

Abstract
Background
Rare data are available on the molecular responses of higher plants to low pH. Seedlings of ‘Sour pummelo’ (Citrus grandis) and ‘Xuegan’ (Citrus sinensis) were treated daily with nutrient solution at a pH of 2.5, 3, or 6 (control) for nine months. Thereafter, we first used 2-dimensional electrophoresis (2-DE) to investigate low pH-responsive proteins in Citrus leaves. Meanwhile, we examined low pH-effects on leaf gas exchange, carbohydrates, ascorbate, dehydroascorbate and malondialdehyde. The objectives were to understand the adaptive mechanisms of Citrus to low pH and to identify the possible candidate proteins for low pH-tolerance.
Results
Our results demonstrated that Citrus were tolerant to low pH, with a slightly higher low pH-tolerance in the C. sinensis than in the C. grandis. Using 2-DE, we identified more pH 2.5-responsive proteins than pH 3-responsive proteins in leaves. This paper discussed mainly on the pH 2.5-responsive proteins. pH 2.5 decreased the abundances of proteins involved in ribulose bisphosphate carboxylase/oxygenase activation, Calvin cycle, carbon fixation, chlorophyll biosynthesis and electron transport, hence lowering chlorophyll level, electron transport rate and photosynthesis. The higher oxidative damage in the pH 2.5-treated C. grandis leaves might be due to a combination of factors including higher production of reactive oxygen species, more proteins decreased in abundance involved in antioxidation and detoxification, and lower ascorbate level. Protein and amino acid metabolisms were less affected in the C. sinensis leaves than those in the C. grandis leaves when exposed to pH 2.5. The abundances of proteins related to jasmonic acid biosynthesis and signal transduction were increased and decreased in the pH 2.5-treated C. sinensis and C. grandis leaves, respectively.
Conclusions
This is the first report on low pH-responsive proteins in higher plants. Thus, our results provide some novel information on low pH-toxicity and -tolerance in higher plants.
Electronic supplementary material
The online version of this article (10.1186/s12870-018-1413-3) contains supplementary material, which is available to authorized users.
Keywords: Citrus grandis, Citrus sinensis, 2-DE, Leaves, Low pH, Proteomics
Background
Soil acidity is a major factor limiting crop and productivity in many parts of the world, with up to 30% of the world’s ice-free land and 12% of crops affected by soil acidity [1]. What’s worse, soil pH is rapidly decreasing due to acid rain, soil leaching, intensive agriculture and monoculture, poor nutrient cycling, and the acidifying effects of nitrogen (N) fertilizer [2–5].
Usually, multiple stress factors including toxicities of H+, aluminum (Al) and manganese (Mn), lack of nutrients, namely N, phosphorus (P), potassium (K), magnesium (Mg), calcium (Ca) and molybdenum (Mo), decreased uptake of water, and toxic level of phenolic acids are considered to be responsible for poor growth and yield loss of crops on acidic soils [1, 6–8]. Recently, many researchers have paid attention to Al-toxicity and -tolerance, but few data are available on low pH (H+) damage and adaptation in plants [9–11]. Evidence shows that the adaptation of plants to H+ and Al are regulated by separate mechanisms [7, 8, 12]. Obviously, additional research on low pH adaptation is needed in order to a better understanding of plant adaptation to acid soils [13].
In addition to inhibiting directly or indirectly plant growth and development, low pH (high H+) has negative influences on cellular structure and functions, and physiological and biochemical processes, including the uptake of water and nutrients [8, 14], leaf gas exchange [8, 10, 15], chlorophyll (Chl) biosynthesis, Chl a fluorescence [8, 11, 15, 16], reactive oxygen species (ROS) production and detoxification [4, 16–18], membrane integrity [19], and cell wall structure and functions [20, 21]. Because low pH can inhibit photosynthesis and growth in some higher plants [8, 10, 15], carbohydrates should be altered by low pH. To our best knowledge, such data are very rare.
Although some workers have investigated the physiological and biochemical responses of higher plants to low pH [8, 22, 23], rare data are available on the molecular responses until recently [24]. In a study, Lager et al. investigated the effects of pH on gene expression in roots of Arabidopsis thaliana shifted from a nutrition solution of pH 6 to one of pH 4.5 for 1 h and 8 h, and obtained a total of 277 ‘early-responsive genes’, namely ‘1 h responsive genes’ and a total of 748 ‘late-responsive genes’, namely ‘8 h responsive genes’. The major alterations of gene expression in response to low pH were associated with Ca2+ signaling and cell wall modifications [24]. Howbeit these transcriptome data are very useful, great difference exists between protein level and mRNA level because the abundance of a protein is determined not only by the transcriptional rate of the gene, but also by the transcript stability, nuclear export and location, translational regulation and protein degradation [25, 26]. Because proteins are the ultimate controllers for biological processes, it is imperative to conduct a proteomic analysis in order to fully understand the molecular responses of higher plants to low pH. To our knowledge, data on low pH-responsive proteins in higher plants are very scanty.
Citrus can be cultivated in soils covering a wide range of pH and are tolerant to acidic soils [27]. Recently, we used sand culture to investigate the effects of pH 2.5, 3, 4, 5 and 6 on growth, nutrients, relative water content (RWC), specific leaf weight, total soluble proteins, H2O2 production, electrolyte leakage, photosynthesis and related physiological parameters in C. grandis and C. sinensis seedlings. pH 2.5 greatly inhibited seedling growth; pH 3 slightly inhibited growth; and pH 4 had almost no influence on growth. In addition, most of these parameters [i.e., leaf CO2 assimilation, Chl levels, ribulose bisphosphate carboxylase/oxygenase (Rubisco) activity, overwhelming majority of Chl a fluorescence parameters and specific leaf weight; root and leaf RWC and electrolyte leakage; and root, stem and leaf N and K concentrations] were altered only at pH 2.5, with slightly greater changes in the C. grandis seedlings than those in the C. sinensis seedlings. Evidently, C. grandis and C. sinensis were tolerant to low pH, and the latter was slightly more tolerant to low pH [8]. Most of soils used for Citrus production in China are acidic and strong acidic. Moreover, Citrus orchard soil pH is rapidly decreasing in the last decade [28].
In this study, we first used a 2-dimensional electrophoresis (2-DE)-based mass spectrometry (MS) approach to investigate low pH-responsive proteins in C. sinensis and C. grandis leaves. Also, we examined low pH-effects on leaf gas exchange, carbohydrates, ascorbate (ASC), dehydroascorbate (DHA) and malondialdehyde (MDA). The objectives were (a) to understand the adaptive mechanisms of Citrus to low pH and (b) to identify the possible candidate proteins for tolerance to low pH in Citrus.
Methods
Citrus seedling culture and pH treatments
Seedling culture and pH treatments were carried out according to Long et al. [8]. Briefly, four week-old uniform seedlings of ‘Xuegan’ (C. sinensis) and ‘Sour pummelo’ (C. grandis) with single stem were chosen and transplanted to 6 L pots (two seedlings per pot) filled with ~ 0.6 cm in diameter clean river sand washed thoroughly with tap water, then grown in a glasshouse under natural photoperiod at Fujian Agriculture and Forestry University (FAFU), Fuzhou (26°5’ N, 119°14′), China, until the end of the experiment. Seven weeks after transplanting, each pot was supplied daily with nutrient solution containing 2.5 mM KNO3, 2.5 mM Ca(NO3)2, 1 mM MgSO4, 0.5 mM KH2PO4, 20 μM Fe-EDTA, 10 μM H3BO3, 2 μM ZnSO4, 2 μM MnCl2, 0.5 μM CuSO4 and 0.065 μM (NH4)6Mo7O24 until dripping (~ 500 mL) at a pH of 6 (control), 3 or 2.5 (adjusted by 1 M H2SO4) for nine months, which were selected based on our preliminary experiment and previous study [8] and were suitable for physiological and proteomic analysis. In this study, we focused mainly on the long-term changes that allow homeostatic adjustment to low pH and on the long-term consequences of low pH because there is an opportunity to extend Citrus cultivation to acidic soils. No any precipitates were formed in the nutrient solution. In addition, we measured the concentrations of macroelements (N, P, K, Ca, Mg and S) in the nutrient solution. Analytic results showed that pH did not affect their solubility. Thereafter, recent fully expanded (~ 7-week-old) leaves were used for all measurements. After leaf gas exchange being determined, leaves (midribs, petioles and winged leaves removed) and leaf discs (0.6 cm in diameter) from the same seedlings were harvested at sunny noon and frozen in liquid N2, then stored at − 80 °C until they were used for the extract of proteins.
Leaf nonstructural carbohydrate, ASC, DHA and malondialdehyde
Leaf fructose, glucose, sucrose and starch were assayed using enzymatic methods as previously described by Han et al. [29]. Leaf ASC and DHA were measured using enzymatic methods after being extracted with 6% (v/v) of HClO4 [30]. Leaf malondialdehyde (MDA) was measured as thiobarbituric acid-reactive substances after being extracted with 80% (v/v) of ethanol [31].
Leaf gas exchange
Leaf gas exchange was determined with a CIARS-2 portable photosynthesis system (PP systems, Herts, UK) at a controlled CO2 concentration of ~ 380 μmol mol− 1, a controlled light intensity of ~ 1000 μmol m− 2 s− 1, a relative humidity of 64.0 ± 0.6% and a leaf temperature of 30.8 ± 0.2 °C, between 9 and 11 a.m. on a sunny day.
Leaf protein extraction, 2-DE and image analysis
In order to reduce errors and get reliable and reproducible results, ~ 1 g frozen leaves from four seedlings (one seedling per pot, equal amount of sample per seedling) were mixed as one biological replicate. There were three replicates per treatment (a total of 12 seedlings from 12 pots). Proteins were independently extracted thrice from pH 2.5-, 3- and 6-treated samples using a phenol extraction procedure as described previously [32] and their concentrations were determined according to Bradford [33]. Both 2-DE and image analysis were performed as described by Sang et al. [34, 35] and Yang et al. [36]. Background subtraction, Gaussian fitting, gel alignment, spot detection, matching and normalization were made with PDQuest version 8.0.1 (Bio-Rad, Hercules, CA, USA). A protein spot was considered differentially abundant when it had both a P-value < 0.05 by ANOVA and a fold change > 1.5. All these differentially abundant protein (DAP) spots were visually checked and excised for identification by MALDI-TOF/TOF-MS.
Protein identification by MALDI-TOF/TOF-MS and bioinformatic analysis
MALDI-TOF/TOF-MS-based protein identification was conducted on an AB SCIEX 5800 TOF/TOF plus MS (AB SCIEX, Shanghai, China) as described previously [26, 34]. All acquired spectra of samples were processed using TOF/TOF Explorer™ Software (AB SCIEX, Shanghai, China) in a default mode. The data were searched by GPS Explorer (Version 3.6) with the search engine MASCOT (Version 2.3, Matrix Science Inc., Boston, MA) against the C. sinensis databases (http://citrus.hzau.edu.cn/orange/index.php). The search parameters were as follows: trypsin cleavage with one missed, MS tolerance of 100 ppm, and MS/MS tolerance of 0.6 Da. At least two peptides were required to match for each protein. Protein identifications were accepted if MASCOT score was not less than 70, and the number of matched peptides (NMP) was not less than five or the sequence coverage was not less than 20% [35, 37]. Functional categories of DAPs were assigned according to Kyoto Encyclopedia of Genes and Genomes (KEGG; http://www.kegg.jp/), Uniprot (http://www.uniprot.org/) and gene ontology (GO; http://www.geneontology.org/) databases [38, 39].
qRT-PCR analysis
About 300 mg frozen leaves from four seedlings (one seedling per pot, equal amount of sample per seedling) were pooled as one biological replicate. qRT-PCR was made with three biological and two technical replicates for each treatment (a total of 12 seedlings from 12 pots) as described by Zhou et al. [40]. Here, we randomly selected a total of 26 DAPs from the pH 2.5-treated C. sinensis (i.e., S1, S4, S5, S6, S7, S9, S10, S15, S23, S26, S27, S35 and S40) and C. grandis (i.e., G2, G5, G6, G13, G14, G15, G16, G19, G21, G36, G37, G40, and G41) leaves for qRT-PCR. Specific primers were designed from the corresponding sequences of these selected DAPs in Citrus genome (http://citrus.hzau.edu.cn/orange/index.php) using Primer Primier Version 5.0 (PREMIER Biosoft International, CA, USA). The sequences of the F and R primers used were listed in Additional file 1: Table S1. For the normalization of gene expression and reliability of quantitative analysis, two Citrus genes: actin (Ciclev10025866m) and U4/U6 small nuclear ribonucleoprotein PRP31 (PRPF31; Ciclev10031363m), were selected as internal standards and the leaves from the pH 6-treated seedlings were used as reference sample, which was set to 1.
Data analysis
There were 20 pots (40 seedlings) per treatment in a completely randomized design. Experiments were performed with 3–8 replicates. Results represented the mean ± SE. Significant differences among the six treatment combinations were analyzed by two (species) × three (pH levels) ANOVA, and the six means were separated by the Turkey test at P < 0.05.
Results
Physiological and biochemical responses to low pH
Leaf CO2 assimilation and stomatal conductance were significantly lower at pH 2.5 than at pH 3 or pH 6. Intercellular CO2 concentration was similar among the six treatment combinations except that it was slightly higher in the 2.5-treated C. grandis leaves than that in the 3-treated C. grandis and C. sinensis leaves (Fig. 1). Based on the stomatal limited theory in photosynthesis [41], the pH 2.5-induced inhibition of photosynthesis was not explained alone by stomatal limitation. CO2 assimilation, stomatal conductance and intercellular CO2 concentration were similar between C. grandis and C. sinensis leaves (Fig. 1).
Fig. 1.
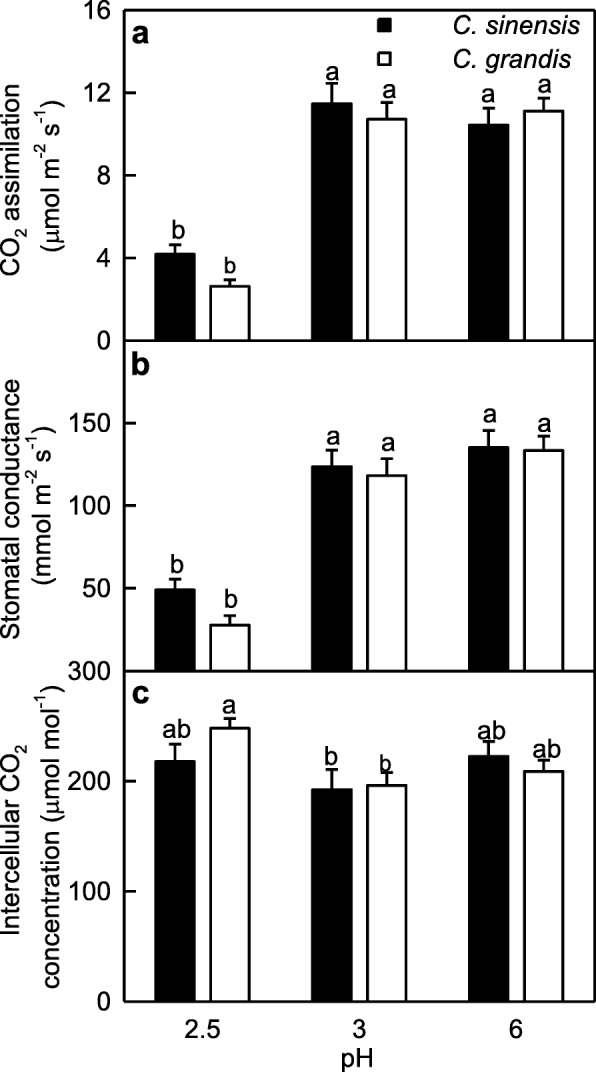
Leaf gas exchange in response to low pH. a CO2 assimilation; b stomatal conductance; c intercellular CO2 concentration. Bars represent means ± SE (n = 5). Different letters above the bars indicate a significant difference at P < 0.05
As shown in Fig. 2, the levels of glucose, fructose, sucrose, total soluble sugars (the summation of glucose, fructose and sucrose), starch, and total nonstructural carbohydrates (TNC, the summation of glucose, fructose, sucrose and starch) in the C. grandis and C. sinensis leaves were elevated at pH 2.5, but unaffected at pH 3. The only exception was that sucrose levels in the C. sinensis leaves did not change as pH decreased from 6 to 2.5. The concentrations of all these nonstructural carbohydrates were higher in the C. grandis leaves than those in the C. sinensis leaves or similar between the two at each given pH with the exceptions that glucose and fructose concentrations were higher in the C. sinensis leaves than those in the C. grandis leaves at pH 2.5.
Fig. 2.

Leaf concentrations of nonstructural carbohydrates in response to low pH. a glucose; b fructose; c sucrose; d total soluble sugars (the summation of glucose, fructose and sucrose); e starch; f total nonstructural carbohydrates (TNC; the summation of glucose, fructose, sucrose and starch). Bars represent means ± SE (n = 8). Different letters above the bars indicate a significant difference at P < 0.05
As shown in Fig. 3, only pH 2.5 decreased the levels of ASC + DHA and ASC, and the ratio of ASC/(ASC + DHA) in C. grandis and C. sinensis leaves, especially in the C. grandis leaves. The levels of ASC + DHA and ASC, and the ratio of ASC/(ASC + DHA) were higher in the C. sinensis leaves than those in the C. grandis leaves at pH 2.5, but similar between the two at pH 3 or pH 6. Interestingly, leaf DHA level did not significantly differ among the six treatment combinations.
Fig. 3.

Leaf ASC + DHA, ASC and DHA concentrations, and ASC/(ASC + DHA) ratio in response to low pH. Bars represent means ± SE (n = 6–8). Different letters above the bars indicate a significant difference at P < 0.05
MDA concentrations in C. grandis and C. sinensis leaves increased as pH decreased from 6 to 2.5. MDA concentrations were higher in the C. grandis leaves than that in the C. sinensis leaves at pH 2.5, but similar between the two at pH 6 or pH 3 (Fig. 4).
Fig. 4.

Leaf MDA concentration in response to low pH. Bars represent means ± SE (n = 7–8). Different letters above the bars indicate a significant difference at P < 0.05
Protein yield and low pH-responsive proteins in leaves
To obtain reliable results, three biological replicates were conducted in this experiment (Fig. 5 and Additional file 2: Figure S1). As shown in Table 1, protein yields and the number of protein spots per gel did not significantly differ among the six treatment combinations (Table 1, Fig. 5 and Additional file 2: Figure S1).
Fig. 5.

Representative 2-DE images of proteins extracted from C. grandis (a-c) and C. sinensis (d-f) leaves
Table 1.
Protein yield, number of variable spots and number of identified DAP spots in C. sinensis and C. grandis leaves
| C. sinensis | C. grandis | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| pH 2.5 | pH 3 | pH 6 | pH 2.5 | pH 3 | pH 6 | |||||
| Protein yield (mg g−1 FW) | 16.4 ± 0.24a | 15.57 ± 0.09a | 15.97 ± 0.35a | 14.35 ± 0.19a | 14.04 ± 0.66a | 14.63 ± 0.10a | ||||
| Number of spots per gel | 714 ± 12a | 717 ± 4a | 733 ± 14a | 727 ± 13a | 737 ± 9a | 755 ± 11a | ||||
| Only present at pH 2.5 | Only present at pH 3 | Present at both pH 2.5 and 3 | Total | Only present at pH 2.5 | Only present at pH 3 | Present at both pH 2.5 and 3 | Total | |||
| Number of DAP spots | ||||||||||
| Increased in abundance | 15 | 0 | 1 | 16 | 5 | 0 | 3 | 8 | ||
| Decreased in abundance | 29 | 1 | 6 | 36 | 14 | 1 | 23 | 38 | ||
| Increased in abundance at pH 2.5 and decreased in abundance at pH 3 | 2 | 2 | 1 | 1 | ||||||
| Decreased in abundance at pH 2.5 and increased in abundance at pH 3 | 1 | 1 | 1 | 1 | ||||||
| Total | 44 | 1 | 10 | 55 | 19 | 1 | 28 | 48 | ||
| Number of identified DAP spots | ||||||||||
| Increased in abundance | 13 | 0 | 1 | 14 | 5 | 0 | 3 | 8 | ||
| Decreased in abundance | 26 | 0 | 6 | 32 | 12 | 1 | 21 | 34 | ||
| Increased in abundance at pH 2.5 and decreased in abundance at pH 3 | 2 | 2 | 1 | 1 | ||||||
| Decreased in abundance at pH 2.5 and increased in abundance at pH 3 | 1 | 1 | 1 | 1 | ||||||
| Total | 39 | 0 | 10 | 49 | 17 | 1 | 26 | 44 | ||
Data are means ± SE (n = 3). Different letters within a row indicate significant differences at P < 0.05
A protein spot having an average fold change > 1.5 and a P-value < 0.05 was considered as differentially abundant. Based on the two criteria, we obtained 55 and 48 DAP spots from the pH 2.5 and/or pH 3-treated C. sinensis and C. grandis leaves, respectively. After submitting these DAP spots to MALDI-TOF/TOF-MS-based identification, we identified 49 and 44 DAP spots in the pH 2.5 and/or pH 3-treated C. sinensis and C. grandis leaves, responsively (Tables 1, 2 and 3, Fig. 5 and Additional file 3: Table S2, Additional file 4: Table S3, Additional file 5: Figure S2). For C. sinensis, 39 identified DAP spots only presented in pH 2.5-treated leaves, and 10 identified DAP spots with the same accession number were shared by the two. For C. grandis, 26 identified DAP spots were shared by pH 2.5- and pH 3-treated leaves. Only 17 or one identified DAP spots presented in pH 2.5- or pH 3-treated leaves, respectively. In short, we identified 16 protein spots increased in abundance and 33 protein spots decreased in abundance, and two protein spots increased in abundance and eight protein spots decreased in abundance from the pH 2.5- and pH 3-treated C. sinensis leaves, respectively, and nine protein spots increased in abundance and 34 protein spots decreased in abundance, and four protein spots increased in abundance and 23 protein spots decreased in abundance from the pH 2.5- and pH 3-treated C. grandis leaves, respectively (Tables 1, 2 and 3 and Fig. 6a-d). The majority of these low pH-responsive proteins were identified only in the C. sinensis or C. grandis leaves, only six pH 2.5-responsive proteins (i.e., Cs7g31800, Cs3g01420, Cs1g25510, Cs8g19010, Cs3g11320 and orange1.1 t04488) and two pH 3-responsive proteins (i.e., Cs7g31800 and Cs1g25510) with the same accession number were simultaneously identified in the two species (Tables 2 and 3 and Fig. 6e-f). These low pH-responsive proteins were mainly associated with carbohydrate and energy metabolism, antioxidation and detoxification, stress response, protein and amino acid metabolisms, lipid metabolism, cellular transport, signal transduction and nucleic acid metabolism (Tables 2 and 3 and Fig. 6a-d).
Table 2.
DAP spots and their identification by MALDI-TOF/TOF-MS in leaves from C. sinensis seedlings submitted to pH 2.5, pH 3 or pH 6 for 9 months
| Spot No.a | Protein identity | Accession No. | Mr (kDa)/pI theor. | Mr (kDa)/pI exp. | Protein score | Peptide ions | NMPb | Ratioc | Coveredd sequence (%) | Charge | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH 2.5 | pH 3 | pH 6 | ||||||||||
| Antioxidation and detoxification | ||||||||||||
| S6 | L-ascorbate peroxidase 1, cytosolic | Cs8g17370.1 | 27.57/5.55 | 44.86/5.80 | 584 | 1 26 | 24 | 2.16 ± 0.09a | 1.07 ± 0.12b | 1.00 ± 0.03b | 22 | 1 |
| S4 | L-ascorbate peroxidase 3, peroxisomal | Cs3g19810.2 | 47.33/8.59 | 53.31/5.73 | 573 | 144 | 25 | 2.94 ± 0.80a | 1.11 ± 0.21b | 1.00 ± 0.15b | 23 | 1 |
| S5 | Probable aldo-keto reductase 1 | Cs3g10670.1 | 38.42/5.50 | 62.64/5.68 | 295 | 90 | 15 | 2.41 ± 0.26a | 0.84 ± 0.05b | 1.00 ± 0.17b | 14 | 1 |
| S8 | Isoflavone reductase-like protein | Cs2g16220.1 | 34.25/6.40 | 56.42/6.23 | 155 | 56 | 17 | 2.03 ± 0.29a | 0.80 ± 0.16b | 1.00 ± 0.11b | 15 | 1 |
| S1 | Peroxidase 15 | orange1.1 t02046.1 | 37.43/4.52 | 71.41/4.46 | 178 | 90 | 9 | 0.37 ± 0.07b | 0.45 ± 0.06b | 1.00 ± 0.04a | 8 | 1 |
| S9 | Glutathione S-transferase U19 | Cs5g15190.1 | 25.56/7.56 | 42.77/6.64 | 148 | 71 | 14 | 0.30 ± 0.05b | 0.83 ± 0.10a | 1.00 ± 0.09a | 13 | 1 |
| S10 | Thioredoxin-2 | Cs7g13660.1 | 21.44/8.44 | 21.81/4.87 | 178 | 62 | 7 | 0.47 ± 0.10b | 0.87 ± 0.11a | 1.00 ± 0.06a | 6 | 1 |
| Stress response | ||||||||||||
| S11 | Heat shock protein 90–1 | Cs5g03150.1 | 80.52/5.03 | 90.75/5.15 | 186 | 96 | 41 | 1.62 ± 0.19a | 0.53 ± 0.09c | 1.00 ± 0.02b | 37 | 1 |
| S12 | Putative uncharacterized protein Sb02g035950 | Cs1g06050.1 | 27.76/5.56 | 52.74/6.00 | 265 | 87 | 11 | 1.29 ± 0.04a | 0.51 ± 0.14b | 1.00 ± 0.10a | 10 | 1 |
| S13 | Abscisic stress ripening-like protein | Cs3g21500.1 | 20.05/5.75 | 45.68/6.08 | 411 | 92 | 14 | 0.20 ± 0.05c | 1.59 ± 0.11a | 1.00 ± 0.06b | 13 | 1 |
| S17 | Thiamine thiazole synthase 1, chloroplastic | Cs4g11090.1 | 37.60/5.40 | 53.52/5.22 | 304 | 95 | 15 | 1.64 ± 0.12a | 0.75 ± 0.02b | 1.00 ± 0.18b | 14 | 1 |
| S39 | S-norcoclaurine synthase; Pathogenesis-related (PR)-10-related norcoclaurine synthase-like protein | Cs6g03210.1 | 17.29/4.89 | 28.52/5.12 | 485 | 90 | 17 | 4.45 ± 0.27a | 2.23 ± 0.16b | 1.00 ± 0.13c | 15 | 1 |
| Carbohydrate and energy metabolism | ||||||||||||
| S14 | Chlorophyll a-b binding protein 215 | Cs1g06360.1 | 28.93/5.13 | 41.21/4.90 | 71 | 56 | 6 | 2.22 ± 0.10a | 0.46 ± 0.03c | 1.00 ± 0.16b | 5 | 1 |
| S20 | Chlorophyll a-b binding protein 4, chloroplastic | Cs3g06180.1 | 29.52/6.84 | 40.18/5.42 | 167 | 88 | 11 | 0.13 ± 0.02c | 0.71 ± 0.07b | 1.00 ± 0.11a | 10 | 1 |
| S16 | Oxygen-evolving enhancer protein 1–1, chloroplastic | Cs1g23450.1 | 35.38/5.83 | 49.12/5.09 | 371 | 110 | 14 | 2.20 ± 0.59a | 0.82 ± 0.11b | 1.00 ± 0.02b | 13 | 1 |
| S19 | Oxygen-evolving enhancer protein 1–1, chloroplastic | Cs1g23450.1 | 35.38/5.83 | 48.88/5.44 | 523 | 135 | 19 | 2.67 ± 0.76a | 0.99 ± 0.19b | 1.00 ± 0.03b | 17 | 1 |
| S18e | Ribulose bisphosphate carboxylase/oxygenase activase 1, chloroplastic | Cs7g31800.4 | 50.90/5.33 | 68.00/5.12 | 534 | 91 | 23 | 0.30 ± 0.06b | 0.44 ± 0.06b | 1.00 ± 0.09a | 21 | 1 |
| S21 | Ribulose bisphosphate carboxylase/oxygenase activase 1, chloroplastic | Cs7g31800.3 | 46.96/5.94 | 65.74/5.36 | 511 | 109 | 20 | 0.21 ± 0.02b | 0.91 ± 0.02a | 1.00 ± 0.08a | 18 | 1 |
| S22 | Rubisco subunit binding-protein β-2 subunit; Chaperonin 60 subunit β 1 | Cs9g03300.1 | 64.78/5.85 | 81.19/5.36 | 798 | 107 | 39 | 0.33 ± 0.04b | 1.10 ± 0.10a | 1.00 ± 0.13a | 35 | 1 |
| S35 | Ferredoxin-NADP reductase, leaf isozyme, chloroplastic | Cs1g25510.1 | 40.48/8.68 | 55.49/6.40 | 258 | 80 | 22 | 0.45 ± 0.04c | 0.65 ± 0.08b | 1.00 ± 0.02a | 20 | 1 |
| S37 | Ferredoxin-NADP reductase, leaf isozyme, chloroplastic | Cs1g25510.4 | 40.48/8.68 | 55.29/6.70 | 240 | 82 | 21 | 0.45 ± 0.03b | 0.91 ± 0.11a | 1.00 ± 0.13a | 19 | 1 |
| S27 | Phosphoglycerate kinase 1, chloroplastic | orange1.1 t03280.1 | 49.45/8.20 | 67.19/5.91 | 567 | 95 | 25 | 0.25 ± 0.10b | 0.87 ± 0.06a | 1.00 ± 0.11a | 23 | 1 |
| S29 | Phosphoglycerate kinase 1, chloroplastic | orange1.1 t03280.1 | 49.45/8.20 | 80.46/5.95 | 346 | 92 | 13 | 0.06 ± 0.02b | 0.95 ± 0.10a | 1.00 ± 0.14a | 12 | 1 |
| S23 | 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase, chloroplastic | Cs3g01420.1 | 32.59/8.29 | 47.43/5.72 | 415 | 121 | 14 | 0.31 ± 0.04b | 0.80 ± 0.08a | 1.00 ± 0.15a | 13 | 1 |
| S24 | Succinyl-CoA ligase [ADP-forming] subunit beta, mitochondrial | Cs5g29390.1 | 45.26/5.98 | 63.82/5.62 | 286 | 78 | 27 | 0.52 ± 0.13b | 1.10 ± 0.06a | 1.00 ± 0.06a | 25 | 1 |
| S28 | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial | Cs1g17930.1 | 59.37/8.43 | 77.39/5.80 | 390 | 94 | 23 | 0.30 ± 0.06c | 0.77 ± 0.04b | 1.00 ± 0.05a | 21 | 1 |
| S32 | Dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex 1, mitochondrial | Cs2g21190.1 | 51.06/9.07 | 72.17/6.03 | 181 | 135 | 11 | 0.19 ± 0.04b | 0.86 ± 0.04a | 1.00 ± 0.07a | 10 | 1 |
| S3 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 8-A, mitochondrial | Cs8g06410.1 | 26.11/5.68 | 42.59/5.12 | 238 | 48 | 20 | 2.58 ± 0.27a | 1.10 ± 0.10b | 1.00 ± 0.09b | 18 | 1 |
| S26 | ATP synthase gamma chain 1, chloroplastic | Cs2g03080.1 | 40.62/6.08 | 59.62/5.82 | 208 | 103 | 10 | 0.19 ± 0.04c | 0.44 ± 0.06b | 1.00 ± 0.06a | 9 | 1 |
| S30 | Probable ATP synthase 24 kDa subunit, mitochondrial | Cs1g04030.1 | 27.52/8.90 | 46.81/6.24 | 368 | 96 | 22 | 0.22 ± 0.06b | 0.55 ± 0.11b | 1.00 ± 0.14a | 20 | 1 |
| S36 | Probable ATP synthase 24 kDa subunit, mitochondrial | Cs1g04030.1 | 27.52/8.90 | 45.64/6.86 | 543 | 102 | 27 | 0.29 ± 0.07b | 0.68 ± 0.08ab | 1.00 ± 0.24a | 25 | 1 |
| S47 | DNA-damage-repair/toleration protein DRT102; ribose-5-phosphate isomerase B | Cs3g11320.1 | 33.52/5.25 | 53.47/5.39 | 393 | 85 | 19 | 0.44 ± 0.07b | 1.02 ± 0.10a | 1.00 ± 0.16a | 17 | 1 |
| Protein and amino acid metabolism | ||||||||||||
| S40 | Peptidyl-prolyl cis-trans isomerase CYP37, chloroplastic | Cs1g06710.1 | 50.39/6.42 | 60.41/5.11 | 579 | 139 | 21 | 3.06 ± 0.91a | 1.36 ± 0.22ab | 1.00 ± 0.11b | 19 | 1 |
| S43 | T-complex protein 1 subunit beta | Cs3g26890.2 | 57.19/5.56 | 79.99/5.91 | 226 | 73 | 22 | 0.30 ± 0.13b | 1.04 ± 0.05a | 1.00 ± 0.19a | 20 | 1 |
| S41 | Proteasome subunit beta type-6-A like protein | Cs7g07630.1 | 25.57/5.21 | 40.62/5.73 | 254 | 86 | 12 | 0.23 ± 0.01c | 0.68 ± 0.04b | 1.00 ± 0.10a | 11 | 1 |
| S44 | 26S proteasome non-ATPase regulatory subunit 11A | Cs4g04180.1 | 47.05/5.79 | 70.34/6.16 | 367 | 97 | 24 | 0.22 ± 0.028b | 0.83 ± 0.091a | 1.00 ± 0.06a | 22 | 1 |
| S50 | Unknown protein DS12 from 2D-PAGE of leaf, chloroplastic; ACT domain-containing protein, putative, expressed | orange1.1 t04488.1 | 30.69/5.59 | 51.57/4.75 | 333 | 105 | 15 | 0.35 ± 0.07b | 0.89 ± 0.02a | 1.00 ± 0.04a | 14 | 1 |
| S51 | Kynurenine formamidase | Cs8g05140.1 | 30.62/5.32 | 55.58/5.48 | 270 | 75 | 13 | 0.44 ± 0.01b | 1.04 ± 0.03a | 1.00 ± 0.08a | 12 | 1 |
| Cell wall and cytoskeleton | ||||||||||||
| S45 | Tubulin alpha-1 chain | Cs9g03120.1 | 49.75/4.99 | 73.13/5.41 | 487 | 85 | 20 | 0.40 ± 0.05b | 0.93 ± 0.12a | 1.00 ± 0.13a | 18 | 1 |
| S46 | Caffeic acid 3-O-methyltransferase 1 | orange1.1 t05216.1 | 29.98/5.73 | 63.37/6.19 | 194 | 66 | 11 | 0.53 ± 0.09b | 1.18 ± 0.04a | 1.00 ± 0.04a | 10 | 1 |
| Nucleic acid metabolism | ||||||||||||
| S48 | Hypoxanthine-guanine phosphoribosyltransferase | Cs3g21990.1 | 20.48/5.54 | 38.60/5.61 | 143 | 94 | 8 | 0.41 ± 0.03b | 1.07 ± 0.17a | 1.00 ± 0.08a | 7 | 1 |
| Lipid metabolism | ||||||||||||
| S2 | Plastid lipid-associated protein 2, chloroplastic; Chromoplast-specific carotenoid-associated protein, chromoplast | Cs2g02520.1 | 43.18/6.08 | 50.48/4.67 | 503 | 92 | 24 | 1.68 ± 0.09a | 0.79 ± 0.18b | 1.00 ± 0.02b | 22 | 1 |
| S7 | Epoxide hydrolase 4 | Cs2g06360.1 | 36.94/5.90 | 59.20/6.19 | 352 | 73 | 21 | 2.51 ± 0.27a | 0.90 ± 0.10b | 1.00 ± 0.18b | 19 | 1 |
| S31 | Cinnamoyl-CoA reductase 1 | Cs8g20610.1 | 35.48/5.57 | 55.34/6.06 | 220 | 77 | 14 | 0.31 ± 0.02b | 0.66 ± 0.21ab | 1.00 ± 0.12a | 13 | 1 |
| Cellular transport | ||||||||||||
| S15 | Ferritin-1, chloroplastic | Cs6g09150.2 | 28.97/5.46 | 43.81/4.92 | 88 | 64 | 8 | 1.97 ± 0.34a | 0.69 ± 0.09b | 1.00 ± 0.11b | 7 | 1 |
| Signal transduction | ||||||||||||
| S38 | 14–3-3-like protein GF14 kappa; General regulatory factor 8 | Cs1g20220.1 | 27.69/4.87 | 45.64/4.90 | 339 | 106 | 20 | 4.71 ± 0.32a | 1.20 ± 0.15b | 1.00 ± 0.06b | 18 | 1 |
| Others | ||||||||||||
| S52 | Putative uncharacterized protein Sb02g003450 | Cs5g09380.2 | 27.69/7.84 | 45.27/5.58 | 193 | 60 | 14 | 0.38 ± 0.11b | 1.00 ± 0.09a | 1.00 ± 0.12a | 13 | 1 |
| S42 | Putative uncharacterized protein Sb09g010000 | Cs8g19010.1 | 55.68/7.66 | 79.38/5.66 | 248 | 102 | 19 | 0.19 ± 0.06b | 0.73 ± 0.13a | 1.00 ± 0.22a | 17 | 1 |
| S34 | Flavoprotein WrbA | Cs4g11860.1 | 22.29/5.75 | 40.45/6.51 | 256 | 136 | 10 | 0.44 ± 0.05b | 0.67 ± 0.07ab | 1.00 ± 0.14a | 9 | 1 |
| S49 | NAD(P)H-dependent 6′-deoxychalcone synthase | orange1.1 t00001.2 | 67.34/9.06 | 57.67/6.61 | 247 | 100 | 20 | 0.46 ± 0.07b | 0.67 ± 0.11b | 1.00 ± 0.07a | 18 | 1 |
| Unidentified protein spots | ||||||||||||
| S33 | Probable phosphoglucomutase, cytoplasmic 1 | orange1.1 t05474.1 | 16.20/5.32 | 84.77/6.25 | 100 | 91 | 4 | 0.40 ± 0.05b | 0.77 ± 0.08ab | 1.00 ± 0.17a | 4 | 1 |
| S25 | Glucose-6-phosphate 1-dehydrogenase 1, chloroplastic | Cs7g11110.1 | 66.95/7.68 | 25.37/5.76 | 65 | 23 | 21 | 0.46 ± 0.04b | 1.34 ± 0.08a | 1.00 ± 0.23b | 19 | 1 |
| S53 | Endo-1,3;1,4-beta-D-glucanase | Cs9g05910.3 | 28.00/7.07 | 43.84/5.09 | 43 | / | 12 | 3.32 ± 0.29a | 1.02 ± 0.07b | 1.00 ± 0.09b | 11 | 1 |
| S54 | E3 ubiquitin-protein ligase MARCH9 | Cs9g05100.7 | 32.53/8.56 | 16.93/6.1 | 44 | / | 12 | 4.26 ± 0.19a | 0.45 ± 0.02b | 1.00 ± 0.18b | 11 | 1 |
| S55 | Cytochrome b6-f complex iron-sulfur subunit, chloroplastic | Cs2g22650.3 | 22.32/8.76 | 30.95/8.76 | 52 | 36 | 6 | 0.33 ± 0.04c | 0.75 ± 0.05b | 1.00 ± 0.09a | 5 | 1 |
aSpot number corresponds to the 2-DE imagines in Fig. 5
bNMP means the number of matched peptides
cRatio means the ratio of pH 2.5 and pH 3.0 to pH 6.0; Different letters within a row indicate significant differences at P < 0.05
dCovered sequence (%) means the ratio of the number of amino of the matched peptides to the number of amino acids of the full-length protein
eLow pH-responsive proteins shared by the two Citrus species were highlighted in bold
Table 3.
DAP spots and their identification by MALDI-TOF/TOF-MS in leaves from C. grandis seedlings submitted to pH 2.5, pH 3 or pH 6 for 9 months
| Spot No.a | Protein identity | Accession No. | Mr (kDa)/pI theor. | Mr (kDa)/pI exp. | Protein score | Peptide ions | NMPb | Ratioc | Coveredd sequence (%) | Charge | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH 2.5 | pH 3 | pH 6 | ||||||||||
| Antioxidation and detoxification | ||||||||||||
| G2 | Copper/zinc superoxide dismutase 2, chloroplastic | Cs8g15520.1 | 26.00/6.52 | 22.72/6.05 | 341 | 119 | 8 | 2.51 ± 0.06a | 2.42 ± 0.24a | 1.00 ± 0.17b | 7 | 1 |
| G11 | Aldehyde dehydrogenase family 2 member B4, mitochondrial | Cs5g05240.1 | 58.91/7.11 | 75.4/5.31 | 409 | 115 | 27 | 2.48 ± 0.20a | 0.33 ± 0.06c | 1.00 ± 0.08b | 25 | 1 |
| G40 | Apolipoprotein D | Cs4g01600.1 | 21.52/6.33 | 32.64/6.84 | 264 | 106 | 11 | 1.57 ± 0.22a | 0.62 ± 0.04b | 1.00 ± 0.10b | 10 | 1 |
| G15 | Phosphomannomutase 1 | orange1.1 t00331.1 | 28.16/6.19 | 59.09/5.85 | 561 | 122 | 25 | 0.48 ± 0.05b | 1.01 ± 0.02a | 1.00 ± 0.05a | 23 | 1 |
| G1 | NADP-dependent alkenal double bond reductase P2 | Cs5g21010.1 | 36.04/6.16 | 82.02/5.3 | 435 | 116 | 18 | 0.18 ± 0.04b | 0.38 ± 0.07b | 1.00 ± 0.29a | 16 | 1 |
| G3 | Thioredoxin-like protein CDSP32, chloroplastic | Cs3g26690.1 | 39.02/8.59 | 49.64/6.62 | 472 | 106 | 18 | 0.36 ± 0.05c | 0.66 ± 0.09b | 1.00 ± 0.08a | 16 | 1 |
| G4 | Thioredoxin-like protein CDSP32, chloroplastic | Cs3g26690.1 | 39.02/8.59 | 49.46/6.85 | 477 | 106 | 20 | 0.22 ± 0.01b | 0.83 ± 0.15a | 1.00 ± 0.22a | 18 | 1 |
| G8 | Ferredoxin-thioredoxin reductase catalytic chain, chloroplastic | Cs6g20130.1 | 16.57/6.29 | 24.68/5.07 | 130 | 61 | 7 | 0.25 ± 0.05b | 0.42 ± 0.10b | 1.00 ± 0.08a | 6 | 1 |
| G23 | Protein disulfide isomerase-like 1–1 | Cs3g19790.1 | 55.62/4.84 | 83.51/4.91 | 625 | 95 | 31 | 0.43 ± 0.06b | 0.80 ± 0.09ab | 1.00 ± 0.16a | 28 | 1 |
| G39 | Annexin A13 | Cs3g18360.1 | 36.10/5.55 | 28.12/6.6 | 422 | 82 | 23 | 0.18 ± 0.05c | 0.52 ± 0.04b | 1.00 ± 0.07a | 21 | 1 |
| G36 | Nucleoside diphosphate kinase II, chloroplastic | Cs5g06840.1 | 25.53/9.35 | 25.98/6.32 | 467 | 106 | 13 | 0.07 ± 0.02b | 0.25 ± 0.02b | 1.00 ± 0.15a | 12 | 1 |
| G5 | Betaine aldehyde dehydrogenase 1, chloroplastic | Cs5g04880.1 | 54.95/5.40 | 24.08/4.24 | 236 | 116 | 12 | 0.10 ± 0.05c | 0.52 ± 0.10b | 1.00 ± 0.01a | 11 | 1 |
| Carbohydrate and energy metabolism | ||||||||||||
| G7 | RuBisCO large subunit-binding protein subunit alpha, chloroplastic; Chaperonin 60 subunit alpha 1 | Cs8g16040.1 | 61.50/5.23 | 46.22/5.1 | 215 | 77 | 23 | 2.75 ± 0.28a | 1.58 ± 0.11b | 1.00 ± 0.06b | 21 | 1 |
| G10 | RuBisCO large subunit-binding protein subunit alpha, chloroplastic; Chaperonin 60 subunit α 1 | Cs8g16040.1 | 61.50/5.23 | 82.35/5.02 | 783 | 126 | 37 | 0.35 ± 0.09b | 0.99 ± 0.13a | 1.00 ± 0.09a | 34 | 1 |
| G12e | Ribulose bisphosphate carboxylase/oxygenase activase 1, chloroplastic | Cs7g31800.2 | 41.38/5.07 | 64.04/5.4 | 219 | 97 | 10 | 0.14 ± 0.02b | 0.31 ± 0.05b | 1.00 ± 0.08a | 9 | 1 |
| G19 | Ferredoxin--NADP reductase, leaf isozyme, chloroplastic | Cs1g25510.1 | 40.48/8.68 | 54.2/6.92 | 289 | 70 | 23 | 0.44 ± 0.05b | 0.53 ± 0.06b | 1.00 ± 0.12a | 21 | 1 |
| G13 | Rhodanese-like domain-containing protein 4A, chloroplastic | orange1.1 t00475.2 | 46.75/5.59 | 69.08/5.26 | 507 | 130 | 24 | 0.41 ± 0.06b | 1.00 ± 0.12a | 1.00 ± 0.18a | 22 | 1 |
| G16 | 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase, chloroplastic | Cs3g01420.1 | 32.59/8.29 | 46.72/5.75 | 321 | 95 | 12 | 0.34 ± 0.06b | 0.88 ± 0.13a | 1.00 ± 0.15a | 11 | 1 |
| G17 | Probable 6-phosphogluconolactonase 2 | orange1.1 t02542.1 | 35.38/6.24 | 81.27/5.72 | 454 | 133 | 15 | 0.34 ± 0.04b | 2.61 ± 0.45a | 1.00 ± 0.01b | 14 | 1 |
| G18 | Dihydroxyacetone kinase 2 | orange1.1 t02644.1 | 61.75/5.74 | 81.59/6.13 | 342 | 95 | 19 | 2.30 ± 0.56a | 1.66 ± 0.14ab | 1.00 ± 0.04b | 17 | 1 |
| G9 | Probable fructokinase-1 | Cs5g22920.1 | 35.11/4.98 | 59.41/5.02 | 718 | 137 | 22 | 0.49 ± 0.10b | 0.58 ± 0.05b | 1.00 ± 0.07a | 20 | 1 |
| G33 | DNA-damage-repair/toleration protein DRT102; ribose-5-phosphate isomerase B | Cs3g11320.1 | 33.52/5.25 | 52.44/5.28 | 279 | 104 | 15 | 0.27 ± 0.06b | 0.79 ± 0.05a | 1.00 ± 0.20a | 14 | 1 |
| Protein and amino acid metabolism | ||||||||||||
| G26 | Peptidyl-prolyl cis-trans isomerase FKBP62 | Cs7g16620.3 | 63.83/5.19 | 87.75/5.46 | 344 | 81 | 21 | 0.23 ± 0.02c | 0.79 ± 0.07b | 1.00 ± 0.03a | 19 | 1 |
| G21 | Peptidyl-prolyl cis-trans isomerase CYP38, chloroplastic | Cs2g28260.1 | 48.02/5.00 | 64.55/4.61 | 231 | 80 | 18 | 0.20 ± 0.06c | 0.47 ± 0.04b | 1.00 ± 0.05a | 16 | 1 |
| G30 | Ankyrin repeat domain-containing protein 2 | orange1.1 t02270.1 | 64.18/5.29 | 64.12/4.57 | 261 | 110 | 16 | 0.16 ± 0.04b | 0.72 ± 0.09a | 1.00 ± 0.11a | 15 | 1 |
| G31 | Ankyrin repeat domain-containing protein 2 | orange1.1 t02270.1 | 64.18/5.29 | 65.14/4.51 | 228 | 120 | 13 | 0.30 ± 0.13b | 1.13 ± 0.16a | 1.00 ± 0.21a | 12 | 1 |
| G20 | SKP1-like protein 10 | Cs3g26510.1 | 17.82/4.56 | 33.74/4.58 | 105 | 60 | 11 | 0.16 ± 0.02b | 0.27 ± 0.04b | 1.00 ± 0.13a | 10 | 1 |
| G29 | Subtilisin-like protease SDD1 | Cs1g17350.1 | 82.90/5.86 | 49.48/6.95 | 311 | 107 | 12 | 0.25 ± 0.06b | 0.07 ± 0.01b | 1.00 ± 0.09a | 11 | 1 |
| G24 | 26S protease regulatory subunit 6B homolog | Cs1g08770.1 | 46.49/5.42 | 72.36/5.68 | 425 | 87 | 27 | 1.24 ± 0.12a | 0.42 ± 0.05b | 1.00 ± 0.12a | 25 | 1 |
| G6 | Proline iminopeptidase | Cs8g03250.1 | 44.67/5.70 | 60.15/5.58 | 463 | 116 | 23 | 3.44 ± 0.42a | 2.21 ± 0.41b | 1.00 ± 0.05c | 21 | 1 |
| G25 | Glutamine synthetase cytosolic isozyme | Cs9g05680.1 | 47.86/6.29 | 65.88/5.51 | 181 | 82 | 16 | 0.07 ± 0.02c | 0.61 ± 0.12b | 1.00 ± 0.14a | 15 | 1 |
| G34 | Dihydropyrimidine dehydrogenase [NADP(+)] | orange1.1 t02649.1 | 46.01/6.29 | 66.1/5.75 | 189 | 96 | 16 | 0.28 ± 0.04b | 0.50 ± 0.02b | 1 ± 0.17a | 15 | 1 |
| G27 | Argininosuccinate synthase, chloroplastic | Cs5g07120.1 | 54.38/6.50 | 70.63/5.94 | 371 | 112 | 26 | 2.36 ± 0.13a | 0.90 ± 0.14b | 1.00 ± 0.18b | 24 | 1 |
| G28 | 3-isopropylmalate dehydratase large subunit 1 | Cs5g35290.1 | 55.66/6.68 | 76.63/6.29 | 124 | 72 | 15 | 3.06 ± 0.05a | 1.55 ± 0.12b | 1.00 ± 0.08c | 14 | 1 |
| G22 | Unknown protein DS12 from 2D-PAGE of leaf, chloroplastic; ACT domain-containing protein, putative, expressed | orange1.1 t04488.3 | 26.98/4.94 | 51.32/4.76 | 318 | 89 | 16 | 0.41 ± 0.11b | 1.08 ± 0.26a | 1.00 ± 0.02a | 15 | 1 |
| Signal transduction | ||||||||||||
| G32 | Plasma membrane-associated cation-binding protein 1 | Cs2g12010.1 | 23.34/5.03 | 51.44/5.25 | 113 | 37 | 12 | 0.19 ± 0.04b | 0.39 ± 0.04b | 1.00 ± 0.09a | 11 | 1 |
| Nucleic acid metabolism | ||||||||||||
| G35 | Adenylosuccinate synthetase 1, chloroplastic | orange1.1 t03395.1 | 53.23/6.40 | 70.64/6.01 | 568 | 101 | 25 | 0.25 ± 0.01b | 0.42 ± 0.09b | 1 ± 0.22a | 23 | 1 |
| Lipid metabolism | ||||||||||||
| G37 | Lipase/lipooxygenase, PLAT/LH2 family protein | Cs1g01370.1 | 19.62/4.80 | 22.28/4.22 | 213 | 134 | 7 | 0.15 ± 0.03c | 0.46 ± 0.07b | 1 ± 0.03a | 6 | 1 |
| G38 | Lipase/lipooxygenase, PLAT/LH2 family protein | Cs1g01370.1 | 19.62/4.80 | 44.79/6.6 | 222 | 137 | 7 | 0.33 ± 0.07b | 0.48 ± 0.06b | 1 ± 0.003a | 6 | 1 |
| G14 | Allene oxide cyclase 1, chloroplastic | Cs6g18900.1 | 27.51/8.63 | 34.12/5.73 | 371 | 83 | 13 | 0.19 ± 0.06c | 0.64 ± 0.03b | 1.00 ± 0.11a | 12 | 1 |
| Cellular transport | ||||||||||||
| G41 | V-type proton ATPase catalytic subunit A isoform 1 | Cs1g10270.1 | 68.68/5.29 | 59.4/6.96 | 635 | 100 | 40 | 1.50 ± 0.16a | 0.65 ± 0.07b | 1.00 ± 0.08b | 36 | 1 |
| Others | ||||||||||||
| G42 | Putative uncharacterized protein Sb01g045410 | Cs4g06170.1 | 26.81/8.90 | 31.73/5.39 | 228 | 82 | 11 | 0.13 ± 0.04b | 0.22 ± 0.03b | 1.00 ± 0.14a | 10 | 1 |
| G43 | Putative uncharacterized protein Sb09g010000 | Cs8g19010.1 | 55.68/7.66 | 80.33/6.05 | 285 | 118 | 20 | 0.46 ± 0.05c | 1.18 ± 0.06a | 1.00 ± 0.02b | 18 | 1 |
| G44 | Uncharacterized protein At2g37660, chloroplastic | Cs6g06590.1 | 36.01/9.43 | 45/6.92 | 485 | 136 | 14 | 0.19 ± 0.03c | 0.58 ± 0.07b | 1.00 ± 0.15a | 13 | 1 |
| Unidentified protein spots | ||||||||||||
| G45 | Kinesin-like protein KIF21A | Cs3g17220.3 | 108.17/6.47 | 25.06/4.49 | 52 | / | 28 | 0.27 ± 0.07b | 0.39 ± 0.07b | 1.00 ± 0.25a | 25 | 1 |
| G46 | Actinidain | Cs3g23180.1 | 51.42/5.37 | 52.44/4.62 | 55 | 55 | 2 | 0.35 ± 0.01b | 0.97 ± 0.25a | 1.00 ± 0.13a | 2 | 1 |
| G47 | Peptide methionine sulfoxide reductase B2, chloroplastic | Cs8g03090.1 | 17.59/10.55 | 20.63/5.27 | 51 | 24 | 8 | 0.10 ± 0.009b | 0.34 ± 0.14b | 1.00 ± 0.13a | 7 | 1 |
| G48 | Putative uncharacterized protein | Cs7g03150.1 | 15.56/6.22 | 85.49/5.56 | 55 | 29 | 7 | 0.35 ± 0.06b | 0.85 ± 0.05a | 1.00 ± 0.13a | 6 | 1 |
aSpot number corresponds to the 2-DE imagines
bNMP means the number of matched peptides in Fig. 5
cRatio means the ratio of pH 2.5 and pH 3 to pH 6.0; Different letters within a row indicate significant differences at P < 0.05
dCovered sequence (%) means the ratio of the number of amino of the matched peptides to the number of amino acids of the full-length protein
eLow pH-responsive proteins shared by the two Citrus species were highlighted in bold
Fig. 6.

Classification of low pH-responsive proteins (a-d) and venn diagram analysis of low pH-responsive proteins (e-f). For e, 37 or 43 identified DAPs only presented in the pH 2.5-treated C. grandis or C. sinensis leaves, respectively, and only six identified DAPs with the same accession number were shared by the two. For f, 25 or 9 identified DAPs only presented in the pH 3-treated C. grandis or C. sinensis leaves, respectively, and only two identified DAPs with the same accession number were shared by the two
qRT-PCR analysis of genes for some low pH-responsive proteins
To understand the correlation between gene expression levels and 2-DE data, we used qRT-PCR to assay the transcript levels of genes for a total of 26 DAPs from the C. sinensis (i.e., S1, S4, S5, S6, S7, S9, S10, S15, S23, S26, S27, S35 and S40) and C. grandis (i.e., G2, G5, G6, G13, G14, G15, G16, G19, G21, G36, G37, G40 and G41) leaves. Actin and PRPF31 were selected as the internal standards (Fig. 7). The transcript levels of all these genes with the exceptions of G5, G14, G19, G37, S23, S35 and S40 matched well with our 2-DE data, regardless of which gene was used as the internal standard (Tables 2 and 3). In addition, there was a positive linear correlation between qRT-PCR results and 2-DE data, regardless of actin or PRPF31 was used as the internal standard (Fig. 7e-f). Thus, it is reasonable to assume that these DAPs were mainly regulated in the transcriptional level.
Fig. 7.

qRT-PCR analysis of 26 low-pH responsive protein genes. Relative expression levels of genes encoding 26 low-pH responsive proteins identified in C. grandis (a-b) and C. sinensis (c-d) leaves using actin (a, c) and PRPF31 (b, d) as internal standards, and the correlation analysis of qRT-PCR results and 2-DE data (e-f). For a-d, bars represent means ± SE (n = 3). For the same genes, different letters above the bars indicate a significant difference at P < 0.05. For e and f, 2-DE data from Tables 2 and 3
Discussion
Citrus sinensis and C. grandis were tolerant to low pH
Our results clearly showed that only pH 2.5 led to significant decreases in leaf CO2 assimilation and stomatal conductance (Fig. 1a-b), ASC + DHA and ASC concentrations, and ASC/(ASC + DHA) ratio (Fig. 3a, b and d), and significant increases in leaf levels of nonstructural carbohydrates (Fig. 2). Based on these results, we concluded that C. sinensis and C. grandis seedlings were tolerant to low pH. Similar results have been obtained in grafted Citrus unshiu plants [27] and C. sinensis and C. grandis seedlings (seedlings should be normal type) [8]. Thus, Citrus are ideal materials for studying low pH-tolerance of higher plants.
We found that the pH 2.5-induced decreases of both ASC + DHA and ASC concentrations and ASC/(ASC + DHA) ratio (Fig. 3a, b and d), and increases of MDA (Fig. 4), sucrose, starch and TNC concentrations (Fig. 2c, e and f) were greater in the C. grandis leaves than those in the C. sinensis leaves. This agrees with our previous finding that C. sinensis seedlings were slightly tolerant to low pH than C. grandis ones [8]. We identified slightly more pH 2.5-responsive proteins in the C. sinensis leaves (49) than those in the C. grandis leaves (43), but much more pH 3-responsive proteins in the C. grandis leaves (27) than those in the C. sinensis leaves (11) (Tables 1, 2 and 3 and Fig. 6). The observed fewer pH 3-responsive proteins in the C. sinensis leaves could be explained by the slightly higher low pH-tolerance.
As shown in Tables 1, 2 and 3 and Fig. 5, we identified more pH 2.5-responsive proteins than pH 3-responsive proteins in the C. sinensis and C. grandis leaves. This agrees with our results that only pH 2.5 significantly affected leaf gas exchange, ratio of ASC/(ASC + DHA), and levels of ASC + DHA, ASC, nonstructural carbohydrates and MDA (Figs. 1, 2, 3 and 4), and the previous report that most of the physiological and biochemical indexes were altered only at pH 2.5, but almost unaltered at pH 3 or more [8]. Here, we focused mainly on the effects of pH 2.5 on Citrus leaf protein profiles in this paper.
Low pH-responsive proteins related to carbohydrate and energy metabolism
As shown in Figs. 1a and 2, pH 2.5 significantly inhibited leaf photosynthesis and increased leaf accumulation of nonstructural carbohydrates. Thus, the abundances of proteins involved in carbohydrate and energy metabolism might be altered at pH 2.5. As expected, we identified four DAPs increased in abundance and 16 DAPs decreased in abundance, and two DAPs increased in abundance and eight DAPs decreased in abundance in the pH 2.5-treated C. sinensis and C. grandis leaves, respectively. Also, we obtained five DAPs increased in abundance from the pH 3-treated C. sinensis leaves, and one DAP increased in abundance and three DAPs decreased in abundance from the C. grandis leaves (Tables 2 and 3 and Fig. 6a-d). Evidently, low pH-responsive proteins related to carbohydrate and energy metabolism greatly differed between the two Citrus species.
In higher plants, Chl a/b-binding protein (CAB) binds to Chl and forms light harvesting complex (LHC), which functions as a light receptor. Lhca proteins are associated with the photosystem I (PSI) light-harvesting complexes (LHCI) and the Lhcb proteins are associated with the LHCII. Damkjær et al. observed that the maximum photosystem II (PSII) efficiency of dark-adapted leaves (Fv/Fm) dropped more in the A. thaliana T-DNA knockout plants lacking Lhcb3 (koLhcb3) than that in the wild type, indicating the involvement of Lhcb3 in photoacclimation [42]. We found that the abundance of Chl a-b binding protein 215 (Lhcb3, S14) was increased in the pH 2.5-treated C. sinensis leaves, but not in the pH 2.5-treated C. grandis leaves (Tables 2 and 3). This agrees with our report that the decreases of both Fv/Fm and the electron transport rate (ETR) through PSII in response to pH 2.5 was slightly lower in the C. sinensis leaves than that in the C. grandis leaves [8]. Similarly, the abundances of oxygen-evolving enhancer protein 1–1 (PSBO2, S16 and 19) were enhanced only in the pH 2.5-treated C. sinensis leaves (Tables 2 and 3). Previous studies showed that PSBO2 was necessary for the stability of Mn cluster, the primary site of water splitting [43], and played a role in D1 dephosphorylation and turnover [44]. The increased abundances of PSBO2 might contribute to the stability of oxygen evolving complexes (OEC), as indicated by the less pronounced ΔK-band (a specific indicator of OEC) [45] in the pH 2.5-treated C. sinensis leaves than that in the pH 2.5-treated C. grandis leaves [8]. However, the abundance of Chl a-b binding protein 4 (S20, Lhca3) was decreased in the pH 2.5-treated C. sinensis leaves (Table 2).
The abundances of Rubisco activase 1 (S18 and S21) catalyzed the activation of Rubisco and of Rubisco subunit binding-protein β-2 subunit (60 kDa chaperonin 1, S22) involved in protein folding and stabilization were decreased in the pH 2.5-treated C. sinensis leaves (Table 2). Transgenic plants showed that the activation of Rubisco by Rubisco activase is necessary for CO2 assimilation at atmospheric CO2 concentrations [46–48]. Suzuki et al. observed that moderate decrease of plastid chaperonin Cpn60 level led to impaired plastid division and reduced Chl level, suggesting that plastid chaperonins Cpn60α and Cpn60β were necessary for plastid division in A. thaliana [49]. The observed decreases in the abundances of Rubisco activase 1 (S18 and 21) and chaperonin 60 subunit beta 1 (S21) agrees with our report that pH 2.5 decreased Rubisco activity, photosynthesis, and Chl a and Chl b levels in the C. sinensis leaves [8]. Similarly, Rubisco activase 1 (G12) and chaperonin 60 subunit α 1 (G10) abundances (Table 3), Rubisco activity, photosynthesis, and Chl a and Chl levels [8] were decreased in the pH 2.5-treated C. grandis leaves.
The abundances of ferredoxin-NADP reductase, leaf isozyme, chloroplastic (FNR2; S35 and S37) in the C. sinensis leaves (Table 2), and the abundances of FNR2 (G19) and Rhodanese-like domain-containing protein 4A, chloroplastic (TROL; G13) in the C. grandis leaves (Table 3) were decreased at pH 2.5. FNR mediates the final step of line electron flow by transferring electron from reduced ferredoxin and NADP+, providing NADPH for a number of reactions, including carbon fixation, Chl biosynthesis and stromal redox regulation. In A. thaliana, FNR exists as two isoforms: AtLFNR1 and AtLFNR2. The Arabidopsis fnr2 RNAi mutants had lower levels of Chl and photosynthetic thylakoid proteins, decreased rate of carbon fixation than the wild type (WT) plants [50]. TROL is necessary for the maintenance of efficient linear electron flow via mediating the binding of FNR to the thylakoids. The TROL-deficient Arabidopsis plants had decreased ETR at high-light intensities accompanied with increased non-photochemical quenching (NPQ) [51]. The decreased abundance of TROL in the pH 2.5-treated C. grandis leaves agrees with the report that pH 2.5 led to decreased ETR, and increased NPQ in the C. grandis leaves [8]. In addition, the abundances of phosphoglycerate kinase 1 (PGK1) (S27 and S29) involved in Calvin cycle and 2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase (MECT) (S23 and G16) involved in chloroplast development [52] were decreased in the pH 2.5-treated C. sinensis and C. grandis leaves (Tables 2 and 3). Based on these results, we concluded that pH 2.5 decreased the abundances of proteins related to Rubisco activation, Calvin cycle, carbon fixation, chloroplast development, Chl biosynthesis and electron transport, thus lowering Chl level, ETR and photosynthesis. Here, we first found that pH 2.5 increased the accumulation of nonstructural carbohydrates in the C. grandis and C. sinensis leaves despite decreased CO2 assimilation due to the prevented sink growth caused by blocked export with the exception that pH did not significantly alter sucrose level in the C. sinensis leaves (Figs. 1a and 2). High levels of soluble sugars, particularly hexoses, can inhibit the expression of photosynthetic genes, especially of the nuclear-encoded small subunit of Rubisco, thus lowering Rubisco level and photosynthesis [53]. The pH 2.5-induced increases of glucose and fructose levels were greater in the C. sinensis leaves than those in the C. grandis leaves, and sucrose level only increased in the pH 2.5-treated C. grandis leaves (Fig. 2a-c), while both CO2 assimilation and Rubisco activity were similar between the pH 2.5-treated C. grandis and C. sinensis leaves [8]. Thus, the pH 2.5-induced decreases of leaf Rubisco activity and CO2 assimilation could not explained alone by the pH 2.5-induced accumulation of soluble sugars. Interestingly, the pH 2.5-induced accumulation of starch was higher in the C. grandis leaves than that in the C. sinensis leaves (Fig. 2e). Excessive accumulation of starch can damage chloroplastic structure, thus resulting in lower CO2 assimilation and Chl level [54]. This agrees with the report that the pH 2.5-induced decreases of Chl a and Chl b levels were greater in the C. grandis leaves than those in the C. sinensis leaves, and that mottled bleached leaves occurred only in some pH 2.5-treated C. grandis leaves [8].
We found that the abundances of proteins related to tricarboxylic acid cycle (S24 and S32), glycolysis (S28) and ATP biosynthesis (S26, S30 and S36) were decreased in the pH 2.5-treated C. sinensis leaves (Table 2), suggesting that ATP production might be reduced in these leaves, thus resulting in a decrease in energy (ATP) level. In addition to producing ATP from ADP via utilizing proton gradient formed by photosynthetic electron transport, chloroplastic ATP synthase can catalyze ATP hydrolysis, when the transmembrane electrochemical potential gradient is small [55]. Thus, the observed lower abundance of chloroplastic ATP synthase might be of advantage to the maintenance of ATP homeostasis.
Low pH-responsive proteins related to antioxidation and detoxification
Under high light, the amount of excess absorbed light energy was greater in the low pH-treated Citrus leaves, because these leaves used only less of the absorbed light energy in electron transport due to decreased ETR and CO2 assimilation [8]. Excess absorbed light energy can potentially trigger ROS generation. Indeed, the production of ROS (H2O2) was elevated in the low pH-treated C. sinensis and C. grandis leaves, especially in the latter [8]. To scavenge the increased production of ROS, the abundances of some proteins related to the scavenging of ROS might be increased in these leaves. Here, the abundances of four protein spots [i..e., L-ascorbate peroxidase (APX) 1, cytosolic (S6), L-ascorbate peroxidase 3, peroxisomal (S4), probable aldo-keto reductase 1 (AKR1, S5) and isoflavone reductase-like protein (IRL, S8), and of three protein spots [i.e., copper/zinc superoxide dismutase (Cu/Zn SOD) 2, chloroplastic (G2), aldehyde dehydrogenase family 2 member B4, mitochondrial (G11) and apolipoprotein D (G40)] involved in antioxidation and detoxification were increased in the pH 2.5-treated C. sinensis and C. grandis leaves, respectively (Tables 2 and 3). However, MDA concentration (Fig. 4) and electrolyte leakage [8] were elevated in the pH 2.5-treated C. grandis and C. sinensis leaves, especially in the former. Obviously, the antioxidant and detoxification system as a whole did not provide considerable protection to the pH 2.5-treated C. grandis and C. sinensis leaves against oxidative damage, which was greater in the former. This is also supported by our data that the pH 2.5-induced decrease of ASC/(ASC + DHA) ratio was greater in the C. grandis leaves than that in the C. sinensis leaves (Fig. 3d), because the ratio of ASC/(ASC + DHA) decreases in higher plants when exposed to oxidative stress [56–58]. The greater oxidative damage in the C. grandis leaves than that in the C. sinensis leaves might be related to the findings that the pH 2.5-induced production of H2O2 was greater in the C. grandis leaves than that in the C. sinensis leaves [8], that the abundances of more protein species involved in antioxidation and detoxification were decreased by pH 2.5 in the C. grandis (G15, G1, G3, G4, G8, G23, G39, G36 and G5) leaves than those in the C. sinensis leaves (S1, S9 and S10) (Tables 2 and 3), and that ASC level was lower in the pH 2.5-treated C. grandis leaves than that in the pH 2.5-treated C. sinensis leaves (Fig. 4b). As shown in Table 3, the abundance of phosphomannomutase (PMM) 1, which plays a crucial role in ASC biosynthesis in plants, was decreased in the pH 2.5-treated C. grandis leaves. Studies showed that ASC level in Nicotiana benthamiana and Arabidopsis leaves were decreased or increased by suppressing or overexpressing PMM, respectively [59, 60]. Thus, ASC biosynthesis might be impaired in the pH 2.5-treated C. grandis leaves. This is supported by our data that DHA + ASC and ASC levels were substantially decreased in the pH 2.5-treated C. grandis and C. sinensis leaves, especially in the former (Fig. 3a, b).
Low pH-responsive proteins related to protein and amino acid metabolism
As shown in Tables 2 and 3, we identified one DAP increased in abundance (S40) and one DAP decreased in abundance (S43) involved in protein folding and stability, and two DAPs decreased in abundance (S41and S44) involved in proteolytic degradation in the pH 2.5-treated C. sinensis leaves; and five DAPs decreased in abundance involved in protein folding and stability (G23, G26 and G21) and mainly in mediating protein-protein interactions (G30 and G31), and two DAPs decreased in abundance (G20 and G29) and two DAPs increased in abundance (G24 and G6) involved in proteolytic degradation in the pH 2.5-treated C. grandis leaves. Evidently, pH 2.5 affected protein metabolism more in the C. grandis leaves than that in the C. sinensis leaves. This is also support by our report that the pH 2.5-induced decrease of total soluble protein level was greater in the C. grandis leaves than that in the C. sinensis leaves [8]. Similarly, amino acid metabolism was more affected by pH 2.5 in the C. grandis leaves than that in the C. sinensis leaves, as indicated by more pH 2.5-responsive proteins isolated from the C. grandis leaves (G25, G27, G28, G34 and G22) than those from the C. sinensis leaves (S50 and S51; Tables 2 and 3 and Fig. 6a-b).
Low pH-responsive proteins related to cellular transport
Chloroplastic ferritin-1 (FER1) plays an important role in Fe homeostasis because of its ability to store large amounts of free Fe in a non-toxic form. FER1 and FER2, two nuclear genes of Clamydomonas reinhardtii were upregulated when its cells were shifted to Fe-deficient conditions [61]. The increased abundance of FER1 (S15) in the pH 2.5-treated C. sinensis leaves (Table 2) might contribute to the Fe homeostasis. This is also supported by our result that pH 2.5 decreased Fe level in the C. grandis leaves, but not in the C. sinensis leaves [8]. The increased abundance of FER1 in the pH 2.5-treated C. sinensis leaves also agrees with the reports that AtFER1 was induced in P-deficient Arabidopsis roots and leaves [62], because P level was decreased in the pH 2.5-treated C. sinensis leaves [8]. We found that the abundance of V-type proton ATPase (V-ATPase) catalytic subunit A isoform 1 (G41) was increased in the pH 2.5-treated C. grandis leaves (Table 2) accompanied by greatly decreased N, P, Ca and Mg levels and slightly decreased K level [8], as found in the P-deficient C. grandis and C. sinensis roots [63]. Transport across the tonoplast is energized by two proton pumps, the V-ATPase and the vacuolar H+-pyrophosphatase. Evidence shows that V-ATPase is a key regulator of intracellular ion homeostasis [64–66]. Therefore, the pH 2.5-induced increases of FER1 and V-ATPase abundances might contribute to the tolerance of Citrus plants to H+-toxicity.
Low pH-responsive proteins related to signal transduction and jasmonic acid biosynthesis
As shown in Tables 2 and 3, the abundances of 14–3-3-like protein GF14 kappa (GRF8; S38) and plasma membrane-associated cation-binding protein 1 PCAP1 (G32) involved in signal transduction were increased and decreased in the pH 2.5-treated C. sinensis and C. grandis leaves. Similarly, we found one DAP increased in abundance and three DAPs decreased in abundance involved in jasmonic acid (JA) biosynthesis in the pH 2.5-treated C. sinensis (S2) and C. grandis (G37, G38 and G14) leaves, respectively (Tables 2 and 3). Thus, the pH 2.5-induced alterations of signal transduction and JA biosynthesis might differ between C. sinensis and C. grandis leaves.
Conclusions
Our results demonstrated that C. sinensis and C. grandis were tolerant to low pH, with a slightly higher low pH-tolerance in the former. We first used 2-DE to investigate low pH-responsive proteins in Citrus leaves and identified 49 and 44 DAP spots in the pH 2.5- and/or pH 3-treated C. sinensis and C. grandis leaves, respectively. These DAPs are mainly involved in carbohydrate and energy metabolism, antioxidation and detoxification, stress response, protein and amino acid metabolisms, lipid metabolism, cellular transport, signal transduction and nucleic acid metabolism. Further analysis showed that pH 2.5 decreased the abundances of proteins related to Rubisco activation, Calvin cycle, carbon fixation, chloroplast development, Chl biosynthesis and electron transport, hence lowering Chl level, ETR and photosynthesis. The higher oxidative damage in the pH 2.5-treated C. grandis leaves might be due to a combination of factors including higher production of ROS, more proteins decreased in abundance involved in antioxidation and detoxification, and lower level of ASC. Protein and amino acid metabolisms were less affected in the pH 2.5-treated C. sinensis leaves than those in the pH 2.5-treated C. grandis leaves. The abundances of proteins related to JA biosynthesis and signal transduction were increased and decreased in pH 2.5-treated C. sinensis and C. grandis leaves, respectively. However, the abundances of cellular transport-related proteins: FER1 in the C. sinensis leaves and of V-ATPase in the C. grandis leaves, were enhanced at pH 2.5. Our investigation of low pH-responsive proteins and related physiological responses in Citrus leaves will increase our understanding of the mechanisms on low pH-toxicity and -tolerance in higher plants.
Additional files
Table S1. Specific primer pairs used for qRT-PCR analysis. (DOC 57 kb)
Figure S1. Two-DE images of proteins extracted from pH 2.5- (a, d, g, j), pH 3.0- (b, e, h, k) and pH 6.0-treated (c, f, i, l) C. grandis (a-f) and C. sinensis (g-l) leaves for the other two replicates. (PDF 317 kb)
Table S2. Master list of proteins identified in MALDI TOF/TOF MS from pH 2.5 and/or pH 3-treated C. sinensis leaves using 2DE and DIGE experiments. (DOC 1635 kb)
Table S3. Master list of proteins identified in MALDI TOF/TOF MS from pH 2.5 and/or pH 3-treated C. grandis leaves using 2DE and DIGE experiments. (DOC 1467 kb)
Figure S2. Close-up views of 22 DAP spots in pH 2.5, pH 3 and pH 6-treated C. grandis and C. sinensis leaves. (PDF 132 kb)
Acknowledgments
Funding
This study was supported by the National Natural Science Foundation of China (No. 31772257), the earmarked fund for Science and Technology Innovation of Fujian Agriculture and Forestry University (Nos. CXZX2017232 and CXZX2016148), and the earmarked fund for China Agriculture Research System (No. CARS27). The funding bodies had no role in the design, collection and interpretation of data or the manuscript writing.
Availability of data and materials
All data analyzed in this study are included in this published article and its additional files. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD010426.
Abbreviations
- 2-DE
2-dimensional electrophoresis
- AKR
Aldo-keto reductase
- APX
ASC peroxidase
- ASC
Ascorbate
- CAB
Chl a/b-binding protein
- Chl
Chlorophyll
- Cpn
Chaperonin
- DAP
Differentially abundant protein
- DHA
Dehydroascorbate
- ETR
Electron transport rate
- FER1
Ferritin-1
- FNR
Ferredoxin-NADP reductase
- Fv/Fm
Maximum PSII efficiency of dark-adapted leaves
- GRF8
14–3-3-like protein GF14 kappa
- IRL
Isoflavone reductase-like protein
- JA
Jasmonic acid
- LHC
Light harvesting complex
- MDA
Malondialdehyde
- MECT
2-C-methyl-D-erythritol 4-phosphate cytidylyltransferase
- MS
Mass spectrometry
- NPQ
Non-photochemical quenching
- OEC
Oxygen evolving complexes
- PGK
Phosphoglycerate kinase
- PMM
Phosphomannomutase
- PRPF31
U4/U6 small nuclear ribonucleoprotein PRP31
- PSBO2
Oxygen-evolving enhancer protein 1–1
- PSI
Photosystem I
- PSII
Photosystem II
- ROS
Reactive oxygen species
- Rubisco
Ribulose bisphosphate carboxylase/oxygenase
- RWC
Relative water content
- SOD
Superoxide dismutase
- TNC
Total nonstructural carbohydrates
- TROL
Rhodanese-like domain-containing protein 4A, chloroplastic
Authors’ contributions
JZ carried out most of the experiment, analyzed the data, and drafted the manuscript; QL, WH, NL and XY participated in the measurements of leaf gas exchange, qRT-PCR and data analysis; YQ and LY participated in the direction of this study; LC designed and directed the study and revised the manuscript. All authors have read and approved the final manuscript.
Ethics approval and consent to participate
Citrus sinensis and Citrus grandis seeds, which were public and available for non-commercial purpose, were collected from Minan village, Tingjiang town, Mawei district, Fuzhou city, China and Fujian Academy of Forestry Sciences, Fuzhou city, China, respectively. Collection of seeds complied with the institutional, national and international guidelines. No specific permits were required.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests. The author Li-Song Chen is an Associate Editor of BMC Plant Biology.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Jiang Zhang, Email: 771030197@qq.com.
Qiang Li, Email: 1554962076@qq.com.
Yi-Ping Qi, Email: qiyiping2008@hotmail.com.
Wei-Lin Huang, Email: 1527204185@qq.com.
Lin-Tong Yang, Email: talstoy@sina.com.
Ning-Wei Lai, Email: 20244715@qq.com.
Xin Ye, Email: yexin1000@163.com.
Li-Song Chen, Email: lisongchen2002@hotmail.com, Email: lisongchen@fafu.edu.cn.
References
- 1.von Uexküll HR, Mutert E. Global extent, development and economic impact of acid soils. Plant Soil. 1995;171:1–15. doi: 10.1007/BF00009558. [DOI] [Google Scholar]
- 2.Ferguson B, Lin MH, Gresshoff PM. Regulation of legume nodulation by acidic growth conditions. Plant Signal Behav. 2013;8:e23426. doi: 10.4161/psb.23426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Guo JH, Liu XJ, Zhang Y, Shen JL, Han WX, Zhang WF, Christie P, Goulding KWT, Vitousek PM, Zhang FS. Significant acidification in major Chinese croplands. Science. 2010;327:1008–1010. doi: 10.1126/science.1182570. [DOI] [PubMed] [Google Scholar]
- 4.Shi QH, Zhu ZJ, Juan LI, Qian QQ. Combined effects of excess Mn and low pH on oxidative stress and antioxidant enzymes in cucumber roots. Agri Sci China. 2006;5:767–772. doi: 10.1016/S1671-2927(06)60122-3. [DOI] [Google Scholar]
- 5.Yang LT, Qi YP, Jiang HX, Chen LS. Roles of organic acid anion secretion in aluminum tolerance of higher plants. Biomed Res Int. 2013;2013:173682. doi: 10.1155/2013/173682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bian M, Zhou M, Sun D, Li C. Molecular approaches unravel the mechanism of acid soil tolerance in plants. Crop J. 2013;1:91–104. doi: 10.1016/j.cj.2013.08.002. [DOI] [Google Scholar]
- 7.Kidd PS, Proctor J. Why plants grow poorly on very acid soils: are ecologists missing the obvious? J Exp Bot. 2001;52:791–799. doi: 10.1093/jexbot/52.357.791. [DOI] [PubMed] [Google Scholar]
- 8.Long A, Zhang J, Yang LT, Ye X, Lai NW, Tan LL, Lin D, Chen LS. Effects of low pH on photosynthesis, related physiological parameters and nutrient profile of Citrus. Front Plant Sci. 2017;8:185. doi: 10.3389/fpls.2017.00185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Koyama H, Toda T, Hara T. Brief exposure to low-pH stress causes irreversible damage to the growing root in Arabidopsis thaliana: pectin-ca interaction may play an important role in proton rhizotoxicity. J Exp Bot. 2001;52:361–368. [PubMed] [Google Scholar]
- 10.Yang M, Tan L, Xu Y, Zhao Y, Cheng F, Ye S, Jiang W. Effect of low pH and aluminum toxicity on the photosynthetic characteristics of different fast-growing Eucalyptus vegetatively propagated clones. PLoS One. 2015;10:e0130963. doi: 10.1371/journal.pone.0130963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Martins N, Osório ML, Gonçalves S, Osório J, Palma T, Romano A. Physiological responses of Plantago algarbiensis and P. almogravensis shoots and plantlets to low pH and aluminum stress. Acta Physiol Plant. 2013;35:615–625. doi: 10.1007/s11738-012-1102-z. [DOI] [Google Scholar]
- 12.Lazof DB, Holland MJ. Evaluation of the aluminium-induced root growth inhibition in isolation from low pH effects in Glycine max, Pisum sativum and Phaseolus vulgaris. Aust J Plant Physiol. 1999;26:147–157. doi: 10.1071/PP98072. [DOI] [Google Scholar]
- 13.Samac DA, Tesfaye M. Plant improvement for tolerance to aluminum in acid soils: a review. Plant Cell Tissue Organ Cult. 2003;75:189–207. doi: 10.1023/A:1025843829545. [DOI] [Google Scholar]
- 14.Kamaluddin M, Zwiazek JJ. Effects of root medium pH on water transport in paper birch (Betula papyrifera) seedlings in relation to root temperature and abscisic acid treatments. Tree Physiol. 2004;24:1173–1180. doi: 10.1093/treephys/24.10.1173. [DOI] [PubMed] [Google Scholar]
- 15.Zhang CP, Meng P, Li JZ, Wan XC. Interactive effects of soil acidification and phosphorus deficiency on photosynthetic characteristics and growth in Juglans regia seedlings. Chin J Plant Ecol. 2014;38:1345–1355. doi: 10.3724/SP.J.1258.2014.00129. [DOI] [Google Scholar]
- 16.Martins N, Osório ML, Gonçalves S, Osório J, Romano A. Differences in Al tolerance between Plantago algarbiensis and P. almogravensis reflect their ability to respond to oxidative stress. Biometals. 2013;26:427–437. doi: 10.1007/s10534-013-9625-3. [DOI] [PubMed] [Google Scholar]
- 17.Martins N, Gonçalves S, Palma T, Romano A. The influence of low pH on in vitro growth and biochemical parameters of Plantago almogravensis and P. Algarbiensis. Plant Cell Tissue Organ Cult. 2011;107:113–121. doi: 10.1007/s11240-011-9963-1. [DOI] [Google Scholar]
- 18.Martins N, Gonçalves S, Romano A. Metabolism and aluminum accumulation in Plantago almogravensis and P. algarbiensis in response to low pH and aluminum stress. Biol Plant. 2013;57:325–331. doi: 10.1007/s10535-012-0271-3. [DOI] [Google Scholar]
- 19.Yang M, Huang SX, Fang SZ, Huang XL. Response of seedling growth of four Eucalyptus clones to acid and aluminum stress. Plant Nutr Fert Sci. 2011;17:195–201. [Google Scholar]
- 20.Graças JP, Ruiz-Romero R, Figueiredo LD, Mattiello L, Peres LEP, Vitorello VA. Root growth restraint can be an acclimatory response to low pH and is associated with reduced cell mortality: a possible role of class III peroxidases and NADPH oxidases. Plant Biol. 2016;18:658–668. doi: 10.1111/plb.12443. [DOI] [PubMed] [Google Scholar]
- 21.Kobayashi Y, Ohyama Y, Kobayashi Y, Ito H, Iuchi S, Fujita M, Zhao CR, Tanveer T, Ganesan M, Kobayashi M, Koyama H. STOP2 activates transcription of several genes for Al- and low pH-tolerance that are regulated by STOP1 in Arabidopsis. Mol Plant. 2014;7:311–322. doi: 10.1093/mp/sst116. [DOI] [PubMed] [Google Scholar]
- 22.Anugoolprasert O, Kinoshita S, Naito H, Shimizu M, Ehara H. Effect of low pH on the growth, physiological characteristics and nutrient absorption of sago palm in a hydroponic system. Plant Prod Sci. 2012;15:125–131. doi: 10.1626/pps.15.125. [DOI] [Google Scholar]
- 23.Shavrukov Y, Hirai Y. Good and bad protons: genetic aspects of acidity stress responses in plants. J Exp Bot. 2016;67:15–30. doi: 10.1093/jxb/erv437. [DOI] [PubMed] [Google Scholar]
- 24.Lager IDA, Andréasson O, Dunbar TL, Andreasson E, Escobar MA, Rasmusson AG. Changes in external pH rapidly alter plant gene expression and modulate auxin and elicitor responses. Plant Cell Environ. 2010;33:1513–1528. doi: 10.1111/j.1365-3040.2010.02161.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Wang ZF, Wang ZH, Shi L, Wang LJ, Xu FS. Proteomic alterations of Brassica napus root in response to boron deficiency. Plant Mol Biol. 2010;74:265–278. doi: 10.1007/s11103-010-9671-y. [DOI] [PubMed] [Google Scholar]
- 26.Peng HY, Qi YP, Lee J, Yang LT, Guo P, Jiang HX, Chen LS. Proteomic analysis of Citrus sinensis roots and leaves in response to long-term magnesium-deficiency. BMC Genomics. 2015;16:253. doi: 10.1186/s12864-015-1462-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Yuda E, Okamoto S. The effect of soil reaction on the growth of young citrus plants. I. Forms of nitrogen fertilizer and kinds of pH adjusting agent. J Jap Soc Hort Sci. 1965;34:177–186. doi: 10.2503/jjshs.34.177. [DOI] [Google Scholar]
- 28.Li Y, Han MQ, Lin F, Ten Y, Lin J, Zhu DH, Guo P, Weng YB, Chen LS. Soil chemical properties, ‘Guanximiyou’ pummelo leaf mineral nutrient status and fruit quality in the southern region of Fujian province, China. J Soil Sci Plant Nutr. 2015;15:615–628. [Google Scholar]
- 29.Han S, Chen LS, Jiang HX, Smith BR, Yang LT, Xie CY. Boron deficiency decreases growth and photosynthesis, and increases starch and hexoses in leaves of Citrus seedlings. J Plant Physiol. 2008;165:1331–1341. doi: 10.1016/j.jplph.2007.11.002. [DOI] [PubMed] [Google Scholar]
- 30.Chen LS, Qi YP, Liu XH. Effects of aluminum on light energy utilization and photoprotective systems in Citrus leaves. Ann Bot. 2005;96:35–41. doi: 10.1093/aob/mci145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hodges DM, DeLong JM, Forney CF, Prange RK. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta. 1999;207:604–611. doi: 10.1007/s004250050524. [DOI] [PubMed] [Google Scholar]
- 32.You X, Yang LT, Lu YB, Li H, Zhang SQ, Chen LS. Proteomic changes of citrus roots in response to long-term manganese toxicity. Trees Struct Funct. 2014;28:1383–1399. doi: 10.1007/s00468-014-1042-x. [DOI] [Google Scholar]
- 33.Bradford MM. A rapid and sensitive method for quantitation of microgram and quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- 34.Sang W, Huang ZR, Qi YP, Yang LT, Guo P, Chen LS. An investigation of boron-toxicity in leaves of two citrus species differing in boron-tolerance using comparative proteomics. J Proteome. 2015;123:128–146. doi: 10.1016/j.jprot.2015.04.007. [DOI] [PubMed] [Google Scholar]
- 35.Sang W, Huang ZR, Yang LT, Guo P, Ye X, Chen LS. Effects of high toxic boron concentration on protein profiles in roots of two citrus species differing in boron-tolerance revealed by a 2-DE based MS approach. Front Plant Sci. 2017;8:180. doi: 10.3389/fpls.2017.00180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yang LT, Liu JW, Wu YM, Qi YP, Wang JL, Lai NW, Ye X, Chen LS. Proteome profile analysis of boron-induced alleviation of aluminum-toxicity in Citrus grandis roots. Ecotoxicol Environ Saf. 2018;162:488–498. doi: 10.1016/j.ecoenv.2018.07.028. [DOI] [PubMed] [Google Scholar]
- 37.Figueiredo A, Martins J, Sebastiana M, Guerreiro A, Silva A, Matos AR, Monteiro F, Pais MS, Roepstorff P, Coelho AV. Specific adjustments in grapevine leaf proteome discriminating resistant and susceptible grapevine genotypes to Plasmopara viticola. J Proteome. 2017;152:48–57. doi: 10.1016/j.jprot.2016.10.012. [DOI] [PubMed] [Google Scholar]
- 38.Yang LT, Qi YP, Lu YB, Guo P, Sang W, Feng H, Zhang HX, Chen LS. iTRAQ protein profile analysis of Citrus sinensis roots in response to long-term boron-deficiency. J Proteome. 2013;93:179–206. doi: 10.1016/j.jprot.2013.04.025. [DOI] [PubMed] [Google Scholar]
- 39.Guo P, Qi YP, Huang WL, Yang LT, Huang ZR, Lai NW, Chen LS. Aluminum-responsive genes revealed by RNA-Seq and related physiological responses in leaves of two Citrus species with contrasting aluminum-tolerance. Ecotoxicol Environ Saf. 2018;158:213–222. doi: 10.1016/j.ecoenv.2018.04.038. [DOI] [PubMed] [Google Scholar]
- 40.Zhou CP, Qi YP, You X, Yang LT, Guo P, Ye X, Zhou XX, Ke FJ, Chen LS. Leaf cDNA-AFLP analysis of two citrus species differing in manganese tolerance in response to long-term manganese-toxicity. BMC Genomics. 2013;14:621. doi: 10.1186/1471-2164-14-621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Jones HG. Limiting factors in photosynthesis. New Phytol. 1973;72:1089–1094. doi: 10.1111/j.1469-8137.1973.tb02086.x. [DOI] [Google Scholar]
- 42.Damkjær JT, Kereiche S, Johnson MP, Kovacs L, Kiss AZ, Boekema EJ, Ruban AV, Horton P, Jansson S. The photosystem II light-harvesting protein Lhcb3 affects the macrostructure of photosystem II and the rate of state transitions in Arabidopsis. Plant Cell. 2009;21:3245–3256. doi: 10.1105/tpc.108.064006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Yi X, McChargue M, Laborde S, Frankel LK, Bricker TM. The manganese-stabilizing protein is required for photosystem II assembly/stability and photoautotrophy in higher plants. J Biol Chem. 2005;280:16170–16174. doi: 10.1074/jbc.M501550200. [DOI] [PubMed] [Google Scholar]
- 44.Lundin B, Hansson M, Schoefs B, Vener AV, Spetea C. The Arabidopsis PsbO2 protein regulates dephosphorylation and turnover of the photosystem II reaction Centre D1 protein. Plant J. 2007;49:528–539. doi: 10.1111/j.1365-313X.2006.02976.x. [DOI] [PubMed] [Google Scholar]
- 45.Srivastava A, Guisse B, Greppin H, Strasser RJ. Regulation of antenna structure and electron transport in photosystem II of Pisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP. Biochim Biophys Bioenergetics. 1997;1320:95–106. doi: 10.1016/S0005-2728(97)00017-0. [DOI] [Google Scholar]
- 46.Salvucci ME, Portis AR, Jr, Ogren WL. A soluble chloroplast protein catalyzes ribulose bisphosphate carboxylase/oxygenase activation in vivo. Photosynth Res. 1985;7:193–201. doi: 10.1007/BF00037012. [DOI] [PubMed] [Google Scholar]
- 47.Mate CJ, von Caemmerer S, Evans JR, Hudson GS, Andrews TJ. The relationship between CO2-assimilation rate, Rubisco carbamylation and Rubisco activase content in activase-deficient transgenic tobacco suggests a simple model of activase action. Planta. 1996;198:604–613. doi: 10.1007/BF00262648. [DOI] [PubMed] [Google Scholar]
- 48.Portis AR., Jr Rubisco activase-Rubisco’s catalytic chaperone. Photosynth Res. 2003;75:11–27. doi: 10.1023/A:1022458108678. [DOI] [PubMed] [Google Scholar]
- 49.Suzuki K, Nakanishi H, Bower J, Yoder DW, Osteryoung KW, Miyagishima SY. Plastid chaperonin proteins Cpn60α and Cpn60β are required for plastid division in Arabidopsis thaliana. BMC Plant Biol. 2009;9:38. doi: 10.1186/1471-2229-9-38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Lintala M, Allahverdiyeva Y, Kangasjärvi S, Lehtimäki N, Keränen M, Rintamäki E, Aro EM, Mulo P. Comparative analysis of leaf-type ferredoxin-NADP+ oxidoreductase isoforms in Arabidopsis thaliana. Plant J. 2009;57:1103–1115. doi: 10.1111/j.1365-313X.2008.03753.x. [DOI] [PubMed] [Google Scholar]
- 51.Jurić S, Hazler-Pilepić K, Tomašić A, Lepeduš H, Jeličić B, Puthiyaveetil S, Bionda T, Vojta L, Allen JF, Schleiff E, Fulgosi H. Tethering of ferredoxin: NADP+ oxidoreductase to thylakoid membranes is mediated by novel chloroplast protein TROL. Plant J. 2009;60:783–794. doi: 10.1111/j.1365-313X.2009.03999.x. [DOI] [PubMed] [Google Scholar]
- 52.Hsieh HM, Chang CY, Hsu SJ, Chen JJ. Chloroplast localization of methylerythritol 4-phosphate pathway enzymes and regulation of mitochondrial genes in ispD and ispE albino mutants in Arabidopsis. Plant Mol Biol. 2008;66:663–673. doi: 10.1007/s11103-008-9297-5. [DOI] [PubMed] [Google Scholar]
- 53.Sheen J. Feedback control of gene expression. Photosynth Res. 1994;39:427–438. doi: 10.1007/BF00014596. [DOI] [PubMed] [Google Scholar]
- 54.Cave G, Tolley LC, Stain BR. Effect of carbon dioxide enrichment on chlorophyll content, starch content and starch grain structure in Trifolium subterraneum leaves. Physiol Plant. 1981;51:171–174. doi: 10.1111/j.1399-3054.1981.tb02694.x. [DOI] [Google Scholar]
- 55.Hisabori T, Konno H, Ichimura H, Strotmann H, Bald D. Molecular devices of chloroplast F1-ATP synthase for the regulation. Biochim Biophys Acta Bioenerg. 2002;1555:140–146. doi: 10.1016/S0005-2728(02)00269-4. [DOI] [PubMed] [Google Scholar]
- 56.Gossett DR, Millhollon EP, Lucas MC. Antioxidant responses to NaCl stress in salt-tolerant and salt-sensitive cultivars of cotton. Crop Sci. 1994;34:706–714. doi: 10.2135/cropsci1994.0011183X003400030020x. [DOI] [Google Scholar]
- 57.Guo P, Qi YP, Cai YT, Yang TY, Yang LT, Huang ZR, Chen LS. Aluminum effects on photosynthesis, reactive oxygen species and methylglyoxal detoxification in two Citrus species differing in aluminum tolerance. Tree Physiol. 2018; 10.1093/treephys/tpy035. [DOI] [PubMed]
- 58.Guo P, Li Q, Qi YP, Yang LT, Ye X, Chen HH, Chen LS. Sulfur-mediated-alleviation of aluminum-toxicity in Citrus grandis seedlings. Int J Mol Sci. 2017;18:2570. doi: 10.3390/ijms18122570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Hoeberichts FA, Vaeck E, Kiddle G, Coppens E, van de Cotte B, Adamantidis A, Ormenese S, Foyer CH, Zabeau M, Inzé D, Périlleux C, Breusegem FV, Vuylsteke M. A temperature-sensitive mutation in the Arabidopsis thaliana phosphomannomutase gene disrupts protein glycosylation and triggers cell death. J Biol Chem. 2008;283:5708–5718. doi: 10.1074/jbc.M704991200. [DOI] [PubMed] [Google Scholar]
- 60.Qian W, Yu C, Qin H, Liu X, Zhang A, Johansen IE, Wang D. Molecular and functional analysis of phosphomannomutase (PMM) from higher plants and genetic evidence for the involvement of PMM in ascorbic acid biosynthesis in Arabidopsis and Nicotiana benthamiana. Plant J. 2007;49:399–413. doi: 10.1111/j.1365-313X.2006.02967.x. [DOI] [PubMed] [Google Scholar]
- 61.Busch A, Rimbauld B, Naumann B, Rensch S, Hippler M. Ferritin is required for rapid remodeling of the photosynthetic apparatus and minimizes photo-oxidative stress in response to iron availability in Chlamydomonas reinhardtii. Plant J. 2008;55:201–211. doi: 10.1111/j.1365-313X.2008.03490.x. [DOI] [PubMed] [Google Scholar]
- 62.Bournier M, Tissot N, Mari S, Boucherez J, Lacombe E, Briat JF, Gaymard F. Arabidopsis ferritin 1 (AtFer1) gene regulation by the phosphate starvation response 1 (AtPHR1) transcription factor reveals a direct molecular link between iron and phosphate homeostasis. J Biol Chem. 2013;288:22670–22680. doi: 10.1074/jbc.M113.482281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yang LT, Jiang HX, Qi YP, Chen LS. Differential expression of genes involved in alternative glycolytic pathways, phosphorus scavenging and recycling in response to aluminum and phosphorus interactions in Citrus roots. Mol Biol Rep. 2012;39:6353–6366. doi: 10.1007/s11033-012-1457-7. [DOI] [PubMed] [Google Scholar]
- 64.Gaxiola RA, Palmgren MG, Schumacher K. Plant proton pumps. FEBS Let. 2007;581:2204–2214. doi: 10.1016/j.febslet.2007.03.050. [DOI] [PubMed] [Google Scholar]
- 65.Krebs M, Beyhl D, Görlich E, Al-Rasheid KAS, Marten I, Stierhof YD, Hedrich R, Schumacher K. Arabidopsis V-ATPase activity at the tonoplast is required for efficient nutrient storage but not for sodium accumulation. Proc Natl Acad Sci U S A. 2010;107:3251–3256. doi: 10.1073/pnas.0913035107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Tang RJ, Liu H, Yang Y, Yang L, Gao XS, Garcia VJ, Luan S, Zhang HX. Tonoplast calcium sensors CBL2 and CBL3 control plant growth and ion homeostasis through regulating V-ATPase activity in Arabidopsis. Cell Res. 2012;22:1650–1665. doi: 10.1038/cr.2012.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Specific primer pairs used for qRT-PCR analysis. (DOC 57 kb)
Figure S1. Two-DE images of proteins extracted from pH 2.5- (a, d, g, j), pH 3.0- (b, e, h, k) and pH 6.0-treated (c, f, i, l) C. grandis (a-f) and C. sinensis (g-l) leaves for the other two replicates. (PDF 317 kb)
Table S2. Master list of proteins identified in MALDI TOF/TOF MS from pH 2.5 and/or pH 3-treated C. sinensis leaves using 2DE and DIGE experiments. (DOC 1635 kb)
Table S3. Master list of proteins identified in MALDI TOF/TOF MS from pH 2.5 and/or pH 3-treated C. grandis leaves using 2DE and DIGE experiments. (DOC 1467 kb)
Figure S2. Close-up views of 22 DAP spots in pH 2.5, pH 3 and pH 6-treated C. grandis and C. sinensis leaves. (PDF 132 kb)
Data Availability Statement
All data analyzed in this study are included in this published article and its additional files. The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD010426.