

Abstract
Chemotherapy resistance of breast cancer poses a great challenge to the survival of patients. During breast cancer treatment, the development of intrinsic and acquired drug resistance tends to further induce adverse prognosis, such as metastasis. In recent years, the progress of research on cytokine-modulated tumor microenvironment and breast cancer stem cells (BCSCs) has shed light on defining the mechanisms of drug resistance gradually. In this review, we have discussed cytokine regulation on breast cancer chemoresistance. Cytokines can affect tumor cell behavior or reprogram tumor niche through specific signaling pathways, thereby regulating the progress of drug resistance. In addition, we summarized the mutually regulatory networks between cytokines and BCSCs in mediating chemoresistance. Cytokines in the tumor microenvironment can regulate the self-renewal and survival of BCSCs in a variety of ways, sequentially promoting chemotherapeutic resistance. Therefore, the combinational treatment of BCSC targeting and cytokine blockade may have a positive effect on the clinical treatment of breast cancer.
Keywords: Breast cancer stem cells (BCSCs), Cytokine, Chemoresistance, Tumor microenvironment
Introduction
Breast cancer has been seriously endangering the public health because of its high incidence in women [1, 2]. According to the expression of molecular markers (estrogen receptor, progesterone receptor, and HER2), breast cancer can be divided into several subtypes: luminal A, luminal B, HER2+, and triple-negative [3]. According to the different subtypes, there are different therapeutic strategies in clinic. Commonly used treatments for breast cancer include surgery, radiotherapy, chemotherapy, endocrine therapy, targeted therapy and so on. With the development of new drugs, chemotherapy is widely used in the treatment of breast cancer. However, some subtypes of breast cancer are prone to be drug-resistant to chemotherapy, resulting in that the treatment efficacy is very limited, which brings great challenge to clinicians in improving survival of breast cancer patients.
Cancer stem cells are a small population of cells in solid tumors or leukaemia [4, 5]. They are characterized as stem cell-like phenotype, capable of self-renewal and differentiation [6, 7]. Many studies have shown that the role of cancer stem cells can not be ignored in many processes such as tumorigenesis, tumor growth, metastasis and tumor progression [4, 8–13]. In the course of chemotherapy for breast cancer patients, common chemotherapeutic drugs target non-cancer stem cells, but the cancer stem cells can survive and further cause recurrence or even metastasis due to their own characteristics. Therefore, breast cancer stem cells are considered to be the key population leading to drug resistance of breast cancer [14, 15]. With the development of molecular markers of breast cancer stem cells, researchers have found that a small population of cells, selected by markers, can lead to the initiation of tumors, which further confirms the vital role of cancer stem cells in tumor initiation [16, 17].
Cytokine plays important roles in the development of multiple cancers in addition to regulating innate immunity and adaptive immunity, blood cell generation, cell growth and repair of damaged tissue. On one hand, some cytokines can directly regulate the behavior of tumor cells in an either autocrine or paracrine way, and regulate tumor progression, including chemotherapeutic resistance [18, 19]. On the other hand, cytokines can function through affecting other types of cells, such as endothelial cells, fibroblasts and immune cells in the tumor niche. It can further reprogram the tumor microenvironment indirectly, resulting in tumor promotion or tumor suppression and affecting sensitivity to chemotherapeutic agents [20–22]. In addition, studies have shown that some cytokines can also regulate cancer stem cells and further affect the drug resistance of tumor cells [23, 24]. Cancer stem cells can also secrete some specific cytokines and promote their own resistance to chemotherapy drugs to survive [22, 23, 25].
In this review, we summarized recent studies on the roles of cytokines and BCSCs in mediating breast cancer chemoresistance, and discussed the potential of cytokines as therapeutic target to provide new strategies for clinicians to improve the treatment to breast cancer patients.
Cytokines and chemoresistance
Direct regulation of cytokines on tumor cells to promote chemoresistance
With the progression of cancer, tumor cells may also express specific cytokine receptors to receive signal stimuli from the corresponding cytokine ligands in the manner of autocrine or paracrine, regulate intracellular signal transduction, and promote the resistance of tumor cells to chemotherapeutic drugs.
IL-6
Interleukin-6 (IL-6) was initially identified as B cell stimulating factor 2, enhancing immunoglobulin synthesis through activating B cells and was a prototypical cytokine with pleiotropic and redundant activities of a wide range in immune regulation, hematopoiesis, inflammation and oncogenesis. It helps the host to defend against infection and tissue damage as an inflammatory and immunomodulatory cytokine. However, the persistent IL-6 synthesis leads to the development of various diseases, including cancers [26].
The IL-6 signaling is aberrantly hyper activated in many types of cancer and is generally associated with a poor clinical prognosis [27]. IL-6 was the most highly expressed cytokine in the human colorectal cancer-derived mesenchymal stem cells conditioned medium, and promoted the progression of colorectal cancer cells through IL-6/JAK2/STAT3 signaling, which activated PI3K/AKT signaling. Besides, anti-IL-6 antibody abolished the migration and invasion of colon cancer cells induced by IL-6-activated pathway [28]. Human liver cancer tissues contained high ratio of Tim-3-expressing hepatocytes and HBV involved in Tim-3 upregulation in malignant hepatocytes. The hepatocyte-Tim-3 receptor activates NF-kappa B phosphorylation, which in turn stimulates IL-6 secretion and STAT3 phosphorylation, resulting in tumor growth both in vitro and in vivo [29]. In head and neck cancer, IL-6 can induce its expression as upstream of OPN, and OPN promotes the growth, migration and invasion of cancer cells through activating integrin αvβ3-NF-kappa B axis [30].
IL-6 also participated in breast cancer chemoresistance. Local IL-6 paracrine loop act as exogenous IL-6 rich niche for chemo-sensitive breast cancer cells, leading to de novo acquired drug resistance [31]. As one of the principal oncogenic molecules, IL-6 treatment could induce upregulation of HIF-1 alpha via the activation of STAT3, which consequently contributed to its effect against chemotherapeutic drug-induced cytotoxicity and cell apoptosis [32]. In the established MDA-MB-231 metastatic breast cancer cell line, knockdown of endogenous VCAM-1 expression reduced cell proliferation and inhibited IL-6 mediated cell migration, and increased chemosensitivity [33]. Treatment with a genotoxic drug combination (5-fluorouracil, doxorubicin, and cyclophosphamide) activated an NF-kappaB-IL6-dependent inflammatory signaling that imparted stemness to nonstem cancer cells, induced multidrug resistance in breast cancer [34]. The role of JAK-STAT3 in mediating the promotion of IL-6 on drug resistance has been confirmed [26]. IL-6 mainly promotes the expression of multiple genes through STAT3, thus regulating breast cancer drug resistance (Fig. 1a).
Fig. 1.

The IL-6 and IL-8 signal transduction pathways. a The binding of IL-6 to its receptor IL-6R leads to homodimerization of gp130, resulting in phosphorylation of JAKs which further phosphorylates and activates STAT3. The activated STAT3 binds with other STAT proteins (STATs) to form homodimers or heterodimers, which facilitates the transcription of diverse downstream genes. b IL-8 binds to its receptor CXCR1 or CXCR2 (belonging to GPCR superfamily), activating G protein. The G protein subunits activate PLC and PI3K, which further lead to the phosphorylation of PKC and AKT, respectively. Besides, the Rho-GTPase family and non-receptor tyrosine kinases (such as Src and FAK) can be activated by IL-8 signaling. And activated MAPK signaling cascade (Raf-1/MEK/ERK) also transduces the IL-8 stimuli
IL-8
Interleukin-8 (IL-8), also known as chemokine CXCL8, is a cytokine secreted by a wide range of cell populations. IL-8 has cellular chemotaxis to neutrophils to regulate the inflammatory response through combining with chemokine receptor interleukin-8 receptor alpha (IL8RA, also called CXCR1) and interleukin-8 receptor beta (IL8RB/CXCR2), and has multiple cell targets in addition to neutrophils. IL-8 involves in various human diseases, such as atherosclerosis, cancer, inflammatory bowel disease, infection, sepsis, chronic obstructive pulmonary disease, psoriasis and rheumatoid arthritis [35].
The chemokine IL-8 is overexpressed in multiple cancer types, including triple-negative breast cancer (TNBC), where it promotes the acquisition of mesenchymal features, stemness, and chemoresistance. The current research explores the utility of a clinical-stage monoclonal antibody that neutralizes IL-8 as a potential therapeutic option for TNBC [36]. IL-8 showed increased expressions in paclitaxel-treated advanced breast cancer and this over-production effect was inhibited in TLR4-silenced cells. The acquired TLR4-mediated paclitaxel resistance in advanced breast cancer is explained partly by the paracrine effect of IL-8 release [37]. Cytokines produced by breast cancer cells after chemotherapy withdrawal activate both Wnt/beta-catenin and NF-kappa-B pathways, which in turn further promote breast cancer cells to produce and secrete cytokines, forming an autocrine inflammatory forward-feedback loop to facilitate the enrichment of drug-resistant breast cancer cells [38]. Moreover, such an autocrine forward-feedback loop can also be diminished by IL-8 neutralizing antibody or blockade of IL-8 receptors CXCR1/2 with reparixin, and, in a human xenograft model, administration of reparixin after chemotherapy withdrawal effectively attenuates tumor masses [38]. Breast cancer patients treated with chemotherapeutic drugs exhibited poor survival rate and shorter disease-free survival time if their tumor samples expressed high level of IL-8, or its receptor, CXCR1, CXCR2 [38].
The downstream signals activated by IL-8-CXCR1/2 are context-specific, and the effector proteins that mediate IL-8 signals are also pleiotropic, and these effectors may also affect each other. The activation of GPCR (CXCR1 and CXCR2) activated by IL-8 promotes the activation of a variety of downstream pathways, including but may not be limited to PLC-PKC, PI3K-AKT, Rho-GTPase family, FAK/Src of non-receptor tyrosine kinases and MAPK cascade signals (Fig. 1b), which ultimately affect the progression of cancer, angiogenesis, metastasis, and cancer stem cell activation [39]. Although Src and NF-κB have been found to be drug-resistant causes in prostate and colon cancer respectively, the specific mechanism in chemoresistance and critical downstream pathways of IL-8 signal in breast cancer are still not well described, and more research is needed to clarify this point.
TGF-β
In addition to the essential roles in germ-layer specification and patterning during embryonic development, transforming growth factor-β (TGF-β) signaling is also involved in human diseases including fibrosis, cardiovascular, cancer, reproductive or wound-healing disorders through control of cellular functions in proliferation, adhesion, invasion, differentiation, apoptosis, and cellular microenvironment [40, 41].
Through analyzing RNA expression in matched pairs of primary breast cancer biopsies before and after chemotherapy, researchers found that biopsies after chemotherapy displayed increased RNA transcripts of TGF-β signaling. And also, in TNBC cell lines and mouse xenografts, the chemotherapeutic drug paclitaxel upregulated autocrine TGF-β signaling [42]. In addition, treatment of TNBC xenografts with LY2157299, the TGF-β type I receptor kinase inhibitor, prevented relapse of tumors after paclitaxel treatment, suggesting that chemotherapy-induced TGF-β signaling enhances tumor recurrence and that TGF-β pathway inhibitors prevent the development of drug-resistant breast cancer cells [42]. Vinorelbine is one of the most active cytotoxic agents in breast cancer, especially metastatic breast cancer and bioinformatics analysis indicated that TGF-β signaling pathway may associate with drug resistance of breast cancer cells to vinorelbine [43].
The strong implication of TGF-β in mammary epithelial-to-mesenchymal transition (EMT) promotion is becoming increasingly accepted. PARP3 was upregulated in the course of TGF-β-induced EMT and PARP3 responded to TGF-β-induced signaling to enhance the TG2-Snail-E-cadherin axis during EMT, which demonstrates the critical role of PARP3 in mediating the promotion of TGF-β to EMT [44]. The phenotype switch is now indicated as an important contributor to the acquisition of drug resistance, a clinically relevant issue involved in the preservation of high mortality rates among breast cancer cases [45]. As the regulators of TGF-β, numerous miRNAs are involved in TGF-β signaling [46]. They can intervene in the progression of drug resistance in breast cancer and function as enhancers or inhibitors for chemotherapeutic agents. On the contrary, TGF-β can also regulate the micro-RNA signaling to affect the drug resistance of breast cancer (Fig. 2). TGF-β can induce miR-21 expression, which targeted the 3’ untranslated region of MSH2 mRNA and downregulated its expression. Furthermore, by downregulating MSH2, TGF-β contributed to resistance to DNA-damaging chemotherapy agents (cisplatin, methyl methanesulfonate, and doxorubicin) in breast cancer cells [47].
Fig. 2.
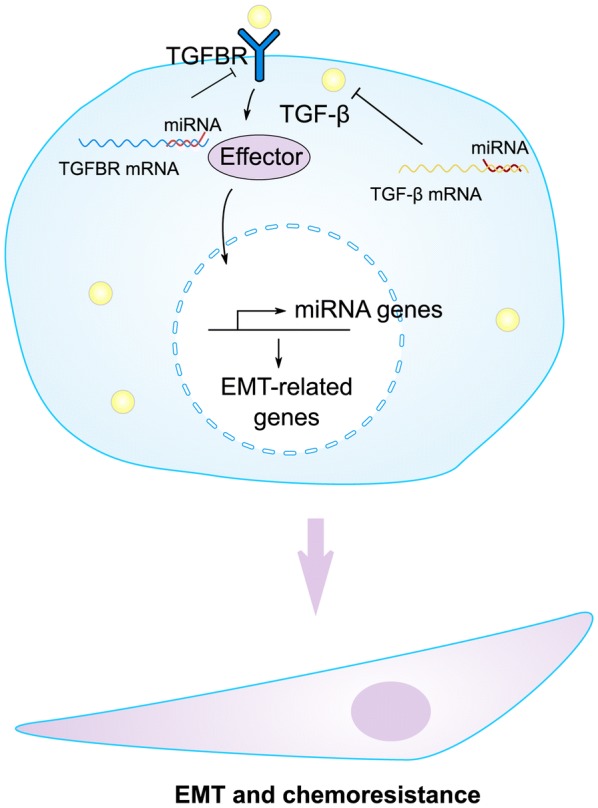
The interplay between microRNAs and TGF-β signaling in regulating chemoresistance of cancer cells. There exists reciprocal modulation between TGF-β and diverse microRNAs. On one hand, TGF-β can affect the transcription of some microRNAs via specific effectors; On the other hand, TGF-β ligand or its receptor TGFBR are the targets of the corresponding microRNAs. These processes control the expression of EMT-related genes, leading to the morphology alteration accompanied with drug resistance in breast cancer
In addition to IL-6, IL-8 and TGF-β, cytokines which can directly determine the effect of chemotherapy for breast cancer, also includes other factors such as M-CSF, TNFα, IL-1β and so on [18]. They lead to breast cancer resistance to various chemotherapeutic agents including paclitaxel, cisplatin, anthracycline and doxorubicin through differential signaling pathways.
Clinical trials in cancer therapies through targeting cytokine signals
These cytokines, closely related to chemotherapeutic resistance, have also attracted the interest of pharmacologists and clinicians, who are trying to develop monoclonal antibodies or small molecular inhibitors targeting these factors to improve the effectiveness of chemotherapies. Therapy through targeting IL-6 signals has been widely applied in inflammatory diseases such as Rheumatoid arthritis [48]. Although no dose-related or cumulative toxicity was apparent in the phase I, open-label study of Siltuximab, an anti-IL-6 monoclonal antibody, in patients of B-cell Non-Hodgkin’s Lymphoma or multiple myeloma (NCT00412321), no responses were seen in patients with relapsed or refractory multiple myeloma treated with single-agent Siltuximab (Table 1). And combining Siltuximab with the bortezomib–melphalan–prednisone (VMP) regimen did not improve complete response (CR), progression free survival, or overall survival but gained very good partial response in multiple myeloma (NCT00911859). In addition, no objective responses were seen in the phase 1/2, multiple-dose, dose-escalation study of siltuximab, suggesting that IL-6 inhibition alone is insufficient in treating advanced or refractory solid tumors including ovarian, pancreatic, colorectal, head & neck cancer and non-small-cell lung carcinoma (NSCLC) (Table 1).
Table 1.
Clinical trials in cancer therapies related to IL-6, IL-8 and TGF-β signals
| Compound and strategy | Cancer type | Phase | Description | Trial numbers | References |
|---|---|---|---|---|---|
| Siltuximab; anti-IL-6 mAb | B-cell Non-Hodgkin’s lymphoma, multiple myeloma | Phase I | No dose-related or cumulative toxicity was apparent across all disease indications | NCT00412321 | [49] |
| Siltuximab; anti-IL-6 mAb | Multiple myeloma | Phase II | Randomized study of bortezomib–melphalan–prednisone with or without siltuximab (anti-IL-6) in multiple myeloma | NCT00911859 | [50] |
| Siltuximab; anti-IL-6 mAb | Multiple myeloma | Phase II | The safety and efficacy of siltuximab with or without dexamethasone for patients with relapsed or refractory multiple myeloma | N/A | [51] |
| Siltuximab; anti-IL-6 mAb | Advanced solid tumors | Phase I/II | Siltuximab monotherapy appears to be well tolerated but without clinical activity in solid tumors | N/A | [52] |
| Reparixin; CXCR1/2 antagonist | HER-2 negative metastatic breast cancer (MBC) | Phase Ib | Weekly paclitaxel plus reparixin in MBC appeared to be safe and tolerable | NCT02001974 | [53] |
| Reparixin; CXCR1/2 antagonist | Early breast cancer | Phase II | Reparixin 1000 mg t.i.d. for 21 consecutive days appeared to be well tolerated | NCT01861054 | [54] |
| Galunisertib; TGFBR1 inhibitor | Advanced cancer and glioma | Phase I | Based on the safety, PK and antitumor activity in glioma patients, the intermittent administration of LY2157299 at 300 mg/day is safe for future clinical investigation | N/A | [55, 56] |
| Galunisertib; TGFBR1 inhibitor | Advanced hepatocellular carcinoma (HCC) | Phase II | HCC patients with normal AFP and with TGFβ1 reduction showed improvement in OS compared to patients with non-TGFβ1 reduction | NCT01246986 | [57] |
| Galunisertib; TGFBR1 inhibitor | Recurrent glioblastoma | Phase II | Galunisertib + lomustine failed to demonstrate improved OS relative to placebo + lomustine | NCT01582269 | [58, 59] |
| Galunisertib; TGFBR1 inhibitor | Advanced solid tumors | Phase I | Galunisertib had an acceptable tolerability and safety profile in Japanese patients with advanced cancers | NCT01722825 | [60] |
| Galunisertib; TGFBR1 inhibitor | Pancreatic cancer | Phase II | GG (galunisertib + gemcitabine) resulted in improvement of OS and PFS in patients with PC, with a manageable toxicity profile as compared to GP (gemcitabine + placebo) | NCT01373164 | [61] |
| PF-03446962; Anti-ALK1 mAb | Urothelial cancer | Phase II | They do not recommend further investigation outside of the combination with agents targeting the VEGF receptor axis | NCT01620970 | [62] |
The data are summarized based on the clinical studies from 2013 to 2018. ALK1 is a member of transforming growth factor-beta (TGF-β) receptor I. Galunisertib, also known as LY2157299 monohydrate
N/A not available
CXCR1 is thought to be a receptor selectively expressed in breast cancer stem cells (BCSCs). Reparixin is an allosteric inhibitor of IL-8 (CXCL8) receptor CXCR1/2 has the activity against BCSCs in xenografts of breast cancer [53]. CXCR1 is thought to be a receptor selectively expressed in breast cancer stem cells. Reparixin is an allosteric inhibitor of IL-8 (CXCL8) receptor CXCR1/2 and has the activity against BCSCs in xenografts of breast cancer. It was confirmed that reparixin monotherapy or paclitaxel plus reparixin were appeared to be safe and tolerable in early or metastatic breast cancer (MBC), respectively (NCT01861054, NCT02001974) (Table 1). However, further studies in the clinical trial to observe the action of reparixin on cancer therapy is still needed.
Small molecule inhibitor, LY215799 monohydrate also known as galunisertib, blocks TGF-beta signaling through inhibiting TGFβ receptor I and reduce tumor progression in preclinical models [55]. Besides, Galunisertib has acceptable tolerability and safety in advanced cancer patients (NCT01722825). In advanced hepatocellular carcinoma (HCC), patients treated with Galunisertib showed improvement in overall survival in a phase 2 study (NCT01246986). However, the mono-antibody of ALK1 [a member of transforming growth factor-beta (TGF-β) receptor I], PF-03446962, had no activities as a single drug in refractory urothelial cancer (NCT01620970). The combination of galunisertib and gemcitabine showed improvement of OS and PFS in patients with unresectable pancreatic cancer (PC) compared to gemcitabine + placebo (NCT01373164). Unfortunately, in patients with recurrent glioblastoma, Galunisertib plus lomustine failed to demonstrate improved OS relative to placebo plus lomustine (NCT01582269) (Table 1). In general, it is still very promising to improve the therapeutic effect of cancers via blockade of TGF-β signaling, which requires more clinical studies to confirm.
Indirect influence of cytokines on tumor chemoresistance via remodeling tumor microenvironment
The tumor microenvironment (TME) comprises immune system elements (such as macrophages and lymphocytes), fibroblast, cells composing blood vessels, myofibroblast, mesenchymal stem cells, adipocytes and extracellular matrix (ECM). Tumor microenvironment (or the tumor niche) plays a vital role in the progression of cancer [63–68], and affects many processes such as tumor growth, metastasis, relapse and drug resistance [69–73].
Cytokines and macrophages
Tumor-associated macrophages (TAM) are the prominent components of TME in breast cancers. Macrophages exhibit a high plasticity in response to various external signals and participate in innate and adoptive immune responses to control numerous factors of TME [74]. Depending on the microenvironmental signal present, macrophages undergo different types of activation, including the “classic” pro-inflammatory phenotype (also called M1) and the “alternative” anti-inflammatory phenotype (also called M2) or even in the transitional state between these two kinds of macrophages. TAMs closely resemble the M2-polarized. Clinicopathological studies have suggested that TAM accumulation in tumors correlates with a poor clinical outcome [74]. However, the characteristics of tumor-infiltrated macrophages are complex. TAMs show pleiotropic effects on tumor behavior due to be stimulated by differential cytokines. Some chemokines may increase the infiltration of TAM and form suitable conditions for tumor outgrowth. Once infiltrated, macrophages may also be regulated by cytokines, changing the gene expression, releasing factors that are beneficial to the progression of tumor and the factors associated with immunosuppression. Finally, multiple behaviors of macrophages affected by cytokines can remodel the tumor microenvironment and promote breast cancer chemotherapy resistance.
Breast cancer-associated macrophages express high levels of insulin-like growth factors 1 and 2 (IGFs) and are the main source of IGFs within both primary and metastatic tumors [73]. In total, 75% of breast cancer patients show activation of insulin/IGF-1 receptor signaling and this correlates with increased macrophage infiltration and advanced tumor stage. In addition, blockade of IGF in combination with paclitaxel showed a significant increase in chemosensitivity of tumor compared to paclitaxel monotherapy [73]. TAMs and its supernatants significantly prevent breast tumor cells from apoptosis caused by paclitaxel and the high level of IL-10 secreted by TAMs was responsible for drug resistance of breast cancer [75]. The possible TAMs-modulated drug resistance mechanism involved may be associated with elevation of bcl-2 gene expression and up-regulation of STAT3 signaling in tumor cells, forming IL-10/STAT3/bcl-2 signaling axis accounting for chemoresistance of breast cancer [75].
Cytokines and fibroblasts
The presence of cancer-associated fibroblasts (CAFs) was found in almost all solid tumors. However, their abundance varies widely among different types of cancer. For example, breast, prostate and pancreatic cancer contain more CAFs, while CAFs in brain, kidney and ovarian tumors is rare. CAFs are markedly different from tumorigenic malignant cells [76]. They have undergone epithelial-to-mesenchymal transition and show fibroblast-like morphology. In addition, there is a great difference between cancer cells and CAF cells that the karyotype is relatively stable, and there are few genetic alterations in CAFs. They are defined as all the fibroblastic, nonneoplastic, nonvascular, nonepithelial, and noninflammatory cells found in a tumor [77].
CAFs may lead to the resistance of cancer cells to anti-tumor drugs by means of soluble factors. Chemokine- or cytokine-signaling pathway perhaps drive CAF to remodel the extracellular matrix (ECM) in the tumor microenvironment and the CAF-remodeled ECM can provide favorable soil for the change of EMT characteristics of tumor cells, conferring chemotherapeutic resistance [78, 79]. The EMT process usually promotes chemoresistance by inducing cell cycle arrest or altering the expression of transporters enabling chemotherapeutic drugs uptake of tumor cells [80–82]. Stimuli from proinflammatory cytokine IL-6 expressed by tumor cells, is sufficient to induce the expression of Twist1 in normal fibroblasts and transdifferentiate them into CAFs through the activation of STAT3 pathway [83]. In the xenograft tumor model of breast cancer, ectopic expression of IL-6 can significantly increase the infiltration of Twist1-positive CAFs, and Twist1 is necessary and sufficient for the trans-differentiation formation of CAFs. In addition, CXCL12 is the target gene of Twist1 transcriptional regulation as the downstream signaling [83]. These studies elucidate that cytokine (IL-6) can promote the remodeling of tumor microenvironment by promoting CAFs formation, infiltration and promoting downstream cytokines expression.
The collagen type I secreted by CAFs was found to reduce the uptake of chemotherapeutic drugs in the tumor, and therefore plays a vital role in regulating the chemosensitivity of cancer to multiple forms of chemotherapy [84]. Accordingly, targeting CAFs can increase the drug absorption inside the tumor, inhibit growth and metastasis of primary tumor cells, and the growth of breast cancer that are resistant to multiple drugs in mice [84]. Taken together, cytokines affect the formation and infiltration of CAFs, and CAFs can reshape the characteristics of ECM in microenvironment and subsequently influence the drug resistance of breast cancer cells. On the other hand, CAFs can secrete specific cytokines to further reprogram the microenvironment.
Synergic regulation of multiple cytokines on chemoresistance
Due to the complexity of the tumor microenvironment, the diversity of cytokines and the existence of pleiotropic regulatory networks among them, the effects of cytokines on tumor progression are often not independent on each other [85–87]. Similarly, in the regulation of cytokines on chemotherapy resistance in breast cancer, some cytokines will exhibit synergistic or inter-dependent phenomena. It is possible that specific context-dependent cytokines can simultaneously regulate the same or different signaling pathways in breast cancer to promote drug resistance; or a cytokine from breast cancer cells or stromal cells in microenvironment promotes the production of another cytokine from the tumor cells; it may also be the combination of the two cytokines above and the more complex cascade response.
Previous studies have shown that cisplatin treatment can significantly alter the secretory phenotype and behavior of mesenchymal stromal cells (MSCs). After cisplatin treatment, the MSCs not only changed the phosphorylation level of many kinases, but also increased the secretion of IL-6 and IL-8, which increased the chemotherapeutic resistance of breast cancer cells to cisplatin [88]. TGF-β can promote paclitaxel resistance in triple-negative breast cancer which is mediated by SMAD4 [42]. In addition, SMAD4 regulates taxane resistance by promoting the expression of IL-8 [42]. The formation of TGF-β-SMAD4-IL-8 axis in breast cancer cells shows a synergistic regulation of multiple cytokines in chemotherapeutic resistance of breast cancer. In brief, in promoting breast cancer chemotherapy resistance, sometimes cytokines are not “fighting” alone. In response to the various effects (necrosis, apoptosis and senescence) on tumor cells produced by chemotherapeutic stimulation, the tumor cells and other cells in TME secrete pleiotropic cytokines, which act on themselves or other cells in manner of autocrine or paracrine, and jointly promote the survival of tumor cells under the treatment of anti-tumor drugs.
Interplay between cytokines and BCSCs on chemoresistance
BCSCs drive chemoresistance
BCSCs are a special group of cells in breast cancer, which can maintain self-renewal and differentiation. The therapeutic effect of treatments to advanced breast cancer, such as radiotherapy and chemotherapy, is prone to be limited because of the presence of BCSCs in the tumor, and they may further lead to recurrence, metastasis and chemoresistance [89–94].
In 2003, CD24−CD44+lineage− cells were identified as BCSCs for the first time, which had a strong ability to initiate breast tumor, which was confirmed and used as a biomarker for BCSCs repeatedly [95]. Aldehyde dehydrogenase (ALDH) is a family of enzymes performing the function of the oxidation of intracellular aldehydes to carboxylic acids and retinoic acid (RA). Ginestier et al. found that in normal and malignant mammary cells, about 8% of normal mammary epithelial cells showed the activity of ALDH identified by ALDEFLUOR assay [16]. In addition, breast cancer cells with the ALDH activity (ALDH+) identified by ALDEFLUOR assay can initiate the xenograft tumor with as little as 500 cells. Even 50,000 ALDH-negative breast cancer cells from the same tumor can not initiate the xenograft tumor. Moreover, if the ALDH+CD24−CD44+ phenotype, as little as 20 breast tumor cells can form breast tumor [16]. Besides, many evidences from other solid tumors also indicate that ALDH can serve as the marker of CSCs [89, 96–100]. Therefore, CD24−CD44+ and ALDH+ are the most commonly used biomarkers for identification and isolation of BCSCs.
RA produced from aldehyde oxidation catalyzed by ALDH can bind to the nuclear retinoic acid receptors (RARs) or retinoid X receptors (RXRs). The heterodimer formed by RARs and RXRs recognizes a direct repeat of a 6 base pair DNA sequence motif with a 2 or 5 base pair spacer at the promoter region of many retinoid-inducible genes [101, 102]. In addition, RXR also can bind to DNA and regulate gene expression and cell differentiation as a homodimer [101]. ALDH can regulate gene expression through RAR signaling pathway, explaining the reason why ALDH plays an important role in the survival and differentiation of BCSCs (Fig. 3). Furthermore, the reprogramming of ALDH on genes expression profiling related to drug effects (genes involved in ABC transporter, DNA damage repair, clearance of reactive oxygen species) also explains the effect of ALDH+ BCSC on chemotherapeutic resistance [103].
Fig. 3.
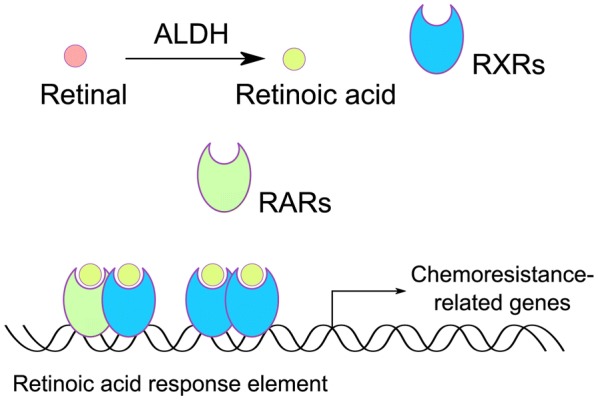
Retinoic acid signaling catalyzed by ALDH modulates the chemoresistance of cancer cells. Retinal is oxidized into retinoic acid by ALDH and then binds with RARs or RXRs. Activated RAR and RXR forms heterodimer to bind with retinoic acid response element, promoting the transcription of retinoic acid-responsive genes of which some play a vital role in driving chemotherapeutic resistance. In addition, RXRs can assemble into a homodimer to modulate the transcription while RARs cannot
A lot of signaling pathways in cells increasing stemness property, cell re-population, and self-renewal of BCSCs, indirectly promote BCSC-mediated chemoresistance of breast cancer [90, 93, 94, 104–109]. These studies provide evidences that in promoting breast cancer chemoresistance, BCSCs resemble a “hub” integrating drug resistance signalling. It can directly defend against the cytotoxic effects of anticancer drugs, or it can also be triggered by other signals to drive breast cancer cell resistance.
Cytokines promote chemotherapy resistance through enhancing BCSCs
Several studies have shown that IL-6 can regulate the self-renewal of BCSCs [110]. Furthermore, a stable equilibrium between BCSC and non-stem breast cancer cells was maintained by the amount of IL-6 secreted by BCSCs, expression level of IL-6 receptor and the whole response of non-BCSC to IL-6 [110]. Mechanistically, IL-6-JAK1-STAT3 signaling axis plays a key role in the conversion from non-BCSCs to BCSCs through regulating the expression of OCT4 gene [111]. In the study of Ibrahim et al, Syndecan-1 (CD138), a heparan sulfate proteoglycan, promotes the breast cancer stem cell phenotype via regulation of IL-6/STAT3 signaling pathways [112]. And Lin et al showed that tanshinone IIA has the potential of targeting and killing BCSCs, and can inhibit BCSCs growth in vivo and in vitro by downregulating the IL-6/STAT3/NF-κB signaling pathway [113]. These studies suggest that the promotion of IL-6 to BCSCs is generally achieved through activation of STAT3, but the effector signaling associated with the BCSCs property caused by the STAT3 activity are variegated because of the differences in the research object and the experimental system.
In addition, BCSCs rely on activated Notch signals to maintain cell survival and proliferation, and the activation of this pathway is closely related to the growth of breast tumors [114]. Inhibition of Notch1 in tumors can significantly inhibit tumor outgrowth, which is caused by promoting apoptosis and reducing the frequency of BCSCs [115]. In addition to Notch signaling, other embryonic signals are also important for stem cell regulation, such as Hedgehog and Wnt [116–119]. These pathways are closely related to the regulation of IL-6 and their interactions with IL-6 also affect the characteristics of breast cancer stem cells. In conclusion, given the critical role of BCSCs in conferring chemoresistance, enhancement of IL-6 to BCSCs population reveals the clew of that it can promote the resistance of breast cancer cells to chemotherapeutic drugs [34].
In addition to IL-6, some studies suggested that IL-8, CCL2 and TGF-β can also promote the characteristics of BCSCs and give rise to chemotherapeutic resistance [38, 42, 120–123].
Therapeutic implications of interfering the interactions between cytokines and BCSCs in chemoresistance
The interaction between cytokines and BCSCs is pivotal for the formation of chemotherapy resistance. Cytokines can act on the surface receptors on breast cancer stem cell membrane through autocrine or paracrine, and promote the proliferation, survival and self-renewal of BCSCs. In response, BCSCs may also increase the secretion of specific factors under the stress of chemotherapeutic drugs to regulate their survival signaling. The augment in the levels of specific cytokines will trigger the response of non-tumor cells in tumor microenvironment, such as secreting cytokines to remodel the BCSCs niche, which may be more conducive to the survival of BCSCs (Fig. 4). When BCSCs are influenced by cytokines, the expression of genes affiliated with resistance may be altered, such as the increase in the level of transporter protein, expression of anti-apoptotic genes, and activation of DNA repair genes. Eventually, BCSCs and specific cytokines collude in doing evil as chemotherapy resistance through reciprocal modulation.
Fig. 4.

The interplay of cytokines and BCSCs on regulating chemotherapeutic resistance. Under the stress of chemotherapeutic drugs, a variety of different sources of cytokines can regulate the self-renewal and promote the survival of BCSCs, leading to drug resistance. Cytokines are a convenient “bridge” between BCSCs and chemoresistance (left panel). If the regulatory signaling to BCSCs is interfered by cytokine blockade, the support of cytokine signals for BCSC chemo-resistance will be broken, increasing the sensitivity of breast cancer chemotherapy (right panel)
Therefore, interfering the interaction between BCSC and cytokines may bring great benefits to the improvement of chemotherapy resistance in breast cancer. In view of the role of IL-8 in mediating BCSC self-renewal and breast cancer chemoresistance, several IL-8 releasing inhibitors, small-molecule CXCR1/2 inhibitors and neutralizing antibodies against IL-8 and CXCR1/2 have been reported during the past two decades [68]. Combination of blocking IGF signaling and paclitaxel can significantly reduce tumor cell proliferation and pulmonary metastasis compared with treatment of paclitaxel alone [73]. This provides a theoretical basis for the further development of combinatorial strategy of paclitaxel and IGF blockers in the treatment of invasive breast cancer. Moreover, IGF-positive stromal cells may be designated as potential biomarker for auxiliary diagnosis [73].
Conclusion and perspective
Chemotherapy resistance in breast cancer is often accompanied with alterations of specific cytokine levels in tumor tissues. The mutual regulation between these cytokines and BCSCs has weakened the effect of chemotherapy and reduced the survival of breast cancer patients.
There also exists great heterogeneity in the populations of BCSCs, which leads to limited therapy efficacy of targeting BCSCs directly [124]. Taking into account the necessity of cytokines for BCSC self-renewal and survival, and the combinational therapeutic strategy of targeting BCSCs and neutralizing cytokines may bring great hope to the improvement of the survival for breast cancer patients (Fig. 4).
Authors’ contributions
WLC designed, drafted, wrote, and reviewed the manuscript. YYQ wrote and reviewed the manuscript. SLL reviewed, modified and improved the manuscript. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
The work was supported by NSFC Grant [81530075], the National Key Research and Development Program of China [Stem Cell and Translational Research 2016YFA0101202], the MOST Grant [2015CB553800], NSFC Grant [81472741].
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- TME
tumor microenvironment
- BCSCs
breast cancer stem cells
- JAK2
Janus kinase 2
- STAT3
signal transducer and activator of transcription 3
- PI3K
phosphatidylinositol-4,5-bisphosphate 3-kinase
- AKT
AKT serine/threonine kinase
- Tim-3
Hepatitis A virus cellular receptor 2
- HBV
Hepatitis B virus
- NF-kappa B
nuclear factor kappa B
- OPN
secreted phosphoprotein 1
- HIF-1
hypoxia inducible factor 1
- VCAM-1
vascular cell adhesion molecule 1
- TLR4
toll like receptor 4
- CXCR
C-X-C motif chemokine receptor
- GPCR
G protein-coupled receptors
- TGF-ß
transforming growth factor beta
- EMT
epithelial-to-mesenchymal transition
- MSH2
MutS homolog 2
- M-CSF
colony stimulating factor 1
- TNFα
tumor necrosis factor
- ECM
extracellular matrix
- TAM
tumor-associated macrophages
- IGFs
insulin-like growth factors
- CAF
cancer-associated fibroblasts
- Twist1
twist family bHLH transcription factor 1
- CXCL12
C-X-C motif chemokine ligand 12
- MSCs
mesenchymal stromal cells
- SMAD4
SMAD family member 4
- ALDH
aldehyde dehydrogenase
- RA
retinoic acid
- RARs
retinoic acid receptors
- RXRs
retinoid X receptors
- ABC
ATP binding cassette
Contributor Information
Weilong Chen, Email: cwl123@mail.ustc.edu.cn.
Yuanyuan Qin, Email: qinyy@mail.ustc.edu.cn.
Suling Liu, Phone: 086-21-34771023, Email: suling@fudan.edu.cn.
References
- 1.Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7–30. doi: 10.3322/caac.21442. [DOI] [PubMed] [Google Scholar]
- 2.Tao Y, Li M, Kim B, Auguste DT. Incorporating gold nanoclusters and target-directed liposomes as a synergistic amplified colorimetric sensor for HER2-positive breast cancer cell detection. Theranostics. 2017;7(4):899–911. doi: 10.7150/thno.17927. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Hwang KT, Kim EK, Jung SH, Lee ES, Kim SI, Lee S, et al. Tamoxifen therapy improves overall survival in luminal A subtype of ductal carcinoma in situ: a study based on nationwide Korean Breast Cancer Registry database. Breast Cancer Res Treat. 2018 doi: 10.1007/s10549-018-4681-6. [DOI] [PubMed] [Google Scholar]
- 4.Greten FR. Cancer: tumour stem-cell surprises. Nature. 2017;543(7647):626–627. doi: 10.1038/543626a. [DOI] [PubMed] [Google Scholar]
- 5.Ford AM, Mansur MB, Furness CL, van Delft FW, Okamura J, Suzuki T, et al. Protracted dormancy of pre-leukemic stem cells. Leukemia. 2015;29(11):2202–2207. doi: 10.1038/leu.2015.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kakarala M, Wicha MS. Implications of the cancer stem-cell hypothesis for breast cancer prevention and therapy. J Clin Oncol. 2008;26(17):2813–2820. doi: 10.1200/JCO.2008.16.3931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Brooks MD, Wicha MS. Tumor twitter: cellular communication in the breast cancer stem cell niche. Cancer Discov. 2015;5(5):469–471. doi: 10.1158/2159-8290.CD-15-0327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gilbertson RJ, Graham TA. Cancer: resolving the stem-cell debate. Nature. 2012;488(7412):462–463. doi: 10.1038/nature11480. [DOI] [PubMed] [Google Scholar]
- 9.Chan CH, Morrow JK, Li CF, Gao Y, Jin G, Moten A, et al. Pharmacological inactivation of Skp2 SCF ubiquitin ligase restricts cancer stem cell traits and cancer progression. Cell. 2013;154(3):556–568. doi: 10.1016/j.cell.2013.06.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Cordenonsi M, Zanconato F, Azzolin L, Forcato M, Rosato A, Frasson C, et al. The Hippo transducer TAZ confers cancer stem cell-related traits on breast cancer cells. Cell. 2011;147(4):759–772. doi: 10.1016/j.cell.2011.09.048. [DOI] [PubMed] [Google Scholar]
- 11.Pece S, Tosoni D, Confalonieri S, Mazzarol G, Vecchi M, Ronzoni S, et al. Biological and molecular heterogeneity of breast cancers correlates with their cancer stem cell content. Cell. 2010;140(1):62–73. doi: 10.1016/j.cell.2009.12.007. [DOI] [PubMed] [Google Scholar]
- 12.Sharma B, Varney ML, Saxena S, Wu L, Singh RK. Induction of CXCR2 ligands, stem cell-like phenotype, and metastasis in chemotherapy-resistant breast cancer cells. Cancer Lett. 2016;372(2):192–200. doi: 10.1016/j.canlet.2015.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gallego-Ortega D, Ledger A, Roden DL, Law AM, Magenau A, Kikhtyak Z, et al. ELF5 drives lung metastasis in luminal breast cancer through recruitment of Gr1+ CD11b+ myeloid-derived suppressor cells. PLoS Biol. 2015;13(12):e1002330. doi: 10.1371/journal.pbio.1002330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Guo L, Zheng P, Fan H, Wang H, Xu W, Zhou W. Ultrasound reverses chemoresistance in breast cancer stem cell like cells by altering ABCG2 expression. Biosci Rep. 2017 doi: 10.1042/BSR20171137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Wang T, Fahrmann JF, Lee H, Li YJ, Tripathi SC, Yue C, et al. JAK/STAT3-regulated fatty acid beta-oxidation is critical for breast cancer stem cell self-renewal and chemoresistance. Cell Metab. 2018;27(1):136–150. doi: 10.1016/j.cmet.2017.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, et al. ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome. Cell Stem Cell. 2007;1(5):555–567. doi: 10.1016/j.stem.2007.08.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Liu S, Cong Y, Wang D, Sun Y, Deng L, Liu Y, et al. Breast cancer stem cells transition between epithelial and mesenchymal states reflective of their normal counterparts. Stem Cell Rep. 2014;2(1):78–91. doi: 10.1016/j.stemcr.2013.11.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Tan C, Hu W, He Y, Zhang Y, Zhang G, Xu Y, et al. Cytokine-mediated therapeutic resistance in breast cancer. Cytokine. 2018;108:151–159. doi: 10.1016/j.cyto.2018.03.020. [DOI] [PubMed] [Google Scholar]
- 19.Zhao M, Liu Y, Liu R, Qi J, Hou Y, Chang J, et al. Upregulation of IL-11, an IL-6 family cytokine, promotes tumor progression and correlates with poor prognosis in non-small cell lung cancer. Cell Physiol Biochem. 2018;45(6):2213–2224. doi: 10.1159/000488166. [DOI] [PubMed] [Google Scholar]
- 20.Koyama S, Akbay EA, Li YY, Aref AR, Skoulidis F, Herter-Sprie GS, et al. STK11/LKB1 deficiency promotes neutrophil recruitment and proinflammatory cytokine production to suppress T-cell activity in the lung tumor microenvironment. Cancer Res. 2016;76(5):999–1008. doi: 10.1158/0008-5472.CAN-15-1439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kulbe H, Chakravarty P, Leinster DA, Charles KA, Kwong J, Thompson RG, et al. A dynamic inflammatory cytokine network in the human ovarian cancer microenvironment. Cancer Res. 2012;72(1):66–75. doi: 10.1158/0008-5472.CAN-11-2178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Korkaya H, Liu S, Wicha MS. Breast cancer stem cells, cytokine networks, and the tumor microenvironment. J Clin Invest. 2011;121(10):3804–3809. doi: 10.1172/JCI57099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bharti R, Dey G, Mandal M. Cancer development, chemoresistance, epithelial to mesenchymal transition and stem cells: a snapshot of IL-6 mediated involvement. Cancer Lett. 2016;375(1):51–61. doi: 10.1016/j.canlet.2016.02.048. [DOI] [PubMed] [Google Scholar]
- 24.Cortini M, Massa A, Avnet S, Bonuccelli G, Baldini N. Tumor-activated mesenchymal stromal cells promote osteosarcoma stemness and migratory potential via IL-6 secretion. PLoS ONE. 2016;11(11):e0166500. doi: 10.1371/journal.pone.0166500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.He G, Dhar D, Nakagawa H, Font-Burgada J, Ogata H, Jiang Y, et al. Identification of liver cancer progenitors whose malignant progression depends on autocrine IL-6 signaling. Cell. 2013;155(2):384–396. doi: 10.1016/j.cell.2013.09.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kishimoto T. IL-6: from its discovery to clinical applications. Int Immunol. 2010;22(5):347–352. doi: 10.1093/intimm/dxq030. [DOI] [PubMed] [Google Scholar]
- 27.Johnson DE, O’Keefe RA, Grandis JR. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat Rev Clin Oncol. 2018;15(4):234–248. doi: 10.1038/nrclinonc.2018.8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhang X, Hu F, Li G, Li G, Yang X, Liu L, et al. Human colorectal cancer-derived mesenchymal stem cells promote colorectal cancer progression through IL-6/JAK2/STAT3 signaling. Cell Death Dis. 2018;9(2):25. doi: 10.1038/s41419-017-0176-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang H, Song Y, Yang H, Liu Z, Gao L, Liang X, et al. Tumor cell-intrinsic Tim-3 promotes liver cancer via NF-kappaB/IL-6/STAT3 axis. Oncogene. 2018 doi: 10.1038/s41388-018-0140-4. [DOI] [PubMed] [Google Scholar]
- 30.Qin X, Yan M, Wang X, Xu Q, Wang X, Zhu X, et al. Cancer-associated fibroblast-derived IL-6 promotes Head and neck cancer progression via the osteopontin-NF-kappa B signaling pathway. Theranostics. 2018;8(4):921–940. doi: 10.7150/thno.22182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Li J, He K, Liu P, Xu LX. Iron participated in breast cancer chemoresistance by reinforcing IL-6 paracrine loop. Biochem Biophys Res Commun. 2016;475(2):154–160. doi: 10.1016/j.bbrc.2016.05.064. [DOI] [PubMed] [Google Scholar]
- 32.Wang K, Zhu X, Zhang K, Yin Y, Chen Y, Zhang T. Interleukin-6 contributes to chemoresistance in MDA-MB-231 cells via targeting HIF-1alpha. J Biochem Mol Toxicol. 2018;32(3):e22039. doi: 10.1002/jbt.22039. [DOI] [PubMed] [Google Scholar]
- 33.Wang PC, Weng CC, Hou YS, Jian SF, Fang KT, Hou MF, et al. Activation of VCAM-1 and its associated molecule CD44 leads to increased malignant potential of breast cancer cells. Int J Mol Sci. 2014;15(3):3560–3579. doi: 10.3390/ijms15033560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Saha S, Mukherjee S, Khan P, Kajal K, Mazumdar M, Manna A, et al. Aspirin suppresses the acquisition of chemoresistance in breast cancer by disrupting an NF-kappa B-IL6 signaling axis responsible for the generation of cancer stem cells. Cancer Res. 2016;76(7):2000–2012. doi: 10.1158/0008-5472.CAN-15-1360. [DOI] [PubMed] [Google Scholar]
- 35.Russo RC, Garcia CC, Teixeira MM, Amaral FA. The CXCL8/IL-8 chemokine family and its receptors in inflammatory diseases. Expert Rev Clin Immunol. 2014;10(5):593–619. doi: 10.1586/1744666X.2014.894886. [DOI] [PubMed] [Google Scholar]
- 36.Dominguez C, McCampbell KK, David JM, Palena C. Neutralization of IL-8 decreases tumor PMN-MDSCs and reduces mesenchymalization of claudin-low triple-negative breast cancer. JCI Insight. 2017;2(21):e94296. doi: 10.1172/jci.insight.94296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Sootichote R, Thuwajit P, Singsuksawat E, Warnnissorn M, Yenchitsomanus PT, Ithimakin S, et al. Compound A attenuates toll-like receptor 4-mediated paclitaxel resistance in breast cancer and melanoma through suppression of IL-8. BMC Cancer. 2018;18(1):231. doi: 10.1186/s12885-018-4155-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Jia D, Li L, Andrew S, Allan D, Li X, Lee J, et al. An autocrine inflammatory forward-feedback loop after chemotherapy withdrawal facilitates the repopulation of drug-resistant breast cancer cells. Cell Death Dis. 2017;8(7):e2932. doi: 10.1038/cddis.2017.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Liu Q, Li A, Tian Y, Wu JD, Liu Y, Li T, et al. The CXCL8–CXCR1/2 pathways in cancer. Cytokine Growth Factor Rev. 2016;31:61–71. doi: 10.1016/j.cytogfr.2016.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Meulmeester E, Ten Dijke P. The dynamic roles of TGF-beta in cancer. J Pathol. 2011;223(2):205–218. doi: 10.1002/path.2785. [DOI] [PubMed] [Google Scholar]
- 41.Santibanez JF, Quintanilla M, Bernabeu C. TGF-beta/TGF-beta receptor system and its role in physiological and pathological conditions. Clin Sci (Lond). 2011;121(6):233–251. doi: 10.1042/CS20110086. [DOI] [PubMed] [Google Scholar]
- 42.Bhola NE, Balko JM, Dugger TC, Kuba MG, Sanchez V, Sanders M, et al. TGF-beta inhibition enhances chemotherapy action against triple-negative breast cancer. J Clin Invest. 2013;123(3):1348–1358. doi: 10.1172/JCI65416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhong S, Ma T, Zhang X, Lv M, Chen L, Tang J, et al. MicroRNA expression profiling and bioinformatics analysis of dysregulated microRNAs in vinorelbine-resistant breast cancer cells. Gene. 2015;556(2):113–118. doi: 10.1016/j.gene.2014.11.046. [DOI] [PubMed] [Google Scholar]
- 44.Karicheva O, Rodriguez-Vargas JM, Wadier N, Martin-Hernandez K, Vauchelles R, Magroun N, et al. PARP3 controls TGFβ and ROS driven epithelial-to-mesenchymal transition and stemness by stimulating a TG2-Snail-E-cadherin axis. Oncotarget. 2016;7(39):64109. doi: 10.18632/oncotarget.11627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gulei D, Mehterov N, Ling H, Stanta G, Braicu C, Berindan-Neagoe I. The, “good-cop bad-cop” TGF-beta role in breast cancer modulated by non-coding RNAs. Biochim Biophys Acta. 2017;1861(7):1661–1675. doi: 10.1016/j.bbagen.2017.04.007. [DOI] [PubMed] [Google Scholar]
- 46.Butz H, Racz K, Hunyady L, Patocs A. Crosstalk between TGF-beta signaling and the microRNA machinery. Trends Pharmacol Sci. 2012;33(7):382–393. doi: 10.1016/j.tips.2012.04.003. [DOI] [PubMed] [Google Scholar]
- 47.Yu Y, Wang Y, Ren X, Tsuyada A, Li A, Liu LJ, et al. Context-dependent bidirectional regulation of the MutS homolog 2 by transforming growth factor beta contributes to chemoresistance in breast cancer cells. Mol Cancer Res. 2010;8(12):1633–1642. doi: 10.1158/1541-7786.MCR-10-0362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Jones SA, Scheller J, Rose-John S. Therapeutic strategies for the clinical blockade of IL-6/gp130 signaling. J Clin Invest. 2011;121(9):3375–3383. doi: 10.1172/JCI57158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kurzrock R, Voorhees PM, Casper C, Furman RR, Fayad L, Lonial S, et al. A phase I, open-label study of siltuximab, an anti-IL-6 monoclonal antibody, in patients with B-cell non-Hodgkin’s lymphoma, multiple myeloma, or Castleman’s disease. Clin Cancer Res. 2013 doi: 10.1158/1078-0432.CCR-12-3349. [DOI] [PubMed] [Google Scholar]
- 50.San-Miguel J, Bladé J, Shpilberg O, Grosicki S, Maloisel F, Min C-K, et al. Phase 2 randomized study of bortezomib–melphalan–prednisone with or without siltuximab (anti–IL-6) in multiple myeloma. Blood. 2014;123(26):4136–4142. doi: 10.1182/blood-2013-12-546374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Voorhees PM, Manges RF, Sonneveld P, Jagannath S, Somlo G, Krishnan A, et al. A phase 2 multicentre study of siltuximab, an anti-interleukin-6 monoclonal antibody, in patients with relapsed or refractory multiple myeloma. Br J Haematol. 2013;161(3):357–366. doi: 10.1111/bjh.12266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Angevin E, Tabernero J, Elez ME, Cohen SJ, Bahleda R, Van Laethem J-L, et al. A phase 1/2, multiple-dose, dose-escalation study of siltuximab, an anti-interleukin-6 monoclonal antibody, in patients with advanced solid tumors. Clin Cancer Res. 2014 doi: 10.1158/1078-0432.CCR-13-2200. [DOI] [PubMed] [Google Scholar]
- 53.Schott AF, Goldstein LJ, Cristofanilli M, Ruffini PA, McCanna S, Reuben JM, et al. Phase Ib pilot study to evaluate reparixin in combination with weekly paclitaxel in patients with HER-2—negative metastatic breast cancer. Clin Cancer Res. 2017 doi: 10.1158/1078-0432.CCR-16-2748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Goldstein LJ, Perez RP, Yardley DA, Han LK, Reuben JM, McCanna S, et al. Abstract CT057: a single-arm, preoperative, pilot study to evaluate the safety and biological effects of orally administered reparixin in early breast cancer patients who are candidates for surgery. Cancer Res. 2016 doi: 10.1158/1538-7445.AM2016-CT057. [DOI] [Google Scholar]
- 55.Rodon J, Carducci MA, Sepulveda-Sánchez JM, Azaro A, Calvo E, Seoane J, et al. First-in-human dose study of the novel transforming growth factor-β receptor I kinase inhibitor LY2157299 monohydrate in patients with advanced cancer and glioma. Clin Cancer Res. 2014 doi: 10.1158/1078-0432.CCR-14-1380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Rodón J, Carducci M, Sepulveda-Sánchez JM, Azaro A, Calvo E, Seoane J, et al. Pharmacokinetic, pharmacodynamic and biomarker evaluation of transforming growth factor-β receptor I kinase inhibitor, galunisertib, in phase 1 study in patients with advanced cancer. Invest New Drugs. 2015;33(2):357–370. doi: 10.1007/s10637-014-0192-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Faivre SJ, Santoro A, Gane E, Kelley RK, Hourmand IO, Assenat E, et al. A phase 2 study of galunisertib, a novel transforming growth factor-beta (TGF-β) receptor I kinase inhibitor, in patients with advanced hepatocellular carcinoma (HCC) and low serum alpha fetoprotein (AFP) J Clin Oncol. 2016 doi: 10.1200/JCO.2016.34.15_suppl.4070. [DOI] [Google Scholar]
- 58.Brandes AA, Carpentier AF, Kesari S, Sepulveda-Sanchez JM, Wheeler HR, Chinot O, et al. A phase II randomized study of galunisertib monotherapy or galunisertib plus lomustine compared with lomustine monotherapy in patients with recurrent glioblastoma. Neuro-oncology. 2016;18(8):1146–1156. doi: 10.1093/neuonc/now009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Brandes AA, Carpentier AF, Kesari S, Sepulveda J, Wheeler H, Chinot OL, et al. A phase II study of galunisertib monotherapy or galunisertib plus lomustine compared to lomustine monotherapy in recurrent glioblastoma. J Clin Oncol. 2015 doi: 10.1200/jco.2015.33.15_suppl.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fujiwara Y, Nokihara H, Yamada Y, Yamamoto N, Sunami K, Utsumi H, et al. Phase 1 study of galunisertib, a TGF-beta receptor I kinase inhibitor, in Japanese patients with advanced solid tumors. Cancer Chemother Pharmacol. 2015;76(6):1143–1152. doi: 10.1007/s00280-015-2895-4. [DOI] [PubMed] [Google Scholar]
- 61.Melisi D, Garcia-Carbonero R, Macarulla T, Pezet D, Deplanque G, Fuchs M, et al. A phase II, double-blind study of galunisertib + gemcitabine (GG) vs gemcitabine + placebo (GP) in patients (pts) with unresectable pancreatic cancer (PC) J Clin Oncol. 2016 doi: 10.1200/JCO.2016.34.15_suppl.4019. [DOI] [Google Scholar]
- 62.Necchi A, Giannatempo P, Mariani L, Fare E, Raggi D, Pennati M, et al. PF-03446962, a fully-human monoclonal antibody against transforming growth-factor β (TGFβ) receptor ALK1, in pre-treated patients with urothelial cancer: an open label, single-group, phase 2 trial. Invest New Drugs. 2014;32(3):555–560. doi: 10.1007/s10637-014-0074-9. [DOI] [PubMed] [Google Scholar]
- 63.Menacho-Marquez M, Garcia-Escudero R, Ojeda V, Abad A, Delgado P, Costa C, et al. The Rho exchange factors Vav2 and Vav3 favor skin tumor initiation and promotion by engaging extracellular signaling loops. PLoS Biol. 2013;11(7):e1001615. doi: 10.1371/journal.pbio.1001615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Gligorijevic B, Bergman A, Condeelis J. Multiparametric classification links tumor microenvironments with tumor cell phenotype. PLoS Biol. 2014;12(11):e1001995. doi: 10.1371/journal.pbio.1001995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Camorani S, Hill BS, Fontanella R, Greco A, Gramanzini M, Auletta L, et al. Inhibition of bone marrow-derived mesenchymal stem cells homing towards triple-negative breast cancer microenvironment using an Anti-PDGFRbeta aptamer. Theranostics. 2017;7(14):3595–3607. doi: 10.7150/thno.18974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.McGranahan N, Swanton C. Cancer evolution constrained by the immune microenvironment. Cell. 2017;170(5):825–827. doi: 10.1016/j.cell.2017.08.012. [DOI] [PubMed] [Google Scholar]
- 67.Iida N, Dzutsev A, Stewart CA, Smith L, Bouladoux N, Weingarten RA, et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013;342(6161):967–970. doi: 10.1126/science.1240527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Ha H, Debnath B, Neamati N. Role of the CXCL8–CXCR1/2 axis in cancer and inflammatory diseases. Theranostics. 2017;7(6):1543–1588. doi: 10.7150/thno.15625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Xie HY, Shao ZM, Li DQ. Tumor microenvironment: driving forces and potential therapeutic targets for breast cancer metastasis. Chin J Cancer. 2017;36(1):36. doi: 10.1186/s40880-017-0202-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Xing F, Liu Y, Sharma S, Wu K, Chan MD, Lo HW, et al. Activation of the c-Met pathway mobilizes an inflammatory network in the brain microenvironment to promote brain metastasis of breast cancer. Cancer Res. 2016;76(17):4970–4980. doi: 10.1158/0008-5472.CAN-15-3541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Guo C, Chen Y, Gao W, Chang A, Ye Y, Shen W, et al. Liposomal nanoparticles carrying anti-IL6R antibody to the tumour microenvironment inhibit metastasis in two molecular subtypes of breast cancer mouse models. Theranostics. 2017;7(3):775–788. doi: 10.7150/thno.17237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Phuengkham H, Song C, Um SH, Lim YT. Implantable synthetic immune niche for spatiotemporal modulation of tumor-derived immunosuppression and systemic antitumor immunity: postoperative immunotherapy. Adv Mater. 2018;30:1706719. doi: 10.1002/adma.201706719. [DOI] [PubMed] [Google Scholar]
- 73.Ireland L, Santos A, Campbell F, Figueiredo C, Hammond D, Ellies LG, et al. Blockade of insulin-like growth factors increases efficacy of paclitaxel in metastatic breast cancer. Oncogene. 2018;37(15):2022–2036. doi: 10.1038/s41388-017-0115-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Choi J, Gyamfi J, Jang H, Koo JS. The role of tumor-associated macrophage in breast cancer biology. Histol Histopathol. 2018;33(2):133–145. doi: 10.14670/HH-11-916. [DOI] [PubMed] [Google Scholar]
- 75.Yang C, He L, He P, Liu Y, Wang W, He Y, et al. Increased drug resistance in breast cancer by tumor-associated macrophages through IL-10/STAT3/bcl-2 signaling pathway. Med Oncol. 2015;32(2):352. doi: 10.1007/s12032-014-0352-6. [DOI] [PubMed] [Google Scholar]
- 76.Ohlund D, Elyada E, Tuveson D. Fibroblast heterogeneity in the cancer wound. J Exp Med. 2014;211(8):1503–1523. doi: 10.1084/jem.20140692. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Smith NR, Baker D, Farren M, Pommier A, Swann R, Wang X, et al. Tumor stromal architecture can define the intrinsic tumor response to VEGF-targeted therapy. Clin Cancer Res. 2013;19(24):6943–6956. doi: 10.1158/1078-0432.CCR-13-1637. [DOI] [PubMed] [Google Scholar]
- 78.Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16(9):582–598. doi: 10.1038/nrc.2016.73. [DOI] [PubMed] [Google Scholar]
- 79.Chang JT, Mani SA. Sheep, wolf, or werewolf: cancer stem cells and the epithelial-to-mesenchymal transition. Cancer Lett. 2013;341(1):16–23. doi: 10.1016/j.canlet.2013.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Zheng X, Carstens JL, Kim J, Scheible M, Kaye J, Sugimoto H, et al. Epithelial-to-mesenchymal transition is dispensable for metastasis but induces chemoresistance in pancreatic cancer. Nature. 2015;527(7579):525. doi: 10.1038/nature16064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.LeBleu VS, O’Connell JT, Gonzalez Herrera KN, Wikman H, Pantel K, Haigis MC, et al. PGC-1alpha mediates mitochondrial biogenesis and oxidative phosphorylation in cancer cells to promote metastasis. Nat Cell Biol. 2014;16(10):992–1003. doi: 10.1038/ncb3039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Ozdemir BC, Pentcheva-Hoang T, Carstens JL, Zheng X, Wu CC, Simpson TR, et al. Depletion of carcinoma-associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell. 2014;25(6):719–734. doi: 10.1016/j.ccr.2014.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Lee KW, Yeo SY, Sung CO, Kim SH. Twist1 is a key regulator of cancer-associated fibroblasts. Cancer Res. 2015;75(1):73–85. doi: 10.1158/0008-5472.CAN-14-0350. [DOI] [PubMed] [Google Scholar]
- 84.Luo H, Tu G, Liu Z, Liu M. Cancer-associated fibroblasts: a multifaceted driver of breast cancer progression. Cancer Lett. 2015;361(2):155–163. doi: 10.1016/j.canlet.2015.02.018. [DOI] [PubMed] [Google Scholar]
- 85.Lippitz BE. Cytokine patterns in patients with cancer: a systematic review. Lancet Oncol. 2013;14(6):e218–e228. doi: 10.1016/S1470-2045(12)70582-X. [DOI] [PubMed] [Google Scholar]
- 86.Esquivel-Velázquez M, Ostoa-Saloma P, Palacios-Arreola MI, Nava-Castro KE, Castro JI, Morales-Montor J. The role of cytokines in breast cancer development and progression. J Interf Cytok Res. 2015;35(1):1–16. doi: 10.1089/jir.2014.0026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Newman G, Gonzalez-Perez RR. Leptin–cytokine crosstalk in breast cancer. Mol Cell Endocrinol. 2014;382(1):570–582. doi: 10.1016/j.mce.2013.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Skolekova S, Matuskova M, Bohac M, Toro L, Durinikova E, Tyciakova S, et al. Cisplatin-induced mesenchymal stromal cells-mediated mechanism contributing to decreased antitumor effect in breast cancer cells. Cell Commun Signal. 2016;14:4. doi: 10.1186/s12964-016-0127-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Abdullah LN, Chow EK. Mechanisms of chemoresistance in cancer stem cells. Clin Transl Med. 2013;2(1):3. doi: 10.1186/2001-1326-2-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Tam WL, Lu H, Buikhuisen J, Soh BS, Lim E, Reinhardt F, et al. Protein kinase C α is a central signaling node and therapeutic target for breast cancer stem cells. Cancer Cell. 2013;24(3):347–364. doi: 10.1016/j.ccr.2013.08.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Lin X, Chen W, Wei F, Zhou BP, Hung MC, Xie X. Nanoparticle delivery of miR-34a eradicates long-term-cultured breast cancer stem cells via targeting C22ORF28 directly. Theranostics. 2017;7(19):4805–4824. doi: 10.7150/thno.20771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Park EY, Chang E, Lee EJ, Lee H-W, Kang H-G, Chun K-H, et al. Targeting of miR34a–NOTCH1 axis reduced breast cancer stemness and chemoresistance. Cancer Res. 2014;74(24):7573–7582. doi: 10.1158/0008-5472.CAN-14-1140. [DOI] [PubMed] [Google Scholar]
- 93.Bartucci M, Dattilo R, Moriconi C, Pagliuca A, Mottolese M, Federici G, et al. TAZ is required for metastatic activity and chemoresistance of breast cancer stem cells. Oncogene. 2015;34(6):681–690. doi: 10.1038/onc.2014.5. [DOI] [PubMed] [Google Scholar]
- 94.Samanta D, Gilkes DM, Chaturvedi P, Xiang L, Semenza GL. Hypoxia-inducible factors are required for chemotherapy resistance of breast cancer stem cells. Proc Natl Acad Sci USA. 2014;111(50):E5429–E5438. doi: 10.1073/pnas.1421438111. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 95.Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF. Prospective identification of tumorigenic breast cancer cells. Proc Natl Acad Sci USA. 2003;100(7):3983–3988. doi: 10.1073/pnas.0530291100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Giudice FS, Pinto DS, Jr, Nör JE, Squarize CH, Castilho RM. Inhibition of histone deacetylase impacts cancer stem cells and induces epithelial-mesenchyme transition of head and neck cancer. PLoS ONE. 2013;8(3):e58672. doi: 10.1371/journal.pone.0058672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kim SK, Kim H, D-h Lee, T-s Kim, Kim T, Chung C, et al. Reversing the intractable nature of pancreatic cancer by selectively targeting ALDH-high, therapy-resistant cancer cells. PLoS ONE. 2013;8(10):e78130. doi: 10.1371/journal.pone.0078130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Sun J-H, Luo Q, Liu L-L, Song G-B. Liver cancer stem cell markers: progression and therapeutic implications. World J Gastroenterol. 2016;22(13):3547. doi: 10.3748/wjg.v22.i13.3547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Nishida S, Hirohashi Y, Torigoe T, Inoue R, Kitamura H, Tanaka T, et al. Prostate cancer stem-like cells/cancer-initiating cells have an autocrine system of hepatocyte growth factor. Cancer Sci. 2013;104(4):431–436. doi: 10.1111/cas.12104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Huang C-P, Tsai M-F, Chang T-H, Tang W-C, Chen S-Y, Lai H-H, et al. ALDH-positive lung cancer stem cells confer resistance to epidermal growth factor receptor tyrosine kinase inhibitors. Cancer Lett. 2013;328(1):144–151. doi: 10.1016/j.canlet.2012.08.021. [DOI] [PubMed] [Google Scholar]
- 101.Elizondo G, Medina-Diaz IM, Cruz R, Gonzalez FJ, Vega L. Retinoic acid modulates retinaldehyde dehydrogenase 1 gene expression through the induction of GADD153-C/EBPbeta interaction. Biochem Pharmacol. 2009;77(2):248–257. doi: 10.1016/j.bcp.2008.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Alvarez S, Germain P, Alvarez R, Rodriguez-Barrios F, Gronemeyer H, de Lera AR. Structure, function and modulation of retinoic acid receptor beta, a tumor suppressor. Int J Biochem Cell Biol. 2007;39(7–8):1406–1415. doi: 10.1016/j.biocel.2007.02.010. [DOI] [PubMed] [Google Scholar]
- 103.Shima H, Yamada A, Ishikawa T, Endo I. Are breast cancer stem cells the key to resolving clinical issues in breast cancer therapy? Gland Surg. 2017;6(1):82–88. doi: 10.21037/gs.2016.08.03. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.W-x Chen, X-m Liu, M-m Lv, Chen L, J-h Zhao, S-l Zhong, et al. Exosomes from drug-resistant breast cancer cells transmit chemoresistance by a horizontal transfer of microRNAs. PLoS ONE. 2014;9(4):e95240. doi: 10.1371/journal.pone.0095240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Schwarzenbacher D, Balic M, Pichler M. The role of microRNAs in breast cancer stem cells. Int J Mol Sci. 2013;14(7):14712–14723. doi: 10.3390/ijms140714712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Liu P, Kumar I, Brown S, Kannappan V, Tawari P, Tang J, et al. Disulfiram targets cancer stem-like cells and reverses resistance and cross-resistance in acquired paclitaxel-resistant triple-negative breast cancer cells. Br J Cancer. 2013;109(7):1876. doi: 10.1038/bjc.2013.534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Luo M, Brooks M, Wicha MS. Epithelial-mesenchymal plasticity of breast cancer stem cells: implications for metastasis and therapeutic resistance. Curr Pharm Des. 2015;21(10):1301–1310. doi: 10.2174/1381612821666141211120604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Ma X, Zhou J, Zhang C-X, Li X-Y, Li N, Ju R-J, et al. Modulation of drug-resistant membrane and apoptosis proteins of breast cancer stem cells by targeting berberine liposomes. Biomaterials. 2013;34(18):4452–4465. doi: 10.1016/j.biomaterials.2013.02.066. [DOI] [PubMed] [Google Scholar]
- 109.Mallini P, Lennard T, Kirby J, Meeson A. Epithelial-to-mesenchymal transition: what is the impact on breast cancer stem cells and drug resistance. Cancer Treat Rev. 2014;40(3):341–348. doi: 10.1016/j.ctrv.2013.09.008. [DOI] [PubMed] [Google Scholar]
- 110.Dethlefsen C, Hojfeldt G, Hojman P. The role of intratumoral and systemic IL-6 in breast cancer. Breast Cancer Res Treat. 2013;138(3):657–664. doi: 10.1007/s10549-013-2488-z. [DOI] [PubMed] [Google Scholar]
- 111.Kim SY, Kang JW, Song X, Kim BK, Yoo YD, Kwon YT, et al. Role of the IL-6–JAK1–STAT3-Oct-4 pathway in the conversion of non-stem cancer cells into cancer stem-like cells. Cell Signal. 2013;25(4):961–969. doi: 10.1016/j.cellsig.2013.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Ibrahim SA, Hassan H, Vilardo L, Kumar SK, Kumar AV, Kelsch R, et al. Syndecan-1 (CD138) modulates triple-negative breast cancer stem cell properties via regulation of LRP-6 and IL-6-mediated STAT3 signaling. PLoS ONE. 2013;8(12):e85737. doi: 10.1371/journal.pone.0085737. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Lin C, Wang L, Wang H, Yang L, Guo H, Wang X. Tanshinone IIA inhibits breast cancer stem cells growth in vitro and in vivo through attenuation of IL-6/STAT3/NF-κB signaling pathways. J Cell Biochem. 2013;114(9):2061–2070. doi: 10.1002/jcb.24553. [DOI] [PubMed] [Google Scholar]
- 114.Suman S, Das TP, Damodaran C. Silencing NOTCH signaling causes growth arrest in both breast cancer stem cells and breast cancer cells. Br J Cancer. 2013;109(10):2587–2596. doi: 10.1038/bjc.2013.642. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Qiu M, Peng Q, Jiang I, Carroll C, Han G, Rymer I, et al. Specific inhibition of Notch1 signaling enhances the antitumor efficacy of chemotherapy in triple negative breast cancer through reduction of cancer stem cells. Cancer Lett. 2013;328(2):261–270. doi: 10.1016/j.canlet.2012.09.023. [DOI] [PubMed] [Google Scholar]
- 116.Fu Y, Chang H, Peng X, Bai Q, Yi L, Zhou Y, et al. Resveratrol inhibits breast cancer stem-like cells and induces autophagy via suppressing Wnt/beta-catenin signaling pathway. PLoS ONE. 2014;9(7):e102535. doi: 10.1371/journal.pone.0102535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Zhou M, Hou Y, Yang G, Zhang H, Tu G, Du YE, et al. LncRNA-Hh strengthen cancer stem cells generation in twist-positive breast cancer via activation of hedgehog signaling pathway. Stem Cells. 2016;34(1):55–66. doi: 10.1002/stem.2219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Fan P, Fan S, Wang H, Mao J, Shi Y, Ibrahim MM, et al. Genistein decreases the breast cancer stem-like cell population through Hedgehog pathway. Stem Cell Res Ther. 2013;4(6):146. doi: 10.1186/scrt357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Takebe N, Miele L, Harris PJ, Jeong W, Bando H, Kahn M, et al. Targeting Notch, Hedgehog, and Wnt pathways in cancer stem cells: clinical update. Nat Rev Clin Oncol. 2015;12(8):445–464. doi: 10.1038/nrclinonc.2015.61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Chin AR, Wang SE. Cytokines driving breast cancer stemness. Mol Cell Endocrinol. 2014;382(1):598–602. doi: 10.1016/j.mce.2013.03.024. [DOI] [PubMed] [Google Scholar]
- 121.Fang WB, Yao M, Brummer G, Acevedo D, Alhakamy N, Berkland C, et al. Targeted gene silencing of CCL2 inhibits triple negative breast cancer progression by blocking cancer stem cell renewal and M2 macrophage recruitment. Oncotarget. 2016;7(31):49349. doi: 10.18632/oncotarget.9885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Singh JK, Simões BM, Clarke RB, Bundred NJ. Targeting IL-8 signalling to inhibit breast cancer stem cell activity. Expert Opin Ther Targets. 2013;17(11):1235–1241. doi: 10.1517/14728222.2013.835398. [DOI] [PubMed] [Google Scholar]
- 123.Singh JK, Simões BM, Howell SJ, Farnie G, Clarke RB. Recent advances reveal IL-8 signaling as a potential key to targeting breast cancer stem cells. Breast Cancer Res. 2013;15(4):210. doi: 10.1186/bcr3436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Mannello F. Understanding breast cancer stem cell heterogeneity: time to move on to a new research paradigm. BMC Med. 2013;11(1):169. doi: 10.1186/1741-7015-11-169. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.