.14,15,16,17 Inca1 is involved in regulating cell cycle, detecting and repairing gene damage, and preventing abnormal proliferation.18,19 Targeted mutation of Upp1 causes disruption of cell proliferation, uracil DNA damage, and p53 activation.20 Utf1, Pou5f1, Inca1 and Upp1 are all associated with cell cycle progression and cell proliferation. It can be speculated that Stra8 cooperates with these genes in stable proliferation and self-renewal of spermatogonia.
In male mice, Nanos3 is fully downregulated before differentiation of type B spermatogonia and plays important roles in both the survival and maintenance of undifferentiated spermatogonia.21,22 Moreover, Egr4 was highly expressed in SSCs, and decreased rapidly during differentiation, which indicated the involvement of Egr4 in maintaining the SSC population.23 With the first appearance of Stra8 expression in the late undifferentiated spermatogonia,1 the expression of Nanos3 and Egr4 increased significantly in Stra8−/− testes. It can be speculated that this pattern of expression has an effect on the maintenance of undifferentiated SSCs. Furthermore, it is likely that Stra8 performs functions in the maintenance of the undifferentiated state through the regulation of Nanos3 and Egr4, although the precise details of the regulatory network remain to be established.
The role of Stra8 in the differentiating spermatogonia
When spermatogonia begin to differentiate, the activity of pluripotent genes is downregulated and the self-renewal ability is lost. Once differentiation is initiated, it is irreversible. The expression level of Sohlh1 was significantly increased in Stra8 KO testes. Sohlh1 is critical for spermatogonial differentiation and is expressed mainly in differentiating spermatogonia. Sohlh1 and Sohlh2 function cooperatively and directly to repress Stra8 expression by binding to E-Boxes in its promoter.8,24 Rhox10 is detected during the whole process of spermatogonial differentiation.13,25 Stra 8-deficient testes contain a significantly reduced proportion of type B spermatogonia.1,3,4 Stra8 may promote differentiation of spermatogonia through regulation of Sohlh1, Rhox10, and other molecules to ensure Stra8 downregulation during the mid-end stages of meiotic prophase I.
The role of Stra8 in meiotic prophase I
In mammalian spermatogenesis, meiosis is a specialized cellular process. Stra8, a marker of meiosis initiation, is required for germ cells to undergo meiotic prophase. Stra8−/− testes exhibit heterosynapsis or extensive asynapsis.11 In addition, Stra8 is reported to be a potential component of an unidentified cell cycle surveillance system, such as a checkpoint, in response to errors in chromosome synapsis or recombination.12 Many studies have demonstrated the critical role of Stra8 in meiosis; however, the underlying mechanism of spermatogonial differentiation and meiotic initiation remains to be elucidated. In the current study, we found that the expression levels of some meiosis-related genes changed significantly during spermatogenesis, which may provide a theoretical basis for further investigations of the functions of Stra8.
Stra 8-deficiency results in arrest of spermatogenesis in leptotene prophase and failure of the transition to the zygotene/pachytene stages. The expression levels of Prdm9 and Msh5 were significantly reduced in Stra8 KO testes. Prdm9, which plays a key role in the initiation of meiotic events, was highly expressed in preleptonema and early leptonema. Male Prdm9-deficient mice are infertile, with meiosis arrested before the mid-pachytene stage.26,27 Msh5 is highly expressed at the early preleptotene to zygotene stages, with lower expression at the pachytene stage. In Msh5 mutant mice, meiosis is arrested around the zygotene stage, with a high level of asynapsis and persistent unrepaired DNA double-strand breaks.28,29 The Stra8 expression pattern is consistent with that of Prdm9 and Msh5 in spermatocytes. We postulate that Stra8 directly or indirectly targets the regulation of Prdm9 and Msh5 to ensure the successful progress of meiosis prophase I.
We also identified significant changes in the expression of some other meiosis-associated genes. Both Egr423,30 and Mtl5 (tesmin)31 are cytoplasmic and nuclear shuttle proteins that predominantly function in the early stage of pachytene spermatocytes. Genecards software analysis shows that Egr4 binds to the promoter of the CCDC155 gene. Ccdc155 (Kash5) is localized predominantly at the telomere from the leptotene to diplotene stages in spermatocytes. Kash5 combines with SUN1 to form the SUN-Kash nuclear envelope bridge, which mediates the meiotic homolog pairing.32 Dmc1 is essential for the correct execution of meiotic recombination, and spermatogenesis in Dmc1-mutated testes progresses only to the late leptotene and early pachytene.33,34 Pxt1 interacts with the apoptosis regulator BAT3, and c-myc-Pxt1 transgenic mouse testes revealed arrest of spermatogenesis in pachytene spermatocytes.35 Asb9, a specific marker of active spermatogenesis, is strongly expressed in pachytene spermatocytes and round spermatids. However, its function in spermatocytes has not yet been reported.36
In Stra8 KO testes, the expression of Egr4, Pxt1, and Asb9 was significantly upregulated, while Ccdc155, Dmc1, and Mtl5 expression was downregulated. Interestingly, Stra8 is not present in pachytene spermatocytes and there is no evidence that Stra8 directly regulates these genes. We hypothesize that a defect in meiosis I indirectly results in changes in the expression of these genes. A recent study37 offered another possible explanation for this change based on the known ability of RA to regulate spermatogonial differentiation and meiotic initiation. Endo et al.37 demonstrated that RA also regulates the initiation of spermatid elongation and release of spermatozoa. The indirect regulation of pachytene spermatocyte-related genes by Stra8 may be due, in part, to RA produced by pachytene spermatocytes.
The role of Stra8 in physiological processes
Phlda2 is negatively correlated with cell migration and invasion and inhibits glycogen accumulation.38 Foxf1 promotes angiogenesis and regulates embryonic and pluripotent stem cell differentiation.39 Energy homeostasis is regulated mainly by factors produced in the hypothalamus and the brainstem. Nkx1-1 (homeobox gene Sax2), which is a key regulator of organ development, maintains the steady state of energy homeostasis.40
Studies to date have focused on the functions of Phlda2, Foxf1, and Nkx1-1 in embryonic development, angiogenesis, and energy homeostasis. Currently, there is no evidence of a direct link between Phlda2, Foxf1, Nkx1-1, and survival or self-renewal of spermatogonia. However, expression of these genes at the mRNA level increased in Stra8−/− mice, leading us to postulate a role in attachment of spermatogonia to the seminiferous tubule membrane, provision of energy for cell proliferation in mitosis, and maintenance of spermatogonia survival, with a possible indirect role in spermatogenesis.
Moreover, although Mobp,41 Rrad,42 and Clic643,44 were not directly related to spermatogenesis, their expression was directly related to Stra8, with an obvious increase in Stra 8-deficent testes. Spermatogenesis is a complex and highly precise physiological processes, which may involve many unknown mechanisms that require these molecules although this remains to be established.
CONCLUSION
In this study, we identified 21 DEGs related to spermatogenesis that possibly coordinate with Stra8 in this process. Combined with analysis of relevant reports, we provide evidence indicating that Stra8 is a multifunctional protein involved in spermatogenesis. In addition to the major function in spermatogonial differentiation, Stra8 is also implicated in meiotic initiation, the establishment and maintenance of SSCs, cell proliferation, self-renewal, undifferentiation, and other physiological processes. The identification of Stra8-related genes and establishment of its regulatory network provides an improved understanding of the role of Stra8 in spermatogenesis, as well as a theoretical basis for its involvement in mammalian reproduction and the implications of dysregulation on germline tumors and infertility.
AUTHOR CONTRIBUTIONS
HTM designed and carried out experiments; CMN wrote article and performed the experiments; JX, XYS, and MMX collected literatures and analyzed data; YQH participated in the design of the study; and YZ designed the study. All authors read and approved the final version of the manuscript.
COMPETING INTERESTS
All authors declared no competing interests.
ACKNOWLEDGMENTS
This work was supported by the National Natural Science Foundation of China (Number 31371174), the Natural Science Foundation of Jiangsu Province, China (Number BK20131230), and the Postgraduate Research and Practice Innovation Program of Jiangsu Province, China (KYCX17-1893). We thank Dr. Wilkinson (University of California, USA) for kindly providing Rhox10 antibody.
REFERENCES
- 1.Endo T, Romer KA, Anderson EL, Baltus AE, de Rooij DG, et al. Periodic retinoic acid-STRA8 signaling intersects with periodic germ-cell competencies to regulate spermatogenesis. Proc Natl Acad Sci U S A. 2015;112:2347–56. doi: 10.1073/pnas.1505683112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhou Q, Nie R, Li Y, Friel P, Mitchell D, et al. Expression of stimulated by retinoic acid gene 8 (Stra8) in spermatogenic cells induced by retinoic acid: an in vivo study in Vitamin A-sufficient postnatal murine testes. Biol Reprod. 2008;79:35–42. doi: 10.1095/biolreprod.107.066795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Oulad-Abdelghani M, Bouillet P, Décimo D, Gansmuller A, Heyberger S, et al. Characterization of a premeiotic germ cell-specific cytoplasmic protein encoded by Stra8, a novel retinoic acid-responsive gene. J Cell Biol. 1996;135:469–77. doi: 10.1083/jcb.135.2.469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Feng CW, Bowles J, Koopman P. Control of mammalian germ cell entry into meiosis. Mol Cell Endocrinol. 2014;382:488–97. doi: 10.1016/j.mce.2013.09.026. [DOI] [PubMed] [Google Scholar]
- 5.Potter SJ, DeFalco T. Role of the testis interstitial compartment in spermatogonial stem cell function. Reproduction. 2017;153:151–62. doi: 10.1530/REP-16-0588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Saba R, Wu Q, Saga Y. CYP26B1 promotes male germ cell differentiation by suppressing STRA8-dependent meiotic and STRA8-independent mitotic pathways. Dev Biol. 2014;389:173–81. doi: 10.1016/j.ydbio.2014.02.013. [DOI] [PubMed] [Google Scholar]
- 7.Chen W, Jia W, Wang K, Si X, Zhu S, et al. Distinct roles for CBP and p300 on the RA-mediated expression of the meiosis commitment gene Stra8 in mouse embryonic stem cells. PLoS One. 2013;8:e66076. doi: 10.1371/journal.pone.0066076. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Desimio MG, Campolo F, Dolci S, De Felici M, Farini D. SOHLH1 and SOHLH2 directly down-regulate STIMULATED BY RETINOIC ACID 8 (STRA8) expression. Cell Cycle. 2015;14:1036–45. doi: 10.1080/15384101.2015.1007721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhang T, Murphy MW, Gearhart MD, Bardwell VJ, Zarkower D. The mammalian doublesex homolog DMRT6 coordinates the transition between mitotic and meiotic developmental programs during spermatogenesis. Development. 2014;141:3662–71. doi: 10.1242/dev.113936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McCarrey JR. Toward a more precise and informative nomenclature describing fetal and neonatal male germ cells in rodents. Biol Reprod. 2013;89:47. doi: 10.1095/biolreprod.113.110502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Anderson EL, Baltus AE, Roepers-Gajadien HL, Hassold TJ, de Rooij DG. Stra8 and its inducer, retinoic acid, regulate meiotic initiation in both spermatogenesis and oogenesis in mice. Proc Natl Acad Sci U S A. 2008;105:14976–80. doi: 10.1073/pnas.0807297105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Baltus AE, Menke DB, Hu YC, Goodheart ML, Carpenter AE, et al. In germ cells of mouse embryonic ovaries, the decision to enter meiosis precedes premeiotic DNA replication. Nat Genet. 2006;38:1430–4. doi: 10.1038/ng1919. [DOI] [PubMed] [Google Scholar]
- 13.Song HW, Bettegowda A, Lake BB, Zhao AH, Skarbrevik D, et al. The homeobox transcription factor RHOX10 drives mouse spermatogonial stem cell establishment. Cell Rep. 2016;17:149–64. doi: 10.1016/j.celrep.2016.08.090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kooistra SM, van den Boom V, Thummer RP, Johannes F, Wardenaar R, et al. Undifferentiated embryonic cell transcription factor 1 regulates ESC chromatin organization and gene expression. Stem Cells. 2010;28:1703–14. doi: 10.1002/stem.497. [DOI] [PubMed] [Google Scholar]
- 15.Lin CH, Yang CH, Chen YR. UTF1 deficiency promotes retinoic acid-induced neuronal differentiation in P19 embryonal carcinoma cells. Int J Biochem Cell Biol. 2012;44:350–7. doi: 10.1016/j.biocel.2011.11.008. [DOI] [PubMed] [Google Scholar]
- 16.Niwa H, Masui S, Chambers I, Smith AG, Miyazaki J. Phenotypic complementation establishes requirements for specific POU domain and generic transactivation function of Oct-3/4 in embryonic stem cells. Mol Cell Biol. 2002;22:1526–36. doi: 10.1128/mcb.22.5.1526-1536.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Nishimoto M, Miyagi S, Yamagishi T, Sakaguchi T, Niwa H, et al. Oct-3/4 maintains the proliferative embryonic stem cell state via specific binding to a variant octamer sequence in the regulatory region of the UTF1 locus. Mol Cell Biol. 2005;25:5084–94. doi: 10.1128/MCB.25.12.5084-5094.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bäumer N, Tickenbrock L, Tschanter P, Lohmeyer L, Diederichs S, et al. Inhibitor of cyclin-dependent kinase (CDK) interacting with cyclin A1 (INCA1) regulates proliferation and is repressed by oncogenic signaling. J Biol Chem. 2011;286:28210–22. doi: 10.1074/jbc.M110.203471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li XB, Chen J, Deng MJ, Wang F, Du ZW, et al. Zinc finger protein HZF1 promotes K562 cell proliferation by interacting with and inhibiting INCA1. Mol Med Rep. 2011;4:1131–7. doi: 10.3892/mmr.2011.564. [DOI] [PubMed] [Google Scholar]
- 20.Cao Z, Ma J, Chen X, Zhou B, Cai C, et al. Uridine homeostatic disorder leads to DNA damage and tumorigenesis. Cancer Lett. 2016;372:219–25. doi: 10.1016/j.canlet.2016.01.007. [DOI] [PubMed] [Google Scholar]
- 21.Suzuki H, Sada A, Yoshida S, Saga Y. The heterogeneity of spermatogonia is revealed by their topology and expression of marker proteins including the germ cell-specific proteins Nanos2 and Nanos3. Dev Biol. 2009;336:222–31. doi: 10.1016/j.ydbio.2009.10.002. [DOI] [PubMed] [Google Scholar]
- 22.Suzuki H, Tsuda M, Kiso M, Saga Y. Nanos3 maintains the germ cell lineage in the mouse by suppressing both Bax-dependent and -independent apoptotic pathways. Dev Biol. 2008;318:133–42. doi: 10.1016/j.ydbio.2008.03.020. [DOI] [PubMed] [Google Scholar]
- 23.Hogarth CA, Mitchell D, Small C, Griswold M. EGR4 displays both a cell- and intracellular-specific localization pattern in the developing murine testis. Dev Dyn. 2010;239:3106–14. doi: 10.1002/dvdy.22442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Toyoda S, Yoshimura T, Mizuta J, Miyazaki J. Auto-regulation of the Sohlh1 gene by the SOHLH2/SOHLH1/SP1 complex: implications for early spermatogenesis and oogenesis. PLoS One. 2014;9:e101681. doi: 10.1371/journal.pone.0101681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Song HW, Dann CT, McCarrey JR, Meistrich ML, Cornwall GA. Dynamic expression pattern and subcellular localization of the Rhox10 homeobox transcription factor during early germ cell development. Reproduction. 2012;143:611–24. doi: 10.1530/REP-11-0479. [DOI] [PubMed] [Google Scholar]
- 26.Sun F, Fujiwara Y, Reinholdt LG, Hu J, Saxl RL, et al. Nuclear localization of PRDM9 and its role in meiotic chromatin modifications and homologous synapsis. Chromosoma. 2015;124:397–415. doi: 10.1007/s00412-015-0511-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Parvanov ED, Tian H, Billings T, Saxl RL, Spruce C, et al. PRDM9 interactions with other proteins provide a link between recombination hotspots and the chromosomal axis in meiosis. Mol Biol Cell. 2017;28:488–99. doi: 10.1091/mbc.E16-09-0686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Lu X, Liu X, An L, Zhang W, Sun J, et al. The arabidopsis MutS homolog AtMSH5 is required for normal meiosis. Cell Res. 2008;18:589–99. doi: 10.1038/cr.2008.44. [DOI] [PubMed] [Google Scholar]
- 29.Mahadevaiah SK, Bourc’his D, de Rooij DG, Bestor TH, Turner JM, et al. Extensive meiotic asynapsis in mice antagonises meiotic silencing of unsynapsed chromatin and consequently disrupts meiotic sex chromosome inactivation. J Cell Biol. 2008;182:263–76. doi: 10.1083/jcb.200710195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tourtellotte WG, Nagarajan R, Auyeung A, Mueller C, Milbrandt J. Infertility associated with incomplete spermatogenic arrest and oligozoospermia in Egr4-deficient mice. Development. 1999;126:5061–71. doi: 10.1242/dev.126.22.5061. [DOI] [PubMed] [Google Scholar]
- 31.Olesen C, Møller M, Byskov AG. Tesmin transcription is regulated differently during male and female meiosis. Mol Reprod Dev. 2004;67:116–26. doi: 10.1002/mrd.20007. [DOI] [PubMed] [Google Scholar]
- 32.Morimoto A, Shibuya H, Zhu X, Kim J, Ishiguro K, et al. A conserved KASH domain protein associates with telomeres, SUN1, and dynactin during mammalian meiosis. J Cell Biol. 2012;198:165–72. doi: 10.1083/jcb.201204085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Borgogno MV, Monti MR, Zhao W, Sung P, Argaraña CE, et al. Tolerance of DNA mismatches in Dmc1 recombinase-mediated DNA strand exchange. J Biol Chem. 2016;291:4928–38. doi: 10.1074/jbc.M115.704718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Cloud V, Chan YL, Grubb J, Budke B, Bishop DK. Rad51 is an accessory factor for Dmc1-mediated joint molecule formation during meiosis. Science. 2012;337:1222–5. doi: 10.1126/science.1219379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Forand A, Bernardino-Sgherri J. A critical role of PUMA in maintenance of genomic integrity of murine spermatogonial stem cell precursors after genotoxic stress. Cell Res. 2009;19:1018–30. doi: 10.1038/cr.2009.50. [DOI] [PubMed] [Google Scholar]
- 36.Lee MR, Kim SK, Kim JS, Rhim SY, Kim KS. Expression of murine Asb-9 during mouse spermatogenesis. Mol Cells. 2008;26:621–4. [PubMed] [Google Scholar]
- 37.Endo T, Freinkman E, de Rooij DG, Page DC. Periodic production of retinoic acid by meiotic and somatic cells coordinates four transitions in mouse spermatogenesis. Proc Natl Acad Sci U S A. 2017;114:10132–41. doi: 10.1073/pnas.1710837114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Tunster SJ, Van De Pette M, John RM. Isolating the role of elevated Phlda2 in asymmetric late fetal growth restriction in mice. Dis Model Mech. 2014;7:1185–91. doi: 10.1242/dmm.017079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kalinichenko VV, Gusarova GA, Shin B, Costa RH. The forkhead box F1 transcription factor is expressed in brain and head mesenchyme during mouse embryonic development. Gene Expr Patterns. 2003;3:153–8. doi: 10.1016/s1567-133x(03)00010-3. [DOI] [PubMed] [Google Scholar]
- 40.Simon R, Lufkin T, Bergemann AD. Homeobox gene Sax2 deficiency causes an imbalance in energy homeostasis. Dev Dyn. 2007;236:2792–9. doi: 10.1002/dvdy.21320. [DOI] [PubMed] [Google Scholar]
- 41.Laursen KB, Wong PM, Gudas LJ. Epigenetic regulation by RARα maintains ligand-independent transcriptional activity. Nucleic Acids Res. 2012;40:102–15. doi: 10.1093/nar/gkr637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Manning JR, Yin G, Kaminski CN, Magyar J, Feng HZ, et al. Rad GTPase deletion increases L-type calcium channel current leading to increased cardiac contraction. J Am Heart Assoc. 2013;2:e000459. doi: 10.1161/JAHA.113.000459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Friedli M, Guipponi M, Bertrand S, Bertrand D, Neerman-Arbez M, et al. Identification of a novel member of the CLIC family, CLIC6, mapping to 21q22.12. Gene. 2003;320:31–40. doi: 10.1016/s0378-1119(03)00830-8. [DOI] [PubMed] [Google Scholar]
- 44.Griffon N, Jeanneteau F, Prieur F, Diaz J, Sokoloff P. CLIC6, a member of the intracellular chloride channel family, interacts with dopamine D (2)-like receptors. Brain Res Mol Brain Res. 2003;117:47–57. doi: 10.1016/s0169-328x(03)00283-3. [DOI] [PubMed] [Google Scholar]