

Abstract
To perform their function, transcription factors and DNA-repair/modifying enzymes must first locate their targets in the vast presence of nonspecific, but structurally similar sites on genomic DNA. Before reaching their targets, these proteins stochastically scan DNA and dynamically move from one site to another on DNA. Solution NMR spectroscopy provides unique atomic-level insights into the dynamic DNA-scanning processes, which are difficult to gain by any other experimental means. In this review, we provide an introductory overview on the NMR methods for the structural, dynamic, and kinetic investigations of target DNA search by proteins. We also discuss advantages and disadvantages of these NMR methods over other methods such as single-molecule techniques and biochemical approaches.
Keywords: DNA scanning, dynamics, kinetics, NMR, protein-DNA interactions, target search
1. Introduction: DNA scanning by proteins
Protein-DNA interactions are vital for life. Living organisms regulate expression of genes and maintain integrity of the genome through protein-DNA interactions involving transcription factors or DNA-repair/modifying enzymes. These DNA-binding proteins must first locate their specific target sites in the vast presence of nonspecific but structurally similar sites on genomic DNA. The genome of higher eukaryotes contains billions of base pairs [1] and can potentially provide ~109 nonspecific sites on DNA, whereas functional target sites of each transcription factor or DNA-repair/modifying enzyme are far fewer (typically, ~102–103 sites) [2]. Functionality of these proteins should depend on their efficiency in locating targets on DNA through stochastic search processes. Therefore, it is important to understand at molecular and atomic levels how these proteins scan DNA, recognize sequences, and locate the targets.
In the target search processes, the proteins nonspecifically interact with DNA. Although affinity for nonspecific DNA is weaker than that for targets, the vast quantity of nonspecific sites compensates for the weak affinity, making profound overall impacts on these proteins [3–5]. It should be noted that DNA density in cell nuclei is as high as ~100 mg/ml [6]. This high density is understandable, considering that a total length of ~2 m of human genomic DNA (= 2 × [3.2 × 109 bp] × [0.34 × 10−9 m/bp]) is confined in the nucleus for which a typical diameter is ~6 μm [1]. Histones make this condensation possible, occupying ~80% of genomic DNA as nucleosomes. The average length of linker DNA segments between nucleosomes is ~56 bp [7]. The overall concentration of linker DNA segments is estimated to be as high as ~500 μM in the nuclei. Because this concentration alone is far higher than typical apparent dissociation constants of sequence-specific or structure-specific DNA-binding proteins for nonspecific DNA, the majority of these proteins must be nonspecifically bound to genomic DNA before reaching their targets.
When nonspecifically interacting with DNA, these proteins rapidly move from one site to another, scanning DNA to locate their targets (Figure 1). Berg and von Hippel conceptually defined three major mechanisms for protein translocation on DNA: sliding, dissociation & re-association, and intersegment transfer [8]. Sliding corresponds to one-dimensional (1D) diffusion along DNA and involves random walks of protein being nonspecifically bound to DNA. Binding to nonspecific sites near targets can accelerate target association because sliding from nonspecific sites allows proteins to efficiently reach a target through 1D diffusion [2, 9–11]. The dissociation & re-association mechanism is often called three-dimensional (3D) search. When DNA concentration is high (e.g., in the nuclei), dissociation from DNA is the rate-limiting step in this translocation mechanism. When translocation through this mechanism occurs between two sites in close proximity (e.g. within ~1–7 nm [12]), it is called hopping. Intersegment transfer (also known as direct transfer) is a unique mechanism that allows protein to directly transfer from one DNA duplex to another without going through the intermediary of free protein. This involves an intermediate where a protein molecule transiently bridges two DNA duplexes. Intersegment transfer mechanism may facilitate bypassing of nucleosome because its two ends are close in the three-dimensional structure [13]. These distinct translocation mechanisms coexist in solution, and their relative contributions to the overall efficiency of target DNA search should depend on various factors such as ionic strength, protein structure, DNA density, DNA geometry, and the presence of other proteins. However, details of the DNA scanning mechanisms are not well understood.
Figure 1.

Protein translocation on DNA through nonspecific interactions.
Our goal in this review article is to provide general readers with an introductory overview on nuclear magnetic resonance (NMR) methods for investigating the target DNA search processes of proteins. These NMR methods allow us to gain a great deal of information regarding how proteins nonspecifically interact with DNA during the search processes and how they move from one site to another on DNA. We first compare these NMR methods with biochemical and single-molecule methods for investigating target DNA search processes and explain unique strengths of these methods. Then, we will elaborate on practical details of the NMR methods that provide dynamic and kinetic information on target DNA search processes.
2. Advantages of NMR over other methods for investigating target DNA search of proteins
In the target DNA search processes, proteins dynamically move on DNA, changing their locations. This movement creates a challenge in studying these processes. Because proteins can change their locations through distinct mechanisms that are simultaneously present in the same system, characterizing a particular translocation mechanism is difficult, particularly when molecular ensembles are measured in solution. In the 21st century, there have been substantial advances in methodology for research on the target DNA search processes. Progress in single-molecule biophysics techniques is particularly remarkable, allowing direct observations of protein translocation on DNA in vitro and even in living cells. These single-molecule methods are recently overviewed in excellent reviews [14–21]. Elegant biochemical methods were also developed for kinetic investigations of target search and protein translocation on DNA [12, 22–29]. As discussed below, NMR spectroscopy has provided unique insight into how proteins scan DNA at an atomic level. Capability and suitability of these methods for target search research are summarized in Table 1.
Table 1.
Comparison of the experimental approaches for investigating target DNA search of proteins.
| Methods | NMR | Single-molecule | Biochemical |
|---|---|---|---|
| Capability | |||
| Atomic details of DNA scanning | +++ a) | − | − |
| Direct visualization of protein translocation on DNA | − | +++ | − |
| Kinetic analysis of protein translocation on DNA | ++b) | ++ | ++ |
| Kinetic analysis of target DNA association | + | ++ | +++ |
| Analysis of target DNA search process in cells | + | +++ | + |
Symbols are as follows: +++, well suited; ++, feasible; +, possible, but not well explored; -, not possible.
See Section 4.
See Section 5.
The greatest advantage of the NMR methods over other methods is that NMR spectroscopy can provide atomic-level information on the highly dynamic processes whereby proteins scan DNA and locate their targets. NMR spectroscopy is particularly suited to study structural dynamics of biological macromolecules and can provide spatiotemporal information on dynamics [30]. As described in Section 4, there are several NMR methods that can provide different types of information on the DNA-scanning process. Some NMR data give structural information on the proteins scanning DNA. Other NMR data give information on conformational mobility of particular domains or moieties within the proteins bound to DNA. Dynamic behavior of each basic side chain at protein-DNA interfaces can also be analyzed using NMR relaxation data [31–34]. These NMR methods can provide structural and dynamic details on the proteins moving on DNA, which are very difficult to analyze at an atomic level by any other methods.
Through NMR experiments, one can also obtain quantitative information on kinetics of protein translocation on DNA at equilibrium. The range of kinetic rate constants that can be determined by NMR is as wide as 0.1 – 10,000 s−1, though suitable methods should be adopted depending on the timescale of the analyzed processes. As described in Section 5, several NMR methods are available for the kinetic investigations of protein translocation on DNA and differ in the applicable range in terms of analyzable timescale. These NMR experiments for investigating the kinetics of protein-DNA interactions can be conducted under various buffer conditions, offering flexibility in experimental design. A qualitative in-cell NMR study of protein-DNA interactions in E. coli cells has also been demonstrated [35]. NMR spectroscopy can thus serve as a versatile tool for investigating target DNA search processes of proteins.
3. NMR sample preparation for studying protein-DNA interactions
NMR studies on kinetics and dynamics of target DNA search by proteins require solution samples containing protein and DNA in which protein can move from one site to another. Because NMR spectroscopy is not as sensitive as many other spectroscopic techniques, high concentrations of proteins and DNA are required and high solubility of these materials is crucial for NMR experiments. Although high-field magnets and cryogenic probes have increased detection sensitivity for modern NMR spectrometers, many NMR experiments still require 0.1–1 mM proteins and DNA in a 0.25–0.5 ml solution. Milligrams of protein and DNA are necessary to prepare such a sample. Molecular size is also a limiting factor for NMR application. For typical heteronuclear multi-dimensional NMR experiments with backbone NH detection, the total molecular weight should be < 100 kDa. For methyl-TROSY-type experiments with side-chain CH3 detection, the total molecular weight can be bigger. In fact, 200-kDa nucleosome core particles of histone octamer and 167-bp DNA were studied by the methyl TROSY methods [36]. Typical buffers for NMR experiments are at pH 5–8 and an ionic strength less than 150 mM. We typically use 20–100 mM KCl (or NaCl) for NMR measurements of protein-DNA complexes. In general, sensitivity in NMR detection (particularly with cryogenic probes) is higher at a lower conductivity of solution [37, 38]. When a high concentration of salt is required, use of sample tubes with a thinner sample diameter can mitigate the adverse effect of salt [39, 40].
Due to large amounts of materials required, protein-DNA complexes might aggregate or precipitate during preparation of NMR samples, particularly when high concentrations of proteins and DNA are mixed at a low ionic strength. Figure 2 shows our typical procedures of sample preparation for NMR studies of protein-DNA interactions. To avoid the aggregation or precipitation, we initially mix relatively low concentrations (~10–50 μM) of proteins and DNA at a relatively high ionic strength (≥ 300 mM KCl). In our experience, nonspecific DNA complexes of proteins exhibit a stronger tendency to precipitate at low ionic strength. After mixing the protein and DNA components at high ionic strength, we slowly reduce ionic strength. When ionic strength decreases too rapidly, some proteins precipitates (in some cases, the precipitated materials can be recovered by re-dissolving in a high ionic-strength buffer). So, we often reduce ionic strength through dialysis. A gradual decrease in salt concentration seems to generally help prevent aggregation of protein-DNA complexes, as known for nucleosome reconstitutions [41]. After reaching a desired ionic strength (typically, 20–100 mM KCl in our experiments), protein-DNA solutions are concentrated and buffer-exchanged using centrifugal concentrators such as Amicon Ultra-4 (Millipore) or Vivaspin-6 (Sartorius). A deuterated buffer, such as Tris-d11 or succinate-d4, is used at this point if 1H signals from buffer molecules could interfere with planned NMR experiments.
Figure 2.
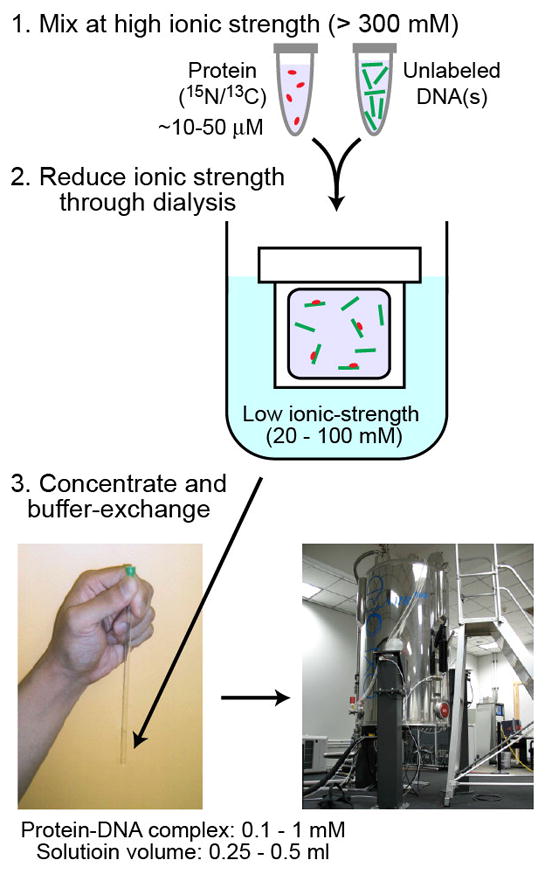
Typical procedures for preparation of protein–DNA solutions for NMR investigations of target DNA search processes.
For NMR studies of nonspecific DNA-protein complexes, concentration of DNA should be sufficiently higher than that of protein. We typically use molar ratios of protein to nonspecific DNA of 1:2–4. A large excess of DNA helps prevent binding of more than one protein molecule to the same DNA molecule. This is important because the multiple bindings would complicate data analysis and also enhance aggregation and precipitation. Use of mM concentrations of nonspecific DNA mimics physiological conditions because it is comparable to the concentration of linker DNA segments in the nuclei, as mentioned in Section 1. DNA length is also an important parameter that affects solubility and stability of protein-DNA complexes in NMR samples. When nonspecific DNA complexes are analyzed, DNA must not contain any high-affinity sequences that are similar to the target sequence. To confirm the absence of any high-affinity site, we use fluorescence-based competitive binding assays [42], which directly provide relative affinity of competitor with respect to that of fluorescent-probe DNA containing a high-affinity site.
4. NMR-based analysis of DNA scanning by proteins
The DNA-scanning processes are highly dynamic, during which proteins rapidly and stochastically change their locations on nonspecific DNA before reaching their targets. Solution NMR spectroscopy is well suited to study such dynamic systems. Although the DNA-scanning processes represent intermediates before the proteins reach their targets, these processes can be investigated most conveniently by using solutions of nonspecific DNA complexes that do not contain any targets. NMR was used to study nonspecific protein-DNA complexes in solution for gene-regulatory proteins (e.g., the lac repressor [43], Egr-1 [13, 44], Ets-1 [45], ETV6 [46], HMGB1 [47], HoxD9 [48], LmrR [49], Oct1 [50], Sox2 [51], and ZNF217[52]) and as well as for DNA-repair/modifying enzymes (e.g., EcoRI [53], M.HhaI [54], and UNG [55, 56]). Because proteins can move on nonspecific DNA, a solution of a nonspecific complex contains many states with the protein being located at different sites. However, NMR samples of nonspecific DNA-protein complexes typically show only a single set of signals because transitions between these states are so rapid that resonances of individual states are averaged (i.e., the “fast exchange” regime in the terminology of NMR spectroscopy). An example is shown in Figure 3. Such samples of nonspecific complexes can be used to study how proteins scan DNA before reaching their targets. The following NMR methods are particularly useful to study the DNA-scanning dynamics.
Figure 3.

1H-15N heteronuclear correlation spectra recorded for the specific and nonspecific DNA complexes of the Egr-1 zinc-finger protein [13]. Although the protein can be located at various sites on nonspecific DNA with a mean residence time being ~1–10 μs at each site [23], the spectrum of the nonspecific complex shows only a single set of signals due to rapid translocation that occurs in the fast exchange regime.
4.1. NMR relaxation
NMR relaxation data for backbone and side-chain moieties are useful to study dynamics of proteins on timescales ranging from ps – ns to μs – ms [57–60]. There are several different types of NMR relaxation parameters (e.g., longitudinal and transverse relaxation rates, cross relaxation rates, cross correlation rates, etc.), which reflect internal motions of various moieties within proteins. Relaxation parameters of 15N, 13C, and 2H nuclei are particularly useful for investigating protein dynamics, whereas 1H relaxation reflects more about structure due to strong 1H-1 H dipole-dipole interactions. Using NMR relaxation data, one can obtain information on 1) conformational flexibility of protein backbone and side chains, 2) conformational equilibrium between different states, 3) inter-domain dynamics, and 4) kinetics of molecular interactions. Analysis of NMR relaxation for nonspecific DNA complexes of proteins allows us to learn how these proteins scan DNA.
Figure 4A shows an example of such an analysis. This figure displays some backbone 15N relaxation data for the nonspecific and specific DNA complexes of the Egr-1 zinc-finger protein. Egr-1 recognizes DNA via three zinc fingers, ZF1, ZF2, and ZF3, each of which contacts with DNA in the crystal structure of the complex with the target DNA [61–63]. Backbone 15N relaxation data were compared with the nonspecific and specific complexes with 28-bp DNA. The specific DNA contains the Egr-1 recognition sequence, GCGTGGGCG, whereas the nonspecific DNA does not contain any high-affinity sequences. The backbone relaxation data for all three zinc fingers ZF1, ZF2, and ZF3 were similar for the specific DNA complex. However, the backbone 15N relaxation data for ZF1 in the nonspecific complex significantly differed from those for ZF2 and ZF3 in the nonspecific complex. These and other data suggested that ZF1 is mainly dissociated from DNA when the Egr-1 zinc-finger protein scans DNA. When ZF1 is locally dissociated from DNA and ZF2 and ZF3 are bound to DNA, ZF1 undergoes almost independent domain motion, making 15N longitudinal (R1) and transverse (R2) relaxation rates of ZF1 differ from those of ZF2 and ZF3. The linker connecting ZF1 to ZF2 shows smaller heteronuclear 15N NOE values in the nonspecific complex than in the specific complex, suggesting that the linker is more flexible in the nonspecific complex. This is also consistent with the independent domain motion of ZF1 in the nonspecific complex.
Figure 4.

DNA-scanning by the Egr-1 zinc-finger protein [13, 44]. (A) Backbone heteronuclear 15N NOE and 15N R1 relaxation data for nonspecific and specific DNA complexes of the Egr-1 zinc-finger protein. 28-bp DNA duplexes were used. Black and magenta data points are data obtained at the 1H frequencies of 600 and 800 MHz, respectively. Blue dotted boxes show data points indicative of local dissociation of zinc finger 1 (ZF1). (B) RDC data for the nonspecific and specific DNA complexes of the Egr-1 zinc-finger protein. RDC 1DNH induced with 8 mg/ml Pf1 phage as a molecular alignment medium were analyzed. The main principal axis and the magnitude Da of the alignment tensor for individual zinc fingers are also shown. (C) Dynamic equilibrium between the recognition and search modes. The search mode facilitates translocation of the protein on DNA. The coarse-grained molecular dynamics simulations also showed the dynamic transitions between these states [13, 44].
4.2. Residual dipolar coupling (RDC)
Residual dipolar coupling (RDC) of nuclei in proteins weakly aligned to the magnetic field provide information about orientations of particular covalent bonds and are useful to study protein structure and dynamics [64–67]. Analysis of RDC data for proteins in the nonspecific and specific DNA complexes with DNA can allow us to assess structural dynamics of the DNA-scanning processes [13, 48, 50, 51]. For example, RDC data for the HoxD9 homeodomain for the nonspecific and specific DNA complexes suggested that this protein interacts with nonspecific DNA in the same manner as in the specific complex [48]. In contrast, RDC data for the Egr-1 zinc-finger protein clearly showed that the three zinc fingers behave differently in the specific complex with target DNA and in the nonspecific DNA complex [13]. As mentioned above, ZF1 in the nonspecific DNA complex is locally dissociated from DNA while ZF2 and ZF3 are bound to DNA. Since ZF1 undergoes independent domain motion with respect to the other part of the complex, the overall magnitude of RDCs for ZF1 in the nonspecific complex was substantially smaller than those for ZF2 and ZF3 in the nonspecific DNA complex (Figure 4B). Prediction of RDC for nonspecific DNA complexes is difficult because the observed samples contain many states with the protein being located at different sites. But if structural models of individual states can be built, the overall profile of RDC for nonspecific complexes can be predicted using structure-based de novo RDC prediction methods [68, 69]. Comparing the experimental RDC data with those predicted for the ensemble of various states in the nonspecific complex allows us to examine models on how proteins scan DNA [13, 48].
4.3. Paramagnetic relaxation enhancement (PRE)
NMR paramagnetic relaxation enhancement (PRE) arising from dipole-dipole interactions between 1H nuclei and unpaired electrons is a powerful tool for structural and dynamic studies of macromolecular complexes [70–72]. There are several methods for site-specific, covalent attachment of an extrinsic paramagnetic group to proteins or nucleic acids. For example, a paramagnetic Mn2+ ion can be site-specifically incorporated into the EDTA conjugated to DNA thymidine (dT-EDTA-Mn2+) [73]. A paramagnetic group can also be incorporated into DNA through conjugation of a nitroxide spin label to a nucleotide base [74–76]. Intermolecular PRE rates can be readily measured using DNA containing a paramagnetic group and 15N- and/or 13C-labeled proteins [77, 78]. In solution, proteins moving on DNA can be located near the paramagnetic group attached to a DNA base. Such a state causes strong PRE for the nuclei that are in close proximity from the Mn2+ ion [47, 48, 79]. Due to their r−6 dependence, PRE rates are sensitive to the presence of states in which the distances (r) between observed nuclei and the unpaired electrons of the paramagnetic group are relatively short. Even if such states are as minor as 1%, they can make a predominant contribution to observed PRE rates, allowing us to detect the low-population states [79]. When protein translocation on DNA occurs in the fast exchange regime, the PRE data can show which parts of the protein can become proximal to the paramagnetic group. This method thus provides structural information on proteins in the target DNA search processes.
4.4. Solvent PRE
Random collisions with paramagnetic co-solute molecules (e.g., gadolinium-diethylenetriamine pentaacettic acid bismethylamide [Gd-DTPA-BME] and 4-hydroxy-2,2,6,6-tetramethylpiperidin-1-oxyl [TEMPOL]) at relatively high concentrations cause sizable PRE for 1H nuclei of proteins [80–84]. This type of PRE, referred to as solvent PRE, is stronger for 1H nuclei near the molecular surface accessible to solvent and can be used to identify molecular interfaces. This method is applicable to highly dynamic complexes such as nonspecific DNA-protein complexes. In the free state, the interfaces are more exposed to the solvent and therefore exhibit larger solvent PRE than in the complex with DNA even when the protein is changing its location on DNA. Although chemical shift perturbation (CSP) upon complex formation may provide similar information on the molecular interface, non-interfacial regions often exhibit significant CSP as well [85]. In principle, solvent PRE is more straightforward than CSP in identifying the molecular interfaces. Relative magnitudes of solvent PRE are predictable from structure, assuming that spatial distribution of the paramagnetic co-solute molecules is uniform [82, 84]. Largely because this assumption is not necessarily valid for commonly used paramagnetic co-solute molecules, prediction of solvent PRE is only qualitatively accurate [81, 84]. Nonetheless, by comparing solvent PRE data for the free protein and for the nonspecific complex, the molecular interfaces during the DNA scanning process can be mapped on the protein surface. This technique was used to analyze how the HoxD9 homeodomain and the uracil DNA glycosylate interact with nonspecific DNA [48, 56].
5. NMR-based kinetic analysis of protein translocation on DNA
NMR spectroscopy can also be used to study kinetics of protein translocation on DNA. Among various spectroscopic techniques for analyzing molecular ensembles in solution, NMR is unique in that it allows investigations of fast kinetics at equilibrium. There are a number of NMR methods for kinetic investigations. Which one is the most appropriate depends on the timescale of the processes of interest [30]. Some are suitable for kinetic analysis of processes on a timescale of 10−2 – 1 s (e.g., z-exchange spectroscopy). Others are suitable for faster processes on a timescale of 10−5 – 10−3 s (e.g., line-shape analysis) or 10−4 – 10−2 s (e.g., CPMG relaxation dispersion methods). Typically, sliding of protein from one nonspecific site to an adjacent site on DNA occurs on a μs timescale. Dissociation from a nonspecific site occurs on a ms timescale, whereas dissociation from a specific, high-affinity site may even require minutes to hours. Furthermore, association and intersegment transfer are second-order processes and therefore their rates depend on the concentration of free DNA.
5.1. Mixture approach
In the NMR sample, protein translocation on DNA occurs within the same DNA molecule (i.e., intramolecular translocation) or between sites on different DNA molecules (i.e., intermolecular translocation). Distinguishing these two is not trivial. A general NMR approach called the “mixture approach” has been developed to facilitate kinetic analysis of intermolecular protein translocation on DNA [13, 48, 86–90]. This approach makes use of two different DNA duplexes a and b of comparable binding affinity and a 15N-labeled protein, from which two individual complexes of protein-DNA a and protein-DNA b and a mixture of these two complexes are prepared (Figure 5). DNA concentrations in these samples should be high enough for the protein to completely bind to either of these DNA. When the 1H-15N heteronuclear spectra are recorded for these three samples, the two samples of the individual complexes with DNA a and DNA b should show different 1H / 15N chemical shifts at least for some interfacial protein residues. For such residues, the spectrum of the mixture should exhibit one of the following features: 1) a single signal on the line between the two corresponding signals of the individual complexes, 2) two separate signals corresponding to the two complexes, or 3) very broad signal somewhere between the two signals. In the terminology of NMR spectroscopy, these three conditions are called fast, slow, and intermediate exchange regimes, respectively. If the intermolecular translocation processes occur in the fast exchange regime (this is usually the case for nonspecific DNA duplexes), NMR line-shape analysis or relaxation dispersion methods can be used to analyze the translocation kinetics (Section 5.2). If translocation of the protein occurs in the slow exchange regime (this is typically the case when DNA duplexes containing a high-affinity site are used), the 15N z-exchange method or the real-time approach can be used to analyze kinetics of intermolecular translocations between two DNA duplexes (Section 5.3).
Figure 5.
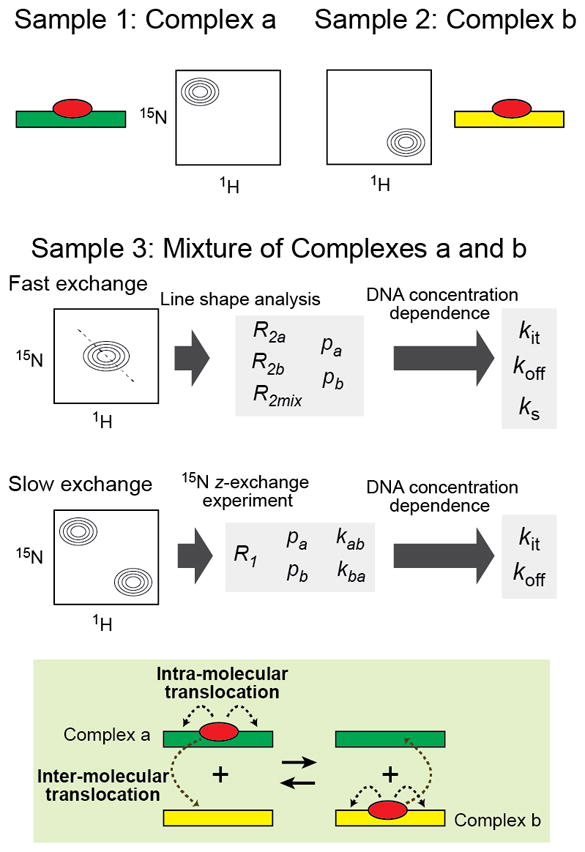
The mixture approach for NMR-based investigations of kinetics of protein translocation on DNA.
5.2. Kinetics of protein translocation on nonspecific DNA
Protein translocation on nonspecific DNA occurs through sliding, dissociation & re-association, and intersegment transfer, typically in the fast exchange regime. A discrete-state kinetics model shown in Figure 6A can represent these translocation processes. This model assumes that a nonspecific DNA duplex contains N distinct sites for binding. For example, for 24-bp DNA and a protein that covers 9 bp at each site, N is calculated to be 16 (=24 − 9 +1). Due to the structural pseudo-C2 symmetry of each DNA site, two opposite orientations are possible for a protein to bind to each site through short-range electrostatic interactions with the DNA backbone. Each nonspecific site is assumed to exhibit the same kinetic properties with the same rate constants for sliding (ks), dissociation (koff), association (kon), and intersegment transfer (kit). These kinetic rate constants are microscopic rate constants defined for each site (not for the entire DNA molecule). The macroscopic association rate constant for the DNA duplex is given by 2Nkon, and thus koff/(2Nkon) corresponds to the macroscopic apparent dissociation constant (Kd,app). The sliding rate constant ks is directly related to the macroscopic one-dimensional diffusion coefficient for sliding (D1) by D1 = l2ks, where l represents the distance (3.4 Å) between adjacent sites along the DNA axis [22]. D1 given in units of bp2 s-1 is equivalent to ks. Because the NMR experiments typically use DNA duplexes shorter than the persistence length (i.e., ~150 bp), intersegment transfers within the same DNA duplex are neglected, and only those between two DNA duplexes are considered. The kit constant is a second-order rate constant for this type of intermolecular intersegment transfer [22].
Figure 6.

NMR-based analysis of kinetics of protein translocation on nonspecific DNA. (A) A discrete-state kinetic model for protein translocation on nonspecific DNA [89]. Note that the rate constants kon and kit are defined as microscopic rate constants for each site. The corresponding macroscopic rate constants are 2Nkon and 2Nkit. (B,C) Mixture approach data on HoxD9 translocation on nonspecific DNA. Panel B shows overlaid spectra recorded for the individual complexes and their mixture. Panel C shows determination of the rate constants koff, kit, and ks from the apparent transverse relaxation rates of the three samples as a function of the concentration of free DNA [89].
In the mixture approach for kinetic analysis of these translocation processes on nonspecific DNA [48, 89], NMR line shapes of protein backbone NH groups are analyzed for three samples: two containing individual nonspecific complexes with DNA duplexes a and b and one containing a mixture of the two nonspecific complexes. Some examples of actual experimental data are shown in Figure 6B. This NMR approach provides accurate kinetic information on the intermolecular translocations of proteins between two DNA molecules, although this approach adopts simple two-state approximation. The validity of this approximation has been confirmed in a recent study using more rigorous McConnell equations that account for 4N + 1 microscopic states for a system containing a protein and two nonspecific DNA duplexes [89]. The rate constants koff and kit can be determined through analysis of apparent transverse relaxation rates from resonance line-shapes as a function of DNA concentration. Interestingly, the same analysis also provides semi-quantitative information on the rate constant ks and the one-dimensional diffusion coefficient D1 for protein sliding on DNA [89]. By the mixture approach, protein translocation on nonspecific DNA was analyzed for the Egr-1, HoxD9, and Sox2 proteins [13, 48, 51, 89].
5.3. Kinetics of protein translocation between high-affinity sites on DNA
Since the residence time of protein at a high-affinity site is far longer than that at nonspecific sites, protein translocation between two high-affinity sites typically occurs in the slow exchange regime. If protein translocation between two DNA duplexes a and b occurs with an exchange rate constant kex (= kab + kba) being roughly ~0.5–50 s−1, the 15N z-exchange method [88, 91, 92] can be used to determine the rate constants for translocation from DNA a to b (kab) and that for translocation from DNA b to a (kba). This method has been applied to intermolecular translocation of proteins between high-affinity sites on two different DNA duplexes [79, 86–88, 90, 93, 94] and also to intramolecular translocation between two high-affinity sites on the same DNA duplex [95]. In a 15N z-exchange experiment, in addition to the signals from the two complexes, signals are observed at the mixed positions with the 1H resonance of the complex a and the 15N resonance of the complex b (and vice versa; see Figure 7A), which are called ‘exchange cross peaks.’ These additional cross peaks arise due to protein translocation between the two DNA duplexes a and b during the mixing time period in which 15N nuclear magnetizations of interest remain along z, the direction of the outer magnetic field. Because the mixing time is present between the 15N and 1H evolution periods in the 15N z-exchange experiment, some proteins interact with DNA a during the 15N evolution period and interact with DNA b during the 1H evolution period, causing the exchange cross peak with the 15N resonance of the complex a and 1H resonance of the complex b. By analyzing signal intensities of the exchange cross peaks and the auto cross peaks as a function of the mixing time, the rate constants kab and kba for protein translocation processes between the two high-affinity sites can be determined. This translocation can occur through the dissociation & re-association mechanism or through the intersegment transfer mechanisms. By measuring the kab and kba rate constants at some different concentrations of free DNA duplexes, the kinetic contributions of these two mechanisms can be determined individually [87].
Figure 7.
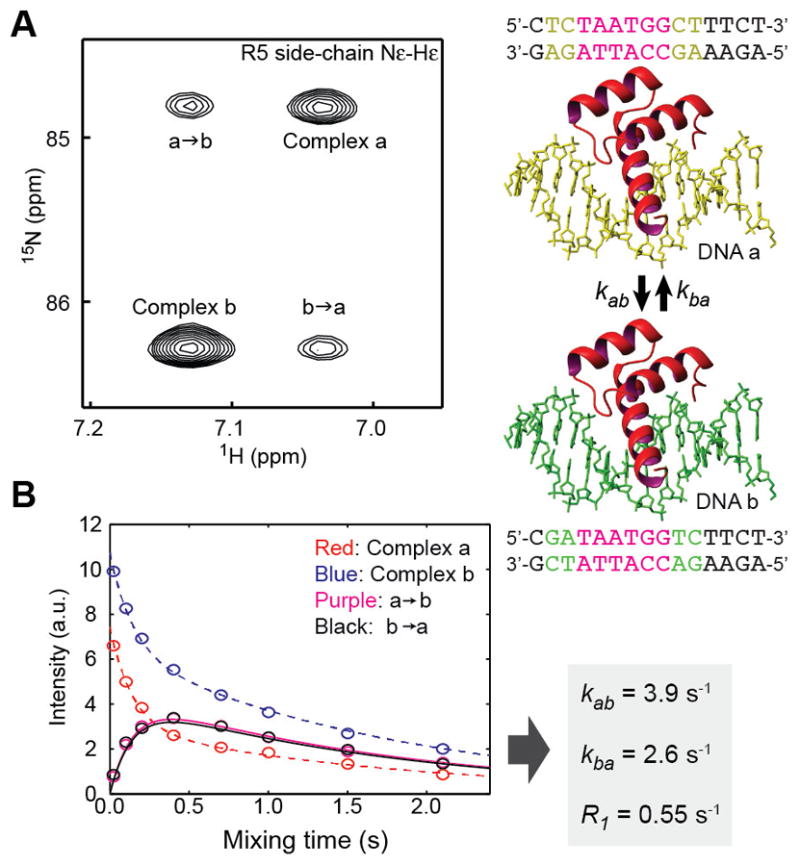
15N z-exchange data on kinetics of translocation of the Antp homeodomain between the two high-affinity sites on the DNA duplexes a and b [94]. (A) A 1H-15N spectrum recorded in 15N z-exchange experiment. (B) Intensities of the auto and exchange cross peaks as a function of the z-exchange mixing time. The rate constants determined from this dataset are also shown.
If the residence time of a protein on a high-affinity site on DNA is on the order of minutes or longer, the kinetics of translocation can be analyzed using a real-time NMR approach [96]. In this approach, a sample of protein-DNA complex is initially prepared. Then, another DNA of comparable affinity is added to the solution and the series of 2D NMR spectra are recorded to monitor translocation of the protein from the original DNA to the added DNA. By analyzing intensities of signals from the two complexes as a function of time, one can determine the kinetic rate constant for translocation between the two DNA duplexes. Fast acquisition methods (e.g. SOFAST-HMQC)[97–99], which allow recording of each 2D 1H-15N heteronuclear correlation spectrum within a minute, are useful to shorten the time interval for the real-time kinetics measurements. Translocation of the Egr-1 zinc-finger protein between two target sites was analyzed using NMR in this manner [96]. It should be noted that slow translocation processes over minutes-hours could readily be measured by biochemical assays requiring far less amounts of proteins and DNA. Nonetheless, when the translocation process turned out to be too slow to analyze with the 15N z-exchange method, NMR-based real-time kinetics approach could be convenient because the same set of materials can be readily tested.
5.4. Self-decoupling of intermolecular hydrogen-bond scalar couplings
The above-mentioned NMR methods for kinetic analysis of protein translocation on DNA require use of two DNA duplexes. Recently, a unique NMR-based kinetic approach that does not require two DNA duplexes has been proposed [94]. This NMR method utilizes intermolecular hydrogen-bond scalar couplings (h3JNP) between protein 15N and DNA 31P nuclei. Ionic-strength dependence of h3JNP could provide information on the residence time of protein at a high-affinity site on DNA [94]. This method does not require a mixture of two DNA duplexes. The observation of intermolecular hydrogen-bond scalar couplings is possible only if the residence time is sufficiently long (> ~10−2 s); otherwise, this coupling disappears through the process called self-decoupling (Figure 8A). Qualitatively, when an intermolecular hydrogen-bond scalar coupling is observed with a magnitude comparable to that of the intrinsic coupling constant hJ, the residence time of the complex should be longer than (2π|hJ|)−1. The intrinsic values of hydrogen-bond scalar couplings can be calculated from structural information by quantum chemical calculations or from the empirical relationship between the coupling constants and the hydrogen-bond geometry [100–102]. Detailed analysis of the self-decoupling of intermolecular hydrogen-bond scalar couplings as a function of ionic strength can provide more quantitative information about the residence time of the complex. For the Antp homeodomain, the exchange rates measured with the self-decoupling-based method were in good agreement with those measured with the 15N z-exchange methods (Figure 8B) [94]. This self-decoupling-based method is unique in that it does not require different signatures for the states involved in the exchange, although such conditions are typically crucial for other methods.
Figure 8.

Kinetic investigation of protein translocation on DNA through analysis of self-decoupling of intermolecular hydrogen-bond scalar coupling between protein 15N and DNA 31P nuclei [94]. (A) Theoretical relationship between the apparent value of intermolecular hydrogen-bond scalar coupling and the residence time of a molecular complex. Four cases simulated with different values of the intrinsic coupling constants are shown. (B) h3JNP coupling constants measured for the hydrogen bonds between Lys side-chain 15N and DNA phosphate 31P nuclei in the Antp homeodomain-DNA complex at some different salt concentrations. The h3JNP coupling constants were measured as described by Anderson et al. [31] (C) Comparison of the residence times of the Antp homeodomain at the recognition sequence measured with the h3JNP method and those measured with 15N z-exchange method.
6. NMR instruments
The above-mentioned NMR methods require a high-field NMR spectrometer (a magnetic field higher than 11 Tesla; 1H frequency ≥ 500 MH) equipped with a multi-channel probe capable of heteronuclear multidimensional experiments. Some quantitative NMR methods require data collection at multiple magnetic fields (e.g., 1H frequencies at 600 and 800 MHz). We use Bruker Avance III NMR spectrometers operated at 1H frequencies of 800, 750, and 600 MHz. Cryogenic 1H, 13C, 15N triple-resonance TCI (800 and 750 MHz) or 1H, 13C, 15N, 31P quadruple-resonance QCI (600 MHz) probes are typically used for these spectrometers. Cryogenic probes achieve ~3-fold higher signal-to-noise ratio in 1H or 13C NMR detection by cooling the detector to a cryogenic temperature for reduction in thermal noise, while retaining a physiological temperature (typically, 2 − 40°C) for measured samples. This high sensitivity is helpful for the quantitative NMR methods for investigating target DNA search processes of proteins. The 1H, 13C, 15N, 31P QCI cryogenic probe is particularly useful for studying protein-DNA interactions because it allows for precise measurements of intermolecular hydrogen-bond scalar couplings between DNA 31P and protein 15N nuclei [31, 32, 42, 44, 94, 103, 104]. Although this cryogenic probe has an additional coil for 31P nuclei, its sensitivity is almost as good as that of a corresponding TCI cryogenic probe (lower only by 11%, which can readily be compensated by a slightly larger number of scans).
7. Combining with other methods
Interpretation of NMR data on the target DNA search process can be greatly facilitated by integrating with other methods. In our lab, we routinely combine NMR and stopped-flow fluorescence data on the target DNA search processes. Our stopped-flow fluorescence kinetic methods allow us to obtain kinetic information on protein translocation on DNA via sliding, dissociation & re-association, and intersegment transfer mechanisms [22, 23]. The kinetic results from the NMR and fluorescence studies were consistent for the Egr-1 zinc-finger protein [13, 23]. The fluorescence data suggested that the Egr-1 zinc-finger protein spends ~1–10 μs at each nonspecific site and then slides to an adjacent site. This was also consistent with NMR observation that protein translocation on nonspecific DNA occurs in the fast exchange regime. The NMR and fluorescence methods can be applicable under the same buffer conditions and data obtained with these methods are complementary. Computational studies are also complementary. The coarse-grained molecular dynamics simulations showed that the search mode of Egr-1 facilitates intersegment transfer between nonspecific DNA duplexes (see Figure 4C) [13]. Applying these experimental and computational methods to the Egr-1 zinc-finger protein, we were able to validate the theoretical model involving the dynamic conformational equilibrium between the recognition and search modes during the target DNA search process [13, 44]. This model was originally proposed by some theoretical researchers to explain how transcription factors can simultaneously achieve two opposing factors: highly specific binding and sufficiently rapid search [105–107]. Through mutagenesis, we modulated the dynamic conformational equilibrium between the search and recognition modes and directly assessed the conformational shifts using NMR spectroscopy [44]. Using fluorescence and biochemical assays, we analyzed how the shifts of the conformational equilibrium influence binding affinity, target search kinetics, and efficiency in displacing other proteins from the target sites. A shift toward the recognition mode caused an increase in affinity for DNA and a decrease in search efficiency. In contrast, a shift toward the search mode caused a decrease in affinity and an increase in search efficiency. This demonstrated that target search by these proteins can be accelerated via engineering based on structural dynamic knowledge of the DNA-scanning process. Thus, NMR spectroscopy can help us deepen our knowledge of target DNA search by proteins and apply the knowledge to engineer proteins that can find targets more efficiently for artificial gene regulation or genome editing.
Highlights.
NMR approaches for investigating DNA-scanning processes of proteins are reviewed.
The NMR methods reveal dynamics of DNA scanning by proteins at an atomic level.
NMR also allows for quantitative kinetic analysis of protein translocation on DNA.
Advantages and disadvantages of NMR over other approaches are discussed.
Acknowledgments
This work was supported by Grant R01-GM107590 from the National Institutes of Health (to J.I.). We thank Ross Luu for proofreading. We also appreciate various supports from the Sealy Center for Structural Biology and Molecular Biophysics.
Abbreviations
- CSP
chemical shift perturbation
- PRE
paramagnetic relaxation enhancement
- RDC
residual dipolar coupling
- ZF
zinc finger
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Alberts B, Johnson A, Lewis J, Morgan D, Martin R, Roberts K, Walter P. DNA, chromosomes, and genomes, Molecular Biology of the Cell. Garland Science; New York: 2014. pp. 173–236. [Google Scholar]
- 2.Mirny L, Slutsky M, Wunderlich Z, Tafvizi A, Leith J, Kosmrlj A. How a protein searches for its site on DNA: the mechanism of facilitated diffusion. J Phys A: Math Theor. 2009;42(43):401335. [Google Scholar]
- 3.Bottani S, Veitia RA. Hill function-based models of transcriptional switches: impact of specific, nonspecific, functional and nonfunctional binding. Biol Rev Camb Philos Soc. 2017;92(2):953–963. doi: 10.1111/brv.12262. [DOI] [PubMed] [Google Scholar]
- 4.Crocker J, Noon EP, Stern DL. The Soft Touch: Low-Affinity Transcription Factor Binding Sites in Development and Evolution. Curr Top Dev Biol. 2016;117:455–69. doi: 10.1016/bs.ctdb.2015.11.018. [DOI] [PubMed] [Google Scholar]
- 5.Gerland U, Moroz JD, Hwa T. Physical constraints and functional characteristics of transcription factor-DNA interaction. Proceedings of the National Academy of Sciences U S A. 2002;99(19):12015–20. doi: 10.1073/pnas.192693599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lewin B. Genes VII. Oxford Univ Press; Oxford: 2000. [Google Scholar]
- 7.Valouev A, Johnson SM, Boyd SD, Smith CL, Fire AZ, Sidow A. Determinants of nucleosome organization in primary human cells. Nature. 2011;474(7352):516–20. doi: 10.1038/nature10002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Berg OG, Winter RB, von Hippel PH. Diffusion-driven mechanisms of protein translocation on nucleic acids. 1. Models and theory. Biochemistry. 1981;20(24):6929–48. doi: 10.1021/bi00527a028. [DOI] [PubMed] [Google Scholar]
- 9.Halford SE, Marko JF. How do site-specific DNA-binding proteins find their targets? Nucleic Acids Research. 2004;32(10):3040–3052. doi: 10.1093/nar/gkh624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kolomeisky AB. Physics of protein-DNA interactions: mechanisms of facilitated target search. Phys Chem Chem Phys. 2011;13(6):2088–95. doi: 10.1039/c0cp01966f. [DOI] [PubMed] [Google Scholar]
- 11.Von Hippel PH, Berg OG. Facilitated target location in biological systems. The Journal of Biological Chemistry. 1989;264(2):675–678. [PubMed] [Google Scholar]
- 12.Schonhoft JD, Stivers JT. Timing facilitated site transfer of an enzyme on DNA. Nat Chem Biol. 2012;8(2):205–10. doi: 10.1038/nchembio.764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Zandarashvili L, Vuzman D, Esadze A, Takayama Y, Sahu D, Levy Y, Iwahara J. Asymmetrical roles of zinc fingers in dynamic DNA-scanning process by the inducible transcription factor Egr-1. Proc Natl Acad Sci U S A. 2012;109(26):E1724–32. doi: 10.1073/pnas.1121500109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cuculis L, Schroeder CM. A Single-Molecule View of Genome Editing Proteins: Biophysical Mechanisms for TALEs and CRISPR/Cas9. Annu Rev Chem Biomol Eng. 2017;8:577–597. doi: 10.1146/annurev-chembioeng-060816-101603. [DOI] [PubMed] [Google Scholar]
- 15.Liu Z, Lavis LD, Betzig E. Imaging live-cell dynamics and structure at the single-molecule level. Mol Cell. 2015;58(4):644–59. doi: 10.1016/j.molcel.2015.02.033. [DOI] [PubMed] [Google Scholar]
- 16.Liu Z, Tjian R. Visualizing transcription factor dynamics in living cells. J Cell Biol. 2018 doi: 10.1083/jcb.201710038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Mueller F, Stasevich TJ, Mazza D, McNally JG. Quantifying transcription factor kinetics: at work or at play? Crit Rev Biochem Mol Biol. 2013;48(5):492–514. doi: 10.3109/10409238.2013.833891. [DOI] [PubMed] [Google Scholar]
- 18.Persson F, Barkefors I, Elf J. Single molecule methods with applications in living cells. Curr Opin Biotechnol. 2013;24(4):737–44. doi: 10.1016/j.copbio.2013.03.013. [DOI] [PubMed] [Google Scholar]
- 19.Presman DM, Ball DA, Paakinaho V, Grimm JB, Lavis LD, Karpova TS, Hager GL. Quantifying transcription factor binding dynamics at the single-molecule level in live cells. Methods. 2017;123:76–88. doi: 10.1016/j.ymeth.2017.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Silverstein TD, Gibb B, Greene EC. Visualizing protein movement on DNA at the single-molecule level using DNA curtains. DNA Repair (Amst) 2014;20:94–109. doi: 10.1016/j.dnarep.2014.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tafvizi A, Mirny LA, van Oijen AM. Dancing on DNA: kinetic aspects of search processes on DNA. Chemphyschem. 2011;12(8):1481–9. doi: 10.1002/cphc.201100112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Esadze A, Iwahara J. Stopped-flow fluorescence kinetic study of protein sliding and intersegment transfer in the target DNA search process. J Mol Biol. 2014;426(1):230–44. doi: 10.1016/j.jmb.2013.09.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Esadze A, Kemme CA, Kolomeisky AB, Iwahara J. Positive and negative impacts of nonspecific sites during target location by a sequence-specific DNA-binding protein: origin of the optimal search at physiological ionic strength. Nucleic Acids Res. 2014;42(11):7039–46. doi: 10.1093/nar/gku418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Esadze A, Rodriguez G, Weiser BP, Cole PA, Stivers JT. Measurement of nanoscale DNA translocation by uracil DNA glycosylase in human cells. Nucleic Acids Res. 2017;45(21):12413–12424. doi: 10.1093/nar/gkx848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Gowers DM, Wilson GG, Halford SE. Measurement of the contributions of 1D and 3D pathways to the translocation of a protein along DNA. Proc Natl Acad Sci U S A. 2005;102(44):15883–8. doi: 10.1073/pnas.0505378102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kemme CA, Esadze A, Iwahara J. Influence of quasi-specific sites on kinetics of target DNA search by a sequence-specific DNA-binding protein. Biochemistry. 2015;54(44):6684–91. doi: 10.1021/acs.biochem.5b00967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kemme CA, Marquez R, Luu RH, Iwahara J. Potential role of DNA methylation as a facilitator of target search processes for transcription factors through interplay with methyl-CpG-binding proteins. Nucleic Acids Res. 2017;45(13):7751–7759. doi: 10.1093/nar/gkx387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Porecha RH, Stivers JT. Uracil DNA glycosylase uses DNA hopping and short-range sliding to trap extrahelical uracils. Proc Natl Acad Sci U S A. 2008;105(31):10791–6. doi: 10.1073/pnas.0801612105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Rau DC, Sidorova NY. Diffusion of the restriction nuclease EcoRI along DNA. J Mol Biol. 2010;395(2):408–16. doi: 10.1016/j.jmb.2009.10.049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cavanagh J, Fairbrother JW, Palmer GA, Rance M, Skelton JN. Protein NMR Spectroscopy: Principles and Practice. 2. Elsevier Academic Press; 2007. [Google Scholar]
- 31.Anderson KM, Esadze A, Manoharan M, Brüschweiler R, Gorenstein DG, Iwahara J. Direct Observation of the Ion-Pair Dynamics at a Protein-DNA Interface by NMR Spectroscopy. J Am Chem Soc. 2013;135(9):3613–9. doi: 10.1021/ja312314b. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chen CY, Esadze A, Zandarashvili L, Nguyen D, Pettitt BM, Iwahara J. Dynamic Equilibria of Short-Range Electrostatic Interactions at Molecular Interfaces of Protein-DNA Complexes. Journal of Physical Chemistry Letters. 2015;6(14):2733–2737. doi: 10.1021/acs.jpclett.5b01134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Esadze A, Chen C, Zandarashvili L, Roy S, Pettitt BM, Iwahara J. Changes in conformational dynamics of basic side chains upon protein-DNA association. Nucleic Acids Res. 2016;44(14):6961–70. doi: 10.1093/nar/gkw531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Nguyen D, Hoffpauir ZA, Iwahara J. Internal Motions of Basic Side Chains of the Antennapedia Homeodomain in the Free and DNA-Bound States. Biochemistry. 2017;56(44):5866–5869. doi: 10.1021/acs.biochem.7b00885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Augustus AM, Reardon PN, Spicer LD. MetJ repressor interactions with DNA probed by in-cell NMR. Proc Natl Acad Sci U S A. 2009;106(13):5065–9. doi: 10.1073/pnas.0811130106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kato H, van Ingen H, Zhou BR, Feng H, Bustin M, Kay LE, Bai Y. Architecture of the high mobility group nucleosomal protein 2-nucleosome complex as revealed by methyl-based NMR. Proc Natl Acad Sci U S A. 2011;108(30):12283–8. doi: 10.1073/pnas.1105848108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kelly AE, Ou HD, Withers R, Dotsch V. Low-conductivity buffers for high-sensitivity NMR measurements. J Am Chem Soc. 2002;124(40):12013–9. doi: 10.1021/ja026121b. [DOI] [PubMed] [Google Scholar]
- 38.Lane AN, Arumugam S. Improving NMR sensitivity in room temperature and cooled probes with dipolar ions. J Magn Reson. 2005;173(2):339–43. doi: 10.1016/j.jmr.2005.01.005. [DOI] [PubMed] [Google Scholar]
- 39.Takeda M, Hallenga K, Shigezane M, Waelchli M, Lohr F, Markley JL, Kainosho M. Construction and performance of an NMR tube with a sample cavity formed within magnetic susceptibility-matched glass. J Magn Reson. 2011;209(2):167–73. doi: 10.1016/j.jmr.2011.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Voehler MW, Collier G, Young JK, Stone MP, Germann MW. Performance of cryogenic probes as a function of ionic strength and sample tube geometry. J Magn Reson. 2006;183(1):102–9. doi: 10.1016/j.jmr.2006.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Luger K, Rechsteiner TJ, Richmond TJ. Expression and purification of recombinant histones and nucleosome reconstitution. Methods Mol Biol. 1999;119:1–16. doi: 10.1385/1-59259-681-9:1. [DOI] [PubMed] [Google Scholar]
- 42.Zandarashvili L, Nguyen D, Anderson KM, White MA, Gorenstein DG, Iwahara J. Entropic Enhancement of Protein-DNA Affinity by Oxygen-to-Sulfur Substitution in DNA Phosphate. Biophys J. 2015;109(5):1026–37. doi: 10.1016/j.bpj.2015.07.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kalodimos CG, Biris N, Bonvin AM, Levandoski MM, Guennuegues M, Boelens R, Kaptein R. Structure and flexibility adaptation in nonspecific and specific protein-DNA complexes. Science. 2004;305(5682):386–9. doi: 10.1126/science.1097064. [DOI] [PubMed] [Google Scholar]
- 44.Zandarashvili L, Esadze A, Vuzman D, Kemme CA, Levy Y, Iwahara J. Balancing between affinity and speed in target DNA search by zinc-finger proteins via modulation of dynamic conformational ensemble. Proc Natl Acad Sci USA. 2015;112(37):E5142–E5149. doi: 10.1073/pnas.1507726112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Desjardins G, Okon M, Graves BJ, McIntosh LP. Conformational Dynamics and the Binding of Specific and Nonspecific DNA by the Autoinhibited Transcription Factor Ets-1. Biochemistry. 2016;55(29):4105–18. doi: 10.1021/acs.biochem.6b00460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.De S, Chan AC, Coyne HJ, 3rd, Bhachech N, Hermsdorf U, Okon M, Murphy ME, Graves BJ, McIntosh LP. Steric mechanism of auto-inhibitory regulation of specific and non-specific DNA binding by the ETS transcriptional repressor ETV6. J Mol Biol. 2014;426(7):1390–406. doi: 10.1016/j.jmb.2013.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Iwahara J, Schwieters CD, Clore GM. Characterization of Nonspecific Protein-DNA Interactions by 1H Paramagnetic Relaxation Enhancement. J Am Chem Soc. 2004;126(40):12800–12808. doi: 10.1021/ja046246b. [DOI] [PubMed] [Google Scholar]
- 48.Iwahara J, Zweckstetter M, Clore GM. NMR structural and kinetic characterization of a homeodomain diffusing and hopping on nonspecific DNA. Proc Natl Acad Sci U S A. 2006;103(41):15062–7. doi: 10.1073/pnas.0605868103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Takeuchi K, Imai M, Shimada I. Dynamic equilibrium on DNA defines transcriptional regulation of a multidrug binding transcriptional repressor, LmrR. Sci Rep. 2017;7(1):267. doi: 10.1038/s41598-017-00257-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Takayama Y, Clore GM. Intra- and intermolecular translocation of the bi-domain transcription factor Oct1 characterized by liquid crystal and paramagnetic NMR. Proc Natl Acad Sci U S A. 2011;108(22):E169–76. doi: 10.1073/pnas.1100050108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Takayama Y, Clore GM. Interplay between minor and major groove-binding transcription factors Sox2 and Oct1 in translocation on DNA studied by paramagnetic and diamagnetic NMR. J Biol Chem. 2012;287(18):14349–63. doi: 10.1074/jbc.M112.352864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Vandevenne M, Jacques DA, Artuz C, Nguyen CD, Kwan AH, Segal DJ, Matthews JM, Crossley M, Guss JM, Mackay JP. New insights into DNA recognition by zinc fingers revealed by structural analysis of the oncoprotein ZNF217. J Biol Chem. 2013;288(15):10616–27. doi: 10.1074/jbc.M112.441451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Sinha K, Sangani SS, Kehr AD, Rule GS, Jen-Jacobson L. Metal Ion Binding at the Catalytic Site Induces Widely Distributed Changes in a Sequence Specific Protein-DNA Complex. Biochemistry. 2016;55(44):6115–6132. doi: 10.1021/acs.biochem.6b00919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhou H, Purdy MM, Dahlquist FW, Reich NO. The recognition pathway for the DNA cytosine methyltransferase M.HhaI. Biochemistry. 2009;48(33):7807–16. doi: 10.1021/bi900502g. [DOI] [PubMed] [Google Scholar]
- 55.Friedman JI, Majumdar A, Stivers JT. Nontarget DNA binding shapes the dynamic landscape for enzymatic recognition of DNA damage. Nucleic Acids Res. 2009;37(11):3493–500. doi: 10.1093/nar/gkp161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Sun Y, Friedman JI, Stivers JT. Cosolute paramagnetic relaxation enhancements detect transient conformations of human uracil DNA glycosylase (hUNG) Biochemistry. 2011;50(49):10724–31. doi: 10.1021/bi201572g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Igumenova TI, Frederick KK, Wand AJ. Characterization of the fast dynamics of protein amino acid side chains using NMR relaxation in solution. Chem Rev. 2006;106(5):1672–99. doi: 10.1021/cr040422h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Kay LE. NMR studies of protein structure and dynamics. J Magn Reson. 2005;173(2):193–207. doi: 10.1016/j.jmr.2004.11.021. [DOI] [PubMed] [Google Scholar]
- 59.Loria JP, Berlow RB, Watt ED. Characterization of enzyme motions by solution NMR relaxation dispersion. Acc Chem Res. 2008;41(2):214–21. doi: 10.1021/ar700132n. [DOI] [PubMed] [Google Scholar]
- 60.Palmer AG., 3rd Nmr probes of molecular dynamics: overview and comparison with other techniques. Annu Rev Biophys Biomol Struct. 2001;30:129–55. doi: 10.1146/annurev.biophys.30.1.129. [DOI] [PubMed] [Google Scholar]
- 61.Elrod-Erickson M, Rould MA, Nekludova L, Pabo CO. Zif268 protein-DNA complex refined at 1.6 A: a model system for understanding zinc finger-DNA interactions. Structure. 1996;4(10):1171–80. doi: 10.1016/s0969-2126(96)00125-6. [DOI] [PubMed] [Google Scholar]
- 62.Pavletich NP, Pabo CO. Zinc finger-DNA recognition: crystal structure of a Zif268-DNA complex at 2.1 A. Science. 1991;252(5007):809–17. doi: 10.1126/science.2028256. [DOI] [PubMed] [Google Scholar]
- 63.Zandarashvili L, White MA, Esadze A, Iwahara J. Structural impact of complete CpG methylation within target DNA on specific complex formation of the inducible transcription factor Egr-1. FEBS Lett. 2015;589(15):1748–53. doi: 10.1016/j.febslet.2015.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bax A. Weak alignment offers new NMR opportunities to study protein structure and dynamics. Protein Sci. 2003;12(1):1–16. doi: 10.1110/ps.0233303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Jensen MR, Markwick PR, Meier S, Griesinger C, Zweckstetter M, Grzesiek S, Bernado P, Blackledge M. Quantitative determination of the conformational properties of partially folded and intrinsically disordered proteins using NMR dipolar couplings. Structure. 2009;17(9):1169–85. doi: 10.1016/j.str.2009.08.001. [DOI] [PubMed] [Google Scholar]
- 66.Lipsitz RS, Tjandra N. Residual dipolar couplings in NMR structure analysis. Annu Rev Biophys Biomol Struct. 2004;33:387–413. doi: 10.1146/annurev.biophys.33.110502.140306. [DOI] [PubMed] [Google Scholar]
- 67.Tolman JR. Dipolar couplings as a probe of molecular dynamics and structure in solution. Curr Opin Struct Biol. 2001;11(5):532–9. doi: 10.1016/s0959-440x(00)00245-1. [DOI] [PubMed] [Google Scholar]
- 68.Zweckstetter M. NMR: prediction of molecular alignment from structure using the PALES software. Nat Protoc. 2008;3(4):679–90. doi: 10.1038/nprot.2008.36. [DOI] [PubMed] [Google Scholar]
- 69.Zweckstetter M, Hummer G, Bax A. Prediction of charge-induced molecular alignment of biomolecules dissolved in dilute liquid-crystalline phases. Biophys J. 2004;86(6):3444–60. doi: 10.1529/biophysj.103.035790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Clore GM. Exploring translocation of proteins on DNA by NMR. J Biomol NMR. 2011;51(3):209–19. doi: 10.1007/s10858-011-9555-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Clore GM. Practical Aspects of Paramagnetic Relaxation Enhancement in Biological Macromolecules. Methods Enzymol. 2015;564:485–97. doi: 10.1016/bs.mie.2015.06.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Clore GM, Iwahara J. Theory, Practice, and Applications of Paramagnetic Relaxation Enhancement for the Characterization of Transient Low-Population States of Biological Macromolecules and Their Complexes. Chem Rev. 2009;109(9):4108–4139. doi: 10.1021/cr900033p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Iwahara J, Anderson DE, Murphy EC, Clore GM. EDTA-derivatized deoxythymidine as a tool for rapid determination of protein binding polarity to DNA by intermolecular paramagnetic relaxation enhancement. J Am Chem Soc. 2003;125(22):6634–5. doi: 10.1021/ja034488q. [DOI] [PubMed] [Google Scholar]
- 74.Cai S, Zhu L, Zhang Z, Chen Y. Determination of the three-dimensional structure of the Mrf2-DNA complex using paramagnetic spin labeling. Biochemistry. 2007;46(17):4943–50. doi: 10.1021/bi061738h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Qin PZ, Hideg K, Feigon J, Hubbell WL. Monitoring RNA base structure and dynamics using site-directed spin labeling. Biochemistry. 2003;42(22):6772–83. doi: 10.1021/bi027222p. [DOI] [PubMed] [Google Scholar]
- 76.Ramos A, Varani G. A new method to detect long-range protein-RNA contacts: NMR detection of electron-proton relaxation induced by nitroxide spin-labeled RNA. J Am Chem Soc. 1998;120(42):10992–10993. [Google Scholar]
- 77.Iwahara J, Schwieters CD, Clore GM. Ensemble approach for NMR structure refinement against (1)H paramagnetic relaxation enhancement data arising from a flexible paramagnetic group attached to a macromolecule. J Am Chem Soc. 2004;126(18):5879–96. doi: 10.1021/ja031580d. [DOI] [PubMed] [Google Scholar]
- 78.Iwahara J, Tang C, Clore GM. Practical aspects of 1H transverse paramagnetic relaxation enhancement measurements on macromolecules. Journal of Magnetic Resonance. 2007;184(2):185–195. doi: 10.1016/j.jmr.2006.10.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Iwahara J, Clore GM. Detecting transient intermediates in macromolecular binding by paramagnetic NMR. Nature. 2006;440(7088):1227–1230. doi: 10.1038/nature04673. [DOI] [PubMed] [Google Scholar]
- 80.Gong Z, Gu XH, Guo DC, Wang J, Tang C. Protein Structural Ensembles Visualized by Solvent Paramagnetic Relaxation Enhancement. Angew Chem Int Ed Engl. 2017;56(4):1002–1006. doi: 10.1002/anie.201609830. [DOI] [PubMed] [Google Scholar]
- 81.Gu XH, Gong Z, Guo DC, Zhang WP, Tang C. A decadentate Gd(III)-coordinating paramagnetic cosolvent for protein relaxation enhancement measurement. J Biomol NMR. 2014;58(3):149–54. doi: 10.1007/s10858-014-9817-3. [DOI] [PubMed] [Google Scholar]
- 82.Hernandez G, Teng CL, Bryant RG, LeMaster DM. O2 penetration and proton burial depth in proteins: applicability to fold family recognition. J Am Chem Soc. 2002;124(16):4463–72. doi: 10.1021/ja017340k. [DOI] [PubMed] [Google Scholar]
- 83.Hocking HG, Zangger K, Madl T. Studying the structure and dynamics of biomolecules by using soluble paramagnetic probes. Chemphyschem. 2013;14(13):3082–94. doi: 10.1002/cphc.201300219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Pintacuda G, Otting G. Identification of protein surfaces by NMR measurements with a pramagnetic Gd(III) chelate. J Am Chem Soc. 2002;124(3):372–3. doi: 10.1021/ja016985h. [DOI] [PubMed] [Google Scholar]
- 85.Williamson MP. Using chemical shift perturbation to characterise ligand binding. Prog Nucl Magn Reson Spectrosc. 2013;73:1–16. doi: 10.1016/j.pnmrs.2013.02.001. [DOI] [PubMed] [Google Scholar]
- 86.Doucleff M, Clore GM. Global jumping and domain-specific intersegment transfer between DNA cognate sites of the multidomain transcription factor Oct-1. Proc Natl Acad Sci U S A. 2008;105(37):13871–13876. doi: 10.1073/pnas.0805050105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Iwahara J, Clore GM. Direct observation of enhanced translocation of a homeodomain between DNA cognate sites by NMR exchange spectroscopy. J Am Chem Soc. 2006;128(2):404–405. doi: 10.1021/ja056786o. [DOI] [PubMed] [Google Scholar]
- 88.Sahu D, Clore GM, Iwahara J. TROSY-Based z-Exchange Spectroscopy: Application to the Determination of the Activation Energy for Intermolecular Protein Translocation between Specific Sites on Different DNA Molecules. J Am Chem Soc. 2007;129(43):13232–13237. doi: 10.1021/ja074604f. [DOI] [PubMed] [Google Scholar]
- 89.Sahu D, Iwahara J. Discrete-State Kinetics Model for NMR-Based Analysis of Protein Translocation on DNA at Equilibrium. J Phys Chem B. 2017;121(41):9548–9556. doi: 10.1021/acs.jpcb.7b07779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Takayama Y, Clore GM. Impact of protein/protein interactions on global intermolecular translocation rates of the transcription factors Sox2 and Oct1 between DNA cognate sites analyzed by z-exchange NMR spectroscopy. J Biol Chem. 2012;287(32):26962–70. doi: 10.1074/jbc.M112.382960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Farrow NA, Zhang O, Forman-Kay JD, Kay LE. A heteronuclear correlation experiment for simultaneous determination of 15N longitudinal decay and chemical exchange rates of systems in slow equilibrium. J Biomol NMR. 1994;4(5):727–734. doi: 10.1007/BF00404280. [DOI] [PubMed] [Google Scholar]
- 92.Li Y, Palmer AG., 3rd TROSY-selected ZZ-exchange experiment for characterizing slow chemical exchange in large proteins. J Biomol NMR. 2009;45(4):357–60. doi: 10.1007/s10858-009-9385-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Walavalkar NM, Cramer JM, Buchwald WA, Scarsdale JN, Williams DC., Jr Solution structure and intramolecular exchange of methyl-cytosine binding domain protein 4 (MBD4) on DNA suggests a mechanism to scan for mCpG/TpG mismatches. Nucleic Acids Res. 2014;42(17):11218–32. doi: 10.1093/nar/gku782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Zandarashvili L, Esadze A, Kemme CA, Chattopadhyay A, Nguyen D, Iwahara J. Residence Times of Molecular Complexes in Solution from NMR Data of Intermolecular Hydrogen-Bond Scalar Coupling. J Phys Chem Lett. 2016;7(5):820–4. doi: 10.1021/acs.jpclett.6b00019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Ryu KS, Tugarinov V, Clore GM. Probing the rate-limiting step for intramolecular transfer of a transcription factor between specific sites on the same DNA molecule by (15)Nz-exchange NMR spectroscopy. J Am Chem Soc. 2014;136(41):14369–72. doi: 10.1021/ja5081585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Takayama Y, Sahu D, Iwahara J. NMR Studies of Translocation of the Zif268 Protein between Its Target DNA Sites. Biochemistry. 2010;49(37):7998–8005. doi: 10.1021/bi100962h. [DOI] [PubMed] [Google Scholar]
- 97.Deschamps M, Campbell ID. Cooling overall spin temperature: protein NMR experiments optimized for longitudinal relaxation effects. J Magn Reson. 2006;178(2):206–11. doi: 10.1016/j.jmr.2005.09.011. [DOI] [PubMed] [Google Scholar]
- 98.Schanda P, Brutscher B. Very fast two-dimensional NMR spectroscopy for real-time investigation of dynamic events in proteins on the time scale of seconds. J Am Chem Soc. 2005;127(22):8014–5. doi: 10.1021/ja051306e. [DOI] [PubMed] [Google Scholar]
- 99.Schanda P, Kupce E, Brutscher B. SOFAST-HMQC experiments for recording two-dimensional heteronuclear correlation spectra of proteins within a few seconds. J Biomol NMR. 2005;33(4):199–211. doi: 10.1007/s10858-005-4425-x. [DOI] [PubMed] [Google Scholar]
- 100.Barfield M. Structural dependencies of interresidue scalar coupling h3JNC, and donor H-1 chemical shifts in the hydrogen bonding regions of proteins. J Am Chem Soc. 2002;124(15):4158–4168. doi: 10.1021/ja012674v. [DOI] [PubMed] [Google Scholar]
- 101.Czernek J, Brüschweiler R. Geometric dependence of 3hJ(31P-15N) and 2hJ(31P-1H) scalar couplings in protein-nucleotide complexes. J Am Chem Soc. 2001;123(44):11079–80. doi: 10.1021/ja011618r. [DOI] [PubMed] [Google Scholar]
- 102.Grzesiek S, Cordier F, Jaravine V, Barfield M. Insights into biomolecular hydrogen bonds from hydrogen bond scalar couplings. Prog in NMR Spect. 2004;45(3–4):275–300. [Google Scholar]
- 103.Anderson KM, Nguyen D, Esadze A, Zandrashvili L, Gorenstein DG, Iwahara J. A chemical approach for site-specific identification of NMR signals from protein side-chain NH3+ groups forming intermolecular ion pairs in protein-nucleic acid complexes. J Biomol NMR. 2015;62(1):1–5. doi: 10.1007/s10858-015-9909-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Chattopadhyay A, Esadze A, Roy S, Iwahara J. NMR Scalar Couplings across Intermolecular Hydrogen Bonds between Zinc-Finger Histidine Side Chains and DNA Phosphate Groups. J Phys Chem B. 2016;120:10679–10685. doi: 10.1021/acs.jpcb.6b08137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Murugan R. Theory of site-specific DNA-protein interactions in the presence of conformational fluctuations of DNA binding domains. Biophys J. 2010;99(2):353–9. doi: 10.1016/j.bpj.2010.04.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Slutsky M, Mirny LA. Kinetics of Protein-DNA Interaction: Facilitated Target Location in Sequence-Dependent Potential. Biophys J. 2004;87(6):4021–4035. doi: 10.1529/biophysj.104.050765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Zhou HX. Rapid search for specific sites on DNA through conformational switch of nonspecifically bound proteins. Proc Natl Acad Sci U S A. 2011;108(21):8651–6. doi: 10.1073/pnas.1101555108. [DOI] [PMC free article] [PubMed] [Google Scholar]