

Abstract
Following 25 years of extensive research by many scientists worldwide, a panel of ten reward gene risk variants, called the Genetic Addiction Risk Score (GARS), has been developed. In unpublished work, when GARS was compared to the Addiction Severity Index (ASI), which has been used in many clinical settings, GARS significantly predicted the severity of both alcohol and drug dependency. In support of early testing for addiction and other RDS subtypes, parents caught up in the current demographic of 127 people, both young and old, dying daily from opiate/opioid overdose, need help. In the past, families would have never guessed that their loved ones would die or could be in real danger due to opiate addiction. Author, Bill Moyers, in Parade Magazine, reported that as he traveled around the United States, he found many children with ADHD and other spectrum disorders like Autism, and noted that many of these children had related conditions like substance abuse. He called for better ways to identify these children and treat them with approaches other than addictive pharmaceuticals.
To our knowledge, GARS is the only panel of genes with established polymorphisms reflecting the Brain Reward Cascade (BRC), which has been correlated with the ASI-MV alcohol and drug risk severity score. While other studies are required to confirm and extend the GARS test to include other genes and polymorphisms that associate with an hypodopaminergic trait, these results provide clinicians with a non-invasive genetic test.
Genomic testing, such as GARS, can improve clinical interactions and decision-making. Knowledge of precise polymorphic associations can help in the attenuation of guilt and denial, corroboration of family gene-o-grams; assistance in risk-severity-based decisions about appropriate therapies, including pain medications and risk for addiction; choice of the appropriate level of care placement (i.e., inpatient, outpatient, intensive outpatient, residential); determination of the length of stay in treatment; determination of genetic severity-based relapse and recovery liability and vulnerability; determination of pharmacogenetic medical monitoring for better clinical outcomes (e.g., the A1 allele of the DRD2 gene reduces the binding to opioid delta receptors in the brain, thus, reducing Naltrexone’s clinical effectiveness); and supporting medical necessity for insurance scrutiny.
Keywords: African-Americans, Benefits, Genetic Addiction Risk Score (GARS), Hypodopaminergia, P450 system, Pro-dopamine regulation, Reward Deficiency Syndrome (RDS)
Introduction
The interaction of neurotransmitters and genes that control the release of dopamine is the Brain Reward Cascade (BRC) [1]. Variations within the BRC, whether genetic or epigenetic, may predispose individuals to addictive behaviors and altered pain tolerance [2]. The Genetic Addiction Risk Score (GARS), the first test to accurately predict vulnerability to pain, addiction, and other compulsive behaviors that are defined as Reward Deficiency Syndrome (RDS), provides benefits for individuals suffering with Substance Use Disorder (SUD). Innovative strategies to combat the opioid epidemic, including iatrogenic prescription drug abuse and death, based on the role of dopaminergic tone in pain pathways, have been previously proposed [3]. Sensitivity to pain may reside in the mesolimbic projection system, where genetic polymorphisms associate with a predisposition to pain vulnerability or tolerance [4]. This system provides unique therapeutic targets that could assist in the treatment of pain, and identify risk for subsequent addiction. Pharmacogenomic testing of candidate genes like DRD1, DRD2, DRD 3, DRD 4; MOA-A; COMT; DAT1; 5HTTLLR; OPRM1; and GABRA3 might result in pharmacogenomic, personalized solutions, and improved clinical outcomes. Genetically identifying risk for all RDS behaviors, especially in compromised populations, may be a frontline tool to assist municipalities to provide better resource allocation
The inter-relationship of at least four important neurochemical pathways: serotonergic, endorphinergic, GABAergic, and dopaminergic constitute the “brain reward cascade” (see Figure 1) a natural sequence of events that produce feelings of well-being. Synthesis, vesicle storage, metabolism, release, and function of these neurochemicals are regulated by gene transcription and RNA-directed translation to proteins. Thus, genetic testing is a potential window that can be used to identify the specific neurochemistry of individuals and formulate the best treatment options for them [5].
Figure 1.
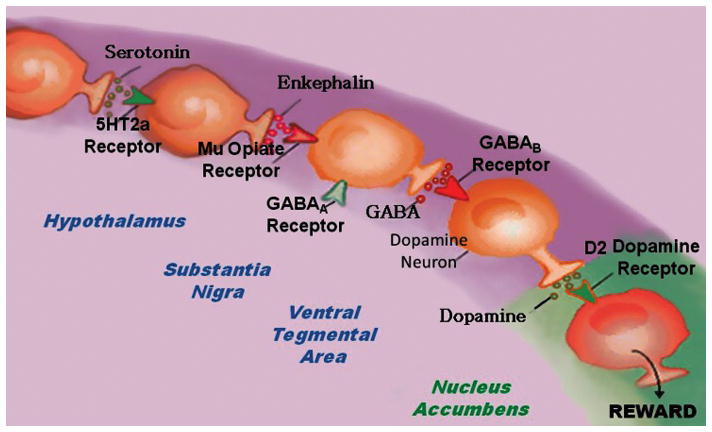
Brain Reward Cascade
In this cascade [6,7], stimulation of the serotonergic system in the hypothalamus leads to the stimulation of delta/mu receptors by serotonin to cause a release of enkephalin. Activation of the enkephalinergic system induces an inhibition of GABA transmission in the midbrain by enkephalin stimulation of mu receptors at GABA neurons. This inhibitory effect allows for the fine-tuning of GABA activity. This provides the normal release of dopamine at the projected area of the NAc.
DA is a neurotransmitter with multiple important functions including behavioral effects such as “pleasure” and “stress reduction.” Without the normal function of this substance, an individual will suffer from cravings and have an inability to cope with stress. Thus, genetic hypodopaminergic brain function predisposes individuals to seek substances and or behaviors that can be used to overcome this craving state by activating the mesolimbic dopaminergic centers [8,9]. Psychoactive substances like alcohol, psychostimulants and opiates, and risky behaviors like gambling, overeating and thrill seeking [10] induce the release of neuronal DA into the synapse at the NAc, to overcome the hypodopaminergic state of that individual.
Temporary relief from the discomfort and a pseudo sense of well-being is the product of this self-medication [11]. However, chronic abuse of psychoactive substances leads to inactivation, or a downregulation, like for example, inhibition of neurotransmitter synthesis, neurotransmitter depletion, formation of toxic pseudo neurotransmitters, and through structural receptor dysfunction. Therefore, both non-dependent substance-seeking and pathological behaviors associated with dependence are both used as a means of providing a feel-good response, a “fix”, to lessen uncontrollable cravings. Individuals who possess reward gene polymorphisms or variations, will, given environmental insult be at risk for impulsive, compulsive, and addictive behaviors. Reward Deficiency Syndrome (RDS) is a term used to embrace and characterize these genetically induced behaviors [12]. Any and all of these pathological behaviors, as well as psychoactive drug-abuse, are candidates for addiction including tolerance and dependence. The behavior or drug of choice by the individual is a function of both genes and environmental factors like availability and peer pressure.
Previously we published on the potential of GARS to predict vulnerability or risk for both drug and alcohol severity as measured by the Addiction-Severity Index (ASI) [13]. However, a frequently raised question relates to what is the benefit of GARS™ testing in known addicts already in treatment programs. We believe that there are many important reasons for GARS testing in people expressing addictive behaviors of all types.
Denial
It is well-known that many patients in treatment programs deny that they have a biological problem and are therefore able to control addictions [14]. Through the “war on drugs,” the just-say-no campaign, and into the early years of this century, the overarching approach to Substance Use Disorders (SUDs) called for a single outcome (abstinence) and a single methodology (spiritual connection with a higher power) as the remedy for SUDs. Those who did not become permanently abstinent or rejected the spiritual approach were seen as “not ready” or “in denial”. A seismic shift in thinking about “addiction” and “recovery” began in earnest in the 1990s. In 2005, the Substance Abuse and Mental Health Services Administration brought together leaders of the treatment and recovery field for the historic National Summit on Recovery to develop broad-based consensus on guiding principles for recovery and elements of recovery-oriented systems of care. Major changes associated with the recovery-oriented approach include viewing SUDs as chronic, rather than acute, problems that require long-term support and a focus on recovery management rather than disease management.
Complete abstinence is not an absolute requirement for wellness for all persons with SUDs. There are “many pathways to recovery,” not only the 12-Step approach [15]. Sustained recovery is self-directed and requires personal choices, the support of peers and allies, and community reinforcement as well as a strength-based approach and the use of research-based interventions. So many in recovery and currently in treatment have denial, and the addiction medicine community is attempting to promote recovery-oriented approaches to reduce misconceptions, labeling, and stigmatization, and to promote recovery for individuals, families, and communities. Providing real evidence genetically (GARS) to predict risk for both substance and non-substance severity helps reduce individuals’ denial of the problem.
Guilt/Shame
A very common response from people already addicted is a profound sense of shame and guilt, not realizing that they are not alone [16]. Addiction is a person-level phenomenon that involves in many cases both guilt and shame. Feeling shame for addiction is not a mistake. It is part of the shape of addiction, part of the normal phenomenology of addiction, and often a source of motivation for the addict to heal. Like other recent attempts in the addiction literature to return normative concepts such as “choice” and “responsibility” to their rightful place in understanding and treating addiction, the ongoing effort to help remove both guilt and shame is compatible with investigation of genetic and neuroscientific causes of addiction. Certainly, there can be shame without blame. However, in our view a plausible key to attenuating addiction is to arrest it, to stop the addict from using. There is the documented view that reveals in many cases that addicts fail to be able to exert normal self-control capacities and are ashamed of both this fact and the fact that they are failing to live a good human life. Understanding the false premise “Just Say No” is normally not enough to stop abusing drugs. For this, we are working toward pharmacological interventions that help addicts to stop using, by induction of “dopamine homeostasis” [17] which is a necessary condition for any and all further healing. Eventually, work in genetics will yield simple interventions that adjust genes for those predisposed. We the authors believe that these novel interventions might work to arrest addiction in the future, and as such acknowledge that epigenetic impact on reward gene expression is a cornerstone cause of addiction and should be integrally considered in any gene-based therapy [18]. Albeit, not as an excuse, providing biological and genetic (GARS) evidence to predict risk for both substance and non-substance severity helps remove both guilt and shame.
Genogram Confirmation
In most chemical and non-chemical dependency programs the patients are usually asked to provide a family history of all kinds of addiction in the form of a family tree called a Genogram [19]. A genogram (also known as a McGoldrick-Gerson study, a Lapidus Schematic or a Family Diagram) is a pictorial display of a person’s family relationships and medical history. It goes beyond a traditional family tree by allowing the user to visualize hereditary patterns and psychological factors that punctuate relationships. A genogram is a multi-generational diagram of a person’s family and social network. It allows users to view multiple relationship dynamics, review developmental influence and identify trends. Each person on a genogram is represented by a symbol. The symbols are then linked with lines to depict various dynamics and significant individual qualities. Genograms are used by professionals in many fields which study and work with people including doctors, researchers, psychiatrists, counselors and psychologists.
The Genogram in addiction provides a non-definitive snapshot for the substance abuse counselor whereby they will be able to:
demonstrate the use of family systems therapy to assist families in recovery
define traits of healthy families and techniques for assisting families to develop those traits
illustrate the process for breaking the cycle of addiction in multi-generational families
identify different family treatment methods based on family structure
Medical and behavioral researchers use the results of multiple genograms to identify recurring patterns. Interviewing individuals from different generations and coding the information provides clues to possible causal or correlation elements, such as genetics or generational learning. Genograms may also be used to study how factors such as the environment or socio-economic status influence personal or family development and functioning. Offering the GARS test to a person’s family in treatment is the best way to confirm the risk of addiction in the family to help confirm the genetic basis of the Genogram.
Medication Assisted Treatment (MATS) Dosing
There is no question that America is experiencing a horrific opiate/opioid epidemic whereby thousands of people are unfortunately dying, and the rate of people seeking treatment is at an all-time high. One major problem can be linked to the fact that legal prescriptions for powerful opioid analgesics reached 297 million in 2016. One company that manufactures Oxycontin generated $3.1 billion in revenue in 2017. Moreover, deaths from prescription drug overdoses have been called the “silent epidemic” for many years. Indeed, approximately one American is dying every 17 minutes from accidental prescription drug overdose [20,21].
There is a plethora of research indicating the successful treatment of opioid dependence with either buprenorphine alone or in combination with naloxone (Suboxone®). However, we encourage caution in long-term maintenance with these drugs, albeit, options are minimal as there is a lack of any other FDA approved opioid maintenance compound to date. Our concern has been supported by severe withdrawal (even with tapering of the dosage of, for example, Suboxone® which is 40 times more potent than morphine) from low dose of buprenorphine (alone or with naloxone). In addition, our findings of a long-term flat affect in chronic Suboxone® patients amongst other unwanted side effects including diversion and suicide attempts provides impetus to reconsider long-term utilization. However, it seems prudent to embrace genetic testing to reveal reward circuitry gene polymorphisms especially those related to dopaminergic pathways as well as opioid receptor(s) as a way of improving treatment outcomes. Understanding the interaction of reward circuitry involvement in buprenorphine effects and respective genotypes provides a novel framework to augment a patient’s clinical experience and benefits during opioid replacement therapy [4].
It is important to realize that clinical outcome in drug addicted patients including alcoholism may depend upon dopaminergic genes and associated polymorphisms. In 1995, Lawford et al. showed in a double-blind study, that when bromocriptine (a DRD2 agonist) or placebo was administered to alcoholics with either the A1 (A1/A1 and A1/A2 genotypes) or only the A2 (A2/ A2 genotype) allele of the DRD2 gene, the greatest improvement in craving and anxiety occurred in the bromocriptine-treated A1 alcoholics. Importantly, the attrition or compliance to treatment was highest in the placebo-treated A1 alcoholics suggesting treatment outcome is a function of genotype [22,23].
The concept of the feasibility of treating RDS based on pharmacogenetics has been further underscored by Blum et al. [24] They found that the DRD2 gene polymorphism (A1 allele vs A2 allele) had a significant Pearson correlation with days in treatment (r=0.42). Compared to the DRD2 A1-carriers the number of days in treatment with the putative natural dopamine agonist KB220 was 51.9 ± 9.9 SE (95%CI, 30.8 to 73.0) and for the DRD2 A1+ carriers the number of days on treatment with KB220 was 110.6 ± 31.1 (95% CI, 38.9 to 182.3). Once again, the attrition was highest in the A1− genotype group. It was suggested that the genotype may be a predictor of treatment persistency and compliance. Moreover, even relapse may depend on the DRD2 A1 allele which could affect treatment response. Dahlgren et al. [25] provided the first report of an association between the TaqI A1 allele and a substantially increased relapse rate in alcohol dependent patients.
Along similar lines, Noble & Ritchie [26] measured [3H] Naloxone binding in frontal gray cortex, caudate nucleus, amygdala, hippocampus and cerebellar cortex obtained post mortem from human alcoholic and non-alcoholic subjects. When subjects were grouped by the presence or absence of the A1 allele of the D2 dopamine receptor gene, [3H]naloxone binding was lower in all brain regions examined of subjects with the A1 allele than in those without this allele, with a significant difference in the caudate nucleus. It was suggested that the decreased [3H] naloxone binding observed in subjects with the A1 allele may be a compensatory response to their decreased dopaminergic modulation of opiate receptor activity.
Interestingly, Gerra et al. [27] provided clear evidence that the dopaminergic system is linked to a buprenorphine treatment response in heroin addicted humans. Surprisingly, they found no difference between responders and non-responders to buprenorphine in the frequency of kappa opioid receptor (OPRK1) 36G>T SNP. However, the frequency of dopamine transporter (DAT) gene polymorphism (SLC6A3/DAT1), allele 10, was much higher in “non-responder” than in “responder” individuals (64.9% vs. 55.93%) whereas the frequency of the category of other alleles was higher in responder than in non-responder individuals (11.02% vs. 2.13% respectively). Our own interpretation of these results dovetail with the work of others [4, 22, 23] that show better treatment outcome and compliance based on dopaminergic polymorphisms whereby hypodopaminergic traits mediate a better response during treatment. We hypothesize that carriers of the 9 alleles of the DAT1 would confer a better treatment response with buprenorphine due to its faster transport activity resulting in a hypodopaminergic trait.
Finally, while Barratt et al. [28] did not show significant differences in methadone or buprenorphine outcomes in terms of maintenance with carriers of the Taq1A A1 allele, methadone subjects did show that significantly fewer A1 allele carriers experienced withdrawal compared to non-A1 carriers (P = 0.04). Moreover, our laboratory [29] found in a genetically determined hypodopaminergic trait patient at 432 days post Suboxone® withdrawal being maintained on a putative dopamine agonist KB220Z, has been urine tested and is opioid free. Genotyping data revealed a moderate genetic risk for addiction showing a hypodopaminergic trait. In agreement with these findings, Makhinson and Gomez-Makhinson [30] observed in a case report that buprenorphine withdrawal syndrome with predominant symptoms of restlessness resistant to clonidine and benzodiazepines, was successfully treated with the dopamine agonist pramipexole.
The constant controversy over either dopamine antagonistic compared to dopamine agonistic therapy or simply put treating the dopaminergic surfeit or deficit has been the recent subject of paper published in Nature Neuroscience [31]. Specifically, Willuhn et al. [31] found that phasic dopamine decreased as the rate of cocaine intake increased, with the decrement in dopamine in the ventromedial striatum (VMS) significantly correlated with the rate of escalation. This work suggests that the “deficit” relative to “surfeit” theory requires dopaminergic agonistic rather than antagonistic treatment.
As has been proposed previously, activation rather than blocking mesolimbic dopaminergic reward circuitry in the long-term treatment of RDS is the preferred modality [32]. Although, the acute treatment should consist of preferential blocking of postsynaptic NAc DA receptors (D1-D5), the long-term mesolimbic activation of the dopaminergic system should involve the release and/or activation of DA at the NAc site. This theory suggests that excessive craving behavior can be attributed to reduced number of DA D2 receptors, an effect of carrying, for example, the DRD2 A1 allelic genotype, whereas a normal or sufficient density of D2 receptors results in reduced craving. A goal, in terms of preventing substance abuse, could be to induce a proliferation of D2 receptors in individuals who are genetically vulnerable. While in vivo experiments that used a typical D2 receptor agonist induce down-regulation [33], in vitro experiments have shown that in spite of genetic antecedents, constant stimulation with a known D2 agonist, bromocriptine, results in significant proliferation of D2 receptors within the DA system. However, chronic treatment with bromocriptine results in down-regulation of D2 receptors, instead of the up-regulation proposed for the KB220Z prodopamine regulation, and that could be a reason for failure in treatment with agonists. In other unpublished work by Chapman et al. it was found that African-Americans carrying CYP3A4 Genotype *1B, the extended metabolic Buprenorphine genotype *1/*1B (43%) and *1B/1B* (42%) compared to ~9000 Caucations (26%) differed significantly [34]. Besides buprenorphine personalized dosing, a candidate GARS panel approach can provide useful information for, preliminary screening for high risk patients in pain clinics and relapse-prevention. Data from our funded 1R41MD012318-01 grant will involve African-Americans carrying CYP3A4 Genotype *1B the extended metabolism Buprenorphine genotype coupled with GARS to address the problem of diversity and addiction risk (see Table 1).
Table 1.
% of CYP3A4 *1B genetic variant. Chapman et al [59].
| CYP3A4 Genotype | CYP3A4 Phenotype | % of patients | |
|---|---|---|---|
| Dr. Chapman’s patient population | ~9000 patients evaluated | ||
| *1/*1B | Extensive Metabolizer | 43% | 17 % |
| *1B/*1B | 42% | 9% | |
It is a fact, that certain genetic variations (as seen in GARS) like that observed in the opioid mu receptor (reduce number of receptors), for example, will result in dosing consequences whereby higher doses of Buprenorphine (a MAT) may be needed to prevent relapse to street heroin.
Resource Allocation
Stepped care models aim at matching treatment intensity to defined patient characteristics in a systematic way, thereby avoiding misplacements and making best use of available treatment resources at the same time. In principle, treatment planning for new patients starts with the least intensive care, progressing to more intensive regimes for non-responders. Such models have been introduced in psychiatry and in other medical fields. Models of stepwise patient placement in addiction treatment are known from Northern America (Sobell model, model of the American Society of Addiction Medicine, ASAM) [34] for adults and adolescents and special models for dual-diagnosis patients. Another model comes from Europe (the Dutch model for triage and evaluation in addiction treatment MATE; special model for judicial patients). Since placement in either Home 1 compared to Home 2 (more intense) requires feasibility, validity, reliability, effectiveness, and cost-effectiveness, GARS testing will negate guessing and provide a genetically based method of real resource allocation methodology.
Opioid Pain Compound Avoidance
Understanding the role of neurogenetics of opioids and its role in pain mechanisms has been extensively studied. Results indicated that both sensitivity and tolerance to morphine were found to be dependent on genotype, with inheritance characterized by dominance or partial dominance and involves many published works [35–44].
Differences in human responses to opioids have been well known for some time, for example, a particular type of opioid may provide better analgesia than other opioids for any one individual. Differences in individual responses to an analgesic effect are not unique; differences can also be seen with other opioid effects such as interactions, side effects, and toxicities. As research gained from databases on knockout rodents, pharmacogenetics, and gene polymorphisms unravels various genetic receptor interactions, and biochemical differences of opioid responses in humans, some of the differences may be exploited to provide better care. Testing will become more readily available and cost-effective as an aid to clinicians. Instead of having to rely solely on patient feedback, clinical judgment and trial and error, clinicians will be able to predict patient responses to doses of specific opioids, individualized opioid analgesic therapy, and devised optimal opioid rotation strategies. In the future, information of this type may translate into improved patient care, as clinicians become adept at tailoring appropriate opioid therapy. Although presently perfect candidate genes for gene-directed opioid therapy are not obvious [45–46], certain candidate genes have been studied, and associations with analgesic requirements for acute and chronic pain states, as well as with sensitivity to the pain, have been found [37].
These associations with analgesia and chronic pain were a consequence of an intense investigation of the candidate genes for the catechol-O-methyl-transferase, melanocortin-1 receptor, guanosine triphosphate glycohydrolase, and the mu-opioid receptor. The genetic variants of drug-metabolizing enzymes, in contrast, have well known and described impacts on responses to pharmacotherapy. The analgesic efficacy of codeine, tramadol, nonsteroidal anti-inflammatory drugs and tricyclic antidepressants are influenced by polymorphisms of the cytochrome P450 enzymes. For example, genetically caused cytochrome P450 (CYP) 2D6 inactivity, renders codeine ineffective due to its non-conversion to morphine, slightly decreases the clearance of methadone and the efficacy of tramadol due to lack of formation of the active O-desmethyl-tramadol [38–40].
In an animal genetic experiment Mogil et al. investigated sensitivity and tolerance to morphine. They used two strains of mice and C57BL/6By and BALB/cBy, and seven recombinant inbred strains of their reciprocal F1 hybrids. Following the administration of 20 mg/kg of morphine hydrochloride or saline, sensitivity was measured using a locomotive activity. The ‘hot plate’ method was used to measure tolerance following the single or repeated administration of 20 mg/kg of morphine hydrochloride or saline. Results indicated that both sensitivity and tolerance to morphine were found to be dependent on genotype, with inheritance characterized by dominance or partial dominance [47]. Ongoing research (GARS testing) will target other candidate gene polymorphisms and drug metabolizing enzyme genetic variants searching for associations between drug response and an individual’s genetic profile (pharmacogenetics).
The mu opioid receptor gene encodes the receptor targets for some endogenous opioids and studies of mu-opioid receptor polymorphisms have contributed substantially to knowledge about genetic influences on cocaine and opiate addiction (including heroin, morphine, and synthetic opioids). Monoaminergic system genes, particularly those encoding the dopamine, serotonin, and norepinephrine transporters, and dopamine β-hydroxylase, as well as those of the endogenous opioid system have also been studied [39].
Methadone is an opiate used in substitution therapy to treat opioid dependence. Variability in individualized responses to methadone dosage, effects program retention rates, due in part to withdrawal symptoms and further heroin craving and use, caused by low, non-optimal dosing. Methadone is a substrate for the P-glycoprotein transporter, encoded by the ABCB1 gene, which regulates central nervous system exposure. Coller et al. demonstrated that ABCB1 genetic variability influenced daily methadone dose requirements. They found that subjects who are carrying two copies of the wild-type haplotype when compared with those carriers of no copies or one copy required higher methadone doses (98.3. ± 10.4., 55.4. ± 26.1. and 58.6. ± 20.9.mg/d, respectively; P = 0.0.29). This is possibly a protective effect. They also found that doses that are significantly lower are required by carriers rather than noncarriers of the AGCTT haplotype (38.0. ± 16.8. and 61.3. ± 24.6. mg/d, respectively; P = 0.0.4). Thus, ABCB1 genetic variability may offer help for clinical methadone dosage individualization [38]. Opioids are among the P-glycoprotein substrates. Opioid pharmacology may be affected by Multi-Drug Resistance Gene (MDR1) mutations. Higher fentanyl doses are required by carriers of the mutated G118 allele. The G118 allele has been associated with decreased analgesic effects including decreased potency of morphine and morphine-6-glucuronide [40,42]. Clinical response to opioid therapy can be altered by genetic variations which may trigger or modify drug interactions. Another example is the inhibition of CYP2D6 paroxetine which in extensive metabolizers of debrisoquine/ sparteine but not in poor metabolizers increases the steady-state plasma concentrations of (R)-methadone [40,42,48].
The clinical consequences of opioid Pharmacogenetics have so far been limited. Genetically precipitated drug interactions that might cause standard opioid doses to be toxic require caution and codeine should not be administered to poor metabolizers of debrisoquine/sparteine. The on-demand administration of opioids would limit the utility of understanding the effects of mutations on opioid receptors, pain perception, and pain processing, to merely explaining why some patients require higher opioid doses, and the adverse effects profile of patients, may indeed, be modified by these mutations. An example is labor analgesia; women with the mu opioid receptor 304G variant demonstrate more responsiveness to opioids and require significantly reduced intrathecal fentanyl [48]. These findings for intrathecal fentanyl pharmacogenetics may have implications for patients receiving opioids in other settings [48–49]. Thus, Pharmacogenetics can be expected to facilitate individualized opioid therapy especially for African-Americans concerning buprenorphine dosing.
The following sampling of the genes involved in the addictive process can also be indicative of which genes are engaged in pain mechanisms, pain sensitivity, and opiate addiction. The list includes the mu opioid receptor, a δ-opioid receptor, metabotropic receptors; mGluR6 and mGluR8, nuclear receptor NR4A2, and photolyase-like cryptochrome 1. The dopamine receptor genes 1 to 5, dopamine transporter gene DAT1, Dopamine Beta-Hydroxylase (DBH), proenkephalin (PENK) and prodynorphin (PDYN) genes are implicated. The CAMKII enzyme, Gonadotrophin-releasing hormone (GnRH), and the CYP2D6, CYP2B6, CYP2C19, and CYP2C9 genes members of the cytochrome P450 superfamily of enzymes have a role. Brain-derived neutropenic factor (BDNF), and Neurotrophin-3 NT-3 are neurotrophic genes, and OPRM1, G-protein alpha subunits, 5q33, GABA (A) gamma2 the GABA receptor subunit genes, and OPRK1, alpha2-adrenoceptor are involved. The TTC12, ANKK1, NCAM1, and TTC12 are important for drug exposure in heroin dependence [51] and morphine stimulates zinc finger CCHC-type, RNA-binding motif 1 (ZCRB1) (10) and RGS-R7 [42–45]. Other genes involved include Interleukin-2, Gbeta5, MAO-A, 287 A/G polymorphism of catechol-O-methyltransferase, serotonin transporter, Ca2+/ cAMP responsive element binding protein, CNR1, ABCB1, P-glycoprotein, UGT2B7, and CREB. Some genes are involved in pain mechanisms and the healing process, the following tables represent a sampling (see Table 2).
Table 2.
Genes involved in pain mechanisms. Taken from Blum et al. [2]
| Gene name | Polymorphism | Pathway (s) | Reference (s) |
|---|---|---|---|
| Human κ opioid receptor gene (OPRK1) | In humans, the 36G > T single nucleotide polymorphism (SNP) on KOR gene | The κ opioid receptor (KOR) system seems to play a role in stress responsivity, opiate withdrawal and responses to psychostimulants, inhibiting mesolimbic dopamine. KOR gene polymorphisms have been reported to contribute to predisposition to voluntary alcohol-drinking behavior in experimental animals. | Gerra G, Leonardi C, Cortese E, D’Amore A, Lucchini A, Strepparola G, et al. Human kappa opioid receptor gene (OPRK1) polymorphism is associated with opiate addiction. Am J Med Genet B Neuropsychiatr Genet 2007;144 (6): 771–775. |
| Mu opioid receptor | A118G SNP of the mu-opioid receptor gene (OPRM1) | Mu opioid receptors are critical for heroin dependence, and A118G SNP of the mu-opioid receptor gene (OPRM1) has been linked to heroin abuse. In our population of European Caucasians (n = 118), approximately 90% of 118G allelic carriers were heroin users. | Drakenberg K, Nikoshkov A, Horváth MC, Fagergren P, Gharibyan A, Saarelainen K, et al. Mu opioid receptor A118G polymorphism in association with striatal opioid neuropeptide gene expression in heroin abusers. Proc Natl Acad Sci USA 2006;16;103 (20): 7883–7888. |
| D (2) dopamine receptor gene (DRD2) | A haplotype block of 25.8. kb was defined by 8 SNPs extending from SNP3 (TaqIB) at the 5′ end to SNP10 site (TaqIA) located 10 kb distal to the 3′ end of the gene | Within this block, specific haplotype cluster A (carrying TaqIB1 allele) was associated with a high risk of heroin dependence in Chinese patients (P = 1.4.25 × 10 (−22); odds ratio, 52.8.0; 95% confidence interval, 7.2.90–382.5. for 8-SNP analysis). A putative recombination ‘hot spot’ was found near SNP6 (intron 6 ins/del G), creating 2 new daughter haplotypes that were associated with a lower risk of heroin dependence in Germans (P = 1.9.4 × 10 (−11) for 8-SNP analysis). Other studies show the relationship of carrying TAq1A1 vs. A2 alleles in the treatment outcomes for heroin abuse. The results indicate that DRD2 variants are predictors of heroin use and subsequent methadone treatment outcome, and suggest a pharmacogenetic approach to the treatment of opioid dependence. Others found an association between nasal inhalation of opiates and DRD2 promoter-141 DeltaC polymorphism. Significantly stronger cue-elicited heroin craving was found in individuals carrying D2 dopamine receptor gene (DRD2) TaqI RFLP A1 allele than the non-carriers (P < 0.0.01). | Xu K, Lichtermann D, Lipsky RH, Franke P, Liu X, Hu Y, et al. Association of specific haplotypes of D2 dopamine receptor gene with vulnerability to heroin dependence in 2 distinct populations. Arch Gen Psychiatry 2004;61 (6):597–606. Lawford BR, Young RM, Noble EP, Sargent J, Rowell J, Shadforth S, et al. The D (2) dopamine receptor A (1) allele and opioid dependence: association with heroin use and response to methadone treatment. Am J Med Genet 2000;96 (5):592–598. Li Y, Shao C, Zhang D, Zhao M, Lin L, Yan P, et al. The effect of dopamine D2, D5 receptor and transporter (SLC6A3) polymorphisms on the cue-elicited heroin craving in Chinese. Am J Med Genet B Neuropsychiatr Genet 2006;141 (3):269–273. |
| ANKK1 gene | With a non-synonymous G to A transition, rs2734849 produces an amino acid change (arginine to histidine) in C-terminal ankyrin repeat domain of ANKK1 | Since DRD2 expression is regulated by the transcription factor NF-κB, we suspect that rs2734849 may indirectly affect dopamine D (2) receptor density. The rs273849 ANNK1 variant alters the expression level of NF-kappaB-regulated genes. | Huang W, Payne TJ, Ma JZ, Beuten J, Dupont RT, Inohara N, et al. Significant association of ANKK1 and detection of a functional polymorphism with nicotine dependence in an African-American sample. Neuropsychopharmacology; 2008. |
| Catechol-O-methyltransferase (COMT) gene | Val (108/158) met polymorphism of the catechol-O-methyltransferase (COMT) gene | Genotyping 38 Israeli heroin addicts and both parents using a robust family based haplotype relative risk (HRR) strategy. There is an excess of the val COMT allele (likelihood ratio = 4.4.8, P = 0.0.3) and a trend for an excess of the val/val COMT genotype (likelihood ratio = 4.9.7, P = 0.0.8, 2 df) in the heroin addicts compared to the HRR control group. | Horowitz R, Kotler M, Shufman E, Aharoni S, Kremer I, Cohen H, et al. Confirmation of an excess of the high enzyme activity COMT Val allele in heroin addicts in a family-based haplotype relative risk study. Am J Med Genet 2000;96 (5):599–603. Cao L, Li T, Xu K, Liu X. Association study of heroin dependence and -287 A/G polymorphism of catechol-O-methyltransferase gene). In: Zhonghua Yi, Xue Yi, Chuan Xue, Za Zhi, editors. 2002;19 (6):499–501. |
| Proenkephalin gene (PENK) | > or =81 bp allele | Among the subjects with opioid dependence, 66% carried the > or =81 bp allele compared with 40% of subjects with other types of substance abuse (χ2 = 11.3.1, p < 0.0.04) and 49% of controls (χ2 = 6.0., p < 0.0.15). These results are consistent with a role of the PENK gene in opioid dependence. In another study, Heroin abuse was significantly associated with PENK polymorphic 3’ UTR dinucleotide (CA) repeats; 79% of subjects homozygous for the 79-bp allele were heroin abusers. Such individuals tended to express higher PENK mRNA than the 81-bp homozygotes, but PENK levels within the nucleus accumbens (NAc) shell were most strongly correlated to catecholamine-O methyltransferase (COMT) genotype. Altogether, the data suggest that dysfunction of the opioid reward system is significantly linked to opiate abuse vulnerability, and that heroin use alters the apparent influence of heritable dopamine tone on mesolimbic PENK and tyrosine hydroxylase function. | Comings DE, Blake H, Dietz G, Gade-Andavolu R, Legro RS, Saucier G, et al. The proenkephalin gene (PENK) and opioid dependence. Neuroreport. 1999;10 (5):1133–1135 Nikoshkov A, Drakenberg K, Wang X, Horvath MC, Keller E, Hurd YL. Opioid neuropeptide genotypes in relation to heroin abuse: dopamine tone contributes to reversed mesolimbic proenkephalin expression. Proc Natl Acad Sci USA 2008;105 (2):786–791. |
| Serotonin transporter (hSERT) | Homozygosity at hSERT (especially 10/10) was associated with early opiate addiction, while genotype 12/10 proved to be protective | Reward system pathway | Galeeva AR, Gareeva AE, Iur’ev EB, Khusnutdinova EK. VNTR polymorphisms of the serotonin transporter and dopamine transporter genes in male opiate addicts. Mol Biol (Mosk). 2002;36 (4):593–598. Bonnet-Brilhault F, Laurent C, Thibaut F, Campion D, Chavand O, Samolyk D, et al. Serotonin transporter gene polymorphism and schizophrenia: an association study. Biol Psychiatry 1997;42 (7):634–636. |
| Dopamine transporter (DAT1) | In the case of DAT1, genotype 9/9 was associated with early opiate addiction. The combination of hSERT genotype 10/10 with DAT1 genotype 10/10 was shown to be a risk factor for opiate abuse under 16 years of age | Reward system pathway | Galeeva AR, Gareeva AE, Iur’ev EB, Khusnutdinova EK. VNTR polymorphisms of the serotonin transporter and dopamine transporter genes in male opiate addicts. Mol Biol (Mosk). 2002;36 (4):593–598. |
| Cannabinoid CB1 (brain) receptor gene (CNR1) | A microsatellite polymorphism (AAT)n at the cannabinoid CB1 (brain) receptor gene (CNR1) consists of 9 alleles. The number of i.v. drugs used was significantly greater for those carrying the > or ≥ or = 5 genotype than for other genotypes (P = 0.0.05) | Cannabinoid receptors in the modulation of dopamine and cannabinoid reward pathways | Comings DE, Muhleman D, Gade R, Johnson P, Verde R, Saucier G, MacMurray J. Cannabinoid receptor gene (CNR1): association with i.v. drug use. Mol Psychiatry. 2000;5 (2):128–130. |
We are faced with an out of control American opioid epidemic. The main legal gateway to opioid addiction and abuse starts in many cases with iatrogenic prescribing of powerful analgesics (e.g. OxyContin®). One way to prevent this legal dilemma is provided by GARS to unravel opioid dependence risk and seek out other non-opioid pain relievers (e.g. electrotherapy and non-steroid analgesics) leading to Opioid pain compound avoidance.
Pro-Dopamine Regulation
“Gene Guided Precision Nutrition™” and KB220 variants (a complex mixture of amino-acids, trace metals, and herbals) are the pioneers and standard-bearers for a state of the art DNA customization. Findings by both, Kenneth Blum, Ph.D. and Ernest Noble, Ph.D and others [50] concerning the role of genes in shaping cravings and pleasure- seeking, opened the doors to comprehension of how genetics control our actions and effect our mental and physical health. Moreover, technology that is related to KB220 variants in order to reduce or eradicate excessive cravings by influencing gene expression is a cornerstone in the pioneering of the practical applications of nutrigenomics/neurogenetics [51]. Continuing discoveries have been an important catalyst for the evolution, expansion, and scientific recognition of the significance of nutrigenomics and its remarkable contributions to human health. Neuro-Nutrigenomics is now a very important field of scientific investigation that offers great promise to improving the human condition. In the forefront is the development of the Genetic Addiction Risk Score (GARS™), which has predictive value for the severity of drug and alcohol abuse as well as other non-substance related addictive behaviors [13]. While customization of neuronutrients has not yet been commercialized, there is emerging evidence that in the future, the concept will be developed and could have a significant impact in addiction medicine.
It has been established that inducing what has been termed “dopamine homeostasis (balance)” across the brain reward circuitry is the best way to treat all addictive-like behaviors [52]. Thus, GARS testing of the already addicted person provides an exact mirror into the brain’s chemical messenger function (receptor number and chemical production) and can lead to personalized addiction medicine based on Pro-dopamine regulation.
Summary
It is now known that in terms of nature (genes) and nurture (environment) and behavioral outcome in Homo sapiens, the contribution is 50% genes and 50% epigenetics. Thus, molecular genetics or DNA testing is very important, especially linking aberrant behaviors to any individual.
Blum’s laboratory proposed that any disturbance along the brain reward cascade due to either gene variations (polymorphisms) or environment (epigenetics) will result in aberrant addictive behaviors or RDS. In spite of a global search to uncover specific or candidate genes, or even clusters of genes characterized from high-density SNP arrays, it is well known that many attempts have not replicated or have been inconclusive. However, Palmer et al. [53] recently showed that between 25–36% of the variance in the generalized vulnerability to substance dependence is attributable to common single nucleotide polymorphisms. Moreover, the additive effect of common single nucleotide polymorphisms is shared across principal indicators of comorbid drug problems. Furthermore, as a result of these studies, more recent evidence has revealed that specific candidate gene variants account for risk prediction.
Adopting a Bayesian approach, earlier studies from Blum’s laboratory [54] determined a Positive Predictive Value (PPV) for the DRD2 A1 variant (low number of D2 receptors) of 74%, indicating that if a child is born with this polymorphism, they have a very high risk of becoming addicted to either drugs, food, or aberrant behaviors at some point in their lives [55]. Over the many years to come since the 1990 finding on the DRD2 gene association of the Taq A1 allele and severe alcoholism [50], laboratories all across the globe including NIDA and NIAAA not only confirmed this early work [56,57], but also extended the magnitude of many other candidate genes, especially genes and second messengers located in the reward circuitry of the brain [2].
An example includes: Moeller et al. [58] who suggested that drug cues contribute to relapse, and their neurogenetic results have identified the DAT1R 9R allele as a vulnerability allele for relapse, especially during early abstinence (e.g., detoxification). The DAT1R 9R allele influences the fast-acting transport of dopamine, sequestered from the synapse, leading to a hypodopaminergic trait.
It is important to be cautious to accept such genetic testing that uncovers reward circuitry gene polymorphisms, particularly those linked to dopaminergic pathways as well as opioid receptor(s) as a method of obtaining better treatment results. Comprehending the relationship between the reward circuitry’s participation in buprenorphine outcomes and its corresponding genotypes deliver an innovative model to enhance a patient’s clinical experience and improvements throughout opioid replacement therapy [4]. In fact, Blum’s group’s genetic risk score represents a panel of known reward genes and associated risk polymorphisms providing genetic risk for addiction and other behaviors, including medical monitoring and clinical outcome response [4,13].
References
- 1.Blum K, Kozlowski GP. Ethanol and neuromodulator interactions: a cascade model of reward. In: Ollat H, Parvez S, Parvez H, editors. Alcohol and behavior. Utrecht, Netherland, Japan: 1990. pp. 131–149. [Google Scholar]
- 2.Chen AL, Chen TJ, Waite RL, Reinking J, Tung HL, et al. Hypothesizing that brain reward circuitry genes are genetic antecedents of pain sensitivity and critical diagnostic and pharmacogenomic treatment targets for chronic pain conditions. Med Hypotheses. 2009;72:14–22. doi: 10.1016/j.mehy.2008.07.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Blum K, Oscar-Berman M, Dinubile N, Giordano J, Braverman ER, et al. Coupling Genetic Addiction Risk Score (GARS) with Electrotherapy: Fighting Iatrogenic Opioid Dependence. J Addict Res Ther. 2013;4:1000163. doi: 10.4172/2155-6105.1000163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Blum K, Oscar-Berman M, Jacobs W, McLaughlin T, Gold MSJ, et al. Buprenorphine Response as a Function of Neurogenetic Polymorphic Antecedents: Can Dopamine Genes Affect Clinical Outcomes in Reward Deficiency Syndrome (RDS)? Addict Res Ther. 2014;5:1000185. doi: 10.4172/2155-6105.1000185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Blum K, Badgaiyan RD, Agan G, Fratantonio J, Simpatico T, et al. Molecular Genetic Testing in Reward Deficiency Syndrome (RDS): Facts and Fiction. Reward Defic Syndr. 2015;1:65–68. doi: 10.17756/jrds.2015-009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Blum K. neurogenetics and nutrigenomics of reward deficiency syndrome. In: Barh D, Blum K, Madigan M, editors. Omics-biomedical perspectives and applications. Boca Raton: 2012. pp. 535–576. [Google Scholar]
- 7.Blum K, Oscar-Berman M, Stuller E, Miller D, Giordano J, et al. Neurogenetics and nutrigenomics of neuro-nutrient therapy for reward deficiency syndrome (RDS): clinical ramifications as a function of molecular neurobiological mechanisms. J Addict Res Ther. 2012;3:139. doi: 10.4172/2155-6105.1000139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Conner BT, Hellemann GS, Ritchie TL, Noble EP. Genetic, personality, and environmental predictors of drug use in adolescents. J Subst Abus Treat. 2010;38:178–190. doi: 10.1016/j.jsat.2009.07.004. [DOI] [PubMed] [Google Scholar]
- 9.Comings DE, Blum K. Reward deficiency syndrome: genetic aspects of behavioral disorders. Prog Brain Res. 2000;126:325–341. doi: 10.1016/S0079-6123(00)26022-6. [DOI] [PubMed] [Google Scholar]
- 10.Gold MS, Redmond DE, Jr, Kleber HD. Clonidine blocks acute opiate-withdrawal symptoms. Lancet. 1978;2:599–602. doi: 10.1016/s0140-6736(78)92823-4. [DOI] [PubMed] [Google Scholar]
- 11.Blum K, Chen AL, Oscar-Berman M, Chen TJ, Lubar J, et al. Generational association studies of dopaminergic genes in reward deficiency syndrome (RDS) subjects: selecting appropriate phenotypes for reward dependence behaviors. Int J Environ Res Public Health. 2011;8:4425–4459. doi: 10.3390/ijerph8124425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Blum K, Chen ALC, Thanos PK, Febo M, Demetrovics Z, et al. Genetic addiction risk score (GARS)™, a predictor of vulnerability to opioid dependence. Front Biosci (Elite Ed) 2018;10:175–196. doi: 10.2741/e816. [DOI] [PubMed] [Google Scholar]
- 13.Gumbley SJ. Recovery in the 21st Century: From Shame to Strength. J Addict Nurs. 2016;27:143–147. doi: 10.1097/JAN.0000000000000125. [DOI] [PubMed] [Google Scholar]
- 14.Blum K, Thompson B, Demotrovics Z, Femino J, Giordano J, et al. The Molecular Neurobiology of Twelve Steps Program & amp; Fellowship: Connecting the Dots for Recovery. J Reward Defic Syndr. 2015;1:46–64. doi: 10.17756/jrds.2015-008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Flanagan O. The shame of addiction. Front Psychiatry. 2013;4:120. doi: 10.3389/fpsyt.2013.00120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Blum K, Thanos PK, Wang GJ, Febo M, Demetrovics Z, et al. The Food and Drug Addiction Epidemic: Targeting Dopamine Homeostasis. Curr Pharm Des. 2017 doi: 10.2174/1381612823666170823101713. [DOI] [PubMed] [Google Scholar]
- 17.Heller EA, Hamilton PJ, Burek DD, Lombroso SI, Peña CJ, et al. Targeted Epigenetic Remodeling of the Cdk5 Gene in Nucleus Accumbens Regulates Cocaine and Stress-Evoked Behavior. J Neurosci. 2016;36:4690–4697. doi: 10.1523/JNEUROSCI.0013-16.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Davies SM1, Rutledge CM, Davies TC. A family physician-counselor program for medical students. Fam Med. 1993;25:327–330. [PubMed] [Google Scholar]
- 19.https://www.cdc.gov/media/releases/2016/p1216-continuing-opioid-epidemic.html
- 20.https://luxuryrehab.co/3000-increase-in-demand-for-opioid
- 21.Noble EP. D2 dopamine receptor gene in psychiatric and neurologic disorders and its phenotypes. Am J Med Genet B Neuropsychiatr Genet. 2013;116B:103–125. doi: 10.1002/ajmg.b.10005. [DOI] [PubMed] [Google Scholar]
- 22.Lawford BR, Young RM, Rowell JA, Qualichefski J, Fletcher BH, et al. Bromocriptine in the treatment of alcoholics with the D2 dopamine receptor A1 allele. Nat Med. 1995;1:337–341. doi: 10.1038/nm0495-337. [DOI] [PubMed] [Google Scholar]
- 23.Blum K, Chen TJH, Chen LC, Rhoades P, Prihoda TJ, et al. Dopamine D2 Receptor Taq A1 allele predicts treatment compliance of LG839 in a subset analysis of pilot study in the Netherlands. Gene Ther Mol Biol. 2008;12:129–140. [Google Scholar]
- 24.Dahlgren A, Wargelius HL, Berglund KJ, Fahlke C, Blennow K, et al. Do alcohol-dependent individuals with DRD2 A1 allele have an increased risk of relapse? A pilot study. Alcohol Alcohol. 2011;46:509–513. doi: 10.1093/alcalc/agr045. [DOI] [PubMed] [Google Scholar]
- 25.Ritchie T, Noble EP. [3H] naloxone binding in the human brain: alcoholism and the TaqI A D2 dopamine receptor polymorphism. Brain Res. 1996;718:193–197. doi: 10.1016/0006-8993(96)00068-6. [DOI] [PubMed] [Google Scholar]
- 26.Gerra G, Somaini L, Leonardi C, Cortese E, Maremmani I, et al. Association between gene variants and response to buprenorphine maintenance treatment. Psychiatry Res. 2014;215:202–207. doi: 10.1016/j.psychres.2013.11.001. [DOI] [PubMed] [Google Scholar]
- 27.Barratt DT, Coller JK, Somogyi AA. Association between the DRD2 A1 allele and response to methadone and buprenorphine maintenance treatments. Am J Med Genet B Neuropsychiatr Genet. 2016;141B:323–331. doi: 10.1002/ajmg.b.30319. [DOI] [PubMed] [Google Scholar]
- 28.Blum K, Oscar-Berman M, Femino J, Waite RL, Benya L, et al. Withdrawal from Buprenorphine/Naloxone and Maintenance with a Natural Dopaminergic Agonist: A Cautionary Note. J Addict Res Ther. 2013:4. doi: 10.4172/2155-6105.1000146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Makhinson M, Gomez-Makhinson J. A successful treatment of buprenorphine withdrawal with the dopamine receptor agonist pramipexole. Am J Addict. 2014;23:475–477. doi: 10.1111/j.1521-0391.2014.12133.x. [DOI] [PubMed] [Google Scholar]
- 30.Willuhn I, Burgeno LM, Groblewski PA, Phillips PE. Excessive cocaine use results from decreased phasic dopamine signaling in the striatum. Nat Neurosci. 2014;17:704–709. doi: 10.1038/nn.3694. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Blum K, Chen AL, Chen TJ, Braverman ER, Reinking J, et al. Activation instead of blocking mesolimbic dopaminergic reward circuitry is a preferred modality in the long-term treatment of reward deficiency syndrome (RDS): a commentary. Theor Biol Med Model. 2008;5:24. doi: 10.1186/1742-4682-5-24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Boundy VA, Pacheco MA, Guan W. Molinoff Agonists and antagonists differentially regulate the high affinity state of the D2L receptor in human embryonic kidney 293 cells. Mol Pharmacol. 1995;48:956–964. [PubMed] [Google Scholar]
- 33.Hartz Sarah M, Pato Carlos N, Medeiros Helena, Cavazos-Rehg Patricia, Sobell Janet L, et al. Comorbidity of severe psychotic disorders with measures of substance use. JAMA Psychiatry. 2014;71:248–254. doi: 10.1001/jamapsychiatry.2013.3726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Woodcock E, Lundahl L, Burmeister M, Greenwald M. Functional mu opioid receptor polymorphism (OPRM1 A (118) G) associated with heroin use outcomes in Caucasian males: A pilot study. Am J Addict. 2015;24:329–335. doi: 10.1111/ajad.12187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ramerstorfer J, Furtmüller R, Vogel E, Huck S, Sieghart W, et al. The point mutation gamma 2F77I changes the potency and efficacy of benzodiazepine site ligands in different GABAA receptor subtypes. Eur J Pharmacol. 2010;636:18–27. doi: 10.1016/j.ejphar.2010.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mogil J, Marek P, Flodman P, Spence M, Sternberg W, et al. One or two genetic loci mediate high opiate analgesia in selectively bred mice. Pain. 1995;60:125–135. doi: 10.1016/0304-3959(94)00098-Y. [DOI] [PubMed] [Google Scholar]
- 37.Coller J, Barratt D, Dahlen K, Loennechen M, Somogyi AAA, et al. ABCB1 genetic variability and methadone dosage requirements in opioid-dependent individuals. Clin Pharmacol Ther. 2006;80:682–690. doi: 10.1016/j.clpt.2006.09.011. [DOI] [PubMed] [Google Scholar]
- 38.McClung C, Nestler E, Zachariou V. Regulation of gene expression by chronic morphine and morphine withdrawal in the locus coeruleus and ventral tegmental area. J Neurosci. 2005;25:6005–6015. doi: 10.1523/JNEUROSCI.0062-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lötsch J, Skarke C, Liefhold J, Geisslinger G. Genetic predictors of the clinical response to opioid analgesics: clinical utility and future perspectives. Clin Pharmacokinet. 2004;43:983–1013. doi: 10.2165/00003088-200443140-00003. [DOI] [PubMed] [Google Scholar]
- 40.Befort K. Interactions of the opioid and cannabinoid systems in reward: Insights from knockout studies. Front Pharmacol. 2015;6:6. doi: 10.3389/fphar.2015.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Lötsch J, Zimmermann M, Darimont J, Marx C, Dudziak R, et al. Does the A118G polymorphism at the mu-opioid receptor gene protect against morphine-6-glucuronide toxicity? Anesthesiology. 2008;97:814–819. doi: 10.1097/00000542-200210000-00011. [DOI] [PubMed] [Google Scholar]
- 42.Landau R, Kern C, Columb M, Smiley R, Blouin J, et al. Genetic variability of the mu-opioid receptor influences intrathecal fentanyl analgesia requirements in laboring women. Pain. 2008;139:5–14. doi: 10.1016/j.pain.2008.02.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nelson EC, Lynskey MT, Heath AC, Wray N, Agrawal A, et al. ANKK1, TTC12, and NCAM1 polymorphisms and heroin dependence: importance of considering drug exposure. JAMA Psychiatry. 2013;70:325–333. doi: 10.1001/jamapsychiatry.2013.282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wang H, Gao MX, Li L, Wang B, Hori N, et al. Isolation, expression, and characterization of the human ZCRB1 gene mapped to 12q12. Genomics. 2007;89:59–69. doi: 10.1016/j.ygeno.2006.07.009. [DOI] [PubMed] [Google Scholar]
- 45.Garzón J, Rodríguez-Muñoz M, López-Fando A, García-España A, Sánchez-Blázquez P, et al. RGSZ1 and GAIP regulate mu- but not delta-opioid receptors in mouse CNS: role in tachyphylaxis and acute tolerance. Neuropsychopharmacology. 2004;29:1091–1094. doi: 10.1038/sj.npp.1300408. [DOI] [PubMed] [Google Scholar]
- 46.Woodcock E, Lundahl L, Burmeister M. M Greenwald: Functional mu opioid receptor polymorphism (OPRM1 A (118) G) associated with heroin use outcomes in Caucasian males: A pilot study. Am J Addict. 2015;24:329–335. doi: 10.1111/ajad.12187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ramerstorfer J, Furtmüller R, Vogel E, Huck S, Sieghart W, et al. The point mutation gamma 2F77I changes the potency and efficacy of benzodiazepine site ligands in different GABAA receptor subtypes. Eur J Pharmacol. 2010;636:18–27. doi: 10.1016/j.ejphar.2010.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Befort K. Interactions of the opioid and cannabinoid systems in reward: Insights from knockout studies. Front Pharmacol. 2015;6:6. doi: 10.3389/fphar.2015.00006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Blum K, Hauser M, Fratantonio J, Badgaiyan RD. Molecular Genetic Testing in Pain and Addiction: Facts, Fiction and Clinical Utility. Addict Genet. 2015;2:1–5. doi: 10.1515/addge-2015-0001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Blum K, Noble EP, Sheridan PJ, Montgomery A, Ritchie T, et al. Allelic association of human dopamine D2 receptor gene in alcoholism. JAMA. 1990;263:2055–2060. [PubMed] [Google Scholar]
- 51.Blum K, Gold MS, Jacobs W, McCall WV, Febo M, et al. Neurogenetics of acute and chronic opiate/opioid abstinence: treating symptoms and the cause. Front Biosci (Landmark Ed) 2017;22:1247–1288. doi: 10.2741/4544. [DOI] [PubMed] [Google Scholar]
- 52.Blum K, Febo M, McLaughlin T, Cronjé FJ, Han D, et al. Hatching the behavioral addiction egg: Reward Deficiency Solution System (RDSS)™ as a function of dopaminergic neurogenetics and brain functional connectivity linking all addictions under a common rubric. J Behav Addict. 2014;3:149–56. doi: 10.1556/JBA.3.2014.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Palmer RH, Brick L, Nugent NR, Bidwell LC, McGeary JE, et al. Examining the role of common genetic variants on alcohol, tobacco, cannabis and illicit drug dependence: genetics of vulnerability to drug dependence. Addiction. 2015;110:530–537. doi: 10.1111/add.12815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Blum K, Wood RC, Braverman ER, Chen TJ, Sheridan PJ, et al. The D2 dopamine receptor gene as a predictor of compulsive disease: Bayes’ theorem. Funct Neurol. 1995;10:37–44. [PubMed] [Google Scholar]
- 55.Ducci F, Goldman D. The genetic basis of addictive disorders. Psychiatr Clin North Am. 2012;35:495–519. doi: 10.1016/j.psc.2012.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Vereczkei Zsolt Demetrovics, Szekely Anna, Sarkozy Peter, et al. Peter Antal, Multivariate Analysis of Dopaminergic Gene Variants as Risk Factors of Heroin Dependence Andrea. Plos One. 2013;8:66592. doi: 10.1371/journal.pone.0066592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Xu K, Lichtermann D, Lipsky RH, Franke P, Liu X, et al. Association of specific haplotypes of D2 dopamine receptor gene with vulnerability to heroin dependence in 2 distinct populations. JAMA Psychiatry. 2004;61:597–606. doi: 10.1001/archpsyc.61.6.597. [DOI] [PubMed] [Google Scholar]
- 58.Moeller SJ, Parvaz MA, Shumay E, Beebe-Wang N, Konova AB, et al. Gene x abstinence effects on drug cue reactivity in addiction: multimodal evidence. J Neurosci. 2013;33:10027–10036. doi: 10.1523/JNEUROSCI.0695-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Chapman EC, Ettienne E, Ofoegbu Clarke M, Dunston GM, et al. The Impact of Pharmacogenomic Testing on Buprenorphine Dosing for Opioid Use Disorder (OUD) Management in an African-American Population. 2nd ASRDS Conference; Baltimore. 2017. [Google Scholar]