

Abstract
Water availability is a major environmental challenge to a variety of terrestrial organisms. In insects, desiccation tolerance varies predictably over spatial and temporal scales and is an important physiological determinant of fitness in natural populations. Here, we examine the dynamics of desiccation tolerance in North American populations of Drosophila melanogaster using: 1) natural populations sampled across latitudes and seasons; 2) experimental evolution in field mesocosms over seasonal time; 3) genome-wide associations to identify SNPs/genes associated with variation for desiccation tolerance; and 4) subsequent analysis of patterns of clinal/seasonal enrichment in existing pooled sequencing data of populations sampled in both North America and Australia. A cline in desiccation tolerance was observed, for which tolerance exhibited a positive association with latitude; tolerance also varied predictably with culture temperature, demonstrating a significant degree of thermal plasticity. Desiccation tolerance evolved rapidly in field mesocosms, although only males showed differences in desiccation tolerance between spring and autumn collections from natural populations. Water loss rates did not vary significantly among latitudinal or seasonal populations; however, changes in metabolic rates during prolonged exposure to dry conditions are consistent with increased tolerance in higher latitude populations. Genome wide associations in a panel of inbred lines identified twenty-five SNPs in twenty-one loci associated with sex-averaged desiccation tolerance, but there is no robust signal of spatially varying selection on genes associated with desiccation tolerance. Together, our results suggest that desiccation tolerance is a complex and important fitness component that evolves rapidly and predictably in natural populations.
Keywords: desiccation tolerance, Drosophila, cline, season, plasticity
Introduction
Insects inhabit a wide range of habitats in which water is often scarce and desiccation represents a major physiological challenge. Insects are particularly vulnerable to water related challenges, due to their small size and thus large surface area to volume ratio (Edney 1977; Hadley 1994; Gibbs & Rajpurohit 2010). Environmental stressors such as desiccation are highly variable among natural habitats, and often vary predictably with such features as latitude, altitude, and season (Endler 1977; Hoffmann & Parsons 1991; Parkash et al. 2005; Parkash et al. 2009). Patterns of variation across these gradients (Telonis-Scott et al. 2011), particularly when genetic/genomic and phenotypic variation are integrated, offer a means to address the extent to which divergence in physiological traits is affected by natural selection and may represent an adaptive response to environmental heterogeneity (Barton 1999; Whitlock & McCauley 1999).
In Drosophila, various species have adapted to diverse climatic conditions during their evolutionary history and thus constitute good models for the integrated study of physiological adaptation and population ecology (Parsons 1983; Lemeunier et al. 1986; Coyne et al. 1983). High levels of desiccation resistance are associated with adaptation to arid habitats (e.g., David et al. 1983; Hoffmann & Parsons 1991; Gibbs et al. 2003). Similarly, geographical variation for desiccation tolerance among populations has been widely observed (Griffiths et al. 2005; Matzkin et al. 2007; Matzkin et al., 2009; Rajpurohit et al. 2013a; Rajpurohit et al. 2013b; Rajpurohit & Nedved 2013; Rajpurohit et al. 2017b). These patterns suggest that desiccation tolerance is an important fitness component that varies predictably within and among taxa.
However, the genetic architecture of desiccation tolerance remains unresolved. In D. melanogaster, tolerance evolves rapidly in response to artificial selection in the laboratory (Gibbs et al. 1997; Bradley et al. 1999; Gefen et al. 2006; Telonis-Scott et al. 2016) and appears associated with reduced water loss rates (Telonis-Scott et al. 2006; Rajpurohit et al. 2016). Selection for increased tolerance results in differential gene expression (Sorensen et al. 2007) and significant allele frequency changes across hundreds of genes (Telonis-Scott et al. 2006, 2012), indicating a complex genetic architecture (Matzkin et al. 2009; Foley & Telonis-Scott 2011; Rajpurohit et al. 2013a). A genome-wide association study in a natural population recently derived from southern Australia identified a number of genetic variants associated with differences in desiccation tolerance, further demonstrating that desiccation tolerance is a highly polygenic trait (Telonis-Scott et al. 2016). However, it remains unclear whether the architecture of this trait is parallel between natural populations exposed to pronounced environmental heterogeneity at various spatial and temporal scales vs. laboratory-based populations subjected to strong artificial selection over shorter time scales.
The observed clines in desiccation tolerance suggest a robust association with climate and imply spatially variable selection on tolerance in natural populations. However, whether the observed phenotypic clines are mirrored by allele frequency clines at identified candidates (e.g., Telonis-Scott et al. 2016) is unknown. Furthermore, clines may be generated by processes other than selection (e.g., Caracristi & Schlöotterer 2003; Kao et al. 2015; Bergland et al. 2016), and the timescale over which clines are generated and maintained is often unclear. If variation in desiccation tolerance is driven by spatially varying selection mediated by climate, and may respond rapidly to climate change (Telonis-Scott et al. 2016), then tolerance might also vary predictably with seasonal variation in climatic parameters in temperate habitats. The results from artificial selection studies suggest that desiccation tolerance can evolve rapidly in the laboratory (<10 generations; Hoffmann & Parsons 1989); however, whether this occurs in natural populations is unknown.
Here, we examine some basic dynamics of desiccation tolerance in natural populations sampled from a range of temperate habitats in North America. We predicted that, as has been shown in Indian populations, desiccation tolerance would vary with latitude (Karan et al. 1998; Parkash et al. 2008; Rajpurohit et al. 2013b), and that such patterns among populations would reflect selection for increased tolerance at higher latitudes that are characterized by increased winter severity and length. We tested this by sampling populations across the latitudinal gradient in the eastern U.S. and examining genetically based variation in tolerance across a range of developmental temperatures. Similarly, we hypothesized that desiccation tolerance would be higher in populations sampled in the spring (following the winter season in which maintenance of water balance in overwintering adults might be an important fitness component) as compared to populations sampled in the fall. This hypothesis stems from recent observations that a variety of fitness-associated traits evolve rapidly in D. melanogaster over seasonal time (Behrman et al. 2015, 2018; Rajpurohit et al. 2017a), and was tested by examining seasonal change in desiccation tolerance for three mid-latitude populations sampled from the eastern U.S. In order to examine whether desiccation tolerance evolves rapidly outside the laboratory environment, we employed a field-based experimental evolution design to examine whether tolerance changes over the course of the agricultural growing season from spring to fall.
Prior work on the genetic basis of variance in desiccation tolerance (Foley & Telonis-Scott, 2011; Telonis-Scott et al. 2012; Telonis-Scott et al. 2016) have provided a wealth of information regarding the architecture of desiccation tolerance, as well as an identification of candidates for subsequent functional analysis (Griffin et al. 2017; Kang et al. 2016). Here, we extend this to include a genome-wide association study (GWAS) using the panel of inbred lines that constitute the Drosophila Genetic Reference Panel (DGRP; Mackay et al. 2012). We compare the variants associated with desiccation tolerance in the DGRP to those identified in Australian populations (Telonis-Scott et al. 2012, 2016) to examine parallelism across continents and datasets. Given the documented latitudinal clines for desiccation tolerance and the inference that such patterns are generated by local adaptation to climatic variables, the simple prediction is that the genes underlying variation in desiccation tolerance will exhibit latitudinal clines in allele frequency and signatures of spatially varying selection. We test this prediction by examining whether the SNPs associated with variation in desiccation tolerance in the DGRP exhibit evidence of spatially varying selection, using existing population-level PoolSeq data (Kolackzowski et al. 2011; Bergland et al. 2014; Telonis-Scott et al. 2016).
Material & Methods
1. Patterns of desiccation tolerance in natural, experimental, and inbred populations
Spatiotemporal variation.
Six natural populations of D. melanogaster were collected from fruit orchards located along the east coast of the United States by a combination of aspiration and baiting/sweeping with aerial nets (see Fig. 1). Gravid females were immediately sorted into isofemale lines in the field; once the resulting progeny eclosed, lines were typed to species. Approximately 150 isofemale D. melanogaster lines were collected from each locale. Populations sampled were: Bowdoin, Maine (ME; 44.03N, 73.20W); Media, Pennsylvania (PA; 40.04N, 76.30W); Charlottesville, Virginia (VA; 38.03N, 78.48W); Athens, Georgia (GA; 32.84N, 83.66W); Jacksonville, Florida (JFL) 30.33N, 81.66W)); and Homestead, Florida (HFL) 25.46N, 80.45W). Independent seasonal collections were done in Media, Pennsylvania (PA), Lancaster, Massachusetts (MA); 42.455N, 71.67W)) and Charlottesville, Virginia (VA) orchards in June and November of 2012 (N = 6 populations). Long-term maintenance of all populations was done using standard, discrete generational culture of isofemale lines at 24 °C, 12:12 light:dark photoperiod, with a generation time of 21d. Flies were maintained on cornmeal-molasses-agar media; prior to use in experiments, density was standardized at 40–50 eggs per vial. All desiccation tolerance assays were conducted on individual isofemale lines from the latitudinal and seasonal collections (see below).
Figure 1:
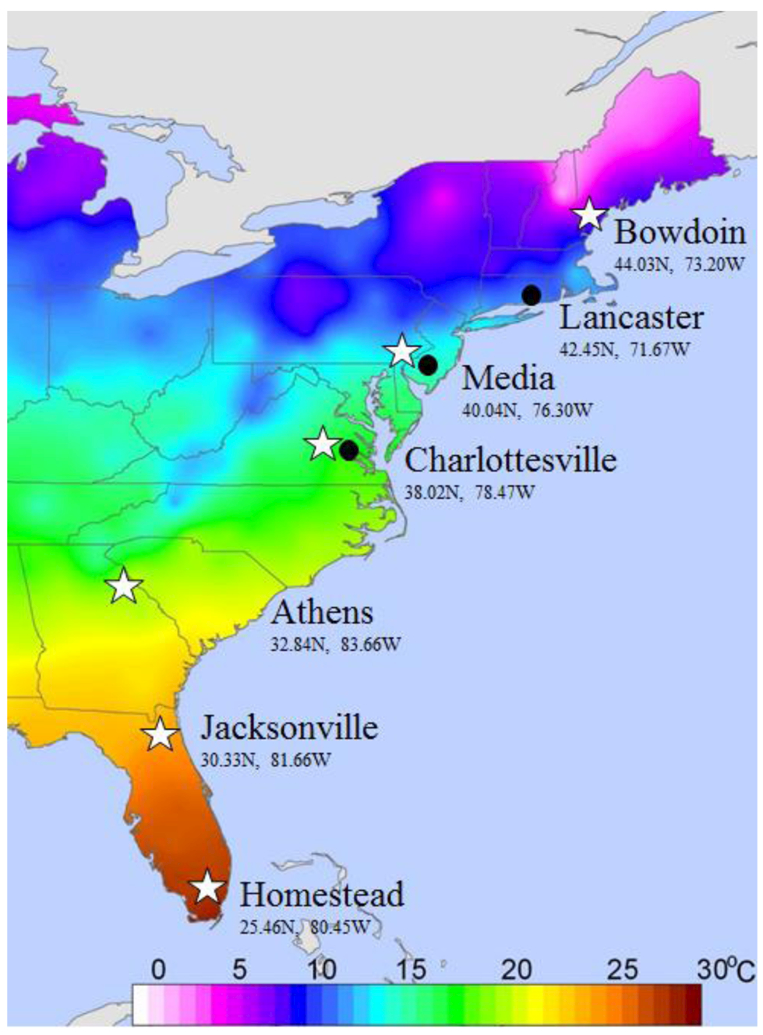
Temperature map of the east coast of the U.S. showing the populations of D. melanogaster that were collected and assayed. Collection sites for the latitudinal & seasonal populations are shown in white stars and black dots respectively. Bowdoin, Maine (ME); Lancaster, Massachusetts (MA); Media, Pennsylvania (PA); Charlottesville, Virginia (VA); Athens, Georgia (GA); Jacksonville, Florida (JFL), Homestead, Florida (HFL).
Geographic clines in fitness traits and their interactions with climatic variables are well known (e.g., Rajpurohit & Nedved 2013). To explore these interactions, we examined the relationship between desiccation tolerance and various climatic parameters (temperature and relative humidity) associated with the origin of the sampled populations. As the investigated populations encounter different temperature and humidity conditions in their natural habitats, we performed a multiple regression analysis of trait values as a simultaneous function of Tmin, Tmax, Tave, RHmin, RHmax and RHave of the origin of populations. Climatic data were obtained from the National Oceanic and Atmospheric Administration (NOAA; www.ncdc.noaa.gov). We used 30-year averages (1980–2010) for all the climatic parameters (Table S1). Statistical analyses were conducted in JMP v12 (SAS Institute, Cary, NC) and Statistica (Statsoft Inc., Release 7.0, Tulsa, OK, USA). We did not perform a sequential regression analysis of climatic factors on desiccation tolerance phenotype, as we did not observe any clear relationship between environmental variables and desiccation resistance or plasticity in tolerance.
Thermal plasticity.
To examine thermal plasticity for desiccation tolerance in the latitudinal collections, we primarily examined the effects of developmental temperature, where populations were cultured from egg to adult at three different temperatures that reflect a standardized culture range from cool to control to warm (18 °C, 25 °C and 29 °C; all at 12L:12D). We also examined desiccation tolerance in adult females that were exposed to environmental conditions that can elicit reproductive dormancy (11˚C, 9L:15D; Saunders et al. 1989). This treatment was included for two reasons. First, reproductive dormancy is associated with overwintering physiology, including an increase in desiccation tolerance (e.g., Denlinger 2002); second, we wished to examine one aspect of short-term, physiological plasticity for desiccation tolerance in the adult stage only.
For developmental thermal plasticity, flies from each line (N = 15–25) from each of the six geographic population were allowed to oviposit for 2–3h at 25 °C in successive culture vials; egg density was standardized at 40–50 eggs per vial by manual removal. Replicate vials (N = 30) of each line were then randomly assigned to one of the three temperature treatments (10 replicate vials per line per population per temperature) and cultured in Percival 136VL incubators. For the dormancy treatment, freshly eclosed adult females from the density controlled cultures at 25˚C were collected in groups of ten from each isofemale line and exposed to low temperature and short-day photoperiods (11˚C, 10L:14D) for three weeks. To further examine spatiotemporal variations, thermal plasticity and their interactions with climatic variables, we performed a principal component analysis (PCA) in XLSTAT using an unconstrained ordination technique.
For the latitudinal, seasonal, and plasticity analyses, four-to-five day old virgin flies from each isofemale line were transferred to empty vials (narrow plastic Drosophila culture vials, Genesee Scientific) in groups of ten and restricted to the lower half of the vials by a foam stopper. Silica gel (5g) was then added above the stopper to maintain low humidity (RH ~ 3%), and the vial was sealed with Parafilm™ (see Gibbs et al. 1997). Mortality was recorded at hourly intervals until all flies were dead. Desiccation tolerance assays were conducted using individual isofemale lines under common garden laboratory conditions of 25˚C, 12L:12D in Percival I36VL incubators. Data on latitudinal, seasonal, and thermal plasticity variation in desiccation tolerance were analyzed using a mixed model, nested ANOVA in JMPv.12 (SAS Institute, Cary, NC), where the response variable was time until death. Isofemale lines were nested within populations and considered a random factor; population, temperature and sex were treated as fixed factors. LT50 means were calculated using sigmoidal logit regression in Origin 9 (Zar 2010) and were used for visual presentation.
Experimental evolution in the field.
To examine whether desiccation tolerance evolves rapidly in the field in response to changing environmental conditions over seasonal time, we established 10 experimental mesocosms at a field site in Philadelphia, PA, U.S. Each mesocosm was an 8m3 outdoor insect rearing enclosure (Bioquip Products, Gardena, CA) surrounding a mature (dwarf) peach tree (Fig. S1). Each mesocosm was seeded with 1000 individuals (500 males, 500 females) derived from a collection made in 2012 from the same PA orchard as described above. 500ml of standard cornmeal-molasses medium in loaf pans was placed in each enclosure every other day for the duration of the experiment (July 13 – November 1, 2014). Flies were allowed to oviposit on the fresh food for a period of 2d, after which point the food was sealed with cotton rope/mesh and larvae allowed to develop; upon eclosion, adults were released into the mesocosms. Thus, the experimental populations were cultured under a natural regime of overlapping generations. Temperature and relative humidity in all experimental cages were recorded using HOBO U23 Pro v2 data loggers (Onset Computer Corp., Bourne, MA, USA). Further experimental details are provided in Rajpurohit et al. (2017a). Over the course of the experiment, samples were taken at four time points for phenotypic characterizations: July 25, August 19, September 11, and October 10, 2014. Approximately 2000 embryos were collected from each of the 10 cages at each time point, brought back to the laboratory, and subsequently passed through two additional generations of common garden, density-controlled culture (25˚C, 12L:12D, 40–50 eggs/vial). In the F3 generation subsequent to each field collection (10 cages x 4 time points), three replicate vials of 10 individuals per sex were assayed and analyzed for desiccation tolerance at 5d of age according to the methods described above. Data were analyzed with a mixed model ANOVA with cage as a random factor, replicate nested within cage as a random factor, and time point treated as an ordinal predictor.
Inbred panel (DGRP).
The panel of DGRP inbred lines (Mackay et al. 2012), derived from a single population in Raleigh, NC, USA, were obtained from the Bloomington Drosophila Stock Center and maintained in the lab on yeast-cornmeal-sucrose medium at 25˚C. For assays, 4–6 day old flies were sorted by sex and held on fresh media for two days. Assays were conducted on ten replicate groups of five flies per sex for each of the independent lines. We analyzed 175 DGRP lines in all.
Desiccation assays were performed in blocks of ~30 lines. Because of the time required to initiate desiccation stress for all lines and to count dead flies, we recorded the exact time when desiccant was added to assay vials and the exact times when each line was checked. Flies that died before the first survival check were assumed to have been injured by handling or other stress and were not included in the data analysis. Any flies dying between two checks were assumed to have died midway between them. To assess potential variation among blocks associated with minor differences in food, incubator temperature, etc., two lines (RAL-315 and RAL-324) were assayed in each block as internal controls. Variation among blocks in desiccation resistance of each line was <10%. For lines assayed multiple times, only data from the first block were included in the overall data analysis. Data are presented as line means ± s.e. The presence of Wolbachia and cosmopolitan chromosomal inversions, both of which can affect a variety of physiological and life history traits (e.g., Brummel et al. 2004; Umina et al. 2005), was included in the examination of among-line variation in desiccation tolerance in the DGRP panel. Line-specific data on inversion and infection status were obtained from Huang et al. (2014).
2. Respirometry
Due to logistical limitations, for respirometry measurements (water loss rate and metabolic rate) we utilized population cages as opposed to individual isofemale lines. We created two population cages for each of the spatial (6 populations x 2 replicates) and temporal (2 seasons x 3 populations x 2 replicates) collections (see above). Each cage was created using independent sets of 25 isofemale lines by releasing ten mated females from each line into 12×12×12 inch insect enclosures (Live Monarch Foundation, Boca Raton, Florida, USA). These lines were maintained in mass culture and allowed to outcross for 5 generations; subsequently, samples were collected for the phenotypic assays described below.
Virgins were collected within 3h of eclosion, separated by sex into sets of 15 individuals, and placed in fresh food vials. A total of 24 sets were assayed for the latitudinal extremes of ME and HFL (2 geographic regions × 2 sexes × 2 replicate populations × 3 experimental replicates), and 24 for seasonal comparisons in Pennsylvania (2 seasons × 2 sexes × 2 biological replicate cages × 3 experimental replicates). Respirometry was carried out at 4–6 days post-eclosion. Flies were transferred directly from their food vials to a 4 mL glass metabolic chamber with aluminum stoppers, which was covered with a black cardboard sleeve to reduce activity in the chamber. Flow-through respirometry at 25°C was carried out using two channels of a flow multiplexer (RM-8, Sable Systems International, Las Vegas, NV, USA), where dry, CO2-free air was supplied to the chambers at 50 mL·min−1 using factory-calibrated mass flow controllers (MC-500 sccm; Alicat Scientific, Tuscon, AZ, USA), and excurrent air from the measured chamber was passed through a LI-7000 CO2/water vapor dual analyzer (Li-Cor Biosciences, Lincoln NE, USA). Prior to measurement the flies were acclimated to the experimental chambers and air flow for 15 minutes during which flies from the alternative chamber were measured. Identical empty chambers were used for baselining. Analyzer voltage output was recorded, stored and analyzed using UI-2 data acquisition interphase and Expedata software (Sable Systems International). Recording rate was set to 1 Hz and only data from the last 5 minutes of each 15-min run were averaged for analysis.
We used an additional experimental approach to compare the HFL and ME populations for which significant differences in desiccation resistance were found (see results). It was previously reported that water loss rates of D. melanogaster under similar experimental conditions stabilize only after >2h of exposure (Gibbs et al. 1997), and therefore we carried out respirometry on additional sets of flies (2 geographic regions × 2 sexes × 2 replicate populations × 6 experimental replicates) that were randomly allocated to six multiplexer channels, with a seventh used for baselining. The measurement sequence of 20 min measurements was as follows: baselining, 3 experimental chambers, baselining, three additional experimental chambers and finally baselining again for a total file recording of 3 h, which was immediately followed by another recording at the same sequence. As each set of flies was added to the respirometry setup 20 min prior to the initial measurement, and only data from the last 10 minutes were analyzed, the flies were assayed 30 min and following additional 3h during which the flies were exposed to experimental temperature and dry air flow conditions. Respirometry data were analyzed with ANOVA when body mass did not vary significantly between experimental groups. On the rare occasion (see results) that body mass did vary, data were analyzed using ANCOVA with body mass as a covariate.
3. GWAS and signatures of clinality
We performed genome-wide association analysis on desiccation tolerance using 175 assayed inbred lines from the Drosophila Genetic Reference Panel (DGRP). Phenotypic line means were uploaded to the DGRP analysis website (http://dgrp2.gnets.ncsu.edu) for genome-wide association analysis following established methods (Mackay et al. 2012; Huang et al. 2014). From this analysis, twenty-five SNPs were associated with sex-averaged desiccation tolerance below the nominal p-value of 1e−5 (Fig. S2; see Table S4). These SNPs were annotated as being in or nearby 20 genes and one non-coding region (Table S4, Table S5); the identified SNP located in the non-coding locus (CR43681) was omitted from further analysis. Visual analysis of the qq-plot (Fig. S2) suggests that the observed p-value distribution is not over-inflated.
We tested whether the set of 20 identified genes were more likely to show signals of spatially varying selection than expected relative to the rest of the genome. We examined patterns of spatially varying selection using whole genome resequencing of populations sampled along the east coasts of North America (Bergland et al. 2014; Bergland et al. 2016) and Australia (Kolaczkowski et al. 2011) following a method outlined in Daub et al. (2013). This method tests whether gene sets identified a priori show stronger signals of spatially varying selection than sets of control genes. To perform this analysis, we first estimated genetic differentiation at approximately 500,000 common SNPs with average minor allele frequency greater than 5% (Bergland et al. 2014) among populations of D. melanogaster along latitudinal transects in North America or Australia using the TFLK statistic (Bonhomme et al. 2010; Bergland et al. 2016). The TFLK statistic is a modified version of the classic Lewontin-Krakauer test for FST outliers (Lewontin & Krakauer 1973) that incorporates certain aspects of population structure and has been shown to have a low false positive rate when sampled populations result from secondary contact, as is likely the case for North American and Australian populations of D. melanogaster (Caracristi & Schlötterer 2003; Duchen et al. 2013; Kao et al. 2015; Bergland et al. 2016). TFLK values were z-transformed and, following Daub et al. (2013), we refer to these transformed TFLK values as z. For each gene in the genome, we calculated the maximum z value, z(g), by considering all SNPs within 10Kb of the beginning and end of the gene. z(g) was normalized by gene length (hereafter zst(g)) by binning all genes with approximately equal length on a log2 scale following equations (1) and (2) of Daub et al. (2013). Next, we generated 5000 sets of control genes matched to the target set associated with desiccation tolerance. These control sets were matched by chromosome, inversion status at the large cosmopolitan inversions that segregate on each chromosome (Corbett-Detig & Hartl 2012), recombination rate (Comeron et al. 2012), and SNP density surrounding each gene, and the number of genes that are wholly or partially within the ±10Kb window surrounding each gene. For each target gene, we calculated the probability that its zst(g) was greater than its specific control genes and report one minus this probability as the boot-strap p-value for that gene. In addition, following Daub et al. (2013), for target and control gene sets we calculated the sum of zst(g), SUMSTAT, and estimated the probability that SUMSTATtarget is greater than SUMSTATcontrol.
Gene ontology enrichment analysis was performed using Gowinda (version 1.12). Because the number of sex-averaged desiccation resistance SNPs was so small, we also included SNPs with p-values below 10−5 for either males or females. This total of 103 candidate SNPs (Table S6) was tested against a background of 4,438,427 SNPs found within the DGRP populations (dgrp2.vcf). Gene annotations were derived by converting a General Feature Format (gff) file from Flybase (version r5.49.gff) into a Gene Transfer Format (gtf) file with the Gff2Gtf.py Python script from Gowinda. Gene ontology associations were downloaded from FuncAssociate 3.0. Gene identifiers between the gene annotation and gene ontology association files were synchronized with the SynchronizeGtfGeneIDs.py script from Gowinda. The gene aliases used for this step were extracted from the most recent gff release on Flybase (version r6.20) using the ExtractGeneAliasesFromGff.py script from Gowinda. Gowinda was set to perform 1e6 simulations, with the --gene-definition and --mode parameters set to ‘gene’.
Results
Spatiotemporal variation in desiccation tolerance
Across the six sampled geographic locales (Fig. 1), desiccation tolerance for both sexes increased positively with latitude (Fig. 2). These patterns were affected by culture temperature, demonstrating an inverse relationship between temperature and desiccation tolerance (Table 1; raw data are archived at Dryad DOI: http://datadryad.org/review?doi=doi:10.5061/dryad.9719r). The significant interaction terms of population by temperature and temperature by sex showed that patterns of thermal plasticity varied among populations and were distinct between the sexes (Table 1; Fig. 2). As culture temperature decreased, in general the magnitude of differentiation among the geographic populations increased. The pattern of population-specific developmental plasticity was qualitatively similar across sexes, but males exhibited a slightly more pronounced response to developmental temperature. Similarly, we observed a strong, positive correlation between desiccation tolerance and latitude in adult females that were exposed to environmental conditions that elicit reproductive dormancy (Fig. 2). This may be driven by geographic variation in the incidence of dormancy (Schmidt et al. 2005), but does suggest that desiccation tolerance may also exhibit short-term, physiological plasticity.
Figure 2:
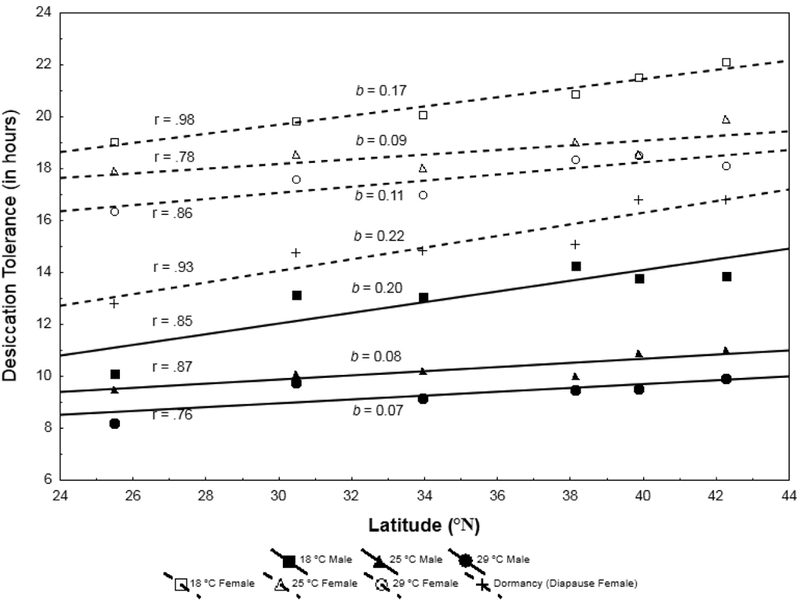
Data on desiccation tolerance for males and females of D. melanogaster from six geographical locations (see Fig. 1) at four different thermal conditions (developmental temperatures of 18 °C, 25 °C, and 29 °C; adult female exposure to dormancy-inducing conditions of 11˚C, 9L:15D). Data are presented as LT50 means. Males and females are denoted as triangles and rectangles, respectively. r = correlation; b = slope. Analysis of developmental temperature is presented in Table 1; for the reproductive dormancy treatment for adult females, latitude is a significant predictor of tolerance (F10,75, df=5, SS=269.99, p < 10−6).
Table 1:
ANOVA for desiccation tolerance of males and females from the six geographically distinct populations. Populations were cultured at three different growth temperatures (18 °C, 25 °C, & 29 °C).
| Parameters | SS | d. f. | MS | F | p |
|---|---|---|---|---|---|
| Population (P) | 532.2 | 5 | 106.4 | 28.74 | < 0.0001 |
| Temperature (T) | 1709.3 | 2 | 854.7 | 230.75 | < 0.0001 |
| Sex (S) | 14039.0 | 1 | 14039.0 | 3790.42 | < 0.0001 |
| P × T | 104.6 | 10 | 10.5 | 2.82 | 0.001866 |
| P × S | 27.5 | 5 | 5.5 | 1.49 | 0.191275 |
| T × S | 33.1 | 2 | 16.6 | 4.47 | 0.011721 |
| P × T × S | 64.3 | 10 | 6.4 | 1.74 | 0.068731 |
| Error | 3229.7 | 872 | 3.7 |
Regression analysis of desiccation tolerance with latitude and associated climatic variables (Tmin, Tmax, Tave, RHmin, RHmax and RHave) for both sexes at all three culture temperatures is given in Table 2. Interestingly, the temperature parameters exhibited stronger associations with desiccation tolerance than did those for relative humidity. In particular, minimum temperature (Tmin) was most strongly associated with the observed variation in desiccation tolerance. To further explore the collinearity of thermal plasticity in desiccation tolerance and various climatic variables, we performed a principal component analysis (Fig. S3). The first ordination axis explained 65.94% of variability and the second axis explained 27.14%. Temperature correlated best with the first axis.
Table 2:
Regression analysis of desiccation tolerance with geographical and climatic parameters. The first row gives correlations (r values), the second line the goodness of fit from ANOVA. Statistical significance is depicted by bold typeface.
| TEMPERATURE | LATITUDE | TMIN | TMAX | TAVE | RHMIN | RHMAX | RHAVE | |
|---|---|---|---|---|---|---|---|---|
| 18 °C MALE | r | 0.8582 | −0.8766 | −0.6603 | −0.7901 | −0.9236 | −0.1267 | −0.6808 |
| p | 0.02874 | 0.0219 | 0.15353 | 0.06148 | 0.00854 | 0.81104 | 0.13654 | |
| 18 °C FEMALE | r | 0.98115 | −0.9531 | −0.9651 | −0.977 | −0.6814 | −0.5908 | −0.7623 |
| p | 0.00053 | 0.00325 | 0.0018 | 0.00079 | 0.13612 | 0.2169 | 0.07804 | |
| 25 °C MALE | r | 0.87516 | −0.8453 | −0.8305 | −0.8544 | −0.6351 | −0.4225 | −0.5673 |
| p | 0.02241 | 0.03405 | 0.04064 | 0.03028 | 0.17541 | 0.40394 | 0.24034 | |
| 25 °C FEMALE | r | 0.78074 | −0.82 | −0.8713 | −0.8602 | −0.3712 | −0.3493 | −0.4461 |
| p | 0.06684 | 0.04567 | 0.02377 | 0.02797 | 0.46884 | 0.4974 | 0.37529 | |
| 29 °C MALE | r | 0.76591 | −0.8095 | −0.6357 | −0.742 | −0.7053 | 0.06338 | −0.3727 |
| p | 0.07579 | 0.05098 | 0.17491 | 0.09126 | 0.11748 | 0.90505 | 0.46687 | |
| 29 °C FEMALE | r | 0.86563 | −0.8094 | −0.7261 | −0.7852 | −0.7537 | −0.3719 | −0.7134 |
| p | 0.02587 | 0.05102 | 0.10226 | 0.06426 | 0.08353 | 0.46788 | 0.11146 |
Seasonal variation in desiccation tolerance for the three sampled seasonal populations (MA, PA, and VA) is depicted in Fig. 3. Again, we observed significant variation across the geographically distinct locales, and predictable differences in tolerance between the sexes (Table 3A). While the effect of season was not significant, the interaction terms incorporating season were significant or approached significance (Table 3A). To further investigate this, we also analyzed the sexes separately (Table 3B). Tolerance for females did not vary with season (Table 3B), whereas males sampled in the spring exhibited higher desiccation relative to males derived from the fall collections (Table 3B). This pattern of male seasonality was driven primarily by the PA populations, as can be seen in Fig. 3.
Figure 3:
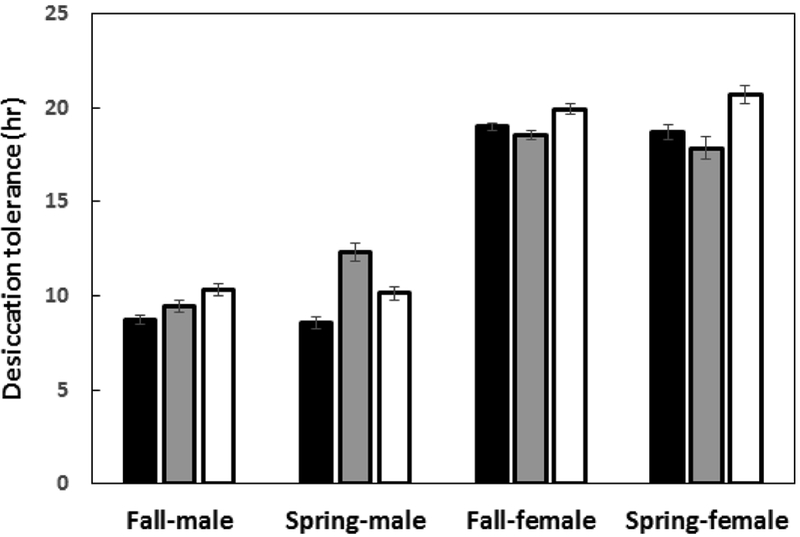
Data (mean ± s.e) on early and late season desiccation tolerance for males and females of D. melanogaster from three geographical locations (Lancaster, MA; Media, PA; Charlottesville, VA). Open, gray and black bars represent MA, PA and VA populations, respectively. The effect of season is significant for males only, and is driven by differences between spring and fall collections in the Media, PA population.
Table 3:
ANOVA for seasonal variations in desiccation tolerance of D. melanogaster adults from the three geographically distinct populations. Sex was included as a predictor in (A); and in (B), males and females were analyzed separately.
| (A) | |||||||||
| Parameters | SS | d.f. | MS | F | p | ||||
| Population (Pop) | 109.66 | 2 | 54.83 | 14.95 | <0.0001 | ||||
| Season (Sea) | 11.06 | 1 | 11.06 | 3.01 | 0.083603 | ||||
| Sex | 6188.14 | 1 | 6188.14 | 1686.67 | <0.0001 | ||||
| Pop*Sea | 21.49 | 2 | 10.74 | 2.93 | 0.055111 | ||||
| Pop*Sex | 133.73 | 2 | 66.87 | 18.23 | <0.0001 | ||||
| Sea*Sex | 14.78 | 1 | 14.78 | 4.03 | 0.045685 | ||||
| Pop*Sea*Sex | 67.72 | 2 | 33.86 | 9.23 | <0.0001 | ||||
| Error | 1034.62 | 282 | 3.67 | ||||||
| (B) | |||||||||
| Parameters | Male | Female | |||||||
| SS | d.f. | MS | F | p | SS | MS | F | p | |
| Population (Pop) | 130.11 | 2 | 65.05 | 34.53 | <0.0001 | 113.29 | 56.64 | 10.38 | <0.0001 |
| Season (Sea) | 25.71 | 1 | 25.71 | 13.64 | 0.000315 | 0.13 | 0.13 | 0.025 | 0.875397 |
| Pop*Sea | 75.65 | 2 | 37.83 | 20.08 | <0.0001 | 13.55 | 6.78 | 1.243 | 0.291793 |
| Error | 265.58 | 141 | 1.88 | 769.03 | 5.45 | ||||
Experimental evolution in the field
For both males and females, desiccation tolerance increased over time in the field mesocosms (Table 4, Fig. 4). As the populations were sampled and then passed through three generations of common garden culture in the laboratory, the observed patterns of increased tolerance over time are inferred to reflect change in the genetic composition of the experimental populations in response to environmental conditions experienced in the field. In females, desiccation tolerance increased rapidly between the first and second sampling time points (a period of 25d), remained constant over the next interval (23d), then exhibited a slight decrease at the last sampling time point (29d) (Table 4). In contrast, desiccation tolerance of males increased at a fairly constant rate; tolerance increased between time points one and two as well as between three and four (Table 4, Fig. 4). Thus, in response to field culture under heterogeneous environmental conditions, desiccation tolerance in outbred populations of D. melanogaster evolved rapidly over a period of approximately 90d (~ 9 generations).
Table 4:
Analysis of desiccation tolerance in field experimental mesocosms. The random variables cage and replicate (cage) are not presented.
| Source | DF | DF Den | F | P |
|---|---|---|---|---|
| Time Point (TP) | 3 | 2357 | 110.99 | <0.001 |
| Sex (S) | 1 | 2357 | 367.74 | <0.001 |
| TP × S | 3 | 2357 | 59.34 | <0.001 |
Figure 4:
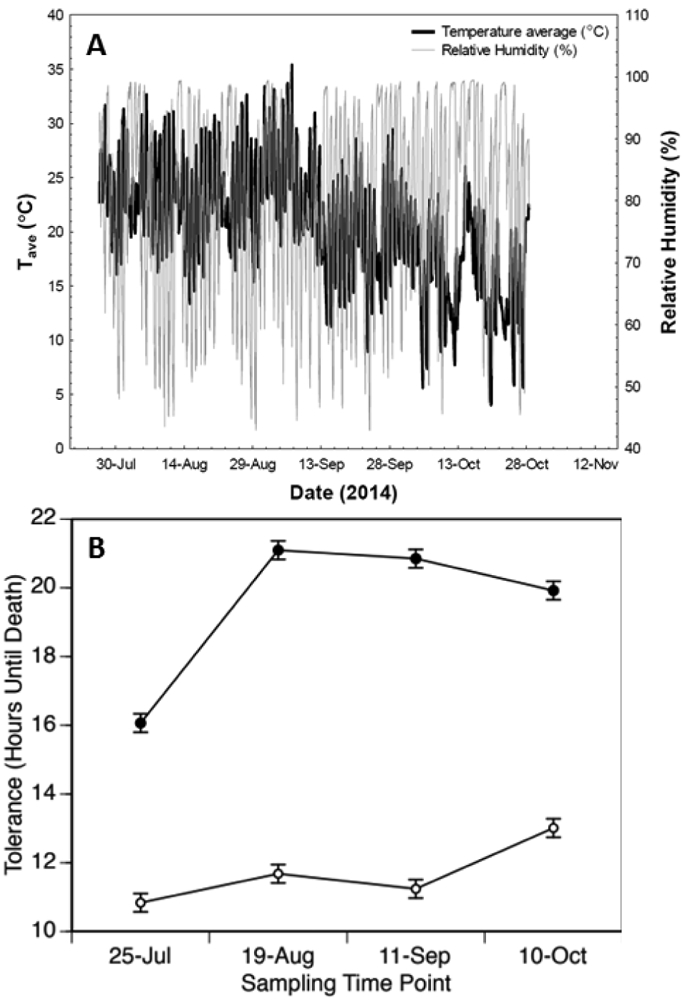
Rapid evolution of desiccation tolerance in field mesocosms. (A) Mean temperature and RH across the 10 replicate mesocosms are plotted over the duration of the experiment. (B) Mean (± s.e.m.) desiccation tolerance for females (closed symbols) and males (open symbols) at the four sampling time points in 2014. Tolerance increased for both females and males from July 25 to August 19 (F1,2357 = 339.91, p < 0.0001; F1,2358 = 9.63, p < 0.002, respectively). Over the period from September 11 to October 10, tolerance decreased in females (F1,2357 = 11.59, p < 0.001) and increased in males (F1,2357 = 41.44, p < 0.0001).
Water loss rate and metabolic rate in natural populations
With the exception of HFL and ME males, we did not find a significant within-sex difference in mean body mass among all our comparisons (data not shown). The mean body mass of male and female flies (ca. 0.7–0.8 and 1.1–1.2 mg, respectively) correlated with the well-documented higher desiccation resistance of females (e.g. Gibbs et al. 1997). We found no significant difference for either sex in mean CO2 or water loss rates (WLRs) between HFL and ME, or the seasonal collections from Pennsylvania (ANOVA, with replication as a random factor, p>0.05; see Table S2). When comparing HFL and ME males, no significant difference was found in mean CO2 (F1,7=1.61, p=0.24) or water loss rates (F1,7=2.76, p=0.21), even when accounting for differences in body size (ANCOVA, with body mass as a covariate).
The limitation of our original respirometry approach in detecting variation in WLRs prompted us to focus on the two most geographically-distant populations for which significant variation in desiccation resistance were detected (i.e. HFL vs. ME; see Fig. 2), using an alternative approach measuring WLR over a longer timeframe (see Methods). Still, no significant difference was found in the WLRs among female (ANOVA; F1,21=0.005, p=0.95) or male (F1,21=0.13, p=0.71) flies even when exposed to dry air flow for 3 h. No significant differences were found in CO2 between HFL and ME females (p=0.68 and 0.34 after 30 min and 3 h, respectively), but similar values for males after 30 minutes (p=0.46) were followed by significantly lower CO2 values for ME compared with HFL males (F1,21=4.34, p=0.049). Interestingly, when testing for temporal changes within sets of flies we found a significant increase in CO2 of HFL males from initial values (3.16±0.15 µL·fly−1·h−1) compared to those recorded after 3 h of exposure to desiccation (3.82±0.13 µL·fly−1·h−1) (paired t-test; t11=5.04, p<0.001). In contrast, values after 30min and 3h of desiccation did not vary significantly for ME male flies (t11=1.89, p=0.09) (Fig. 5A), which could reflect their enhanced desiccation tolerance (Fig. 2). Among female flies, a significant decrease in CO2 during exposure to desiccation was recorded for ME (t11=3.69, p=0.004), but not HFL populations (t11=0.40, p=0.70) (Fig. 5B).
Figure 5:
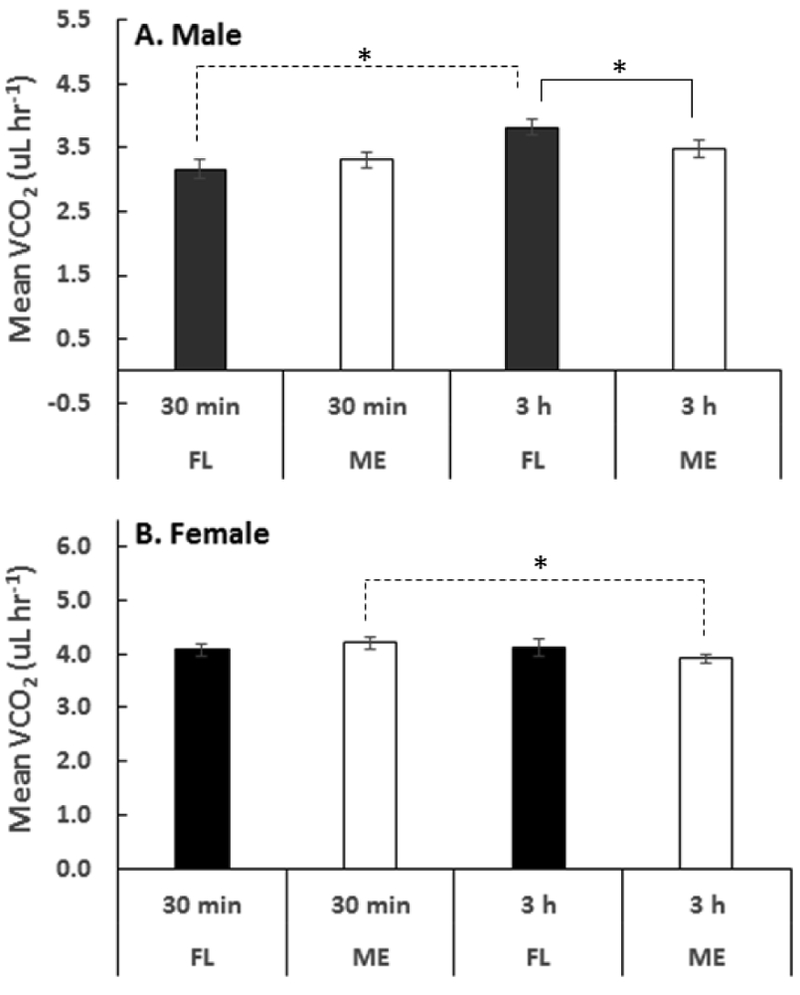
Carbon dioxide emission rates for males (A) and females (B) of D. melanogaster populations from Florida (HFL) and Maine (ME). Measurements were done at two time points (30 min and 3hr). Asterisks indicate a significant difference between measurements conducted at the two timepoints (α=0.05). Within population differences (for FL males and ME females) measured at two-time points (30min and 3hr) are shown with dashed line connections. The differences at the population level are shown with the solid lines connections. After 3h, FL males have higher values than ME males. In comparing FL males 30min to 3h treatment, there is a significant increase in VCO2. In females, ME flies show a decrease in VCO2 going from 30min to 3hr.
GWAS & GO Enrichment
We observed considerable genetic variability in desiccation tolerance among the 175 tested DGRP lines (Table S3). Line means are depicted in Fig. 6. Females survived longer than males under desiccating conditions, as expected. There was significant heterogeneity among lines with respect to sex-specific differences and relative ranking (Fig. 6). Neither Wolbachia infection status nor the presence of chromosomal inversions (Huang et al. 2014) had any effect on desiccation tolerance (Table 5, Table S3, Table S4).
Figure 6:
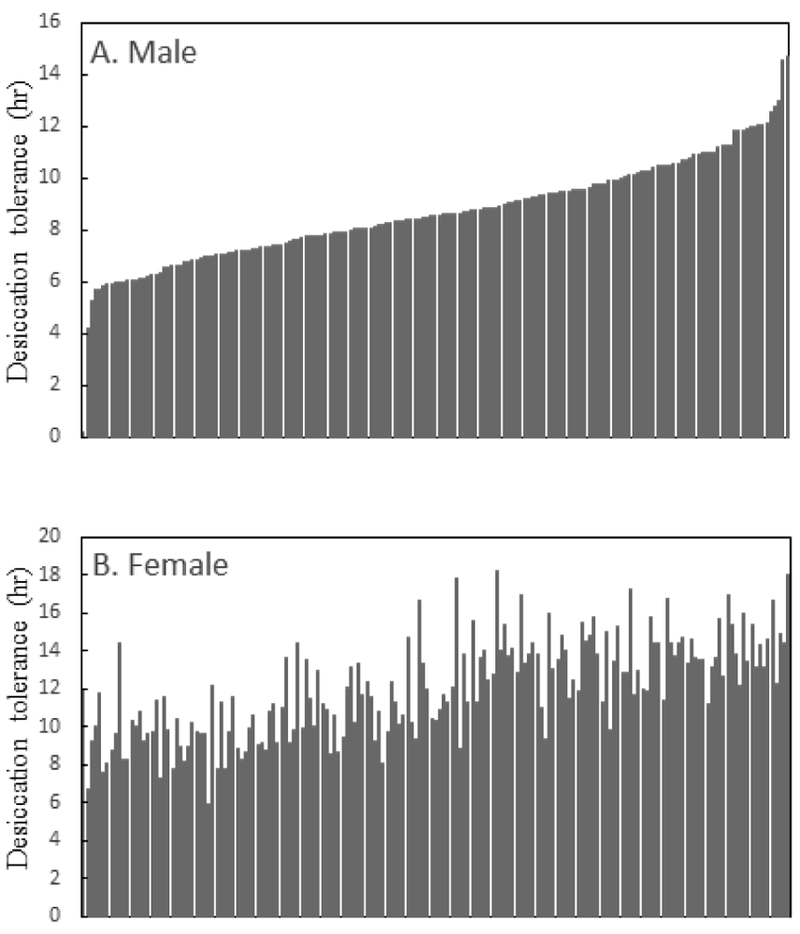
Status of desiccation tolerance (line means) in Drosophila Genetic Resource Panel. A total of 175 lines were considered for this association mapping. A considerable genetic variability in desiccation tolerance was observed across the lines. Males (A) survived shorter under desiccating conditions than females (B). Desiccation tolerance values are sorted in increasing order for males in panel A, and corresponding lines values for females are shown in panel B. The data show extensive within-line differences between the sexes, consistent with the differential response in the field mesocosms (Fig. 4).
Table 5:
ANOVA on desiccation tolerance in the DGRP lines. The Wolbachia infection and inversion status of DGRP lines have also been incorporated in the full model. Sex-specific analyses are presented in the supplementary material (Table S4).
| Parameters | SS | d. f. | MS | F | p |
|---|---|---|---|---|---|
| Sex | 548.29 | 1 | 548.29 | 114.702 | 10−6 |
| Wolbachia | 0.08 | 1 | 0.08 | 0.016 | 0.899487 |
| Inversions | 7.27 | 1 | 7.27 | 1.522 | 0.218271 |
| Sex*Wolbachia | 0.05 | 1 | 0.05 | 0.011 | 0.917563 |
| Sex*Inversions | 0.05 | 1 | 0.05 | 0.011 | 0.915927 |
| Wolbachia*Inversions | 1.14 | 1 | 1.14 | 0.239 | 0.625584 |
| Sex*Wolbachia*Inversions | 1.22 | 1 | 1.22 | 0.254 | 0.614309 |
| Error | 1510.53 | 316 | 4.78 |
Using among line variation in the standard mapping pipeline (Mackay et al. 2012), we identified 25 SNPs that were significantly associated with desiccation tolerance; these SNPs were associated with 20 genes and 1 non-coding locus (Table S5, Table S6). Gene ontology (GO) enrichment analysis placed these 20 genes in various GO biological categories that broadly included metabolism, signaling, cuticle development and transport. However, no enrichment for any biological category was observed (Eden et al. 2009); this was expected, given the small number of genes identified. We did not observe any overlap at the gene level between our association study using the DGRP panel and two studies conducted on D. melanogaster populations from Australia (see Table 1 in Telonis-Scott et al. 2016; see also Telonis-Scott et al. 2012).
Clinal differentiation of SNPs associated with desiccation tolerance
Among North American populations, the genes we identified as being associated with desiccation tolerance in the DGRP did not show increased signatures of spatially varying selection relative to the rest of the genome (Fig. 7A; p=0.769). Although there has not been any latitudinal cline reported for desiccation tolerance in Australian D. melanogaster (Hoffmann et al. 2001; Hoffmann et al. 2005), we observed an indication of increased spatial genetic differentiation at the loci identified in the DGRP (Fig. 7C; p=0.0926) amongst Australian populations. This prompted us to examine per gene zst(g) values relative to sets of genomic matched control genes. Several of the candidate genes (Whamy, CG18302, CG5953) for desiccation tolerance had relatively high zst(g) values, indicated by low bootstrap p-values, in the Australian populations (Fig. 7D). However, none of bootstrap p-values for these genes fell below the Bonferroni correction threshold at α < 0.05.
Figure 7:
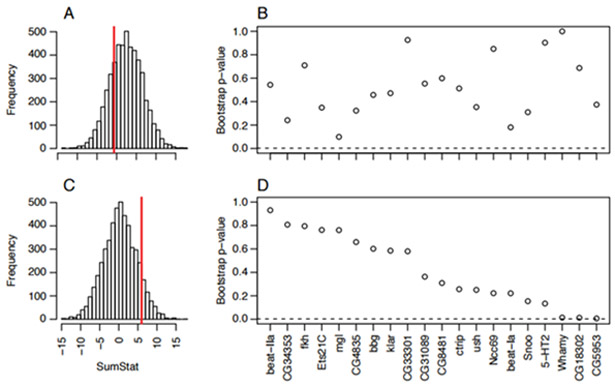
Signals of clinality among North American and Australian populations at genes associated with desiccation tolerance. (A) and (C) represent SumStat scores for 5000 sets of control genes (distribution) and observed SumStat score (dashed red line) for genes associated with desiccation tolerance among populations sampled along the east coast of North America or Australia, respectively. (B) and (D) represent per-gene probabilities that the observed zst(g) values are bigger than expected by chance for North American and Australian populations, respectively. See Materials and Methods for a description of the SumStat score.
Discussion
Desiccation resistance in insects can involve one or more physiological mechanisms, including: (1) increases in total body water content and/or in hemolymph volume (Folk et al. 2001); (2) increased dehydration tolerance (i.e., tolerance of body water loss before death) (Telonis-Scott et al. 2006); (3) reduction in the rate of water loss (Gibbs et al. 1997); or (4), any modification of behavior, including changes in activity level and associated metabolism. Body size plays a significant role in desiccation resistance: in 20 examined Drosophila species, desiccation tolerance varies five-fold among taxa and is associated with interspecific variation in size (Matzkin et al. 2009). Along the coast of eastern Australia, no cline was observed for desiccation tolerance in D. melanogaster (Hoffmann et al. 2001), whereas along the Indian latitudes a robust cline has been observed (Karan et al. 1998). Interestingly, a cline for body size (e.g., thorax and wing length) has been observed on both continents (James et al. 1995; Bhan et al. 2014). In North America, body size also varies clinally and increases with increasing latitude (e.g., Coyne & Milstead 1987). Here, we observed a significant but shallow cline in desiccation tolerance at normal assay temperatures of 25˚C (Fig. 2). While body size may play a role in desiccation tolerance, the relationship between variation in body size and desiccation tolerance among D. melanogaster populations appears complex (Rajpurohit et al. 2016).
Spatiotemporal patterns in desiccation tolerance
Our results, demonstrating a positive cline in desiccation tolerance for populations from the east coast of the U.S., are consistent with previous studies showing higher desiccation tolerance in temperate vs. tropical locales (Hoffmann & Harshman 1999). On the Indian subcontinent, this trend was clearly observed in multiple Drosophila species (Karan et al. 1998; Parkash et al. 2008; see also Rajpurohit et al. 2017b), where parallel clines for this trait have been observed (Rajpurohit et al. 2013b). In India, higher latitudes are characterized by lower temperature and lower humidity during winter, whereas low latitude, southern locations are warm and humid for most of the year. A meta-analysis approach concluded that the coefficient of variance in temperature was a major climatic component to support the observed parallel clines for desiccation tolerance in several Drosophila species on the Indian subcontinent (Rajpurohit et al. 2013b). Along the east coast of the U.S., more temperate populations experience greater environmental fluctuations associated with seasonality and harsher winter conditions. As adult overwintering is associated with long term exposure to a desiccating environment, we predicted that populations collected in the spring would be characterized by relatively higher desiccation tolerance, similar to patterns observed for other stress-related traits (Behrman et al. 2015). However, seasonal variation in relative humidity is not pronounced at the sites we sampled (e.g., Fig. S4). We did not observe robust differences in desiccation tolerance between early and late season collections of D. melanogaster from three temperate populations spanning 38 – 42˚N latitude; only males exhibited seasonal variation in tolerance, and this pattern was pronounced in only one of the three sampled geographic locales.
Although humidity may not vary significantly among seasons (Fig. S4), the regulation of water balance in natural populations may be driven primarily by temperature, or the interaction between temperature and humidity (Kingsolver 1979; Rajpurohit et al. 2013b). In the field mesocosms, relative humidity was highly variable over diurnal timescales, but average humidity was not distinct among sampling time points. Temperature, in contrast, varied predictably over time and was distinct across the sampled intervals from July to October. The patterns of desiccation tolerance observed in the field mesocosms mirror temperature profiles, with some deviation between males and females in the environmental tracking. The evolution of increased desiccation tolerance in the field experiment cannot be explained by response to humidity, as this was consistently higher in the field than under typical conditions of laboratory culture (Heerwaarden & Sgro 2014).
Climatic associations
It is generally assumed that desiccation tolerance is selected during hot and dry conditions, as heat and desiccation stresses often co-occur (Hoffmann & Parsons 1991). In natural habitats on the U.S. east coast, however, such is not the case. Variation in desiccation tolerance is most strongly associated with minimum temperature (and length of the winter season) of the geographic origin of our collections, suggesting that desiccation tolerance in temperate populations may be favored in environments characterized by cold and dry conditions (Leather et al. 1993). Cold and desiccation tolerance may also exhibit correlated responses (e.g., Bubliy & Loeschcke 2005; MacMillan et al. 2009). Thus, it remains unclear whether the latitudinal patterns we observed are driven primarily by selection directly on desiccation tolerance or may reflect an indirect response due to selection on a correlated trait. In contrast to the observed patterns for the geographic collections, our field mesocosm experimental data indicate a role of higher temperature, where desiccation tolerance increases as temperature increases. These contrasting patterns may reflect complex dynamics associated with various spatial and temporal scales, as well as the aforementioned correlations between desiccation tolerance and other traits. It would be of particular interest to generate long term, longitudinal data on desiccation tolerance in natural and experimental populations to further explore the associations between tolerance and various climatic parameters.
Thermal plasticity
We observed that decreasing culture temperature in the laboratory resulted in elevated desiccation tolerance. A recent study on the cold-adapted D. nepalensis from the western Himalayas found that flies grown at 15 °C demonstrate a twofold increase in body size, a greater degree of melanization, higher desiccation resistance, and increased hemolymph and carbohydrate content as compared to flies reared at 25 °C (Parkash et al. 2014). There is a strong possibility that D. melanogaster populations growing at lower temperatures may also exhibit these plastic responses that could subsequently affect desiccation tolerance. We did observe that variation among populations became exacerbated at lower temperatures, and was most distinct following exposure to dormancy inducing conditions. Thus, our data also suggest that patterns of plasticity in D. melanogaster may vary predictably among natural populations and habitats.
Geographic variation in metabolic rate
Desiccation resistance in Drosophila is associated with reduced water loss rates under both natural (Kalra et al. 2014) and laboratory conditions (Hoffmann & Harshman 1999). In contrast, evidence for other potential adaptive mechanisms is more equivocal. Higher body water content was reported for resistant populations in some studies (Gibbs et al. 1997; Chippindale et al. 1998; Folk et al. 2001; Gefen et al. 2006), but not in others (Hoffmann & Parsons 1993). The ability to tolerate dehydration has also been reported to vary between desiccation-selected populations and their controls in one study (Telonis-Scott et al. 2006), but not another (Gibbs et al. 1997). However, this discrepancy could simply reflect an inconsistency in the use of the term dehydration tolerance (Gibbs & Gefen 2009).
We found no evidence for variation in water loss rates that could explain the observed clinal variation in desiccation tolerance. Water-loss rates of the northernmost (ME) and southernmost (HFL) populations did not differ in either males or females. We also recorded similar metabolic rates, expressed as CO2 emission rates (CO2), for females from the two populations; however, higher values for severely desiccated HFL male flies suggest significant differences in the metabolic response of these populations to prolonged exposure to desiccating conditions. This is in agreement with both intraspecific (Hoffmann & Parsons 1993; Gibbs & Gefen 2009) and interspecific (Gibbs et al. 2003) reports that demonstrated desiccation resistance in Drosophila is correlated with reduced activity under stressful conditions; these results are also consistent with pronounced differences in central metabolism between populations at the geographic extremes of the U.S. east coast (e.g., Verrelli & Eanes 2001; Flowers et al. 2007; Lavington et al. 2014). It should be noted that increasing CO2 values could reflect a switch to carbohydrate catabolism under desiccation stress (Marron et al. 2003), independent of changing metabolic rates. However, we did not observe a similar response in females. Instead, the significant decrease in CO2 in the more resistant ME females as they settled to the dry metabolic chamber environment, and absence of this response for HFL females, suggests a difference in behavioral response. Both males and females in these populations do exhibit very distinct behavior in response to thermal variation as well (Rajpurohit & Schmidt 2016).
Variation in activity patterns under stressful conditions is likely to result in correlated differences in respiratory water losses. The similar WLRs reported here for HFL and ME flies can be explained by the considerably higher relative importance of cuticular water loss in insects (Chown 2002), and may suggest that flies across the experimental populations do not vary in their cuticular resistance to water loss. Nevertheless, while results in this study do not confirm an effect of activity level on WLRs and thus on desiccation-resistance, they could well reflect how stressful the exposure to experimental desiccation is to flies from the respective populations. If the more susceptible HFL flies have lower body water contents when hydrated compared with the more resistant ME flies (Chippindale et al. 1998; Folk et al. 2001; Gefen et al. 2006), then at similar WLRs the former would approach the minimum tolerable hydration state earlier, which could elicit an increase in activity levels as a result of attempts to seek more favorable conditions. In addition, a delayed escape response in the more resistant flies could indicate higher dehydration tolerance that would trigger an escape response at lower body water content. We speculate that geographically distinct populations exhibit distinct behaviors in response to desiccation exposure. This hypothesis needs to be tested in future work.
Candidate genes
The GWAS of the DGRP panel identified 20 genes associated with the sex-averaged among-line variance in desiccation tolerance. A Gowinda analysis using 103 SNPs with sex-averaged or sex-specific associations found no significant GO term enrichment (lowest FDR > 0.4). While there was no associated ontology enrichment, these identified genes do represent candidates for subsequent functional analysis. For example, 5-HT2 is a 5-hydroxytyramine (5-HT, serotonin) receptor, and serotonin is an important diuretic hormone in insects (Martini et al. 2004). Two additional candidates (mgl, CG4835) are functionally linked to chitin metabolism. Serotonin also mediates cuticular plasticization (Reynolds 1975); together these findings suggest that cuticular permeability may be an important factor affecting desiccation tolerance. Metabolic differences also appear to be important, as SNPs mapped to or close to three genes implicated in lipid metabolism (klar, CG18302, CG31089). However, we emphasize that the identification of these genes is based on GWA of a single panel of inbred lines and that the generation of a comprehensive list of genes associated with variance in desiccation tolerance would require further investigation as well as integration across datasets and experimental approaches.
Ecological genetics of desiccation tolerance
Relative to other fitness-associated traits in North American populations (e.g., body size, (Coyne & Milstead 1987); reproductive dormancy (Schmidt et al. 2005); cuticular hydrocarbons (Rajpurohit et al. 2017a); and thermal preference (Rajpurohit & Schmidt 2016), we observed a shallow cline for desiccation tolerance across the sampled latitudinal gradient in eastern North America. Our results also demonstrated pronounced patterns of plasticity in response to temperature, both in terms of developmental plasticity as well as adult acclimation and subsequent response. However, the observed patterns suggest that spatially varying selection may be less pronounced on this trait, both in comparison to other traits in North American populations as well as to desiccation tolerance in Australian (Telonis-Scott et al. 2006) and Indian (Karan et al. 1998; Parkash et al. 2008; Rajpurohit et al. 2013a) populations. Our analysis of clinal enrichment demonstrated that genes associated with desiccation tolerance are more strongly enriched for clinality in Australia than in North America. However, it is clear that there is no robust signal, above expectation, that genes associated with desiccation tolerance evidence patterns consistent with spatially variable selection. This may reflect the weak associations between latitude and desiccation tolerance, limitations in the identification of genes underlying variance in the trait, or a combination of the two.
Conclusion:
We observed a shallow cline for desiccation tolerance in populations sampled along the latitudinal gradient in the eastern U.S.; these patterns of variation among populations exhibited both developmental and adult plasticity, suggesting that further analysis of desiccation tolerance should be examined under a range of environmental conditions. Climatic analysis of this cline indicated that observed patterns of desiccation tolerance were most strongly associated with lower temperature conditions, suggesting that selection on this trait in temperate populations may be associated with response to desiccating conditions that co-occur with long-term exposure to reduced temperatures. However, our field experiment also demonstrated that desiccation tolerance increases over short timescales under field conditions of increasing temperature, suggesting that the evolutionary dynamics of desiccation tolerance may vary over distinct temporal and spatial scales. GWAS of the DGRP panel of inbred lines identified 20 genes and one non-protein coding locus associated with desiccation tolerance in North American populations; although underpowered and of perhaps limited scope in associations with patterns in natural populations or in the field, this does provide candidates for subsequent functional analysis and investigation. Overall, patterns of variation for desiccation tolerance appear complex and more fine scaled, mechanistic dissections should shed further light on the evolutionary and ecological dynamics of this trait in natural populations.
Supplementary Material
Schematic representation of field based mesocosm units. Each mesocosm was an 6m3 outdoor insect rearing enclosure surrounding a mature (dwarf) peach tree. A total of 10 cages were established for this experimental work.
qq-plot of genome wide association analysis of intersex averaged desiccation tolerance. Observed p-values are plotted along the y-axis and expected p-values along the x-axis. The solid red line depicts a 1:1 correspondence.
Multiple relationship between thermal plasticity of desiccation tolerance in D. melanogaster and various climatic variables analyzed by unconstrained ordination (Principal Component Analysis). The first ordination axis explained 65.94% of variability and the second axis explained 27.14%. Temperature correlates best with the first axis.
3D Surface plot (A) of Tave and RHave along the latitudes of east coast of the U.S. RHave = Distance weighted least square. The color scale relates to the RHave values distribution along the latitudes. Bar plot (B) is showing seasonal variations (summer vs. winter) in RHave for five latitudinal stations. Relative humidity data for winter and summer are averages for Nov-March and Apr-Oct months respectively. Averaging of relative humidity across an entire season may not be an appropriate level of resolution-relative humidity might vary (biologically) over a shorter timescale. Climatic data were obtained from National Oceanic and Atmospheric Administration (NOAA; www.ncdc.noaa.gov). We used 30-year averages (1980–2010) for all the climatic parameters.
Data on six climatic variables (Tmin: temperature minimum; Tmax: temperature maximum; Tave: temperature average; RHmin: relative humidity minimum; RHmax: relative humidity maximum; RHave: relative humidity average) for latitudinal and seasonal collections used in this study.
Mean (±s.e.m) CO2 and water vapor emission rates among the experimental fly populations (see methods for sample sixes and statistics).
Desiccation survival status in DGRP lines. The status of Wolbachia infection and cytological analysis of polymorphic inversions were updated from Huang et al. 2014 (see Supplemental data file S6 in Huang et al. 2014).
ANOVA on desiccation tolerance assays performed for DGRP lines. The analyses present sex separated and sexes averaged results for effects of Wolbachia infection and inversions status.
SNPs identified from the genome-wide association study for desiccation tolerance in D. melanogaster.
Genes containing SNPs associated with desiccation tolerance in DGRP lines. GO Biological Process information was obtained using DAVID 6.8 (Huang et al. 2014).
Acknowledgements
We would like to thank Kelly Dyer for providing isofemale lines collected from Athens, GA and Emily Behrman for providing seasonal collections from Media, PA. This research was funded by NIH R01GM100366 to P.S. and D.A.P. Experiments using the DGRP lines were supported by NIH R15-GM100395 to A.G. Gibbs and A.J. Andres. We thank many student volunteers for helping with these assays, particularly Michael Brewer, Jessica Gamble, Janilynn Gatpayat, Christensen Hardy, Christopher Hardy, Chelsea Koktavy, Christine Ladaga, Beena Patel, Timothy Saitta, Charmaine Tan, and Lawrence Wu.
Footnotes
Data accessibility
All raw data, including data not presented in the paper are archived at Dryad. DOI: https://doi.org/10.5061/dryad.9719r.
Rajpurohit S, Gefen E, Bergland A, Petrov DA, Gibbs A, Schmidt P. Data from: Spatiotemporal dynamics and genome-wide association analysis of desiccation tolerance in Drosophila melanogaster. Dryad Digital Repository. http://datadryad.org/review?doi=doi:10.5061/dryad.9719r
References
- Barton NH (1999) Clines in polygenic traits. Genetical Research, 74, 223–236. [DOI] [PubMed] [Google Scholar]
- Behrman EL, Watson SS, O’Brien KR, Heschel MS, Schmidt PS (2015) Seasonal variation in life history traits in two Drosophila species. Journal of Evolutionary Biology, 9, 1691–1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergland AO, Berhrman EL, O’Brien KR, Schmidt PS, Petrov DA (2014) Genomic evidence of rapid and stable adaptive oscillations over seasonal time scales in Drosophila. PLoS Genetics, 10, e1004775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bergland AO, Tobler R, Gonzalez J, Schmidt PS, Petrov DA (2016) Secondary contact has contributed to genome-wide patterns of clinal variation in North American and Australian populations of Drosophila melanogaster. Molecular Ecology, 25, 1157–1174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhan V, Parkash R, Aggarwal DD (2014) Effects of body-size variation on flight-related traits in latitudinal populations of Drosophila melanogaster. Journal of Genetics, 93, 103–112. [DOI] [PubMed] [Google Scholar]
- Bonhomme M, Chevalet C, Servin B et al. (2010) Detecting Selection in Population Trees: The Lewontin and Krakauer Test Extended. Genetics, 186, 241–262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradley TJ, Williams AE, Rose MR (1999). Physiological responses to selection for desiccation resistance in Drosophila melanogaster. American Zoologist, 39, 337–345. [Google Scholar]
- Brummel T, Ching A, Seroude L, Simon AF, Benzer S (2004) Drosophila lifespan enhancement by exogenous bacteria. Proceedings of the National Academy of Sciences of the United States of America, 101, 12974–12979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bubliy OA, Loeschcke V (2005) Correlated responses to selection for stress resistance and longevity in a laboratory population of Drosophila melanogaster. Journal of Evolutionary Biology 18, 789–803. [DOI] [PubMed] [Google Scholar]
- Caracristi G, SchlÖtterer C (2003) Genetic differentiation between American and European Drosophila melanogaster populations could be attributed to admixture of African alleles. Molecular Biology and Evolution, 20, 792–799. [DOI] [PubMed] [Google Scholar]
- Chippindale AK, Gibbs AG, Sheik M, Yee K, Djawdan M, Bradley TJ, Rose MR (1998) Resource acquisition and the evolution of stress resistance in Drosophila melanogaster. Evolution, 52, 1342–1352. [DOI] [PubMed] [Google Scholar]
- Chown SL (1993) Desiccation resistance in six sub-Antarctic weevils (Coleoptera: Curculionidae): humidity as an abiotic factor influencing assemblage structure. Functional Ecology, 7, 318–325. [Google Scholar]
- Chown SL (2002) Respiratory water loss in insects. CBP-Part A, 133, 791–804. [DOI] [PubMed] [Google Scholar]
- Chown SL, Le Lagadec MD, Scholtz CH (1999) Partitioning variance in a physiological trait: desiccation resistance in keratin beetles (Coleoptera, Trogidae) Functional Ecology, 13, 838–844. [Google Scholar]
- Comeron JM, Ratnappan R, Bailin S. (2012) The many landscapes of recombination in Drosophila melanogaster. PLoS Genetics, 8, e1002905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Corbett-Detig RB, Hartl DL (2012) Population genomics of inversion polymorphisms in Drosophila melanogaster. PLoS Genetics, 8, e1003056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coyne JA, Bundgaard J, Prout T (1983) Geographic variation of tolerance to environmental stress in Drosophila pseudoobscura. American Naturalist, 122, 474–488. [Google Scholar]
- Coyne JA, Milstead B (1987) Long-distance migration of Drosophila. Dispersal of D. melanogaster alleles from a Maryland orchard. American Naturalist, 130, 70–82. [Google Scholar]
- Daub JT, Hofer T, Cutivet E, Dupanloup I, Quintana-Murci L, Robinson-Rechavi M, Excoffier L (2013) Evidence for polygenic adaptation to pathogens in the human genome. Molecular Biology and Evolution, 30, 1544–1558. [DOI] [PubMed] [Google Scholar]
- David J, Allemand R, Van Herrewege J, Cohet Y (1983) Ecophysiology: abiotic factors. In: Genetics and Biology of Drosophila (Ashburner M, Carson HL, Thompson JN, eds) Academic Press, London, Vol 3, 105–170. [Google Scholar]
- Davidson JK (1988) Extremes of climate and genetic heterogeneity in Australian populations of the dipteran species Drosophila melanogaster. Journal of Biogeography, 15, 481–487. [Google Scholar]
- Denlinger DL, 2002. Regulation of diapause. Annual Review of Entomology 47, 93–122. [DOI] [PubMed] [Google Scholar]
- Duchen P, Zivkovic D, Hutter S, Stephan W, Laurent S (2013) Demographic inference reveals African and European admixture in the North American Drosophila melanogaster population. Genetics, 193, 291–301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eden R, Navon R, Steinfeld I, Lipson D, Yakhini Z (2009) Gorilla: A tool for discovery and visualization of enriched GO terms in ranked genes list. BMC Bioinformatics, 10, 48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edney EB (1977). Water Balance in Land Arthropods (Zoophysiology and Ecology, Vol 9) Springer-Verlag, Berlin, Heidelberg and New York. [Google Scholar]
- Endler JA (1977) Geographic variation, Speciation, and Clines. Princeton University Press, Princeton, USA. [PubMed] [Google Scholar]
- Flowers JM, Sezgin E, Kumagai S, Duvernell DD, Matzkin LM, Schmidt PS, Eanes WF (2007) Adaptive Evolution of Metabolic Pathways in Drosophila. Molecular Biology Evolution, 24, 1347–1354. [DOI] [PubMed] [Google Scholar]
- Foley BR, Telonis-Scott M (2011) Quantitative genetic analysis suggests causal association between cuticular hydrocarbon composition and desiccation survival in Drosophila melanogaster. Heredity, 106, 68–77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Folk DG, Han C, Bradley TJ (2001) Water acquisition and partitioning in Drosophila melanogaster: effects of selection for desiccation-resistance. Journal of Experimental Biology, 204, 3323–3331. [DOI] [PubMed] [Google Scholar]
- Gefen E, Marlon AJ, Gibbs AG(2006) Selection for desiccation resistance in adult Drosophila melanogaster affects larval development and metabolite accumulation. Journal of Experimental Biology 209, 3293–3300. [DOI] [PubMed] [Google Scholar]
- Gibbs AG, Chippindale AK, Rose MR (1997) Physiological mechanisms of evolved desiccation resistance in Drosophila melanogaster. Journal of Experimental Biology, 200, 1821–1832. [DOI] [PubMed] [Google Scholar]
- Gibbs AG, Fukuzato F, Matzkin LM (2003) Evolution of water conservation mechanisms in desert Drosophila. Journal of Experimental Biology, 206, 1183–1192. [DOI] [PubMed] [Google Scholar]
- Gibbs AG, Gefen E (2009) Physiological adaptation and laboratory selection. In: Experimental Evolution Garland T and Rose MR, eds. University of California Press, pp. 523–550. [Google Scholar]
- Gibbs AG, Matzkin LM (2001) Evolution of water balance in the genus Drosophila. Journal of Experimental Biology, 204, 2331–2338. [DOI] [PubMed] [Google Scholar]
- Gibbs AG, Rajpurohit S (2010) Water-proofing properties of cuticular lipids. In Insects Lipids: Biology, Biochemistry and Chemical Biology (eds Blomquist GJ, Bagneres AG), pp. 100–120. Cambridge University Press; Cambridge, UK. [Google Scholar]
- Griffin PC, Hangartner SB, Fournier-Level A, Hoffmann AA (2017) Genomic trajectories to desiccation resistance: convergence and divergence among replicate selected Drosophila lines. Genetics 205:871–890. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths JA, Schiffer M, Hoffmann AA. (2005) Clinal variation and laboratory adaptation in the rainforest species Drosophila birchii for stress resistance, wing size, wing shape and development time. Journal of Evolutionary Biology, 18, 213–222. [DOI] [PubMed] [Google Scholar]
- Hadley NF (1994) Water relations of terrestrial Arthropods. Academic Press Inc; California, USA. [Google Scholar]
- Heerwaarden BV, Sgro CM (2014) Is adaptation to climate change really constrained in niche specialist? Proceedings of The Royal Society B, 281, 20140396 10.1098/rspb.2014.0396 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann AA, Hallas R, Sinclair C, Mitrovski P (2001) Levels of variation in stress resistance in Drosophila among strains, local populations, and geographic regions: Patterns for desiccation, starvation, cold resistance, and associated traits. Evolution, 55, 1621–1630. [DOI] [PubMed] [Google Scholar]
- Hoffmann AA, Harshmann LG (1999) Desiccation and starvation resistance in Drosophila: patterns of variation at the species, populations and intrapopulation levels. Heredity, 83, 637–643. [DOI] [PubMed] [Google Scholar]
- Hoffmann AA, Parsons PA (1989) Selection for increased desiccation resistance in Drosophila melanogaster: Additive genetic control and correlated responses for other stresses. Genetics, 122, 837–845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffmann AA, Parsons PA (1991) Evolutionary Genetics and Environmental Stress. Oxford University Press, Oxford. [Google Scholar]
- Hoffmann AA, Parsons PA (1993) Direct and correlated responses to selection for desiccation resistance: a comparison of Drosophila melanogaster and D. simulans. Journal Evolutionary Biology, 6, 643 –657. [Google Scholar]
- Hoffmann AA, Shirriffs J, Scott M (2005) Relative importance of plastic vs genetic factors in adaptive differentiation: geographical variation for stress resistance in Drosophila melanogaster from eastern Australia. Functional Ecology, 19, 222–227. [Google Scholar]
- Huang W, Massouras A, Inoue Y, Peiffer J, Ràmia M, Tarone AM, Turlapati L, Zichner T, Zhu D, Lyman RF, Magwire MM, Blankenburg K, Carbone MA, Chang K, Ellis LL, Fernandez S, Han Y, Highnam G, Hjelmen CE, Jack JR, Javaid M, Jayaseelan J, Kalra D, Lee S, Lewis L, Munidasa M, Ongeri F, Patel S, Perales L, Perez A, Pu L, Rollmann SM, Ruth R, Saada N, Warner C, Williams A, Wu Y-Q, Yamamoto A, Zhang Y, Zhu Y, Anholt RRH, Korbel JO, Mittelman D, Muzny DM, Gibbs RA, Barbadilla A, Johnston JS, Stone EA, Richards S, Deplancke B, Mackay TFC (2014) Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Research, 24, 1193–1208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- James AC, Azevedo RBR, Partridge L (1995) Cellular basis and developmental timing in a size cline of Drosophila melanogaster. Genetics, 140, 659–666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalra B, Parkash R, Aggarwal DD (2014) Divergent mechanisms for water conservation in Drosophila species. Entomologia Experimentalis et Applicata, 151, 43–56. [Google Scholar]
- Kang L, Aggarwal DD, Rashkovetsky E, Korol AB, Michalak P (2016) Rapid genomic changes in Drosophila melanogaster adapting to desiccation stress in an experimental evolution sustem. BMC Genomics 17:233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kao JY, Zubair A, Salomon MP, Nuzhdin SV, Campo D (2015) Population genomic analysis uncovers African and European admixture in the North American Drosophila melanogaster population. Genetics, 193, 291–301. [DOI] [PubMed] [Google Scholar]
- Karan D, Dahiya N, Munjal AK, Gibert P, Moreteau B, Parkash R, David JR (1998) Desiccation and starvation tolerance of adult Drosophila: opposite latitudinal clines in natural populations of three different species. Evolution 52, 825–831. [DOI] [PubMed] [Google Scholar]
- Kingsolver JG 1979. Thermal and hydric aspects of environmental heterogeneity in the pitcher plant mosquito. Ecological Monographs, 49, 357–376. [Google Scholar]
- Kolaczkowski B, Kern AD, Holloway AK, Begun DJ (2011) Genomic differentiation between temperate and tropical Australian populations of Drosophila melanogaster. Genetics, 187, 245–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lavington E, Cogni R, Kuczynski C, Koury S, Behrman EL, O’Brien KR, Schmidt P, Eanes WF (2014) A small system—high-resolution study of metabolic adaptation in the central metabolic pathway to temperate climates in Drosophila melanogaster. Molecular Biology & Evolution, 31, 2032–2041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leather S, Walters K, Bale J (1993) The Ecology of Insects Overwintering. Cambridge University Press, Cambridge. [Google Scholar]
- Lemeunier F, David JR, Tsacas L, Ashburner M (1986) The melanogaster species group. Pp. 147–256 in ASHBURNER M, CARSON HL, and THOMPSON J, eds. The genetics and biology of Drosophila. Vol. 3e Academic Press, London and Orlando. [Google Scholar]
- Lewontin RC, Krakauer J (1973) Distribution of gene frequency as a test of the theory of the selective neutrality of polymorphisms. Genetics, 74, 175–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mackay TFC, Richards S, Stone EA et al. (2012) The Drosophila melanogaster Genetic Reference Panel. Nature, 482, 173–178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacMillan HA, Walsh JP, Sinclair BJ (2009) The effects of selection for cold tolerance on cross-tolerance to other environmental stressors in Drosophila melanogaster. Insect Science, 16, 263–276. [Google Scholar]
- Marron MT, Markow TA, Kain KJ, Gibbs AG (2003) Effects of starvation and desiccation on energy metabolism in Drosophila. Journal of Insect Physiology, 49, 261–270. [DOI] [PubMed] [Google Scholar]
- Martini SV, Goldenberg RC, Fortes FSA, Campos-de-Carvalho AC, Falkenstein D, Morales MM (2004) Rhodnius prolixus tubule’s aquaporin expression is modulated by 5-hydroxytryptamine. Archives of Insect Biochemistry & Physiology, 57, 133–141. [DOI] [PubMed] [Google Scholar]
- Matzkin LM, Watts TD, Markow TA (2007) Desiccation resistance in four Drosophila species. Fly, 1, 268–273. [DOI] [PubMed] [Google Scholar]
- Matzkin LM, Watts TD, Markow TA (2009) Evolution of stress resistance in Drosophila: interspecific variation in tolerance to desiccation and starvation. Functional Ecology, 23, 521–527. [Google Scholar]
- Parkash R, Lambhod C, Singh D (2014) Thermal developmental plasticity affects body size and water conservation of Drosophila nepalensis from the Western Himalayas. Bulletin of Entomological Research, 104, 504–516. [DOI] [PubMed] [Google Scholar]
- Parkash R, Rajpurohit S, Ramniwas S (2008) Changes in body melanisation and desiccation resistance in highland vs. lowland populations of D. melanogaster. Journal of Insect Physiology, 54, 1050–1056. [DOI] [PubMed] [Google Scholar]
- Parkash R, Sharma V, Kalra B. (2009) Impact of body melanization on desiccation resistance in montane populations of D. melanogaster: Analysis of seasonal variation. Journal of Insect Physiology, 55, 898–908. [DOI] [PubMed] [Google Scholar]
- Parkash R, Tyagi PK, Sharma I, Rajpurohit S(2005) Adaptations to environmental stress in altitudinal populations of two Drosophila species. Physiological Entomology, 30, 353–361. [Google Scholar]
- Parsons PA (1983) The Evolutionary Biology of Colonizing Species. Cambridge University Press, Cambridge. [Google Scholar]
- Rajpurohit S, Nedved O (2013) Clinal variation in fitness related traits in tropical drosophilids of the Indian subcontinent. Journal of Thermal Biology, 38, 345–354. [Google Scholar]
- Rajpurohit S, Nedved O, Gibbs AG (2013b) Meta-analysis of geographical clines in desiccation tolerance of Indian drosophilids. Comparative Biochemistry and Physiology-Part A, 164, 391–398. [DOI] [PubMed] [Google Scholar]
- Rajpurohit S, Oliveira CC, Etges WJ, Gibbs AG (2013a) Functional genomic and phenotypic responses to desiccation in natural populations of a desert drosophilid. Molecular Ecology, 22, 2698–2715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajpurohit S, Peterson LM, Orr AJ, Marlon AJ, Gibbs AG (2016) An experimental evolution test of the relationship between melanism and desiccation survival in insects. PLoS ONE, 11(9): e0163414, doi: 10.1371/journal.pone.0163414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajpurohit S, Schmidt PS (2016) Measuring thermal behavior in smaller insects: A case study in Drosophila melanogaster demonstrates effects of sex, geographic origin, and rearing temperature on adult behavior. Fly, 26, 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajpurohit S, Zhao X, Schmidt PS (2017b) A resource on latitudinal and altitudinal clines of ecologically relevant phenotypes of the Indian Drosophila. Scientific Data - Nature, 4:170066, DOI: 10.1038/sdata.2017.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajpurohit S, Hanus R, Vrkoslav V, Behrman EL, Bergland A, Dmitri P, Cvacka J, Schmidt PS (2017a) Adaptive dynamics of cuticular hydrocarbon profiles in Drosophila. Journal of Evolutionary Biology, 30, 66–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds SE (1975) The mechanical properties of the abdominal cuticle of Rhodnius larvae. Journal of Experimental Biology, 62, 69–80. [DOI] [PubMed] [Google Scholar]
- Saunders DS, Henrich VC, Gilbert LI (1989) Induction of diapause in Drosophila melanogaster: photoperiodic regulation and the impact of arrhythmic clock mutations on time measurement. Proceedings of the National Academy of Sciences of the USA, 86, 3748–3752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schmidt PS, Matzkin LM, Ippolito M, Eanes WF (2005) Geographic variation in diapause incidence, life history traits and climatic adaptation in Drosophila melanogaster. Evolution, 59, 1721–1732. [PubMed] [Google Scholar]
- Sorensen JG, Nielsen MM, Loeschcke V (2007) Gene expression profile analysis of Drosophila melanogaster selected for resistance to environmental stressors. Journal of Evolutionary Biology, 20, 1624–1636. [DOI] [PubMed] [Google Scholar]
- Sorensen JG, Norry FM, Scannapieco AC, Loeschcke V. (2005) Altitudinal variation for stress resistance traits and thermal adaptation in adult Drosophila buzzatii from the new world. Journal of Evolutionary Biology, 18, 829–837. [DOI] [PubMed] [Google Scholar]
- Telonis-Scott M, Gane M, DeGaris S, Sgro CM, Hoffmann AA. (2012) High resolution mapping of candidate alleles for desiccation resistance in Drosophila melanogaster under selection. Molecular Biology and Evolution, 29, 1335–1351. [DOI] [PubMed] [Google Scholar]
- Telonis-Scott M, Guthridge KM, Hoffmann AA (2006) A new set of laboratory-selected Drosophila melanogaster resistance: response to selection, physiology and correlated responses. Journal of Experimental Biology, 209, 1837–1047. [DOI] [PubMed] [Google Scholar]
- Telonis-Scott M, Hoffmann AA, Sgro CM (2011) The molecular genetics of clinal variation: a case study of ebony and thoracic trident pigmentation in Drosophila melanogaster from eastern Australia. Molecular Ecology, 20, 2100–2110. [DOI] [PubMed] [Google Scholar]
- Telonis-Scott M, Sgro CM, Hoffmann AA, Griffin PC (2016) Cross-study comparison reveals common genomic, network, and functional signatures of desiccation resistance in Drosophila melanogaster. Molecular Biology & Evolution, 33, 1053–1067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Umina PA, Weeks AR, Kearney MR, McKenchnie SW, Hoffmann AA (2005) A rapid shift in a classic clinal pattern in Drosophila reflecting climate change. Science, 308, 691–693. [DOI] [PubMed] [Google Scholar]
- Verrelli BC, Eanes WF (2001) The functional impact of Pgm amino acid polymorphism on glycogen content in Drosophila melanogaster. Genetics, 159, 201–210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whitlock MC, McCauley DE (1999) Indirect measures of gene flow and migration: FST≠1/(4Nm+1). Heredity, 82, 117–125. [DOI] [PubMed] [Google Scholar]
- Zar JH (2010) Biostatistical Analysis. Prentice Hall Inc., New Jersey. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Schematic representation of field based mesocosm units. Each mesocosm was an 6m3 outdoor insect rearing enclosure surrounding a mature (dwarf) peach tree. A total of 10 cages were established for this experimental work.
qq-plot of genome wide association analysis of intersex averaged desiccation tolerance. Observed p-values are plotted along the y-axis and expected p-values along the x-axis. The solid red line depicts a 1:1 correspondence.
Multiple relationship between thermal plasticity of desiccation tolerance in D. melanogaster and various climatic variables analyzed by unconstrained ordination (Principal Component Analysis). The first ordination axis explained 65.94% of variability and the second axis explained 27.14%. Temperature correlates best with the first axis.
3D Surface plot (A) of Tave and RHave along the latitudes of east coast of the U.S. RHave = Distance weighted least square. The color scale relates to the RHave values distribution along the latitudes. Bar plot (B) is showing seasonal variations (summer vs. winter) in RHave for five latitudinal stations. Relative humidity data for winter and summer are averages for Nov-March and Apr-Oct months respectively. Averaging of relative humidity across an entire season may not be an appropriate level of resolution-relative humidity might vary (biologically) over a shorter timescale. Climatic data were obtained from National Oceanic and Atmospheric Administration (NOAA; www.ncdc.noaa.gov). We used 30-year averages (1980–2010) for all the climatic parameters.
Data on six climatic variables (Tmin: temperature minimum; Tmax: temperature maximum; Tave: temperature average; RHmin: relative humidity minimum; RHmax: relative humidity maximum; RHave: relative humidity average) for latitudinal and seasonal collections used in this study.
Mean (±s.e.m) CO2 and water vapor emission rates among the experimental fly populations (see methods for sample sixes and statistics).
Desiccation survival status in DGRP lines. The status of Wolbachia infection and cytological analysis of polymorphic inversions were updated from Huang et al. 2014 (see Supplemental data file S6 in Huang et al. 2014).
ANOVA on desiccation tolerance assays performed for DGRP lines. The analyses present sex separated and sexes averaged results for effects of Wolbachia infection and inversions status.
SNPs identified from the genome-wide association study for desiccation tolerance in D. melanogaster.
Genes containing SNPs associated with desiccation tolerance in DGRP lines. GO Biological Process information was obtained using DAVID 6.8 (Huang et al. 2014).