

Abstract
Over 50% of Western Hemisphere shorebird species are in decline due to ongoing habitat loss and degradation. In some regions of high wetland loss, shorebirds are heavily reliant on a core network of remaining human-managed wetlands during migration journeys in the spring and fall. While most refuges have been designed and managed to match the habitat needs of waterfowl, shorebirds typically require much shallower water (<10cm deep). Traditional static habitat modeling approaches at relatively coarse spatial and temporal resolution are insufficient to capture dynamic changes within this narrow water depth range. Our objectives were to: 1) develop a method to quantify shallow water habitat distributions in inland non-tidal wetlands, and 2) to assess how water management practices affect the amount of shorebird habitat in Sacramento National Wildlife Refuge Complex. We produced water depth distributions and modeled optimal habitat (<10 cm deep) within 23 managed wetlands using high-resolution topography and fixed-point water depth records. We also demonstrated that habitat availability, specifically suitable water depth ranges, can be tracked from satellite imagery and high-resolution topography. We found that wetlands with lower topographic roughness may have a higher potential to provide shorebird habitat and that strategically reducing water levels could increase habitat extent. Over 50% of the wetlands measured provided optimal habitat across <10% of their area at the peak of migration in early April, and most provided a brief duration of shallow water habitat. Reducing water volumes could increase the proportion of optimal habitat by 1–1,678% (mean = 294 %) compared to actual volumes measured at peak spring migration in 2016. For wetlands with a high habitat potential, beginning wetland drawdown earlier and extending drawdown time could dramatically improve habitat conditions at the peak of shorebird migration. Our approach can be adapted to track dynamic hydrologic changes at broader spatial scales as additional high-resolution topographic (e.g., lidar, drone imagery photogrammetry) and optical remote sensing data (e.g., Planet imagery, drone photography) become available.
Keywords: habitat modeling, topography, digital elevation model, capacitance sensor, Landsat, remote sensing, shorebird, wetland management, drought, California, Sacramento Valley, Central Valley
Introduction:
Migratory shorebird populations continue to experience population declines, despite substantial effort directed at wetland conservation and restoration programs (Scottish Natural Heritage (Agency) 2006). Shorebirds present a unique challenge for management in that they require suitable available habitat not only within their breeding and wintering grounds, but also along their spring and fall migration corridors, which must temporally-align with migration. Shorebirds’ daily energy requirements increase in preparation for spring migration (Dybala et al. 2017) and the quantity and quality of habitat available during migration can directly affect body condition, migration survival rates, and subsequent breeding season success (Brown et al. 2001). Many factors may affect the quality of available habitat for shorebirds, such as invertebrate prey density (Colwell and Landrum 1993, Farmer and Wiens 1999), vegetation type and density (Colwell 2010, Ma et al. 2010, Stutzman 2012), and the presence of predators (Zharikov et al. 2008, Stutzman 2012). However, previous work has demonstrated that habitat quantity is limited by accessibility, which is driven by a combination of water depth and bird morphology (i.e., bill and tarsus length; (Collazo et al. 2002, Strum et al. 2013).
In regions which have experienced substantial wetland loss, such as the Central Valley of California (>90% wetland loss; Central Valley Joint Venture 2006), managed wetlands and flooded agricultural fields (Taft and Elphick 2007) make up the core habitat network that shorebirds and other waterbirds rely on to rest and feed during migration, particularly under drought conditions (Schaffer-Smith et al. 2017). Wetland conservation and management in the Central Valley have historically focused on waterfowl—mainly ducks—which have very different habitat requirements from shorebirds (Safran et al. 1997, Isola et al. 2000, Taft et al. 2002, Central Valley Joint Venture 2006). In terms of habitat needs, ecologists often distinguish between diving ducks and other waterbirds, dabbling ducks, waders, and smaller shorebirds. In the Central Valley, diving waterbirds include a variety of duck species, cormorants and grebes that use water depths >25 cm, while dabbling ducks use flooded areas from 5–25 cm deep (Taft et al. 2002). Waders, such as herons and egrets, are associated with water depths of <15 cm (Taft et al. 2002). For many shorebird species, foraging is limited to water depths <10 cm deep (Safran et al. 1997, Elphick and Oring 1998, Strum et al. 2013); smaller shorebirds with short bills and short legs such as sandpipers (Calidris sp.) have the most narrow habitat requirements, using saturated wetland soils and regions flooded up to just 5 cm deep (Collazo et al. 2002, Taft et al. 2002, Ma et al. 2010). Shallow water areas are not only more accessible to smaller shorebirds, but facilitate higher foraging efficiency as they recede and concentrate prey (Neckles et al. 1990, Ma et al. 2010). Direct tracking of migratory shorebirds and bioenergetics models are providing mounting evidence that there may not be sufficient shallow water habitat in the Central Valley during spring and fall migration (Barbaree et al. 2015, Dybala et al. 2017).
There is a of lack sufficient information on wetland water depth distributions over broad areas necessary to map optimal shallow water habitat accessible to both smaller and larger shorebirds during migration. Traditional static habitat mapping approaches implemented at coarse spatial and temporal scales (e.g., Guisan and Zimmermann 2000, Elith and Leathwick 2009) do not capture dynamic hydrologic changes taking place across the wetland networks that shorebirds rely on. In recent years, substantial progress has been made in tracking surface water fluctuations around the world from satellite imagery (Alsdorf et al. 2007, Pekel et al. 2016). Although the Landsat archive offers the longest satellite record with >30 years of imagery and provides a snapshot of same location at 16-day intervals under ideal cloud-free conditions, its return interval is not well matched to relatively rapid hydrologic cycles (Alsdorf et al. 2007, Turpie et al. 2015). Landsat’s moderate 30-m spatial resolution also limits the ability to track dynamic water depth changes in smaller wetlands (Alsdorf et al. 2007).
Although water extent and flood duration information provides a proxy for habitat suitability for species that depend on aquatic habitats (Reiter et al. 2015, Schaffer-Smith et al. 2017), optical remote sensing data does not provide water depth information (Alsdorf et al. 2007, Turpie et al. 2015). Incorporating water depth information would offer a tremendous advance over considering all flooded areas as habitat or using expert opinion to derive water depth distributions (Dybala et al. 2017). In-situ water depth can be measured in a variety of ways, including visual monitoring of depth gauges (Strum et al. 2013), or the use of instruments such as water-level loggers or capacitance sensors (Larson and Runyan 2009). However, without topographic or bathymetric data it is not possible to estimate water depth beyond a fixed water depth monitoring location.
Accurate measurement of wetland water depth across wetlands requires knowledge of the water surface elevation as well as the topography of the underlying substrate; however, the relatively low vertical accuracy of existing digital elevation models (DEMs) and the dearth of topographic data for inland non-tidal wetland systems currently limits our ability to measure wetland water depth needed to resolve shorebird habitat suitability on the order of centimeters. A variety of approaches exist to generate digital elevation models, including using photogrammetry taking advantage of stereo-paired (Konecny 1986, Hirano et al. 2003) or multi-angle (Westoby et al. 2012) satellite imagery or aerial photos, terrestrial laser scanning (Brasington et al. 2012) or lidar surveys (Schmugge et al. 2002, Alsdorf et al. 2007). Topographic lidar surveys use lasers with wavelengths 800 nm-1600 nm, but most often near infrared light at 1064 nm (e.g., Liu 2008), which cannot penetrate flooded areas. However, bathymetric lidar surveys using green light at 532 nm can be conducted while wetlands are flooded to map submerged topography (e.g., (Hilldale and Raff 2008). The best available global elevation dataset has a ground resolution of 30-m and an average absolute vertical accuracy of 9 m over the continental U.S. (Tachikawa et al. 2011), while the 10-m National Elevation Dataset for the U.S. has an average absolute vertical accuracy of 1.55 m (U.S. Geological Survey 2015). Unfortunately, both of these DEM products exclude waterbodies and flooded surfaces, considering them as flat regions with a single elevation value. This problem also affects DEMs that rely on topographic lidar (Alsdorf et al. 2007). For example, California’s Central Valley was recently mapped with aerial topographic lidar surveys, however the derived DEM does not include wetland topography due to the presence of active flooding at the time of the surveys (California Department of Water Resources 2015).
Topographic surveys using differential GPS (Los Huertos and Smith 2013) and precise field measurements of water depth can be used to model water depth spatially at a fine-scale appropriate for tracking the presence and duration of suitable flooded habitat for shorebirds within individual wetlands. The integration of satellite-derived water extent information with mapped bathymetry can provide additional information regarding the presence and frequency of flooding within specific water depth ranges over long time scales (Rowe et al. 2002, Munyaneza et al. 2009). Tracking water depth distributions at finer spatial and temporal resolution, and understanding the habitat patterns that result from current management practices can assist in more targeted strategies to optimize habitat across wetlands managed by state and federal fish and wildlife agencies as well as private conservation areas.
Our objectives were: 1) to model fine-scale water depth and habitat suitability at high temporal frequency using high-resolution topography, in-situ water depth measurements and satellite imagery, and 2) to assess the extent and duration of optimal shorebird habitat provided under current management practices in Sacramento National Wildlife Refuge (NWR) Complex managed wetlands, thereby helping to inform wetland restoration and water management planning efforts throughout the Sacramento Valley. Managed wetlands at Sacramento NWR Complex provide the most reliable wetland habitat within the Sacramento Valley (Schaffer-Smith et al. 2017), a globally important stopover site for migratory shorebirds during the spring migration season.
Methods
We estimated water depth and optimal habitat distributions for 23 managed wetlands during spring shorebird migration using a combination of high-resolution topography surveys and water depth measurements. Secondarily, we tracked optimal habitat availability during the peak of spring migration from a satellite-derived historical record of surface water extent (Schaffer-Smith et al. 2017). We used ArcGIS for processing of topography data (ESRI 2014) and R for all data analysis (R Core Team 2012).
Study Area
Sacramento NWR Complex is located within the north Central Valley of California (Fig. 1), a region that supports hundreds of thousands of shorebirds during spring migration between mid-March and mid-April (Shuford et al. 1998, Central Valley Joint Venture 2006). Although 90% of Central Valley wetlands have been converted mainly to agricultural uses (Frayer et al. 1989), the rice and wetland matrix of the Sacramento Valley (northern portion of the Central Valley) is recognized by the Western Hemisphere Shorebird Reserve Network as a site of international importance (Western Hemisphere Shorebird Reserve Network 2009). A network of managed wetlands remains in the Sacramento Valley, in addition to flood-irrigated agricultural fields which support 20% of U.S. rice production (Central Valley Joint Venture 2006, Strum et al. 2013) and provide important surrogate habitat for shorebirds (Elphick 2000, 2010, Barbaree et al. 2015). Sacramento NWR Complex contains approximately 25% of the total emergent non-tidal wetland area in the Sacramento Valley mapped by the National Wetland Inventory (U.S. Fish and Wildlife Service 2016a, 2016b).
Figure 1.
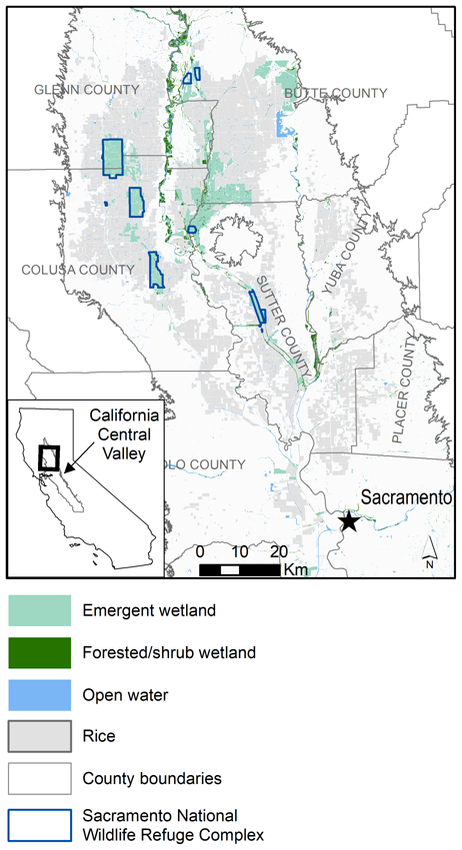
Landcover and land use in the Sacramento Valley of California, USA. Sacramento National Wildlife Refuge Complex boundaries are from U.S. Fish and Wildlife Service (2016b); wetland distributions are from U.S. Fish and Wildlife Service (2016a), and rice production areas are from USDA-NASS (2014).
The majority of managed wetlands in the Sacramento Valley are intentionally flooded from fall through spring to support a variety of wetland dependent plant and wildlife species—this deviates somewhat from historical patterns in which wetlands were fed principally by snow melt runoff from the Sierra Nevada (Wilson and Cronon 2010). The water used in the Sacramento Valley during spring is primarily sourced from reservoirs that capture snowmelt runoff, which is then delivered to wetlands and agricultural fields hundreds of kilometers away through an extensive network of canals, pump stations and levees. Although the Sacramento Valley receives an annual average precipitation of 890 mm, mostly in late fall and winter (NOAA National Centers for Environmental Information 2017), this is highly variable depending on El Niño and other climate cycles. Under strong El Niño conditions in 1998, 1,488 mm of precipitation were recorded, while only 541 mm were recorded for 2014, the third driest year on record in the state (NOAA National Centers for Environmental Information 2017). The extent of emergent wetland water at the peak of spring migration in the Sacramento Valley can fluctuate dramatically, with a 70–80% reduction in water extent within emergent wetlands detected during extremely dry years as compared to extremely wet years (Schaffer-Smith et al. 2017). Ultimately water allocations in the Central Valley are highly managed and more closely tied with annual snowpack and run-off than with local precipitation.
Site selection
For this study, we selected 23 managed wetlands across Sacramento NWR, Delevan NWR, and Colusa NWR (Fig. 2) based on random sampling, stratified by quantiles for wetland size (small, medium, large) and historical shorebird densities (low, medium, high; based on twice-monthly surveys conducted at Sacramento NWR Complex). We included wetlands managed as seasonally-flooded marsh (which receives 70% of shorebird use at Sacramento NWR Complex), semi-permanent, and permanent ponds. Severe drought conditions were in effect when we initiated field data collection in 2015; we therefore limited site selection to those wetlands which would receive water even if the water allocation for Sacramento NWR Complex were to be cut to 40% of the normal amount. Additionally, we worked with managers to choose units without planned maintenance conflicts. Due to these limitations, it was necessary to include three semi-permanent/permanent pond sites. The 23 selected wetlands represent approximately 6% of the total area of Sacramento NWR Complex.
Figure 2.
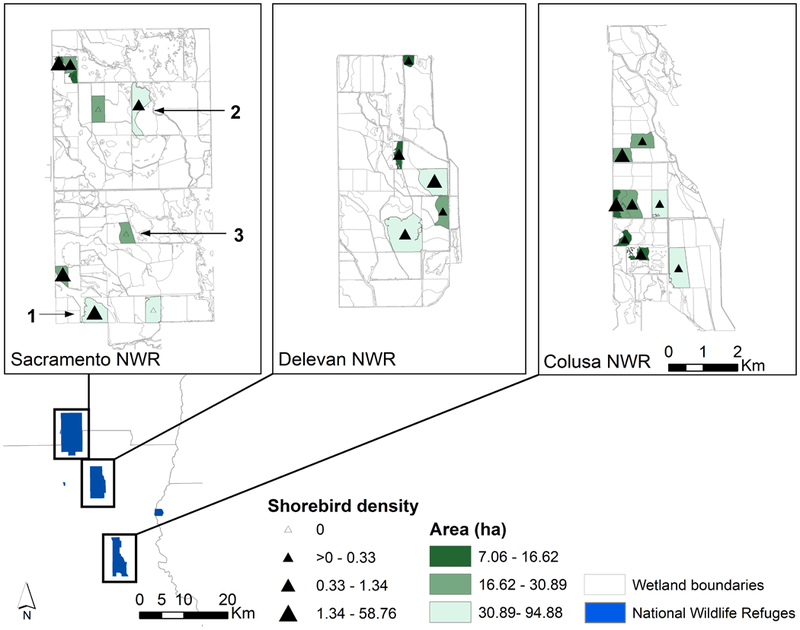
Monitored wetlands in Sacramento National Wildlife Refuge Complex within Sacramento, Delevan, and Colusa National Wildlife Refuges. Study sites were selected using a stratified random sample to capture the range of shorebird densities observed from wildlife surveys (1997–2014) and a range of wetland size (U.S. Fish and Wildlife Service 2016b). Numbered wetlands 1, 2, and 3 are discussed in greater detail in Figs. 3, 6, 7.
Water depth monitoring
Within each wetland, we deployed a 1.5 m Odyssey capacitive water depth sensor (Dataflow Systems, Ltd.). Capacitive sensors record water depth based on the linear relationship between variation in capacitance and the height variation of the water in contact with the Teflon sensor element (Larson and Runyan 2009). Each sensor was contained within a perforated 37.5mm width schedule 40 PVC shroud and mounted on a t-post at a deep point near the wetland flow outlet structure. This positioning ensured that water depth records would track changes during the wetland drawdown in mid-late spring. We recorded water depth (+/−5mm) every 4 hours at each sensor and recalibrated each sensor periodically to ensure accurate water depth readings.
High-resolution topography mapping
We completed a dry season real-time kinematic GPS survey for each wetland to document wetland topography at a high spatial resolution suitable for modeling water depth in centimeters. We used two Trimble AgGPS 442 units to complete surveys; the base GPS unit remained stationary at a reference location established along the wetland perimeter, while the mobile GPS unit automatically sampled elevations with a vertical accuracy of 1–3 cm along the path of an all-terrain vehicle. We surveyed each wetland with a maximum transect width of 5 m. The entirety of wetland levee perimeters were surveyed, and additional transects were covered along the slopes below levees, given that the greatest topographic variation typically occurs at wetland edges. As needed, additional points were collected to capture notable topographic features present within wetland interiors, including islands and swales. For each wetland, we established referenced elevation points at the top and bottom of a cement water control structure, and also recorded the position and base elevation of the water depth sensor.
To create digital elevation models, we first adjusted the relative elevation point clouds we collected in the field to the wetland levee boundary elevations mapped by the California Department of Water Resources’ (DWR) lidar-based DEM (California Department of Water Resources 2015). Some data cleaning was required, including removal of points with poor data quality codes recorded by the GPS. We also removed potentially spurious points using DWR aerial photos collected concurrently with the DWR DEM; points which were inundated in aerial photos, yet were recorded with elevation higher than levees, were removed from the dataset. To generate a 5-m resolution DEM for each wetland, a guided inverse distance weighted interpolation was applied to the final point clouds (average point spacing = 4.16 m), constrained by wetland boundary lines (U.S. Fish and Wildlife Service 2016b).
Daily spring water depth and optimal habitat modeling
We modeled daily water depth across each wetland and the volume of water contained in each wetland using the DEMs and water depth measurements that we collected from February 1 through May 31, 2016 (e.g., Fig. 3). Raw water depth measurements collected at 4-hr intervals were aggregated to a daily mean water depth measurement; we assumed a uniform water surface height across the wetland based on this measurement and the known elevation at the base of the sensor. To estimate daily water depth distributions across the entirety of each wetland, we computed the difference between the water surface and the ground surface elevations from the DEM. Regions mapped up to 10 cm depth were considered to represent the extent of optimal habitat for each day. While a range of depths could be modeled for particular species and habitat types, we chose to use 10 cm given that larger shorebird species are capable of exploiting shallower water depths, while smaller shorebirds would be excluded from deeper water (Elphick and Oring 2003). For each wetland, this data processing resulted in 119 daily water surfaces, water depth maps, and maps of optimal habitat extent in spring 2016.
Figure 3.
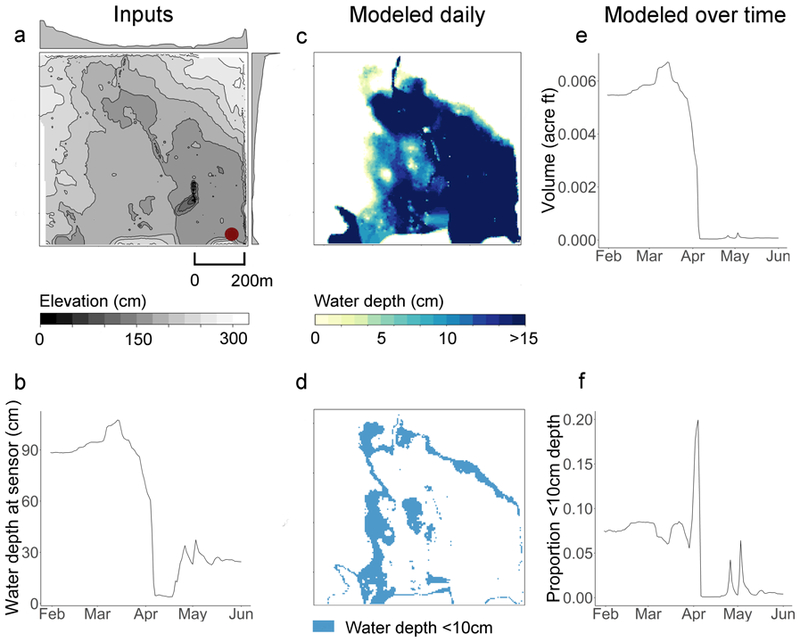
Example of using (a) high-resolution elevation and (b) water depth measurements collected by a sensor at a fixed location (a; red dot) to infer (c, d) daily spatial water depth and optimal habitat distributions (1 April 2016 data shown), and to estimate (f) longitudinal measures of the volume of water and (e) the proportion of optimal habitat in Wetland 1 during spring 2016. Elevation isolines in panel a indicate 25-cm increments and marginal histograms along the figure axes indicate average row and column values. 1 acre foot = 1.233 9 106 L.
Assessing potential for shallow water habitat gains through water management
For each of the 23 wetlands, we determined the water volume that maximized the proportion of optimal habitat available for shorebirds from in-situ water depth records and DEMs (Appendix S1: Table S1). We calculated the increase in the proportion of optimal habitat that could be achieved by managing wetlands at these idealized volumes at the peak of migration as compared to the actual volumes detected at peak migration on April 1, 2016. To explore the influence of topography on potential habitat, we conducted a linear regression predicting maximum habitat proportion using the planimetric-to-surface area ratio (2D:3D area ratio), which is an index of topographic roughness used in habitat assessment applications (Jenness 2004). We also applied linear regression to assess the association between maximum habitat proportion and the idealized water volumes across wetlands.
To illustrate how wetland water level manipulation and topographic characteristics affect habitat suitability spatially, we simulated drawdowns for three wetlands (Wetlands 1, 2, and 3) representing a range of topographic complexity. These three wetlands were selected based on their slope variability, calculated as the difference between the maximum and minimum slope values within a 25 m moving window (Ruszkiczay-Rüdiger et al. 2009). We evaluated drawing down the relative volume of water in each wetland from 100%, down to 75%, 50% or 40% volume. We considered 100% volume to be the maximum volume captured by our spring 2016 water depth data. We did not simulate volumes below 40%, as maintaining wetlands at or below this level is impractical, particularly under warm and dry conditions.
Modeling historical peak migration habitat over drought cycles using satellite imagery
To determine the typical peak of shorebird migration, we analyzed historical bi-weekly census data collected at Sacramento NWR Complex from 1996–2015 from 382 unique survey areas. We summarized the average number of shorebirds for each spring day from February 1 through May 31st across all surveyed wetlands and all years of available spring census data. We then applied local polynomial regression fitting (LOESS; Cleveland et al. 1992) to identify the point at which migration generally peaked.
Using historical Landsat satellite imagery and our high-resolution wetland DEMs, we then examined water distributions and the distribution of optimal habitat within mapped wetlands at the peak of spring migration (early April) from 1996–2015. Although we had previously mapped water, non-water and cloudy (no data) regions across the Sacramento Valley during spring using all available Landsat surface reflectance imagery (Schaffer-Smith et al. 2017), we could only verify that wetland boundaries and topography had been consistent since 1996. We retained Landsat water distribution maps that represented a cloud-free view of at least 90% of the area of each wetland based on the LEDAPS or LaSRC cloud identification algorithms (Zhu and Woodcock 2012, Vermote et al. 2016). Where multiple water extent maps were available for the period from mid-March to mid-April in the same year, we retained the map containing a higher proportion of cloud-free pixels. Each peak migration water distribution map was downscaled to match the 5-m pixels of each wetland DEM, using nearest-neighbor resampling. We then identified the highest elevation cell where water was detected for that year and generated a corresponding water surface height raster. We lastly generated a water depth map by taking the difference between the water surface height and the DEM, and used the 10-cm depth threshold to map the extent of optimal habitat at peak migration for each year.
For the wetlands with historical peak migration habitat information, we examined whether the proportion of optimal habitat observed was related to water availability, reasoning that more habitat would be available in wetter years. We used linear regression to determine whether an index of water availability in the Sacramento River Region (California Department of Water Resources 2016) was a significant predictor of the proportion of optimal habitat observed over time. We also examined trends in drought vs. non-drought years using a t-test to compare the mean proportions of habitat observed across wetlands. We considered the California Department of Water Resources’ designated ‘Critically Dry’, ‘Dry’, and ‘Below Normal’ water years to represent drought conditions, and ‘Above Normal’ or ‘Wet’ years to represent non-drought conditions (California Department of Water Resources 2016).
Results
Habitat suitability patterns during spring shorebird migration
Shorebird abundance during spring in the Sacramento Valley has historically peaked in early to mid-April, yet for most of the wetlands we monitored in spring 2016, the maximum proportion of optimal shallow habitat was slightly delayed relative to bird abundance, with the peak most often occurring in mid-April (Fig. 4). Of the 23 wetlands that we monitored, 13 had only a small proportion (<10%) of flooded habitat within the optimal depth range during the peak of migration between mid-March and mid-April (Fig. 4). Only 7 wetlands provided optimal habitat within >10% of their area for a sustained period of at least 10 days, and only 2 of those provided 30 consecutive days spanning March 15 - April 15. The maximum proportion of optimal habitat observed in one wetland during spring 2016 was 50%, although this occurred in early May, after the peak of migration.
Figure 4.
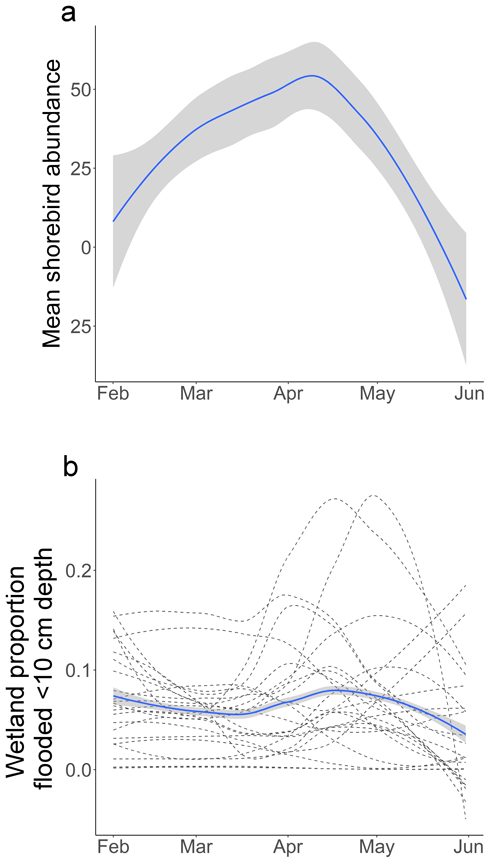
Temporal mismatch between (a) peak shorebird migration and (b) the timing of shallow water habitat availability in monitored wetlands. The peak of migration was estimated using historical once every two weeks shorebird census data collected at Sacramento National Wildlife Refuge Complex 1997–2015 (U.S. Fish and Wildlife Service 2016b), averaged across all surveyed wetlands in all years. Locally weighted smoothing (LOESS) was applied to both shorebird census data and optimal habitat proportions (blue lines with standard error shown in gray). The proportions of optimal habitat in individual wetlands in spring 2016 are represented as dashed lines.
Simulated water management to maximize the proportion of optimal shorebird habitat
We estimated that altering wetland water levels could increase the extent of optimal habitat at peak migration by 1–1,678% (mean = 294%, median = 88%), when compared to actual managed levels captured by water depth records on April 1, 2016 across the 23 wetlands (Appendix S1: Table S1). The wetland with the greatest potential increase in habitat extent is a large seasonally flooded marsh, with an optimal relative volume of 47%, yet was managed at 98% volume on April 1, 2016 (Appendix S1: Table S1). The water volume which optimized habitat was not consistent across the 23 wetlands, but ranged from 16% - 100% of the maximum (mean = 54%, median 48%, Appendix S1: Table S1). Two of the wetlands we monitored had optimal volumes of ~100%, but even under this optimal water management these sites each provided optimal habitat across <2% of their respective areas (Appendix S1: Table S1). For the wetlands that we measured, linear regression results indicate that those with higher mean planimetric-surface area ratios (lower topographic roughness) had more potential to provide optimal habitat (Fig. 5a; R2 = 0.25 p = 0.007). We also found that wetlands with a higher potential to provide habitat would ideally be managed with lower water volumes (Fig. 5b; R2 = 0.58 p < 0.001).
Figure 5.
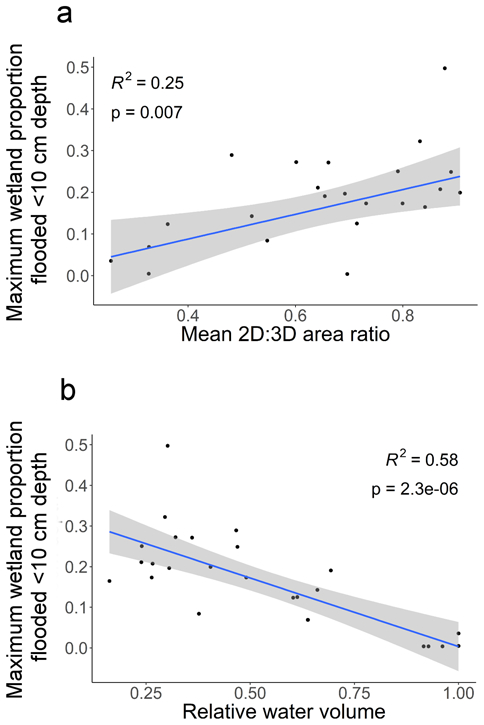
Association between the maximum proportion of optimal shorebird habitat (<10 cm water depth) observed across 23 wetlands and (a) wetland topography and (b) optimal water volume. A higher potential proportion of habitat was observed in wetlands with higher planimetric-to-surface area ratio (2D:3D area ratio). Idealized management conditions for wetlands with higher potential habitat proportions occurred at lower relative water volumes than for wetlands with low potential habitat proportions.
Simulated drawdowns of three representative wetlands with a range of topographic variation (Wetlands 1, 2, 3; Fig. 2; Fig. 6) demonstrated the variability of interactions between topography and water management across wetlands. Wetland 1 represented relatively low slope variability (mean =1.67, sd = 2.03), Wetland 2 represented a mid-range value (mean =4.85, sd=7.93), and Wetland 3 represented a high value (mean =11.43, sd = 12.12). For all three wetlands, reductions in the amount of water from the maximum volume increased the proportion of optimal habitat provided (Fig. 5); Wetland 1 provided a maximum optimal habitat extent across 20% of its area at 40% volume, while Wetland 2 had a maximum optimal habitat across 12.5% of its area at 50% volume, and Wetland 3 provided a maximum optimal habitat across 27% of its area at 38% volume.
Figure 6.
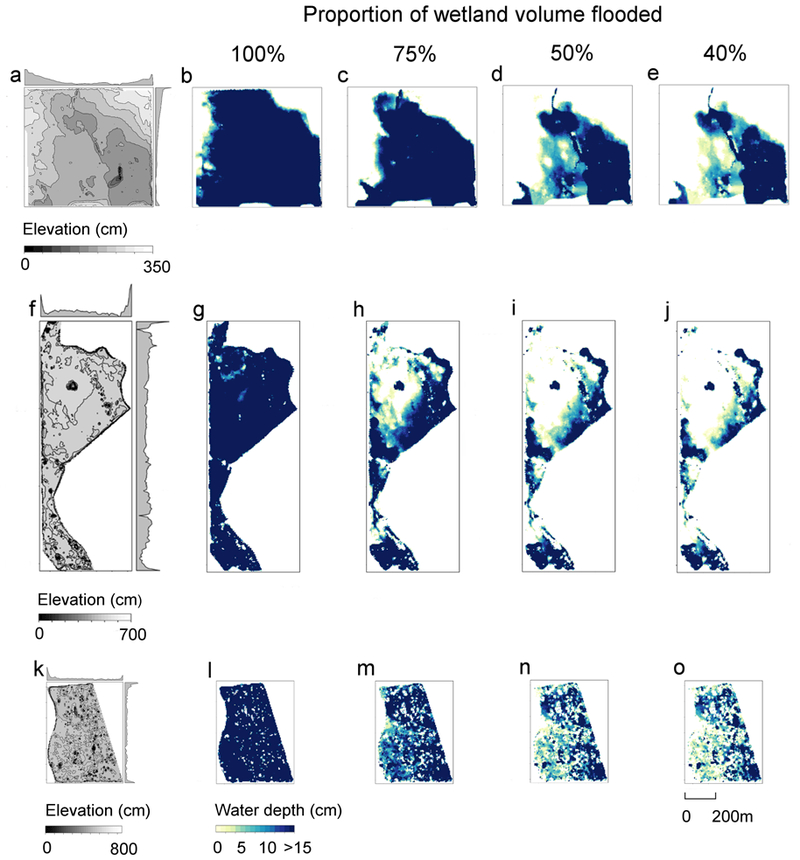
Implications of water level management and topographic variation for optimal shorebird habitat distributions. Wetland drawdowns were simulated for Wetlands 1 (top row, low slope variability), 2 (middle row, medium slope variability), and 3 (bottom row, high slope variability) Relativized elevations for each wetland are shown in the first column (a, f, k), with isolines indicating 25-cm elevation increments and marginal histograms indicating average row and column summaries. Maximum volumes (100%; b, g, l) were calculated based on the maximum water depth reading at a fixed-location capacitance water depth sensor deployed in each wetland, and the difference between water surface height and the digital elevation model for each wetland (e.g., Fig. 3). Drawdowns to 75% (c, h, m), 50% (d, i, n), and 40% (e, j, o) of the maximum volume were then simulated for each wetland.
Historical peak migration habitat distributions over drought cycles
Of the 23 wetlands that we monitored, historical peak migration water extent information from cloud-free Landsat was available for 15 locations. For those 15 wetlands, 12–14 years of water extent data were available (mean = 13.5 years). The average proportion of optimal habitat at peak migration varied from just 0.003% to 6% (mean = 1.58%) across wetlands (Appendix 1: Fig. S1). The frequency of optimal habitat tended to be higher in lower flood frequency portions of wetlands. For example, 51% of Wetland 1’s areal extent was flooded at the peak of migration in >50% of the 14 years of available imagery (Fig. 7a); but only 18.5% of this flooded habitat could be classified as optimal habitat in >10% of years. (Fig. 7b). In contrast, the less reliably flooded zone of Wetland 1 provided optimal habitat in up to 70% of years (Fig.7b). There was no apparent association between drought and optimal habitat proportions in Wetland 1 at the peak of migration from 1996–2015 (Fig. 7c).
Figure 7.
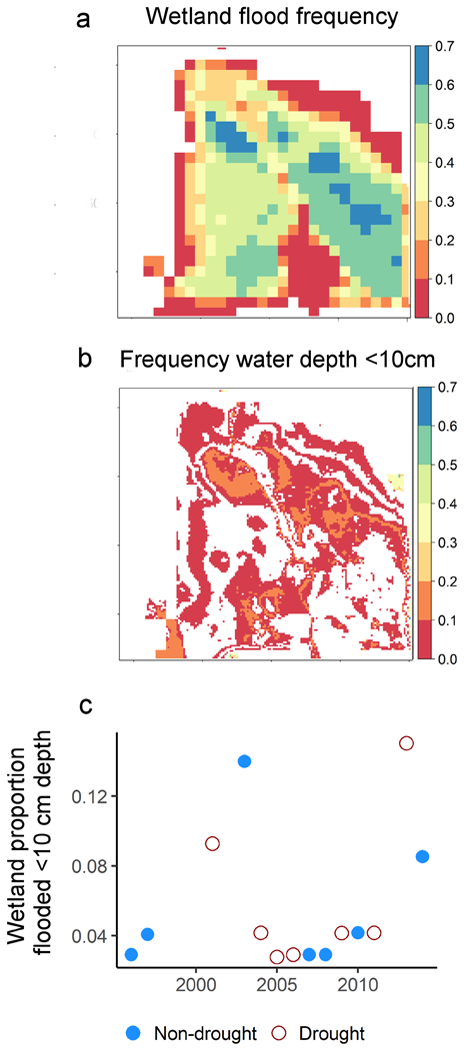
Example of using satellite-based water extent mapping and high-resolution topography data to model optimal habitat distributions in Wetland 1 at the peak of spring migration (early April) from 1996 to 2015. (a) Open water extent and water frequency were mapped during peak migration at Wetland 1 from 1996 to 2015 using Landsat satellite imagery (Schaffer-Smith et al. 2017). (b) We mapped optimal shorebird habitat frequencies by integrating topographic data (water depth <10 cm). Frequencies were computed as the proportion of times that water or habitat was detected out of the total number of cloud-free views of that pixel available over time. (c) The proportion of Wetland 1 that provided optimal habitat from 1996 to 2015 was not associated with drought cycles, based on an index of water availability in the Sacramento River Basin (California Department of Water Resources 2016).
Across all monitored wetlands, we found no relationship between the observed proportion of optimal habitat and drought cycles over time (Appendix S1: Figure S1). Relatively high and low proportions of optimal habitat occurred in both drought and non-drought years from 1996–2015 (Appendix S1: Figure S1). Linear regression results indicated that the index of total water availability in the Sacramento River Basin was not a significant predictor of the detected proportion of optimal habitat in wetlands (R2 = 0.002, p =0.22). A two-sample t-test also revealed no difference between the average proportion of optimal habitat provided across wetlands under drought or non-drought conditions (p=0.48, eight drought years, seven non-drought years).
Discussion
We found that there appears to be even less flooded habitat in the suitable depth range for shorebirds than has been previously estimated. As part of a recent conservation evaluation for Central Valley shorebirds using bioenergetics modeling (Dybala et al. 2017) experts estimated that 75–90% of seasonal wetlands provide shallow water (<10 cm) during the month of April. However, the highest proportional coverage of optimal habitat we observed during peak migration in 2016 was 33%, lasting a duration of 8 days, while 13 of the 23 wetlands (56%) we monitored never had optimal habitat covering greater than 10% of their area. Our analysis also reveals that only a small portion of surface water detected from satellite imagery is likely to be within the optimal depth range for shorebirds. Although previous research indicated that the extent of flooded habitat in the Sacramento Valley is sensitive to climate (Schaffer-Smith et al. 2017), we found no association between drought and the proportion of optimal shorebird habitat at the peak of migration from 1996–2015. This indicates the extent to which human management decisions regarding water allocations and wetland management plans can determine habitat extent, rather than annual weather variation.
The Central Valley Joint Venture Implementation Plan (Central Valley Joint Venture 2006) states that wetlands should provide 100% of shorebird energy needs from April through May, however, our results and those of Dybala et al. (2017) raise concerns that this goal may not be achievable using current management practices. Dybala et al. (2017) recommended increasing shallow water habitat (<10 cm depth) in the Central Valley by an additional 4,692 ha (11,594 ac) during mid-March through late April over the next 10 years. Given that usable habitat may have been overestimated in previous studies, there may be an even greater need for habitat, and thus there is an urgent need to prioritize shallow flooded habitat in future conservation and restoration plans.
Targeted management of wetland water levels may be a particularly important strategy to address the mismatch in the timing of migration and optimal shorebird habitat availability, and to help shorebird populations to survive future droughts. Refuges have received a guaranteed water supply since the Central Valley Project Improvement Act was enacted in 1992, however allocations are subject to reductions during ‘Critically Dry’ years and the water policy landscape could change in the future. Sacramento National Wildlife Refuge Complex received 75% of its full allocation in 2015, resulting in the refuge’s decision to flood fewer wetlands, delay scheduled wetland flooding, reduce total inundation time (earlier drawdowns), and reduce the volume of fresh water flow-through during the inundation period. Modifying drawdown schedules for high habitat potential wetlands to occur earlier in the spring season, and slowing or extending drawdowns, could substantially increase the proportion of optimal habitat that managed wetlands provide during peak migration. Given that idealized water volumes varied substantially across wetlands, we suggest that it is important to manage them each individually to optimize water use and habitat availability.
Practical implementation of wetland water management changes must take into account water availability, local weather, wildlife needs, vegetation management, and maintenance projects. Fortunately, most migratory waterfowl have left Sacramento NWR Complex for their breeding grounds by early April such that modifying water levels and reducing deeper water habitat would not be expected to adversely affect these populations. Although shallower water depths typically boost invertebrate productivity (Gray et al. 2013), reducing total landscape flood extent could have implications for populations of aquatic invertebrates, copepods, crayfish and other species that provide food for resident wildlife. Staggering drawdowns and managing wetlands as a mosaic with a variety of hydroperiods—as is already practiced at Sacramento NWR Complex—would help minimize adverse impacts to resident species.
There are potential conflicts between changing wetland water levels and managing vegetation. Irrigation is a key tool used by wetland managers to either promote the growth of desirable vegetation (e.g., high-yield seed plants that provide food for wildlife) or to prevent the spread of weeds (Mensik and Reid 1995). The timing of spring wetland drawdowns is a major determinant of which plant species will germinate in seasonal wetlands (Strong et al. 1990). At Sacramento NWR Complex, early drawdowns tend to favor cool season emergents, while late drawdowns may result in poor germination and survival of important food plants for wildlife (U.S. Fish and Wildlife Service 2009). Wetland managers must consider how target species’ biological requirements vary throughout the year and consider vegetation management and other logistical constraints when making water level management decisions.
Conservation objectives for shorebirds could also be achieved through other complimentary strategies. New wetland restoration projects could be designed to maximize the amount of shallow water habitat provided. Relatively flat rice fields could be also be shallowly flooded to provide significant additional habitat in lieu of restoration projects, which often involve costly earth-moving operations (Elphick and Oring 1998, Strum et al. 2013). Flooded rice fields provide abundant and accessible food resources, and may be especially attractive to shorebirds due to reduced predation risk compared to semi-natural wetland systems (Elphick and Oring 1998, Barbaree et al. 2015). Recently the highest shorebird densities recorded for agricultural land in the Sacramento Valley were documented in temporary shallow water habitat provided in rice fields through the ‘Bird Returns’ payment for services program (Golet et al. 2017). Such dynamic conservation strategies (Reynolds et al. 2017) could be effective for providing supplemental habitat, so long as they do not conflict with other considerations. For example, Bird Returns fall migration contracts were limited to two week periods due to weed management and mosquito control concerns (Golet et al. 2017). Transferability of such programs to other wetland regions would depend on local conditions, including the availability and cost of water, agricultural practices and the availability of funds to incentivize private landowner participation.
Our estimates of water depth, optimal habitat, and water volume are subject to limitations. There was some error associated with the DEMs that we generated. It was not possible to survey topography where impenetrable wetland vegetation was present (i.e., bulrush). We additionally did not survey areas where an accumulation of tall and dry vegetation was present, due to fire safety concerns. Furthermore, by using water depth to define optimal habitat we likely underestimated the actual extent of suitable areas; our analysis did not capture saturated soils which typically occur beyond the waterline, and represent important shorebird habitat. Although we only measured above-ground water volume and did not account for infiltration or evaporation processes, we do not think this affected comparison among years.
With increased availability of high-resolution DEMs, our approach could lead to better understanding of hydrologic regimes and water depth distributions at a fine spatial and temporal scale. If available, even finer resolution topography (e.g., centimeter scale) would be helpful for resolving water depth distributions across wetlands with low topographic variation. Advancements in lower-cost lidar, drone-based imagery and high-resolution satellite imagery (e.g., <4-m Planet imagery) coupled with new low-cost or open source tools for DEM generation (e.g., Pix4D, GRASS GIS, Boise Center Aerospace Laboratory [BCAL] Tools) offer exciting possibilities for expanding the global coverage of high-resolution DEM data more easily and affordably than before (Sona et al. 2014).
Once DEM data are available for wetlands, they can be combined with in-situ water depth monitoring or water extent from satellite or aerial photo imagery to track water depth over time. More frequent return high-resolution satellite imagery (e.g., near daily Planet satellite constellation imagery) is making it possible to dynamically track water extent with greater precision than ever before. In-situ depth gauges would provide a lower cost alternative to the capacitance water depth sensors used in our study (Strum et al. 2013), provided that the base elevations at gauges are known and that they are visually monitored with sufficient frequency to capture flooding and drying cycles.
To our knowledge this is the first empirical study using water depth sensor technology and high-resolution topography to track availability of suitable shallow water habitat for migratory shorebirds. Quantifying shallow water habitat more accurately with this approach may aid in better understanding the complexities of shorebird habitat selection, which depends on a suite of additional factors such as site latitude, species-specific migration timing, day length, and weather patterns (Colwell 2010). The information we have generated can also be used to guide management of existing wetlands and to inform future wetland habitat restoration projects. Our approach is transferrable to other wetland systems globally, leveraged by the rapid increase in remotely sensed data in the form of bathymetric lidar surveys or multi-angular imagery collected during the dry season. Although we evaluated habitat for shorebirds using a threshold of water depths <10 cm, the determination of habitat suitability can be easily adjusted for other applications according to the requirements of the target species or group of interest.
Conclusions
Wetlands have been widely eliminated and degraded throughout the world, and many remaining wetlands are at risk. Avoiding further loss or modification of wetlands will be crucial for maintaining the important wildlife habitat and other ecosystem services benefits that wetland ecosystems provide. Under ongoing climate change and continued human population growth, it will be ever more important to manage remaining wetlands optimally to meet multiple objectives.
For the managed wetlands that we measured in Sacramento NWR Complex, the extent of optimal shallow water shorebird habitat peaked approximately two weeks after the peak abundance of shorebirds in early April. At peak migration, the wetlands that we measured provided water depths <10 cm within less than 33% of their area, which is substantially lower than previous estimates of habitat availability in managed wetlands across the Central Valley. Although our study was limited to 23 diverse wetlands, there is some evidence that wetlands with lower topographic roughness (i.e., flatter, with more gradually sloped edges) have a higher potential to provide shallow water habitat. It may be possible to boost the amount of optimal shorebird habitat in managed wetlands by altering the drawdown schedules for high habitat potential wetlands to better coincide with the timing of peak spring migration, or to provide additional habitat during droughts.
The methods that we have illustrated in this study using high-resolution topography, in-situ water depth measurements, and satellite-based water extent maps, can be applied to more accurately track hydrologic processes in inland aquatic systems with the proliferation of available high-resolution DEMs. Topography and remote sensing-based water depth time series could contribute greatly to vegetation community mapping and predicting the distributions of plant and wildlife species that depend on specific aquatic habitats and unique hydrologic regimes. Understanding these dynamic fine-scale hydrologic processes over broader scales can inform the acquisition of conservation properties and wetland restoration planning. As semi-arid landscapes such as the Sacramento Valley face changing precipitation regimes, data like those generated here will enable tracking of wetland water depths and volumes, highlight water use optimization opportunities, and help to justify the allocation of limited water resources.
Supplementary Material
Acknowledgements
DSS conducted fieldwork, data processing, and analysis. All authors contributed to study design and manuscript preparation. DSS was supported by a NASA Earth and Space Science Fellowship (NNX13AQ15H) an NSF Doctoral Dissertation Research Improvement Grant (1459226), and a P.E.O. Scholar Award. Sacramento NWR Complex provided housing and access to equipment for elevation surveys, and Jane and Harvey Swenson generously provided a vehicle for use during fieldwork. We thank Todd Smith and the Duke Phytotron for assistance with the design and construction of sensor housings. We thank Sacramento NWR Complex biologists and irrigators for input on site selection and placement of sensors. Scott Currier and Jacob Byers provided valuable technical support during elevation surveys. This research would not have been possible without the support of Mike Wolder, who is greatly missed.
Literature Cited
- Alsdorf DE, Rodríguez E, and Lettenmaier DP. 2007. Measuring surface water from space. Reviews of Geophysics 45. [Google Scholar]
- Barbaree BA, Reiter ME, Hickey CM, and Page GW. 2015. Day and night habitat associations of wintering dunlin (Calidris alpina) within an agriculture-wetland mosaic. Waterbirds 38:40–46. [Google Scholar]
- Brasington J, Vericat D, and Rychkov I. 2012. Modeling river bed morphology, roughness, and surface sedimentology using high resolution terrestrial laser scanning. Water Resources Research 48. [Google Scholar]
- Brown S, Hickey CM, Harrington B, and Gill R. 2001. U.S. Shorebird Conservation Plan, second edition Manomet Center for Conservation Sciences. [Google Scholar]
- California Department of Water Resources. 2015. Central Valley Flood Evaluation Project Topographic Lidar and Orthophoto Dataset. California Department of Water Resources, Sacramento, California. [Google Scholar]
- California Department of Water Resources. 2016. Chronological Reconstructed Sacramento and San Joaquin Valley Water Year Hydrologic Classification Indices. California Data Exchange Center. [Google Scholar]
- Central Valley Joint Venture. 2006. Central Valley Joint Venture Implementation Plan - Conserving Bird Habitat. U.S. Fish and Wildlife Service, Sacramento, CA. [Google Scholar]
- Cleveland WS, Grosse E, and Shyu WM. 1992. Local regression models Page: in Chambers JM and Hastie TJ, editors. Statistical Models in S. [Google Scholar]
- Collazo JA, O’Harra DA, and Kelly CA. 2002. Accessible habitat for shorebirds: factors influencing its availability and conservation implications. The International Journal of Waterbird Biology 25:13–24. [Google Scholar]
- Colwell MA 2010. Shorebird ecology, conservation, and management. University of California Press, Berkeley. [Google Scholar]
- Colwell MA, and Landrum SL. 1993. Nonrandom shorebird distribution and fine-scale variation in prey abundance. The Condor 95:94–103. [Google Scholar]
- Dybala K, Reiter M, Hickey C, Strum K, and Yarris G. 2017. A bioenergetics approach to setting conservation objectives for non-breeding shorebirds in California’s Central Valley. San Francisco Estuary and Watershed Science 15. [Google Scholar]
- Elith J, and Leathwick JR. 2009. Species distribution models: Ecological explanation and prediction across space and time. Annual Review of Ecology, Evolution, and Systematics 40:677–697. [Google Scholar]
- Elphick CS 2000. Functional equivalency between rice fields and seminatural wetland habitats. Conservation Biology 14:181–191. [Google Scholar]
- Elphick CS 2010. Why study birds in rice fields? Waterbirds 33:1–7. [Google Scholar]
- Elphick CS, and Oring LW. 1998. Winter management of Californian rice fields for waterbirds. Journal of Applied Ecology 35:95–108. [Google Scholar]
- Elphick CS, and Oring LW. 2003. Conservation implications of flooding rice fields on winter waterbird communities. Agriculture, Ecosystems & Environment 94:17–29. [Google Scholar]
- ESRI. 2014. ArcGIS Desktop, version 10.3. Environmental Systems Research Institute, Redlands, CA. [Google Scholar]
- Farmer AH, and Wiens JA. 1999. Models and reality: Time-energy trade-offs in pectoral sandpiper (Calidris melanotos) migration. Ecology 80:2566–2580. [Google Scholar]
- Frayer WE, Peters DD, and Pywell HR. 1989. Wetlands of the California Central Valley: Status and trends, 1939 to mid-1980’s. 28pp U.S. Fish and Wildlife Service, Portland, Oregon. [Google Scholar]
- Golet GH, Low C, Avery S, Andrews K, McColl CJ, Laney R, and Reynolds MD. 2017. Using ricelands to provide temporary shorebird habitat during migration. Ecological Applications. [DOI] [PubMed]
- Gray MJ, Hagy HM, Nyman JA, and Stafford JD. 2013. Management of Wetlands for Wildlife Pages 121–180 in Anderson JT and Davis CA, editors.Wetland Techniques. Springer; Netherlands, Dordrecht. [Google Scholar]
- Guisan A, and Zimmermann NE. 2000. Predictive habitat distribution models in ecology. Ecological Modelling 135:147–186. [Google Scholar]
- Hilldale RC, and Raff D. 2008. Assessing the ability of airborne LiDAR to map river bathymetry. Earth Surface Processes and Landforms 33:773–783. [Google Scholar]
- Hirano A, Welch R, and Lang H. 2003. Mapping from ASTER stereo image data: DEM validation and accuracy assessment. ISPRS Journal of Photogrammetry and Remote Sensing 57:356–370. [Google Scholar]
- Isola CR, Colwell MA, Taft OW, and Safran RJ. 2000. Interspecific differences in habitat use of shorebirds and waterfowl in managed wetlands of California’s San Joaquin Valley. Waterbirds: The International Journal of Waterbird Biology 23:196–203. [Google Scholar]
- Jenness JS 2004. Calculating landscape surface area from digital elevation models. Wildlife Society Bulletin 32:829–839. [Google Scholar]
- Konecny G 1986. First Results of the European Spacelab Photogrammetric Camera Mission Pages 115–120 in Szekielda K-H, editor.Satellite Remote Sensing for Resources Development. Springer; Netherlands, Dordrecht. [Google Scholar]
- Larson P, and Runyan C. 2009, September Evaluation of a Capacitance Water Level Recorder and Calibration Methods in an Urban Environment UMBC/CUERE Technical Memo 2009/003. University of Maryland Baltimore County, Center for Urban Environmental Research and Education, Baltimore, MD. [Google Scholar]
- Los Huertos M, and Smith D. 2013. Chapter 2: Wetland Bathymetry and Mapping Pages 49–86 in Anderson JT and Davis CA, editors.Wetland Techniques. Springer; Netherlands, Dordrecht. [Google Scholar]
- Ma Z, Cai Y, Li B, and Chen J. 2010. Managing wetland habitats for waterbirds: An international perspective. Wetlands 30:15–27. [Google Scholar]
- Mensik GJ, and Reid FA. 1995. Managing problem vegetation. Page 8 Ducks Unlimited. [Google Scholar]
- Munyaneza O, Wali UG, Uhlenbrook S, Maskey S, and Mlotha MJ. 2009. Water level monitoring using radar remote sensing data: Application to Lake Kivu, central Africa. Physics and Chemistry of the Earth, Parts A/B/C 34:722–728. [Google Scholar]
- Neckles HA, Murkin HR, and Cooper JA. 1990. Influences of seasonal flooding on macroinvertebrate abundance in wetland habitats. Freshwater Biology 23:311–322. [Google Scholar]
- NOAA National Centers for Environmental Information. 2017. Climate at a Glance: U.S. Time Series.
- Pekel J-F, Cottam A, Gorelick N, and Belward AS. 2016. High-resolution mapping of global surface water and its long-term changes. Nature 540:418–422. [DOI] [PubMed] [Google Scholar]
- R Core Team. 2012. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. [Google Scholar]
- Reiter ME, Elliott N, Veloz S, Jongsomjit D, Hickey CM, Merrifield M, and Reynolds MD. 2015. Spatio-temporal patterns of open surface water in the Central Valley of California 2000–2011: Drought, land cover, and waterbirds. JAWRA Journal of the American Water Resources Association 51:1722–1738. [Google Scholar]
- Reynolds MD, Sullivan BL, Hallstein E, Matsumoto S, Kelling S, Merrifield M, Fink D, Johnston A, Hochachka WM, Bruns NE, Reiter ME, Veloz S, Hickey C, Elliott N, Martin L, Fitzpatrick JW, Spraycar P, Golet GH, McColl C, and Morrison SA. 2017. Dynamic conservation for migratory species. Science Advances 3:e1700707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rowe DK, Shankar U, James M, and Waugh B. 2002. Use of GIS to predict effects of water level on the spawning area for smelt, Retropinna retropinna, in Lake Taupo, New Zealand. Fisheries Management and Ecology 9:205–216. [Google Scholar]
- Ruszkiczay-Rüdiger Z, Fodor L, Horváth E, and Telbisz T. 2009. Discrimination of fluvial, eolian and neotectonic features in a low hilly landscape: A DEM-based morphotectonic analysis in the Central Pannonian Basin, Hungary. Geomorphology 104:203–217. [Google Scholar]
- Safran RJ, Isola CR, and Williams OE. 1997. Benthic invertebrates at foraging locations of nine waterbird species in managed wetlands of the northern San Joaquin Valley, California. Wetlands 17:407–415. [Google Scholar]
- Schaffer-Smith D, Swenson JJ, Barbaree B, and Reiter ME. 2017. Three decades of Landsat-derived spring surface water dynamics in an agricultural wetland mosaic; Implications for migratory shorebirds. Remote Sensing of Environment 193:180–192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmugge TJ, Kustas WP, Ritchie JC, Jackson TJ, and Rango A. 2002. Remote sensing in hydrology. Advances in Water Resources 25:1367–1385. [Google Scholar]
- Scottish Natural Heritage (Agency). 2006. Waterbirds around the world: a global overview of the conservation, management and research of the world’s waterbird flyways. The Stationery Office, Edinburgh. [Google Scholar]
- Shuford WD, Page GW, and Kjelmyr JE. 1998. Patterns and dynamics of shorebird use of California’s Central Valley. The Condor 100:227–244. [Google Scholar]
- Sona G, Pinto L, Pagliari D, Passoni D, and Gini R. 2014. Experimental analysis of different software packages for orientation and digital surface modelling from UAV images. Earth Science Informatics 7:97–107. [Google Scholar]
- Strong MA, Mensik JG, and Walsworth DS. 1990. Converting rice fields to natural wetlands in the Sacramento Valley of California. Transactions of the Western Section of the Wildlife Society 26:29–35. [Google Scholar]
- Strum KM, Reiter ME, Hartman CA, Iglecia MN, Kelsey TR, and Hickey CM. 2013. Winter management of California’s rice fields to maximize waterbird habitat and minimize water use. Agriculture, Ecosystems & Environment 179:116–124. [Google Scholar]
- Stutzman R 2012. Stopover decisions of migratory shorebirds: an assessment of habitat use, food availability, behavior and phenology. MS Thesis, University of Nebraska-Lincoln. [Google Scholar]
- Tachikawa T, Kaku Manabu, Iwasaki Akira, Gesch Dean B., Oimoen Michael J., Zhang Z, Danielson Jeffrey J., Krieger Tabatha, Curtis Bill, Haase Jeff, Abrams Michael, and Carbajal C 2011. ASTER Global Digital Elevation Model Version 2 - summary of validation results. NASA Earth Resources Observation and Science (EROS) Center. [Google Scholar]
- Taft OW, Colwell MA, Isola CR, and Safran RJ. 2002. Waterbird responses to experimental drawdown: implications for the multispecies management of wetland mosaics: Waterbird communities and wetland drawdown. Journal of Applied Ecology 39:987–1001. [Google Scholar]
- Taft OW, and Elphick CS. 2007. Waterbirds on Working Lands: Literature Review and Bibliography Development. Page 284 Technical Report, National Audubon Society. [Google Scholar]
- Turpie KR, Allen DW, Ackelson S, Bell T, Dierssen H, Cavanaugh K, Fisher JB, Goodman J, Guild L, Hochberg E, Klemas VV, Lavender S, Lee C, Muller-Karger F, Ortiz J, Palacios S, Thompson DR, and Zimmerman R. 2015. New Need to Understand Changing Coastal and Inland Aquatic Ecosystem Services.
- U.S. Fish and Wildlife Service. 2009. Sacramento, Delevan, Colusa, and Sutter National Wildlife Refuges, Final Comprehensive Conservation Plan and Environmental Assessment. Sacramento, CA. [Google Scholar]
- U.S. Fish and Wildlife Service. 2016a. National Wetlands Inventory. http://www.fws.gov/wetlands/.
- U.S. Fish and Wildlife Service. 2016b. Sacramento National Wildlife Refuge Complex GIS Database.
- U.S. Geological Survey. 2015. 10m National Elevation Dataset.
- USDA-NASS. 2014. Agricultural statistics data base. USDA-NASS, Washington DC. [Google Scholar]
- Vermote E, Justice C, Claverie M, and Franch B. 2016. Preliminary analysis of the performance of the Landsat 8/OLI land surface reflectance product. Remote Sensing of Environment 185:46–56. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Western Hemisphere Shorebird Reserve Network. 2009. Western Hemisphere Shorebird Reserve Network Sites. http://www.whsrn.org/whsrn-sites.
- Westoby MJ, Brasington J, Glasser NF, Hambrey MJ, and Reynolds JM. 2012. “Structure-from-Motion” photogrammetry: A low-cost, effective tool for geoscience applications. Geomorphology 179:300–314. [Google Scholar]
- Wilson RM, and Cronon W. 2010. Seeking refuge: birds and landscapes of the Pacific flyway. University of Washington Press, Seattle. [Google Scholar]
- Xiaoye Liu. 2008. Airborne LiDAR for DEM generation: some critical issues. Progress in Physical Geography 32:31–49. [Google Scholar]
- Zharikov Y, Elner RW, Shepherd PCF, and Lank DB. 2008. Interplay between physical and predator landscapes affects transferability of shorebird distribution models. Landscape Ecology 24:129–144. [Google Scholar]
- Zhu Z, and Woodcock CE. 2012. Object-based cloud and cloud shadow detection in Landsat imagery. Remote Sensing of Environment 118:83–94. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.