

Abstract
Background
Face processing has been found to be impaired in autism spectrum disorders (ASD). One hypothesis is that individuals with ASD engage in piecemeal compared to holistic face processing strategies. To investigate the role of possible impairments in holistic face processing in individuals with autism, the current study investigated behavioral and electroencephalography (EEG) correlates of face processing (P1/N170 and gamma-band activity) in adolescents with ASD and sex-, age-, and IQ-matched neurotypical controls.
Methods
Participants were presented with upright and inverted Mooney stimuli; black and white low information faces that are only perceived as faces when processed holistically. Participants indicated behaviorally the detection of a face. EEG was collected time-locked to the presentation of the stimuli.
Results
Adolescents with ASD perceived Mooney stimuli as faces suggesting ability to use holistic processing but displayed a lower face detection rate and slower response times. ERP components suggest slowed temporal processing of Mooney stimuli in the ASD compared to control group for P1 latency but no differences between groups for P1 amplitude and at the N170. Increases in gamma-band activity was similar during the perception of the Mooney images by group, but the ASD group showed prolonged temporal elevation in activity.
Conclusion
Overall, our results suggest that adolescents with ASD were able to utilize holistic processing to perceive a face within the Mooney stimuli. Delays in early processing, marked by the P1, and elongated elevation in gamma activity indicate that the neural systems supporting holistic processing are slightly altered suggesting a less automatic and less efficient facial processing system.
Trial registration
Non-applicable.
Electronic supplementary material
The online version of this article (10.1186/s11689-018-9244-y) contains supplementary material, which is available to authorized users.
Keywords: ASD, Gamma-band activity, Holistic face processing, P1, N170
Background
The processing of social information in faces is crucial to communicate effectively with others [1, 2]. Faces possess two types of configural information: first-order information (repeated in every face; e.g., two eyes, above a nose, above a mouth) to enable early face detection [3, 4], and emerging second-order properties (variations in spacing between the features) to extract inter-face variance and to discriminate between faces [3, 5–7]. In configural processing, a face is therefore perceived from lower features to emergent features. In contrast, holistic processing assumes that faces are perceived immediately as undifferentiated wholes without going from first-to second-order features [7]. Configural and holistic processing have been assumed to play parallel roles within face processing [8].
Further interest in face processing is fueled by neurodevelopmental conditions such as autism spectrum disorders (ASDs), which are characterized by early and pervasive social communication and interaction impairments [9]. Individuals with ASD show an enhanced reliance on, or a greater scanning of, unusual face parts (i.e., mouth instead of eyes) [10–12]. The integration of visual information into a meaningful whole may be impeded by processing predominantly first- rather than second-order features leading to a part-based processing style [13–15].
Face inversion paradigms have been used to examine holistic and configural processing in ASD (e.g., [15]). The inversion of a human face may disrupt configural processing [16, 17]. The extraction of first-order information remains intact regardless of stimulus’ orientation [3]. In face inversion tasks, accuracy rates for upright faces compared to inverted faces were higher for neurotypical controls [5]. For upright stimuli, holistic and configural strategies may work together, which contributes to higher accuracy rates, whereas a stronger reliance on first-order features is necessary for inverted faces, which contributes to lower accuracy levels. In contrast, individuals with ASD displayed similar detection rates for upright and inverted faces [18]. This pattern of results supports the idea of a part-based processing strategy in which individuals with ASD predominantly rely on first-order information for upright and inverted face stimuli. Reaction time analyses of face inversion paradigms complement these findings as controls are faster in making their decisions compared to individuals with ASD (e.g., [19]). There have been, however, contrasting results which demonstrate similar face detecting rates in both groups [15, 20, 21] or even better performance in the ASD group compared to controls [20]. In fact, a recent systematic review suggested an intact face inversion effect for the ASD group in most studies [22].
Individuals with ASD may engage in similar face processing strategies as controls [12], but due to a lack of attention to faces from an early age [2], individuals with ASD may develop less expertise in face identification and discrimination [23]. Similar face detection rates for controls and the ASD group were also observed when cueing to relevant parts of the face [10]. Researchers have therefore suggested a quantitative instead of a qualitative difference of face perception in ASD [22, 24].
The disentanglement of holistic and configural processes is another challenge of face inversion tasks [7]. To address this, studies employ the Mooney face task to specifically trigger holistic processes [16, 19, 25, 26]. Mooney stimuli give rise to faces by the two-tone composition of black and white parts [16]. Extensive binding and holistic processes are required to perceive them as faces because they contain few explicit local features [25, 27, 28]. Upright presented Mooney stimuli are thought to recruit more efficient holistic processes, whereas inverted Mooney stimuli severely hinder face abstraction [8, 29]. As before, some studies reported a face inversion effect with Mooney stimuli for the ASD group (e.g., [20]), while others failed to find it (e.g., [19]).
Part of the discrepancy in these result patterns may be related to the inclusion of individuals spanning broad age bands. Holistic face processing was suggested to be impaired in children with ASD (aged 8 to 13 years) who displayed lower accuracy for inverted compared to upright face stimuli [21] and showed less sensitivity to configuration of the faces potentially due to holistic processing deficit or a lack of expertise with faces [30]. McPartland et al. (2004) demonstrated ERP differences in basic face vs. house comparisons in adolescents and adults 15 to 42 years; however, Webb et al. (2012) in adults 18-to-44-year-olds with ASD did not find altered face vs. house ERP activity but did find differences between groups in face inversion processing [23]. In a sample of 9-to 45-year-olds with ASD, O’Connor et al. (2005) found that the younger group with ASD (9 to 15 years) displayed no difference in task performance (compared to the controls), whereas adults with ASD (18 to 45 years) showed deficits across all emotion categories, which the authors suggested reflected indicating a general facial configuration deficit for adults with ASD [17]. One possible source of confound in these papers is the inclusion of the transitional stage of adolescence within either the child or adult groups. It may be of importance to examine manifestations and trajectories of face processing differences separately for adolescents, particularly as orientation processing and some aspects of holistic processing may become mature in childhood (e.g., [31]), but other neural markers of face sensitivity do not become mature until late-adolescence [32].
General face processing differences in adolescents with and without ASD may be represented by altered patterns at the neural level, specifically in the ERP components P1 and N170 which reflect attentional and perceptual aspects of the neural circuitry of face perception [1, 12, 33, 34]. Of importance, based on the latency of these components, this neural activity often precedes behavioral responses about face stimuli. The P1 event-related potential (ERP) component is a positive deflection around 100 ms associated with visual attention [35, 36]. In children and young adolescents with ASD compared to controls, Hileman et al. (2011) found smaller P1 amplitudes (but not latency) for inverted compared to upright faces while Neuhaus et al. (2016) found an inversion effect in the control group for latency (but not amplitude) which was not apparent in children and adolescents with ASD [37]. Within an adult sample, differences in P1 amplitude (but not latency) for inversion across groups were reported [38].
The N170 component reflects face categorical processing (relative to other objects), as well as eye featural sensitivity [39, 40]. In 3- to 4- and 3- to 6-year-old children with ASD, N170 latencies were longer and amplitudes smaller compared to controls in face vs. object perception tasks [2, 41]. Studies of early and late adolescence in ASD displayed a similar pattern of delayed N170 latencies without differences in N70 amplitudes to faces compared to controls [42], but this was not found in another report [37]. It is possible, that the inconsistent finding of a face inversion effect in behavior and ERPs is also associated with the underlying developmental trajectory of holistic processing and with different stimulus types and comparisons altering the extent to which the sources contributing to the P1 or N170 are implicated.
Additional EEG signal properties may inform our understanding of the mechanisms of holistic processing. The rhythmic synchronization of neural discharges in the gamma-band (> 25 Hz) relates to the ‘binding problem’ that is, the question of how various visual features are integrated to a coherent object representation [26, 43, 44]. It is associated with the pyramidal network’s synchronization of excitatory and inhibitory interneurons [43]. Gamma-band activity (GBA) has also been connected to working memory and visual attention processes [43, 45]. The match-and-utilization model (MUM) predicts that meaningful objects such as upright faces lead to stronger GBA compared to inverted faces [46]. GBA in the lower range (25–45 Hz, 150–250 ms) has been shown to be sensitive to inversion of faces with lower activity for inverted compared to upright faces [39] and greater for faces compared to scrambled faces in neurotypical controls around 200 ms [4, 47]. Adults with ASD displayed lower levels of GBA in the lower gamma-band over occipital areas within a passive face viewing task with peak differences between 250 and 450 ms [45] or during a Mooney face inversion task between 100 and 300 ms [19]. GBA of adults with ASD was not sensitive to inversion of face in the lower gamma-band range at frontal sites, whereas controls showed a larger burst for upright faces [48]. These abnormalities in GBA may underlie disruptions in face processing in ASD at a very basic level [45].
Taken together, there are documented differences in face processing in behavioral and neural activity in children and adults with ASD but less is known about holistic face processing during adolescence as most studies have included adolescents either with younger or adult participants rather than as a targeted group. This may be an age period of particular importance as the P1 and N170 (amplitude, latency, and response characteristics) as a marker of early stage face processing becomes adult-like in the quality of the response pattern but still quantitatively differs in amplitude and latency [32]. Therefore, we aimed at investigating behavioral (detection rate/response times) and neurophysiological correlates (P1/N170 component/gamma-band activity) of holistic face processing in a narrow range sample of adolescents with ASD and sex-, age-, and IQ-matched neurotypical controls. EEG was collected while adolescents completed an inversion task with Mooney stimuli.
If adolescents with neurotypical development show effective holistic processes, and in contrast, holistic processing is impaired in the ASD group, then we predict that (1) slower response times as well as reduced face detection rates would occur in the ASD group compared to controls, suggestive of reduced holistic face perception and stronger focus on first-order features. (2) P1 latency would not be modulated by Mooney stimuli detected as faces, whereas P1/N170 amplitudes and N170 latencies to Mooney stimuli detected as faces would be slower and of less amplitude in ASD compared to controls. (3) Controls but not the ASD group would display greater P1/N170 amplitude and faster N170 latency to stimuli perceived as face compared to non-face responses. (4) Gamma power in the lower gamma-band range (25–45 Hz; associated with perceptual binding) would be smaller in the ASD compared to the control group in early and late time windows for anterior and posterior clusters.
Methods
Participants
The local Institutional Review Board approved the protocol, all adolescents provided written assent, and a parent provided written consent for participation. Adolescents with ASD met research diagnostic criteria based on the Autism Diagnostic Observation Schedule (ADOS) [49], criteria on the social and communication domains of the Autism Diagnostic Interview-Revised (ADI-R) [50], and DSM-IV criteria based on expert clinical diagnostic judgment [51]. Adolescents with typical development had no history of developmental delay or concerns about autism-related behaviors. Exclusionary criteria for adolescents with ASD and controls included performance IQ scores < 80 (Wechsler Intelligence Scale III; WISC), known genetic disorders, seizures, significant sensory or motor impairment, major physical abnormalities, serious head injury, and use of anticonvulsant or barbiturate medications. Performance IQ was employed as criteria because the tasks across the full protocol focused on non-verbal visual processing. Additional exclusionary criteria for controls included birth or developmental abnormalities, psychotropic medication usage, and a first-degree relative with ASD. Sixty-eight adolescents were enrolled in the study. Participants were matched based on their age and sex followed by bin-matching with regard to their performance IQ during the screening session. Thirty participants were excluded from the final analysis: 8 participants were disqualified after enrollment (non-compliance or too low IQ), 8 datasets had EEG file errors that resulted in unusable data, 6 had significant EEG artifacts (e.g., excessive movement), and 8 did not show visible ERP components after averaging. The final sample consisted of 19 controls and 19 participants with ASD. No group differences for age, gender, or performance IQ were detected. There were no significant differences in characteristics between those that were included in the analysis and those that were not (ps > .05). Demographic characteristics are provided in Table 1.
Table 1.
Means and standard deviations for gender, age, and IQ scores of controls and the ASD group
| Controls (N = 19) | ASD group (n = 19) | χ 2/t value | p value | |||
|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | |||
| Gender (M:F) | 16:3 | 16:3 | χ 2(1) = 0.000 | 1.000 | ||
| Age (years) | 13.950 | 1.268 | 14.000 | 1.667 | t(36) = 0.110 | .913 |
| P IQ | 112.790 | 16.755 | 115.737 | 14.681 | t(36) = 0.577 | .568 |
| FS IQ | 113.630 | 17.150 | 109.370 | 13.039 | t(36) = − 0.863 | .394 |
Note. ASD = autism spectrum disorder, P IQ = Wechsler Intelligence Scale III Performance IQ, FS IQ = Wechsler Intelligence Scale III Full Performance, SD = standard deviation
Apparatus and stimuli
The current study used a set of 50 Mooney face stimuli (5.9° by 7.9°), which are degraded, 2-tone pictures of human faces [52] (see Fig. 1, Mooney face stimulus examples). They were presented upright and inverted to manipulate holistic face perception.
Fig. 1.
Examples of upright (a) and inverted (b) Mooney face stimuli
Procedure
Adolescents completed a training block, consisting of four trials in which Mooney stimuli were either presented upright (n = 2) or inverted (n = 2). They were asked to indicate whether they perceived a face or not. During the training trials, the goal was to practice mapping the right/left button press to the decision of face/no face. After the mapping was understood, adolescents started with the actual task in which they saw a random sequence of upright and inverted Mooney stimuli. Participants were asked to answer as spontaneously and quickly as possible. Face and no face button position was balanced across participants.
The experiment consisted of 200 trials, presented in four 50 image blocks. A break of participant-determined length separated each block. In each trial, a gray background was presented for 500 ms (baseline) followed by a Mooney stimulus for 500 ms displayed on a gray background. The inter-trial interval (ITI) varied between 2000 and 2500 ms. Adolescents could indicate their decision across the entire stimulus presentation and ITI.
Electrophysiological recordings
EEG was recorded with a 128-channel Geodesic Sensor NetAmps 200 in Net Station 2.0 (Electric Geodesic, Inc. Eugene OR), with a sampling rate of 500 Hz, and experimental control through E-Prime 1.0 software. In a dimly-lit, sound-attenuated room, adolescents sat approximately 24 inches from the stimulus monitor and used buttons 1 (left most button, left index finger) and 5 (right most button, right index finger) on a 5-button box for experimental response.
Processing
All procedures were conducted with MATLAB’s Toolbox EEGLAB (The MathWorks, Natick, MA). Re-sampling of the data to 250 Hz and filtering (0.1 Hz highpass; 100 Hz lowpass; 60 Hz notch) preceded the exclusion of bad channels (impedances over 200 KOhm, drifting channels). Data was re-referenced to average reference, segmented into epochs (− 500 to 1000 ms) for each condition and baseline-corrected to 500 ms pre-stimulus interval. Hand editing was done as a first artifact rejection step to address “non-stereotyped” noise (e.g., pulling the cap) prior to conducting an independent component analysis (ICA). With the help of the EEGLAB plugin SASICA [53], components such as those containing electrical noise, ocular, or head movements were identified. Visual inspection served as final judgment on rejecting bad components. Lastly, excluded channels were interpolated using spherical interpolation.
ERP data
Based on a study of Webb et al. (2012), amplitudes for the P1 and N170 component in adolescents were chosen from a posterior medial left cluster (electrodes 65, 70, 71, and 75) and a posterior medial right cluster (electrodes 83, 84, 90, and 91; also see Additional file 1). The first positive peak was defined as the P1 component and the N170 component was specified as the first negative deflection following the P1. Temporal windows for extracting the ERP components were visually inspected for developmental shifts in latency, amplitude, and morphology [32, 42]. Overall time windows ranged from 70 to 170 ms (P1) and 120 to 220 ms (N170). Amplitudes and latencies were extracted across the selected clusters within the designated time windows separately for the clusters of the left and right hemisphere for the P1 and N170 component. P1 and N170 components had to be present in 50% of the defined electrode cluster to be further included. Data was separately inspected for upright presented stimuli (trials face response: M = 65.210, SD = 11.928; trials no face response: M = 17.820, SD = 9.320) and inverted presented stimuli (trials face response: M = 44.000, SD = 20.254; trials no faces response: M = 37.530, SD = 15.446). At least 20 trials in a condition were necessary to be included in further analyses. Instead of peak amplitude, mean P1 amplitude was calculated to account for the noise level of the waveform [54]. To account for influences of the preceding P1, adjusted N170 amplitudes and latencies were calculated by subtracting the P1 peak amplitude from the N170 peak amplitude, and the P1 peak latency from the N170 peak latency [55]. Lastly, grand average waveforms were calculated for both groups.
Time-frequency-analysis
Gamma-band power (25–45 Hz) was calculated in 50 linear steps using complex Morlet wavelets (c.f. [56]). The wavelets were defined as , with σf = 1/2πσt, where t is time, f0 is frequency, and where σf and σt denote the length of the wavelet in the frequency and time domain. The ratio f0/σf was set to 5. We focused on induced gamma (i.e., non phase-locked gamma power) by obtaining time-frequency transforms of single epochs first and then averaging them across trials for each condition (c.f. [57, 58]). The time-frequency data was normalized to baseline (− 350 to − 50 ms) by applying a Z-transform, where the difference between signal and baseline was divided by the standard deviation of the baseline according to formula (1):
in which Z denotes Z value, t denotes time, f frequency, and n denotes the number of time points in the baseline. Z values from the electrode clusters of interest included the P1/N170 posterior inferior left and right clusters. Based on visual inspection of the scalp map distribution, an anterior left and anterior right cluster was added (left cluster electrodes 19, 23, 24, and 27; right cluster electrodes 2, 3, 9, and 10). Further, based on the time-frequency plots, two time windows were identified for analysis (50–200 ms; 200–350). Signal was averaged separately for clusters across the 25–45 Hz band and for each time window. These values were then averaged across participants for each group.
Statistical analysis
After processing, too few participants had data available for upright Mooney faces not detected as faces. This condition was therefore not included. Thus, we examined the contrasts of face responses to upright and inverted stimuli and face “no face” responses for inverted stimuli within the ERP and gamma analyses.
For face detection rates and reaction times, trials were averaged based on stimulus’ orientation (upright/inverted) for stimuli detected as faces. They were submitted to repeated-measures analyses of variances (ANOVA) with orientation (upright/inverted) as within-factor and group (ASD group/controls) as between-factor.
To contrast ERP responses for face responses to upright and inverted stimuli, mean P1 amplitudes and latencies and adjusted N170 amplitudes and latencies were entered into separate repeated-measures ANOVA including the factors orientation (upright/inverted) and hemisphere (left/right) as within-factors and group (ASD group/controls) as between-factor.
To compare face to no face responses, mean P1 amplitudes and latencies and adjusted N170 amplitudes and latencies were averaged for these categories within the inverted condition. Afterwards, values were submitted to separate repeated-measures ANOVA with percept (face/no face) and hemisphere (left/right) as within-factors and group (ASD group/controls) as between-factor.
To compare GBA responses, a repeated-measures ANOVA with the within factors percept (face/no face), time window (50–200 ms/ 200–350 ms), cluster (anterior/posterior), and hemisphere (left/right) as well as the between factor group (ASD group/controls) was calculated.
All statistical analyses were performed with the IBM (Armank, NY) SPSS Statistics 14.0 software package and MATLAB (The MathWorks, Natick, MA). All analyses were followed up with inclusion of FS IQ or age as a covariate; these covariates did not change the results and findings are reported without the covariates. Significant main effects and interactions were followed by subsequent 1-way ANOVAs for the groups or by post hoc Bonferroni-corrected contrasts. For all analyses, the significance level was set at α < 0.05.
Results
Behavioral performance
Detection rate
As hypothesized, Controls detected significantly more Mooney stimuli as faces compared to participants with ASD (F(1, 36) = 6.272, p < .05, ηp2 = .148). Both groups identified more Mooney stimuli as faces in the upright compared to the inverted presentation (F(1, 36) =37.316, p < .001, ηp2 = .982). There was no orientation × group interaction (F(1, 36) = 0.089, p = .768, ηp2 = .148) (Fig. 2).
Fig. 2.
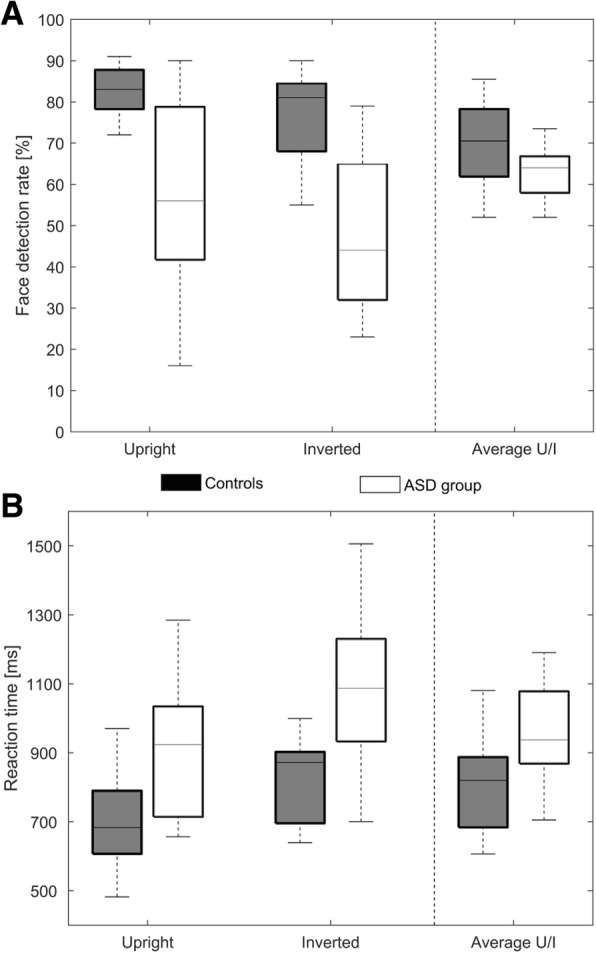
Behavioral performance differences between controls and participants with ASD. Face detection rate in percent (a) and response time in milliseconds (b) for face decisions to upright and inverted Mooney stimuli for controls (black boxplots) and participants with ASD (white boxplots)
Response time
In line with our hypothesis, the control group was faster than the ASD group to detect a face for upright and inverted Mooney stimuli (F(1, 36) = 6.106, p < .05, ηp2 = .145). Both groups were faster to detect an upright Mooney stimulus as opposed to an inverted Mooney stimulus as a face (F(1, 36) = 92.506, p < .001, ηp2 = .720). There was no group x orientation interaction (F(1, 36) = 0.159, p = .692, ηp2 = .004) (Table 2).
Table 2.
Means and standard deviations for face detection rate and reaction times for controls and the participants with ASD
| Controls (n = 19) | ASD group (n = 19) | |||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| Hits upright stimuli (hus) (%) | 81.250 | 8.097 | 75.474 | 12.607 |
| Hits inverted inverted (his) (%) | 56.200 | 21.279 | 47.737 | 18.624 |
| Reaction time hus (ms) | 829.979 | 196.972 | 952.684 | 221.415 |
| Reaction time his (ms) | 1049.970 | 226.513 | 1188.531 | 221.813 |
Note. ASD = autism spectrum disorder, SD = standard deviation
ERP analysis for face decisions in upright vs. inverted stimuli
P1 latency
In contrast to our hypothesis, a between-group comparison revealed a significant effect for group (F(1, 36) = 5.692, p < .05, ηp2 = .137) with controls compared to the ASD group displaying faster P1 latencies for trials with a face decision. There was no effect of orientation (F(1, 36) = 0.907, p = .347, ηp2 = .025) or interaction of group × orientation (F(1, 36) = 0.004, p = .950, ηp2 = .000). When averaged across group and orientation, no hemisphere differences were detected (F(1, 36) = 2.899, p = .097, ηp2 = .075), nor interactions with group (F(1, 36) = 0.450, p = .507, ηp2 = .012) or orientation (F(1, 36) = 0.529, p = .472, ηp2 = .014). The significant 3-way interaction of group × orientation × hemisphere (F(1, 36) = 9.339, p < .05, ηp2 = .206) led to subsequent 1-way ANOVAs separated for group. None of the contrasts (separated by group) displayed significant outcomes.
P1 amplitude
Contrary to expectations, the ASD group showed similar P1 amplitudes compared to controls when Mooney stimuli were detected as faces (F(1, 36) = 1.068, p = .308, ηp2 = .029). There was no main effect of orientation (F(1, 36) = 1.013, p = .321, ηp2 = .027), nor interaction with group (F(1, 36) = 3.014, p = .091, ηp2 = .077). P1 amplitudes did not differ across hemisphere (F(1, 36) = 0.944, p = .338, ηp2 = .026). No significant interactions of hemisphere × group (F(1, 36) = 0.243, p = .625, ηp2 = .007), hemisphere × orientation (F(1, 36) = 0.000, p = .998, ηp2 = .000), nor hemisphere × group × orientation (F(1, 36) = 1.337, p = .255, ηp2 = .036) were observed.
N170 latency
Faster latencies were expected for controls compared to the ASD group when detecting a face. In contrast to our hypothesis, the ASD group showed similar N170 latencies compared to controls (F(1, 36) = 0.796, p = .378, ηp2 = .022), not influenced by orientation (F(1, 36) = 2.191, p = .147, ηp2 = .057). No group × orientation interaction (F(1, 36) = 0.280, p = .600, ηp2 = .008) or differences between hemispheres (F(1, 36) = 2.338, p = .245, ηp2 = .037) were detected. Hemisphere did not interact with group (F(1, 36) = 1.396, p = .245, ηp2 = .037) or orientation (F(1, 36) = 0.121, p = .730, ηp2 = .335). The 3-way interaction of group × orientation × hemisphere was not significant (F(1, 36) = 3.673, p = .063, ηp2 = .093).
N170 amplitude
Contrary to expectations, controls and participants with ASD showed similar N170 amplitudes when detecting faces within the Mooney stimuli (F(1, 36) = 0.492, p = .488, ηp2 = .013). Orientation did not influence N170 amplitudes (F(1, 36) = 0.393, p = .535, ηp2 = .011) or interact with group (F(1, 36) = 0.780, p = .383, ηp2 = .021). A significant difference between hemispheres (F(1, 36) = 18.135, p < .001, ηp2 = .335) indicated larger N170 amplitudes in the right compared to the left cluster (p < .001). There was no interaction of hemisphere × group (F(1, 36) = 2.031, p = .163, ηp2 = .053), hemisphere × orientation (F(1, 36) = 0.001, p = .979, ηp2 = .000), nor hemisphere × orientation × group (F(1, 36) = 0.155, p = .696, ηp2 = .004).
ERP analysis for face vs. no face decisions in inverted Mooney stimuli
P1 latency
Contrary to expectations, a main effect of group (F(1, 36) = 5.349, p < .05, ηp2 = .129) indicated longer latencies for the ASD group compared to controls across conditions. Latencies were not modulated by percept (F(1, 36) = 0.704, p = 407, ηp2 = .019) or a percept × group interaction (F(1, 36) = 0.028, p = .868, ηp2 = .001), indicating that the latency difference was not due to face detection differences. The effect of hemisphere was not significant (F(1, 36) = 3.249, p = .080, ηp2 = .083), nor did hemisphere interact with percept (F(1, 36) = 0.088, p = .769, ηp2 = .002), group (F(1, 36) = 0.767, p = .387, ηp2 = .021) or display 3-way interaction (F(1, 36) = 2.818, p = .102, ηp2 = .073) (Fig. 3).
Fig. 3.

ERP differences for face and no face decisions between groups. ERP plots are represented for face decisions in upright Mooney stimuli (a), face decisions in inverted Mooney stimuli (b), and no face decisions in inverted Mooney stimuli (c) separately for the left posterior cluster (LPC) and right posterior cluster (RPC) for controls (gray line) and the ASD group (black line)
P1 amplitude
In contrast to our hypothesis, there was no significant main effect of group suggesting that P1 amplitudes for face vs. no face decision did not differ between controls and the ASD group (F(1, 36) = 0.558, p = .460, ηp2 = .015). Percept did not yield a significant effect (F(1, 36) = 0.105, p = .748, ηp2 = .003). A significant percept × group interaction was observed (F(1, 36) = 5.699, p < .05, ηp2 = .137); however, post hoc analyses showed no difference of percept for controls (p = .153) or the ASD group (p = .063). The effect of hemisphere was significant (F(1, 36) = 7.953, p < .05, ηp2 = .181). Amplitudes in the right cluster were larger compared to the left cluster (p > .05). Hemisphere did not interact with group (F(1, 36) = 0.026, p = .872, ηp2 = .001), nor percept (F(1, 36) = 0.977, p = .330, ηp2 = .026), nor a 3-way interaction (F(1, 36) = 1.346, p = .254, ηp2 = .036).
N170 latency
We expected similar N170 latencies for face and no face decisions for the ASD group which were hypothesized to be delayed compared to controls. In contrast to our hypothesis, controls and participants with ASD displayed similar N170 latencies (F(1, 36) = 0.155, p = .696, ηp2 = .004). N170 latencies were not modulated by percept (F(1, 36) = 1.670, p = .204, ηp2 = .044) or percept × group interaction (F(1, 36) = 1.670, p = .204, ηp2 = .044). None of the factors (hemisphere: F(1, 36) = 1.604, p = .213, ηp2 = .043; hemisphere × group: F(1, 36) = 1.313, p = .259, ηp2 = .035; hemisphere × percept: F(1, 36) = 0.018, p = .893, ηp2 = .001, hemisphere × percept × group: F(1, 36) = 1.495, p = .229, ηp2 = .040) reached significance.
N170 amplitude
Contrary to expectations, controls showed similar N170 amplitudes compared to the ASD group (F(1, 36) = 0.922, p = .343, ηp2 = .025). There was no effect of percept (F(1, 36) = 0.019, p = .892, ηp2 = .001), nor did percept interact with group (F(1, 36) = 1.049, p = .313, ηp2 = .028). A significant effect for hemisphere (F(1, 36) = 20.744, p < .001, ηp2 = .366) was observed, indicating that larger N170 values were found within the right cluster (p < .001). There was no interaction of hemisphere × group (F(1, 36) = 0.250, p = .620, ηp2 = .007), hemisphere × percept (F(1, 36) = 0.386, p = .539, ηp2 = .011) or hemisphere × percept × group (F(1, 36) = 0.566, p = .121, ηp2 = .065).
Summary ERP analysis
To summarize our ERP results, controls and individuals with ASD showed similar P1 and N170 morphologies. They only differed with regard to their P1 latencies. Controls displayed faster latencies than the ASD group for face decisions across inverted and upright Mooney stimuli and for face vs. no face decision for inverted Mooney stimuli. Across groups, N170 amplitudes were larger in the right hemisphere for face decisions. For the face vs. no face contrast in inverted Mooney stimuli, P1 amplitudes were larger in the right compared to the left cluster.
Time frequency analyses face vs. no face decisions in inverted Mooney stimuli
We hypothesized larger GBA for controls compared to the ASD group. Contrary to expectations, groups did not differ in their general GBA (F(1, 36) = 0.407, p = .528, ηp2 = .011). Whether they detected a face or not did not influence GBA levels (F(1, 36) = 0.049, p = .826, ηp2 = .001), nor was there a percept × group interaction (F(1, 36) = 0.056, p = .814, ηp2 = .002). GBA levels significantly differed across time (F(1, 36) = 7.888, p < .01, ηp2 = .158) with larger activity in the early (50–200 ms) compared to the later time window (200–350 ms). The significant time × group interaction (F(1, 36) = 5.392, p < .05, ηp2 = .110) indicates that controls showed larger GBA levels within the first time window (p < .001), whereas GBA levels for the ASD group were equal across time (p = .742). None of the effects of hemisphere reached significance (hemisphere: F(1, 36) = 0.393, p = .535, ηp2 = .011; hemisphere × group: F(1, 36) = 0.003, p = .986, ηp2 = .000; hemisphere × percept: F(1, 36) = 0.081, p = .778, ηp2 = .002; hemisphere × time: F(1, 36) = 0.079, p = .781, ηp2 = .002). GBA levels were larger for anterior compared to the posterior cluster (F(1, 36) = 8.799, p < .01, ηp2 = .189). There was no significant cluster × group interaction (F(1, 36) = 1.690, p = .202, ηp2 = .036), nor cluster × percept interaction (F(1, 36) = 0.084, p = .774, ηp2 = .002). GBA levels of clusters did, however, differ between time windows (F(1, 36) = 10.389, p < .001, ηp2 = .223). A larger reduction of activity from the early to the later time window in posterior (p < .001), but not within the anterior cluster (p = .368), was detected. Cluster did not interact with hemisphere (F(1, 36) = 0.005, p = .983, ηp2 = .000). No significant 3- or 4-way interactions were observed.
Summary time frequency analysis
Groups did not differ in their general GBA to Mooney stimuli, regardless of orientation or percept. Controls displayed significant decreases in GBA levels in frontal clusters across time. This decrease in GBA did not occur in participants with ASD. For both groups, GBA was larger for the anterior cluster and activity showed larger decreases at the posterior compared to the anterior cluster across time (Fig. 4).
Fig. 4.

Group differences in gamma-band activity for controls and adolescents with ASD. Time-frequency plots for face and no face decisions of the ASD group and controls at the anterior left cluster (a), anterior right cluster (b), posterior left cluster (c), and posterior right cluster (d). The colored scales indicate Z-transformed power values
Discussion
The present study yielded insight into holistic and configural face processing in ASD and neurotypical adolescents by examining behavioral performance (detection rate, response time) and neural correlates (P1, N170, gamma-band activity) with a Mooney stimuli inversion task. The behavioral responses suggest that mechanisms for holistic face processing are in place for both groups. Higher face detection rates for Mooney stimuli in upright orientation compared to inverted were found for both controls and the ASD group. The finding is consistent with previous studies, which also found an intact face inversion effect for individuals with ASD [15, 20–22]. While we cannot eliminate the potential that a part-based processing style influenced face detection rates in the ASD group for prior reports utilizing upright and inverted stimuli [18], our high rates of face identification are unlikely to be due to any type of parts-based system given the nature of the Mooney stimuli. Despite a similar impact of orientation on behavioral decisions of “face-ness” for both groups, adolescents with ASD were less likely to perceive Mooney stimuli as faces than controls and displayed longer reaction times to make a face decision, which is in line with another study that employed Mooney stimuli [19]. Besides intact holistic face processing, the finding also supports a quantitative instead of a qualitative face perception difference in ASD [22, 24].
The ERP results also implicate group differences in the early neural circuitry within the visual processing system thought to reflect attentional (e.g., P1), rather than perceptual (e.g., N170) processes specific to faces. Within the comparison of face responses contrasting upright and inverted Mooney stimuli, controls displayed faster P1 latencies compared to the ASD group, whereas no differences for N170 amplitude or latency could be detected between groups and by orientation. This finding is in contrast to delayed N170 latencies to realistic faces for individuals with ASD [42]. Within the contrast of face and no face responses for inverted stimuli, controls also showed faster P1 latencies compared to the ASD group. Similarly, to the first comparison, no group differences were detected for N170 amplitude or latencies, which is also in contrast to previous findings [23, 42]. Consistent with other studies utilizing facial stimuli, we did find larger N170 amplitudes in the right compared to the left cluster [4].
We investigated the lower gamma-band (25–45 Hz) to examine feature binding processes proposed to underlie deriving a face percept from the black and white Mooney images. No group differences were found in gamma-band activity for groups across clusters, which contrasts previous findings of more GBA in occipital areas for controls compared to adolescents with ASD [45] or more GBA in ASD compared to controls [19]. Both groups showed similar activity in the early window, overlapping the P1 and the start of the N170 component. Former studies found larger GBA for controls in comparison to the ASD group [19]. However, prolonged gamma-band activity for the ASD group was detected in comparison to the control group. Increased temporal activity was also found in similar time windows for adolescents with ASD [45].
General early stage processing of Mooney stimuli was identified by the P1 component activity within both comparisons. The P1 is typically associated with early visual attention [35, 36] and source-localized to the visual association cortex [59]. In our experiment, the task emphasized attention toward the stimulus to determine “face-ness,” while maintaining a 2-button response mapping. Thus, the task protocol required sustained attention and elicited a large P1 component in both groups. It is also possible that our directions provided a strategy that helped to “normalize” engagement of the face processing circuitry, as suggested by consistent morphology of the component across groups and individuals, with quantitative modulation of latency. Thus, the basic attention and processing mechanisms seem to be available in individuals with ASD and can be manipulated to produce greater responses by directing attention [12, 37]. This is in line with another study that found similar face detection rates after directing the attention to parts of the face [10].
Early stage perceptual face processing has been historically assessed by examining response patterns of the N170 component and later GBA (e.g., 150 to 250 ms or 200 to 300 ms) [4, 39]. In contrast to former studies, we did not find any differences in N170 latency or amplitude between adolescents with and without ASD [42]. Although the N170 was right-lateralized in both groups as previously reported [17, 39], it is worthy to note that most studies employ natural faces and previous research suggests that intact natural faces and eyes result in greater and faster N170 responses [60], and schematic face stimuli might induce weaker neural responses [19]. The composition of Mooney stimuli mainly comprises black and white parts that create a 3D shape of a face [26]. As these stimuli did not contain typical first-order face features (e.g., eyes or nose), they may also trigger a weaker or less consistent N170 response [61]. Latinus and Taylor (2005) found that Mooney stimuli elicited a delayed and enhanced N170 component, but only after participants received a training. We also did not find a face inversion for the Mooney images, but this may reflect that our contrast included only those stimuli that were identified as face and the N170 is associated with a general face detection mechanism [4].
Gamma-band activity has been associated with perceptual coherence. In line with the presented behavioral and ERP results, GBA responses were similar for adolescents with ASD and controls over the first 50 to 200 ms. Our groups did differ in later gamma-band activity from 200 to 350 ms, which has been associated with perceptual binding [39].
Besides the association with higher cognitive functions such as memory and attention [43, 45], early GBA has also been associated with the match of bottom-up and top-down information [43]. Within the MUM model, early GBA reflects the matching of bottom-up signals with memory contents and is enhanced when the matching process yields a positive result [62]. Later bursts have been associated with readout processes like action planning, behavioral control, or memory storage [63]. The prolonged GBA for participants with ASD across time might indicate continued activation of the matching processes.
Another explanation of the prolonged activity in ASD could be an imbalance in precision of top-down predictions and bottom-up sensory processing as suggested by the predictive coding framework [64]. Based on Bayesian decision theory, the framework suggests that we perceive our environment by consistently creating inferences. One part of the inference process is prior knowledge which is extracted from earlier sensory events [65]. These priors are consistently updated when presented with sensory evidence (e.g., Mooney stimulus) and these updates are indexed by prediction errors [66]. Individuals with ASD might have hypo-priors, meaning that whenever they saw a Mooney stimulus their system made larger prediction errors [64]. Cortical responses are considered as an index of prediction errors [65]. The prolonged GBA for individuals with ASD across face and no face responses compared to controls might indicate stronger priors in the controls and prolonged updates within the neural network due to larger prediction errors in the ASD group [66]. The lack of differences for GBA might suggest similar bottom-up perceptual binding, assuming that posterior gamma may be more reflective of a posterior-ventral network (e.g., including inferior-occipital gyri; [67]). Additionally, it might indicate an over-reliance on top-down knowledge and less deviation in perceptual areas [8]. The displayed activation patterns of anterior and posterior clusters across time contribute to the idea of different network activations. The significant decrease of GBA levels from the first to the second time window may suggest that perceptual processes are predominant during early perception, whereas networks in anterior areas are constantly active as part of monitoring and decision-making.
Limitations
Due to too few trials, we were unable to analyze the ERP contrasts involving the no face responses for the upright stimuli. Therefore, an enhanced understanding of holistic processing in ASD could be accomplished by a different attentional task. For example, Castelhano et al. (2013) used different perceptual states and presentation angles for the same physical stimulus or Sun et al. (2012) scrambled the Mooney stimuli to make them even less “face-like” [19, 29]. To further delineate effects of face processing from object processing, another option would be to employ Mooney stimuli that are objects or noise as contrasts.
We did not find a face inversion effect in our ERP data. While Mooney stimuli do resemble faces, they are only face-like. The inversion effect for realistic faces not only reflects both a decrement in performance when inverted, but also the efficiency and reliability of processing when upright. Thus, it may be that processing a Mooney stimulus upright compared to inverted (at this age) may require a more similar activation pattern that results in non-significant differences in scalp ERP amplitude and latency.
Adolescent development reflected a research area of less focused attention, given the inclusion of adolescents either in child samples or in adult samples, and a period wherein some qualitative aspects of the face processing system are mature, although potentially quantitatively different. Our results suggest an intact face inversion effect for adolescents with ASD and minor quantitative differences on the neural level.
Longitudinal study designs might be most suitable to detect behavioral changes as well as the time course of P1, N170, and gamma-band abnormalities as Webb, Neuhaus, and Faja [68] have suggested significant improvement and “normalization” of face neural circuitry into adolescence and adulthood in ASD, particularly in relation to first-order processing.
The analysis of binding processes could also be further addressed with phase information and cross-frequency coupling [57, 69, 70]. Understanding connectivity in long-range connections and between sensory areas and attentional systems would be important in understanding how top-down processes related to the task directions influence perceptual responses.
Conclusions
In this paper, we examined behavioral performance, P1, N170, and gamma-band activity in adolescents with ASD and controls during face perception with a carefully selected (IQ-, sex-, and age-matched), narrow range sample. Processing differences may be due to less efficient holistic face processing in ASD, which is required to perceive Mooney stimuli as faces. However, the general similarities between groups suggest that these neural systems are available in individuals with ASD but may be less pronounced or consistently activated. Thus, the fundamental idea of individuals with ASD having an impaired holistic face processing system should be reviewed.
Additional file
Geodesic Sensor Net–128 Channel V 2.0. (PDF 746 kb)
Acknowledgements
The authors thank the participants and their families for their time and effort in participation in this project. Additional support was provided by Adham Atyabi, Geraldine Dawson, Anna Kresse, Tisa Nalty, and Jessica Shook.
Funding
Funding was provided by the Cure Autism Now (Webb) and NIH (R01 MH10028, Pelphrey).
Availability of data and materials
The data that supports the findings of these studies are available from the corresponding author upon reasonable request. The code used in these studies is available from the corresponding author upon reasonable request.
Abbreviations
- ADI
Autism Diagnostic Interview
- ADOS
Autism Diagnostic Observation Schedule
- ASD
Autism spectrum disorder
- EEG
Electroencephalography
- ERP
Event-related potential
- ICA
Independent component analysis
- ITI
Inter-trial interval
- WISC
Wechsler Intelligence Scale
Authors’ contributions
SN directed the data analysis and interpretation of the data and drafted the manuscript. US contributed to the data analysis and interpretation of the data and revised the manuscript. MS contributed to the data analysis and interpretation of the data and revised the manuscript. JM contributed to the coordination and collection of the data and revised the manuscript. SJW conceived of the study; directed in its design, coordination, analysis, and interpretation; and revised the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The study was approved by the University of Washington’s Institutional Review Board Human Subject’s Division. Informed written consent was provided by a custodial parent; the participants provided written assent for participation.
Consent for publication
All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Sandra Naumann, Email: sandra.naumann@hu-berlin.de.
Ulrike Senftleben, Email: Ulrike.Senftleben@tu-dresden.de.
Megha Santhosh, Email: Megha.Santhosh@seattlechildrens.org.
James McPartland, Email: James.McPartland@yale.edu.
Sara Jane Webb, Email: sjwebb@uw.edu.
References
- 1.Jemel B, Mottron L, Dawson M. Impaired face processing in autism: fact or artifact? J Autism Dev Disord. 2006;36(1):91–106. doi: 10.1007/s10803-005-0050-5. [DOI] [PubMed] [Google Scholar]
- 2.Webb SJ, Dawson G, Bernier R, Panagiotides H. ERP evidence of atypical face processing in young children with autism. J Autism Dev Disord. 2006;36(7):881–890. doi: 10.1007/s10803-006-0126-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Taubert J, Apthorp D, Aagten-Murphy D, Alais D. The role of holistic processing in face perception: evidence from the face inversion effect. Vis Res. 2011;51(11):1273–1278. doi: 10.1016/j.visres.2011.04.002. [DOI] [PubMed] [Google Scholar]
- 4.Zion-Golumbic E, Bentin S. Dissociated neural mechanisms for face detection and configural encoding: evidence from N170 and induced gamma-band oscillation effects. Cereb Cortex. 2007;17(8):1741–9. [DOI] [PubMed]
- 5.Freire A, Lee K, Symons LA. The face-inversion effect as a deficit in the encoding of Configural information: direct evidence. Perception. 2000;29(2):159–170. doi: 10.1068/p3012. [DOI] [PubMed] [Google Scholar]
- 6.Kumar SL. Examining the characteristics of Visuospatial information processing in individuals with high-functioning autism. Yale J Biol Med. 2013;86(2):147–156. [PMC free article] [PubMed] [Google Scholar]
- 7.Piepers DW, Robbins RA. A review and clarification of the terms ‘holistic,’ ‘configural,’ and ‘relational’ in the face perception literature. Front Psychol. 2012;3:559. doi: 10.3389/fpsyg.2012.00559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Steinberg Lowe M, Lewis GA, Poeppel D. Effects of part- and whole-object primes on early MEG responses to Mooney faces and houses. Front Psychol. 2016;7(February):147. doi: 10.3389/fpsyg.2016.00147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.American Psychiatric Association . Diagnostic and Statistical Manual of Mental Disorders (DSM-V), no. 1. 2013. [Google Scholar]
- 10.Joseph RM, Tanaka J. Holistic and part-based face recognition in children with autism. J Child Psychol Psychiatry Allied Discip. 2003;44(4):529–542. doi: 10.1111/1469-7610.00142. [DOI] [PubMed] [Google Scholar]
- 11.D’Souza D, Booth R, Connolly M, Happé F, Karmiloff-Smith A. Rethinking the concepts of ‘local or global processors’: evidence from Williams syndrome, Down syndrome, and autism spectrum disorders. Dev Sci. 2016;19(3):452–468. doi: 10.1111/desc.12312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Webb SJ, Neuhaus E, Faja S. Face perception and learning in autism Spectrum disorders. Q J Exp Psychol. 2016;0218(May):1–44. doi: 10.1080/17470218.2016.1151059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Van der Hallen R, Evers K, Brewaeys K, Van den Noortgate W, Wagemans J. Global processing takes time: a meta-analysis on local–global visual processing in ASD. Psychol Bull. 2015;141(3):549–573. doi: 10.1037/bul0000004. [DOI] [PubMed] [Google Scholar]
- 14.Frith U. diagnosis and treatment of autism. New York: Plenum Press; 1989. Autism and ‘theory of mind; pp. 33–52. [Google Scholar]
- 15.Lahaie A, Mottron L, Arguin M, Berthiaume C, Jemel B, Saumier D. Face perception in high-functioning autistic adults: evidence for superior processing of face parts, not for a configural face-processing deficit. Neuropsychology. 2006;20(1):30–41. doi: 10.1037/0894-4105.20.1.30. [DOI] [PubMed] [Google Scholar]
- 16.Latinus M, Taylor MJ. Holistic processing of faces: learning effects with Mooney faces. J Cogn Neurosci. 2005;17(8):1316–1327. doi: 10.1162/0898929055002490. [DOI] [PubMed] [Google Scholar]
- 17.O’Connor K, Hamm JP, Kirk IJ. The neurophysiological correlates of face processing in adults and children with Asperger’s syndrome. Brain Cogn. 2005;59(1):82–95. doi: 10.1016/j.bandc.2005.05.004. [DOI] [PubMed] [Google Scholar]
- 18.Rose FE, Lincoln AJ, Lai Z, Ene M, Searcy YM, Bellugi U. Orientation and affective expression effects on face recognition in Williams syndrome and autism. J Autism Dev Disord. 2007;37(3):513–522. doi: 10.1007/s10803-006-0200-4. [DOI] [PubMed] [Google Scholar]
- 19.Sun L, et al. Impaired gamma-band activity during perceptual Organization in Adults with autism Spectrum disorders: evidence for dysfunctional network activity in frontal-posterior cortices. J Neurosci. 2012;32(28):9563–9573. doi: 10.1523/JNEUROSCI.1073-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Rutherford MD, Clements KA, Sekuler AB. Differences in discrimination of eye and mouth displacement in autism spectrum disorders. Vis Res. 2007;47(15):2099–2110. doi: 10.1016/j.visres.2007.01.029. [DOI] [PubMed] [Google Scholar]
- 21.Scherf KS, Behrmann M, Minshew N, Luna B. Atypical development of face and greeble recognition in autism. J Child Psychol Psychiatry Allied Discip. 2008;49(8):838–847. doi: 10.1111/j.1469-7610.2008.01903.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Weigelt S, Koldewyn K, Kanwisher N. Face identity recognition in autism spectrum disorders: a review of behavioral studies. Neurosci Biobehav Rev. 2012;36(3):1060–1084. doi: 10.1016/j.neubiorev.2011.12.008. [DOI] [PubMed] [Google Scholar]
- 23.McPartland J, Dawson G, Webb SJ, Panagiotides H, Carver LJ. Event-related brain potentials reveal anomalies in temporal processing of faces in autism spectrum disorder. J Child Psychol Psychiatry Allied Discip. 2004;45(7):1235–1245. doi: 10.1111/j.1469-7610.2004.00318.x. [DOI] [PubMed] [Google Scholar]
- 24.Akechi H, Kikuchi Y, Tojo Y, Osanai H, Hasegawa T. Neural and behavioural responses to face-likeness of objects in adolescents with autism spectrum disorder. Sci Rep. 2014;4:1–7. doi: 10.1038/srep03874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.George N, Jemel B, Fiori N, Chaby L, Renault B. Electrophysiological correlates of facial decision: insights from upright and upside-down Mooney-face perception. Cogn Brain Res. 2005;24(3):663–673. doi: 10.1016/j.cogbrainres.2005.03.017. [DOI] [PubMed] [Google Scholar]
- 26.Grützner C, Uhlhaas PJ, Genc E, Kohler A, Singer W, Wibral M. Neuroelectromagnetic correlates of perceptual closure processes. J Neurosci. 2010;30(24):8342–8352. doi: 10.1523/JNEUROSCI.5434-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Otsuka Y, Hill HCH, Kanazawa S, Yamaguchi MK, Spehar B. Perception of Mooney faces by young infants: the role of local feature visibility, contrast polarity, and motion. J Exp Child Psychol. 2012;111(2):164–179. doi: 10.1016/j.jecp.2010.10.014. [DOI] [PubMed] [Google Scholar]
- 28.Verhallen RJ, Bosten JM, Goodbourn PT, Bargary G, Lawrance-Owen AJ, Mollon JD. An online version of the Mooney face test: phenotypic and genetic associations. Neuropsychologia. 2014;63(1):19–25. doi: 10.1016/j.neuropsychologia.2014.08.011. [DOI] [PubMed] [Google Scholar]
- 29.Castelhano J, Rebola J, Leitão B, Rodriguez E, Castelo-Branco M. To perceive or not perceive: the role of gamma-band activity in signaling object percepts. PLoS One. 2013;8(6):35–37. doi: 10.1371/journal.pone.0066363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Gauthier I, Klaiman C, Schultz RT. Face composite effects reveal abnormal face processing in autism Spectrum disorders. Heal (San Fr.) 2010;49(4):470–478. doi: 10.1016/j.visres.2008.12.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jeffery L, Taylor L, Rhodes G. Transfer of figural face aftereffects suggests mature orientation selectivity in 8-year-olds’ face coding. J Exp Child Psychol. 2014;126:229–244. doi: 10.1016/j.jecp.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 32.Taylor MJ, Batty M, Itier RJ. The faces of development: a review of early face processing over childhood. J Cogn Neurosci. 2004;16(8):1426–1442. doi: 10.1162/0898929042304732. [DOI] [PubMed] [Google Scholar]
- 33.Dawson G, Webb SJ, McPartland J. Understanding the nature of face processing impairment in autism: insights from behavioral and electrophysiological studies. Dev Neuropsychol. 2005;27(3):403–424. doi: 10.1207/s15326942dn2703_6. [DOI] [PubMed] [Google Scholar]
- 34.Jeste SS, Nelson CA. Event related potentials in the understanding of autism spectrum disorders: an analytical review. J Autism Dev Disord. 2009;39(3):495–510. doi: 10.1007/s10803-008-0652-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Crist RE, Wu C-T, Karp C, Woldorff MG. Face processing is gated by visual spatial attention. Front Hum Neurosci. 2008;1:10. doi: 10.3389/neuro.09.010.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Jacques C, Rossion B. Electrophysiological evidence for temporal dissociation between spatial attention and sensory competition during human face processing. Cereb Cortex. 2007;17(5):1055–1065. doi: 10.1093/cercor/bhl015. [DOI] [PubMed] [Google Scholar]
- 37.Neuhaus E, Kresse A, Faja S, Bernier RA, Webb SJ. Face processing among twins with and without autism: social correlates and twin concordance. Soc Cogn Affect Neurosci. 2016;11(1):44–54. doi: 10.1093/scan/nsv085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Webb SJ, Merkle K, Murias M, Richards T, Aylward E, Dawson G. ERP responses differentiate inverted but not upright face processing in adults with ASD. Soc Cogn Affect Neurosci. 2012;7(5):578–587. doi: 10.1093/scan/nsp002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Anaki D, Zion-Golumbic E, Bentin S. Electrophysiological neural mechanisms for detection, configural analysis and recognition of faces. Neuroimage. 2007;37(4):1407–1416. doi: 10.1016/j.neuroimage.2007.05.054. [DOI] [PubMed] [Google Scholar]
- 40.Itier RJ, Alain C, Sedore K, McIntosh AR. Early face processing specificity: It’s in the eyes! J Cogn Neurosci. 2007;19(11):1815–1826. doi: 10.1162/jocn.2007.19.11.1815. [DOI] [PubMed] [Google Scholar]
- 41.Webb SJ, Bernier R, Paul M, Dawson G. Further evidence for an abnormality in the temporal processing of faces in autism. 2003. [Google Scholar]
- 42.Hileman CM, Henderson H, Mundy P, Newell L, Jaime M. Developmental and individual differences on the P1 and N170 ERP components in children with and without autism. Dev Neuropsychol. 2011;36(2):214–236. doi: 10.1080/87565641.2010.549870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Güntekin B, Başar E. A review of brain oscillations in perception of faces and emotional pictures. Neuropsychologia. 2014;58(1):33–51. doi: 10.1016/j.neuropsychologia.2014.03.014. [DOI] [PubMed] [Google Scholar]
- 44.Kaiser J, Lutzenberger W. Induced gamma-band activity and human brain function. Neuroscientist. 2003;9(6):475–484. doi: 10.1177/1073858403259137. [DOI] [PubMed] [Google Scholar]
- 45.Wright B, et al. Gamma activation in young people with autism spectrum disorders and typically-developing controls when viewing emotions on faces. PLoS One. 2012;7:7. doi: 10.1371/journal.pone.0041326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Herrmann CS, Fründ I, Lenz D. Human gamma-band activity: a review on cognitive and behavioral correlates and network models. Neurosci Biobehav Rev. 2010;34(7):981–992. doi: 10.1016/j.neubiorev.2009.09.001. [DOI] [PubMed] [Google Scholar]
- 47.Rodriguez E, George N, Lachaux JP, Martinerie J, Renault B, Varela FJ. Perception’s shadow: long-distance synchronization of human brain activity. Nature. 1999;397(6718):430–433. doi: 10.1038/17120. [DOI] [PubMed] [Google Scholar]
- 48.Grice SJ, et al. Disordered visual processing and oscillatory brain activity in autism and Williams syndrome. Neuroreport. 2001;12(12):2697–2700. doi: 10.1097/00001756-200108280-00021. [DOI] [PubMed] [Google Scholar]
- 49.Lord C, Rutter M, Le Couteur A. Autism diagnostic interview-revised: a revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J Autism Dev Disord. 1994;24(5):659–685. doi: 10.1007/BF02172145. [DOI] [PubMed] [Google Scholar]
- 50.Lord C, et al. The autism diagnostic observation schedule-generic: a standard measure of social and communication deficits associated with the spectrum of autism. J Autism Dev Disord. 2000;30(3):205–223. doi: 10.1023/A:1005592401947. [DOI] [PubMed] [Google Scholar]
- 51.APA . Diagnostic and statistical manual of mental disorders, 4th text. Washington, D.C.: Author; 1994. [Google Scholar]
- 52.Mooney CM. Age in the development of closure ability in children. Can J Psychol. 1957;11(4):219–226. doi: 10.1037/h0083717. [DOI] [PubMed] [Google Scholar]
- 53.Chaumon M, Bishop DVM, Busch NA. A practical guide to the selection of independent components of the electroencephalogram for artifact correction. J Neurosci Methods. 2015;250:47–63. doi: 10.1016/j.jneumeth.2015.02.025. [DOI] [PubMed] [Google Scholar]
- 54.Luck SJ. An Introduction to the Event-Related Potential Technique, Second Edition. Cambridge, MA: MIT Press; 2014.
- 55.D’Hondt F, et al. Electrophysiological correlates of emotional face processing after mild traumatic brain injury in preschool children. Cogn Affect Behav Neurosci. 2017;17(1):124–142. doi: 10.3758/s13415-016-0467-7. [DOI] [PubMed] [Google Scholar]
- 56.Tallon-Baudry C, Bertrand O, Delpuech C, Permier J. Oscillatory gamma-band (30-70 Hz) activity induced by a visual search task in humans. J Neurosci. 1997;17(2):722–734. doi: 10.1523/JNEUROSCI.17-02-00722.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Castelhano J, Bernardino I, Rebola J, Rodriguez E, Castelo-Branco M. Oscillations or synchrony? Disruption of neural synchrony despite enhanced gamma oscillations in a model of disrupted perceptual coherence. J Cogn Neurosci. 2015;27(12):2416–2426. doi: 10.1162/jocn_a_00863. [DOI] [PubMed] [Google Scholar]
- 58.Wright B, et al. Gamma activation in young people with autism spectrum disorders and typically-developing controls when viewing emotions on faces. PLoS One. 2012;7(7):e41326. doi: 10.1371/journal.pone.0041326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wong TKW, Fung PCW, McAlonan GM, Chua SE. Spatiotemporal dipole source localization of face processing ERPs in adolescents: a preliminary study. Behav Brain Funct. 2009;5(1):16. doi: 10.1186/1744-9081-5-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Cecchini M, Aceto P, Altavilla D, Palumbo L, Lai C. The role of the eyes in processing an intact face and its scrambled image: a dense array ERP and low-resolution electromagnetic tomography (sLORETA) study. Soc Neurosci. 2013;8(4):314–325. doi: 10.1080/17470919.2013.797020. [DOI] [PubMed] [Google Scholar]
- 61.Sagiv N, Bentin S. Structural encoding of human and schematic faces: holistic and part-based processes. J Cogn Neurosci. 2001;13(7):937–951. doi: 10.1162/089892901753165854. [DOI] [PubMed] [Google Scholar]
- 62.Herrmann CS, Munk MHJ, Engel AK. Cognitive functions of gamma-band activity: memory match and utilization. Trends Cogn Sci. 2004;8(8):347–355. doi: 10.1016/j.tics.2004.06.006. [DOI] [PubMed] [Google Scholar]
- 63.Rojas DC, Wilson LB. Gamma-band abnormalities as markers of autism spectrum disorders. Biomark Med. 2014;8(3):353–368. doi: 10.2217/bmm.14.15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Pellicano E, Burr D. When the world becomes ‘too real’: a Bayesian explanation of autistic perception. Trends Cogn Sci. 2012;16(10):504–510. doi: 10.1016/j.tics.2012.08.009. [DOI] [PubMed] [Google Scholar]
- 65.Stefanics G, Kremlacek J, Czigler I. Visual mismatch negativity: a predictive coding view. Front Hum Neurosci. 2014;8(September):1–19. doi: 10.3389/fnhum.2014.00666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Friston KJ, Lawson R, Frith CD. On hyperpriors and hypopriors: comment on Pellicano and Burr. Trends Cogn Sci. 2013;17(1):1. doi: 10.1016/j.tics.2012.11.003. [DOI] [PubMed] [Google Scholar]
- 67.Gao Z, Goldstein A, Harpaz Y, Hansel M, Zion-Golumbic E, Bentin S. A magnetoencephalographic study of face processing: M170, gamma-band oscillations and source localization. Hum Brain Mapp. 2013;34(8):1783–1795. doi: 10.1002/hbm.22028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Webb SJ, Neuhaus E, Faja S. Face perception and learning in autism spectrum disorders. Q J Exp Psychol. 2017;70(5):970–986. doi: 10.1080/17470218.2016.1151059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Kessler K, Seymour RA, Rippon G. Brain oscillations and connectivity in autism spectrum disorders (ASD): new approaches to methodology, measurement and modelling. Neurosci Biobehav Rev. 2016;71:601–620. doi: 10.1016/j.neubiorev.2016.10.002. [DOI] [PubMed] [Google Scholar]
- 70.David N, Schneider TR, Peiker I, Al-Jawahiri R, Engel AK, Milne E. Variability of cortical oscillation patterns: a possible endophenotype in autism spectrum disorders? Neurosci Biobehav Rev. 2016;71:590–600. doi: 10.1016/j.neubiorev.2016.09.031. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Geodesic Sensor Net–128 Channel V 2.0. (PDF 746 kb)
Data Availability Statement
The data that supports the findings of these studies are available from the corresponding author upon reasonable request. The code used in these studies is available from the corresponding author upon reasonable request.