

Abstract
Background
Geraniol is a monoterpene alcohol that has anti-fungal, anti-cancer and anti-nociceptive properties, but its anti-allergic rhinitis (AR) property is unclear.
Methods
In this study, the anti-inflammatory role and its possible mechanisms of geraniol in human mast cell line (HMC-1) cells stimulated by inflammatory trigger phorbol 12-myristate 13-acetate plus A23187 (PMACI), as well as in ovalbumin (OVA)-induced AR mice models were investigated.
Results
PMACI results in a significant increase in the production of proinflammatory cytokines, such as TNF-α, IL-1β, MCP-1, IL-6 and as well as histamine. Geraniol was found to inhibit both TNF-α, IL-1β and IL-6 protein and mRNA expressions at concentrations of 40, 80, 160 μM. In OVA-induced AR models, geraniol treatment was able to suppress AR biomarkers (OVA-specific IgE and IL-1β as well as histamine) and nasal rub scores. Interestingly, p38, a member of the mitogen-activated protein kinase (MAPK) signaling family, was found to be increasingly hypophosphorylated as geraniol dose was increased. Similar decreases in the nuclear level of p65, a member of the nuclear factor kappa B (NF-κB) signaling pathway, were also observed.
Conclusion
Our data highlights that the anti-inflammatory properties of geraniol on AR-related markers in activated HCM-1 cells and OVA-induced AR models may be mediated through the regulation of the MAPK/NF-κB signaling pathway.
Keywords: geraniol, HMC-1, allergic rhinitis, PMACI
Introduction
An estimated 10%–25% of the global population is believed to suffer from allergic rhinitis (AR), a common form of allergic disease.1 AR encompasses a constellation of symptoms that arise in response to airborne allergens, is marked by perennial or seasonal nasal itching and congestion, rhinorrhea, and sneezing, and is frequently accompanied by pharyngitis and conjunctivitis. AR is an immunological disorder whose pathophysiological processes and disease manifestations are primarily facilitated by type I inflammatory processes and is mediated by innate immune cells (such as mast cells) and T-helper type 2 (Th2) cells.2 Allergic inflammation is initiated by immune cell sensitization, which causes the production of antigen-specific IgE. Early and late phases of the type I immune response occur following sensitization. In combination, these responses drive allergic inflammation and facilitate the spectrum of AR-associated symptoms.3
An exposure to an allergen triggers the early phase within minutes and causes cross-linking of IgE expressed on the surfaces of mast cells in the nasal mucosa. This culminates in a burst of various preformed inflammatory mediators including tryptase, histamine, vascular endothelial growth factor A, tumor necrosis factor (TNF), prostaglandin D2, and leukotrienes (LTB4, LTC4, LTD4, and LTE4).4 Mast cell-derived inflammatory mediators and their respective physiological actions collectively contribute to the late-phase response. Newly synthesized cytokines are also released upon mast cell IgE cross-linking, such as granulocyte–macrophage colony-stimulating factor, TNF, interleukin (IL)-5, and chemokines such as CCL2 and IL-8.3,4 These mediators are vital in recruiting and activating innate and adaptive immune cells, including monocytes, neutrophils, eosinophils, and Th2 cells.
In recent years, several reports regarding the utility of natural herb-based compounds in the treatment of allergic diseases have emerged. Yen et al5 reported that about 63.1% of children in China with newly diagnosed AR expressed a preference for Chinese medicinal herbs, and about 22.4% of these children would be prescribed Xin Yi Qing Fei Tang (Magnolia flower, lung-clearing decoction), the major component of which is Flos Magnoliae (Chinese name: Xin-Yi). Several clinical studies have demonstrated the efficacy of Flos Magnoliae and its mixed herbal formulas in the treatment of AR.6–8 Moreover, Flos Magnoliae and its extracts are able to inhibit several allergic and inflammatory responses by modulating the expression of nitric oxide (NO), platelet activity factor, prostaglandin E2, histamine, and proinflammatory cytokines such as TNF-α and IL-1α.9,10 Extracts of Flos Magnoliae – epimagnolin and fargesin – can decrease NO production (NO being a marker of pulmonary inflammation) and attenuate inducible NO synthase (iNOS) expression in cytokine-stimulated human respiratory epithelial cells by acting on an extracellular signal-regulated kinase (ERK) pathway.10
Geraniol (C10H18O; Figure 1) is a natural acyclic mono-terpenoid extracted from Flos Magnoliae. It has been reported to have anticancer, antifungal, and anti-nociceptive properties;11–13 however, the overall effect of geraniol has not yet been clearly defined. In this study, we sought to examine its utility as a potential AR treatment.
Figure 1.
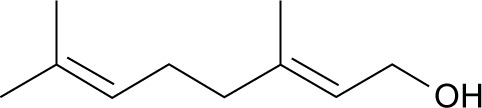
Chemical structure of geraniol.
Materials and methods
Experimental reagents and cell cultures
Geraniol was obtained from the Control of Pharmaceutical and Biological Products (Beijing, China). Its purity was at least 95% as determined by high-performance liquid chromatography (HPLC). Geraniol was then dissolved in dimethyl sulphoxide (DMSO) to produce 160 mmol/L concentrated stock solutions. Subsequently, solutions of 40, 80, and 160 μmol/L concentrations were formed by dissolving geraniol stock solution in Roswell Park Memorial Institute (RPMI) 1640 medium with 10% heat-inactivated fetal bovine serum (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 100 mg/mL streptomycin and 100 unit/mL penicillin (Thermo Fisher Scientific). Phorbol 12-myristate 13-acetate (PMA), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), calcium ionophore (A23187), DMSO, and phosphate-buffered saline (PBS) were procured from Sigma-Aldrich Co. (St Louis, MO, USA). Human mast cell line (HMC-1) cells were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA) and maintained in RPMI 1640 medium in Dulbecco’s Modified Eagle’s Medium (DMEM; Thermo Fisher Scientific) at 95% humidity in 5% CO2 at 37°C. HMC-1 cells were exposed to geraniol for 1 hour before treatment with PMACI (20 nM of PMA plus 1 μM of A23187) and incubated at 37°C.14
Western blot analysis
Total protein extraction was performed on the transfected cells, which were subsequently quantified with a bicinchoninic acid assay kit (Beyotime, Nantong, China). Aliquots of 50 ng proteins were loaded onto 12% sodium dodecyl sulfate poly-acrylamide gel electrophoresis (SDS-PAGE) gel first before transferring the loaded gels onto 0.45 μm pore polyvinylidene difluoride (PVDF) membranes (EMD Millipore, Billerica, MA, USA). A mixture of 5% skim milk in tris-buffered saline in Tween 20 (TBST) was then used to block reactions for 60 minutes at room temperature. These membranes were left to incubate with their corresponding primary antibodies overnight at 4°C. The following day, washing of the membranes was performed thrice with TBST, and membranes were left to incubate further with a secondary antibody for 1 hour at room temperature. Enhanced chemiluminescence (ECL) solution enabled visualization of the immunoreactivity, using synchronously detected GAPHD as a loading control. Antibodies against p38, p-p38, p65, and actin were bought from Santa Cruz Biotechnology Inc. (Dallas, TX, USA).
Cell viability assay
The MTT assay allowed us to determine the viability of HMC-1 cells with or without PMACI stimulation. Briefly, 96-well plates were used for seeding HMC-1 cells at an initial density of 5×104 cells/well. After treatment with geraniol, cell numbers were monitored for 24 hours. Following this observation period, each well received 10 μL of MTT reagent. The cells were then incubated for an additional 2 hours at 37°C. The absorbance of each group of cells was determined at 490 nm with the help of a microplate enzyme-linked immunosorbent assay (ELISA) reader (Bio-Rad Laboratories Inc., Hercules, CA, USA).
Histamine assay
HMC-1 cells (1 × 106 cells/mL) were initially treated with geraniol for 1 hour, followed by a 6-hour incubation period with PMACI. The cell suspension was then centrifuged at 400× g for 5 minutes at 4°C. The supernatant, which contains released HMC-1 cell histamine, was collected and quantified via an o-phthalaldehyde (OPA)-based spectrofluorimetric procedure. The intensity of fluorescence was determined at 460 nm (excitation at 355 nm) with a spectrofluorometer.
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA of each sample was isolated using TRIzol reagent (Thermo Fisher Scientific) according to the manufacturer’s protocol. The quality and concentration of extracted RNAs were identified by measuring the absorbance at 260 nm. First-strand cDNAs were generated with the PrimeScript RT Master Mix Perfect Real Time kit (Takara Biotechnology Co., Ltd., Dalian, China) by mixing the components and incubated at 42°C for 1 hour. All qPCRs were performed with SYBR Premix Ex Taq kit (Takara Biotechnology Co., Ltd.) on an ABI PRISM 7500 Real-Time system (Thermo Fisher Scientific); initial denaturation: 95°C for 5 minutes; annealing: 60°C for 30 seconds; final extension: 72°C for 5 minutes. The cycling count is 40. The primers used are listed in Table 1, and GAPDH was included as the internal control. Each experiment was performed in triplicate at least three times. The primer sequences used are shown in Table 1.
Table 1.
Primer sequences used for the qPCR analysis
| Gene | Forward primer | Reverse primer |
|---|---|---|
| GAPDH | 5′-AGGGCTGCTTTTAACTCTGGT-3 | 5′-CCCCACTTGATTTTGGAGGGA |
| IL-6 | 5′-ACAACCACGGCCTTCCCTACTT | 5′-CACGATTTCCCAGAGAACATGTG |
| MCP-1 | 5′-CCACTCACCTGCTGCTACTCAT | 5′-TGGTGATCCTCTTGTAGCTCTCC |
| TNF-α | 5′-CAGCCTCTTCTCCTTCCTGAT | 5′-GCCAGAGGGCTGATTAGAGA |
| IL-1β | 5′-AACCTGCTGGTGTGTGACGTTC | 5′-CAGCACGAGGCTTTTTTGTTGT |
Abbreviations: IL, interleukin; qPCR, quantitative polymerase chain reaction; TNF, tumor necrosis factor.
ELISA assay
TNF-α, IL-6, IL-1β, and MCP-1 ELISA Duoset kits were purchased from R&D Systems (R&D Systems, Inc., Minneapolis, MN, USA) to analyze cytokine expressions in the supernatant. All procedures were performed following the manufacturer’s protocols. The percentage of inhibition was calculated using the following formula:
Ovalbumin (OVA)-induced AR animal model
Six-to-eight-week-old female Balb/c mice weighing between 18 and 20 g were obtained from the Shanghai Laboratory Animal Center of the Chinese Academy of Sciences (Shanghai, China) and reared in specific pathogen-free (SPF) conditions in strict accordance with the ethics committee of Children Hospital, School of Medicine, Shanghai Fudan University. To create AR mice models, mice were allergen sensitized on the 1st, 5th, and 14th days with intraperitoneal injections of 100 μg emulsified OVA and 20 mg aluminum hydroxide (Sigma-Aldrich Co.) in 100 μL of PBS. AR mice models were then subjected to an intranasal allergen challenge using 2 μL of 1.5 mg OVA, while control groups were given intranasal PBS (n=5). Mice were fed for 10 days with oral geraniol (100 mg/kg) and a control vehicle (0.25 mL of corn oil per 100 g body weight) prior to the intranasal OVA challenge. The number of nasal rubs (which is a marker of AR symptoms) that occurred in 10 minutes was counted after OVA intranasal provocation on day 10 after the challenge. Levels of IgE specific for OVA were also measured in AR models.
Statistical analyses
Results were expressed as mean ± SD. Comparisons between intervention and control data were made with the Student’s t-test. A probability of P<0.05 was determined to confer a statistical significance.
Results
Geraniol inhibits histamine production and mast cell proinflammatory cytokine expressions
We first examined the effects of geraniol and PMACI on HMC-1 viability. Varying concentrations of geraniol (40–160 μmol/L) were used to treat HMC-1 cells for 24 hours with or without PMACI stimulation. Both PMACI and geraniol were found to have no effect on HMC-1 cell viability (Figure 2A). We then explored the effects of geraniol in PMACI-stimulated HMC-1 cells. Mast cell degranulation in the nasal mucosa released pro-inflammatory cytokines such as TNF-α, IL-1β, IL-8, and IL-6, chemokines such as MCP-1, and histamine, a major biogenic amine. In line with previous reports, we demonstrated that PMACI significantly enhanced HMC-1 cell expression of TNF-α, IL-1β, IL-6, and MCP-1 as well as histamine (Figure 2B–F). On the other hand, geraniol concentrations of 40, 80, and 160 μmol/L attenuated expression levels of TNF-α, IL-1β, and IL-6. Thirty percent, 21.6%, and 27.0% of IL-1β, IL-6, and TNF-α, respectively, were inhibited at geraniol concentrations of 160 μmol/L. Geraniol did not significantly alter MCP-1 expression. Geran-iol was also found to inhibit mRNA expression of TNF-α, IL-1β, IL-6, and MCP-1 (Figure 3A–D).
Figure 2.

Geraniol inhibits histamine, TNF-α, IL-1β, IL-6, and MCP-1 expressions.
Notes: (A) The effect of PMACI and geraniol on HMC-1 cell viability using MTT assay. (B–F) HMC-1 cells were treated with geraniol (40, 80, and 160 μmol/L) for 1 hour prior to stimulation with PMACI for the next 6 hours. The histamine content of the supernatant was measured with a histamine assay, while the TNF-α, IL-1β, IL-6, and MCP-1 content of the supernatant was measured with ELISA. *P<0.05; **P<0.01.
Abbreviations: ELISA, enzyme-linked immunosorbent assay; HMC-1, human mast cell line; IL, interleukin; MTT, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; PMACI, phorbol 12-myristate 13-acetate plus A23187; TNF, tumor necrosis factor.
Figure 3.

Geraniol inhibits mRNA expression of TNF-α, IL-1β, and IL-6.
Notes: (A–D) HMC-1 cells were treated with geraniol (40, 80, and 160 μmol/L) for 1 hour prior to stimulation with PMACI for the next 6 hours. The IL-1β content of the supernatant was measured by RT-PCR. Error bars indicate mean ± SE of three independent experiments (*P<0.05; **P<0.01; ***P<0.001).
Abbreviations: IL, interleukin; HMC-1, human mast cell line; PMACI, phorbol 12-myristate 13-acetate plus A23187; RT-PCR, real-time polymerase chain reaction; SE, standard error; TNF, tumor necrosis factor.
Geraniol inhibits p38 phosphorylation and nuclear p65 activation in HMC-1 cells
We analyzed mitogen-activated protein kinase (MAPK) and nuclear factor kappa B (NF-κB) signaling pathway activation to further understand how geraniol modulates the allergic inflammatory response. Western blot analysis showed that treatment with geraniol in HMC-1 cells diminished levels of phosphorylated p38 (an MAPK family member), but maintained total p38 levels. On the other hand, total levels of nuclear p65 (an NF-κB family member) decreased, while there was a significant increase in cytosolic p65 levels (Figure 4). This evidence suggests that geraniol attenuates MAPK and NF-κB signaling in HMC-1 cells.
Figure 4.
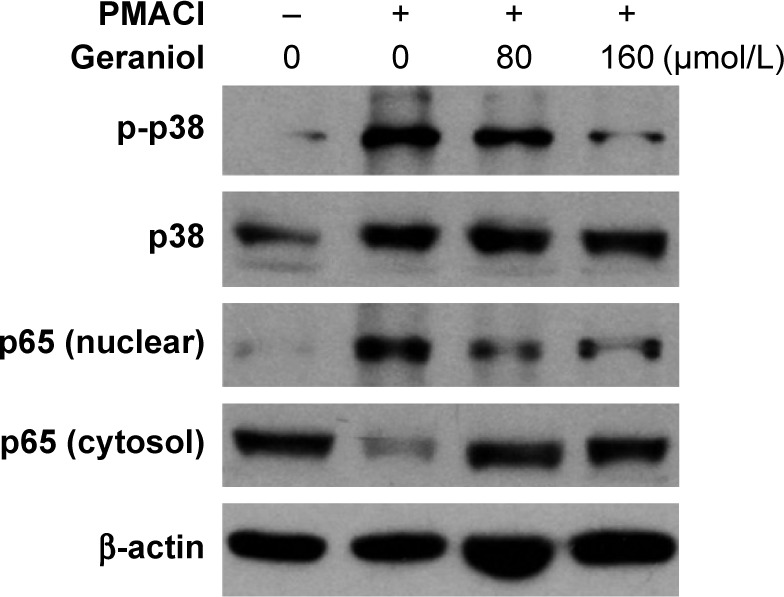
Geraniol mediates activation of NF-κB and MAPK signaling pathways.
Notes: HMC-1 cells were pretreated with various concentrations of geraniol for 1 hour and stimulated with PMACI for 6 hours. The expression of both phosphorylated and non-phosphorylated p38, as well as nuclear and cytosolic p65, was detected by Western blot. Error bars indicate mean ± SE of three independent experiments.
Abbreviations: HMC-1, human mast cell line; MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor kappa B; PMACI, phorbol 12-myristate 13-acetate plus A23187; SE, standard error.
Geraniol reduces clinical symptoms and OVA-specific IgE level in the AR animal model
AR mice models were given different doses of geraniol to assess its in vivo effect on AR. We demonstrated that 100 mg/kg of geraniol treatment in AR mice models was able to significantly inhibit nasal rub scores in comparison to the control group (Figure 5A). OVA-specific IgE level was also attenuated by geraniol in AR mice models, a finding that correlated with the reduction in AR symptoms experienced by mice (Figure 5B). Figure 5C and D depicts serum histamine and IL-1β levels. Overall, these findings support geraniol as an inflammatory inhibitor in AR mice models.
Figure 5.

Geraniol reduced clinical symptoms of AR in an AR mouse model.
Notes: (A) The number of nasal rubs that occurred in 10 minutes after intranasal OVA provocation (n=5). (B) Serum was isolated from blood to measure OVA-specific IgE levels, which were quantified using ELISA (n=5). (C, D) Serum was isolated from blood to measure histamine (C) and IL-1β (D). Error bars indicate mean ± SE of three independent experiments (*P<0.05; **P<0.01; ***P<0.01).
Abbreviations: AR, allergic rhinitis; ELISA, enzyme-linked immunosorbent assay; IL, interleukin; OVA, ovalbumin; PBS, phosphate-buffered saline; SE, standard error.
Discussion
Chinese herbal medicines have been used for thousands of years to manage AR-related symptoms. Indeed, several randomized controlled trials have already demonstrated the safety and efficacy of Chinese herbal medicine when used for treating AR. Geraniol is a herb well known for its antifungal, anticancer, anti-nociceptive, antioxidant, and antidepressant-like effects.11,12,15,16 In this study, we have successfully demonstrated geraniol’s therapeutic effect in in vitro and in vivo AR models with special focus on the underlying anti-inflammatory mechanisms.
The early phase of AR is characterized by mast cell release of protease, cytokines, growth factors, and histamine. Histamine is the primary mediator responsible for enhanced nasal mucus secretion, rhinorrhea, sneezing, and pruritus – all of which are found in AR. As such, AR treatment often involves the use of H1 antihistamines which block histamine release.17 In our study, our results show that both 80 and 160 μmol/L of geraniol are able to stop histamine release by activated HMC-1 cells. This indirectly indicates that geraniol can inhibit mast cell degranulation. Moreover, it is well known that mast cell degranulation gives rise to inflammatory mediators such as TNF-α, IL-1β, and IL-6. Geraniol was also shown to possess the capability of inhibiting proinflammatory cytokines such as TNF-α, IL-1β, and IL-6 in PMACI-simulated HMC-1 cells. Overall, our results strongly suggest that geraniol can hinder mast cell degranulation in response to an allergen.
AR models frequently utilize chicken egg-derived OVA as an allergic trigger, as it is able to induce the classic inflammatory response consisting of inflammatory cell infiltration in nasal mucosa as well as a rise in serum inflammatory markers of histamine, IgE, and other mediators.18 In our study, we used OVA-induced AR mice models to evaluate geraniol’s anti-allergic properties in vivo. We found that geraniol treatment remarkably decreased nasal rub scores in AR mice models in comparison to control groups. Significant attenuations of serum IL-1β, histamine, and OVA-specific IgE were also observed in the AR models. All these observations suggest that geraniol imparted an anti-allergic effect on AR mice models.
The initiation of the inflammatory response relies heavily on MAPK signaling and NF-κB signaling pathways.19 Existing literature reports that p38 MAPK inhibitors are able to suppress monocyte and mast cell production of TNF-α and IL-6. Furthermore, NF-κB has to modulate key components of the inflammatory response, namely leukocyte recruitment, proinflammatory cytokine production, and cell survival. Accordingly, our data showed that levels of phosphorylated mediators belonging to the MAPK signaling pathway and NF-κB signaling pathway in HMC-1 cells were attenuated upon exposure to geraniol treatment, supporting the regulatory effect of geraniol on these pathways.
Conclusion
We found that geraniol has an anti-inflammatory effect in PMACI-stimulated mast cells, as evidenced by its ability to reduce proinflammatory cytokines such as TNF-α, IL-1β, and IL-6 expressions. Geraniol’s anti-inflammatory effects were able to be replicated in vivo in OVA-induced AR models, shown by its capacity to alleviate AR symptoms in mice and diminish levels of serum histamine, OVA-specific IgE, and IL-1β levels. We also successfully demonstrated that these anti-inflammatory effects are due to alterations in MAPK/NF-κB signaling pathways (Figure 6).
Figure 6.
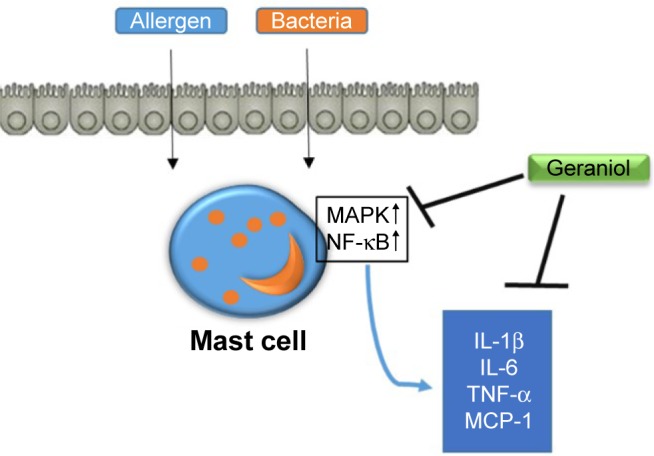
Proposed mechanism of geraniol-mediated anti-inflammatory effect in PMACI-stimulated HMC-1 cells.
Abbreviations: HMC-1, human mast cell line; IL, interleukin; MAPK, mitogen-activated protein kinase; NF-κB, nuclear factor kappa B; PMACI, phorbol 12-myristate 13-acetate plus A23187; TNF, tumor necrosis factor.
Footnotes
Disclosure
The authors report no conflicts of interest in this work.
References
- 1.Licari A, Ciprandi G, Marseglia A, et al. Current recommendations and emerging options for the treatment of allergic rhinitis. Expert Rev Clin Immunol. 2014;10(10):1337–1347. doi: 10.1586/1744666X.2014.955476. [DOI] [PubMed] [Google Scholar]
- 2.Palm NW, Rosenstein RK, Medzhitov R. Allergic host defences. Nature. 2012;484(7395):465–472. doi: 10.1038/nature11047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Galli SJ, Tsai M, Piliponsky AM. The development of allergic inflammation. Nature. 2008;454(7203):445–454. doi: 10.1038/nature07204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Howarth PH, Salagean M, Dokic D. Allergic rhinitis: not purely a histamine-related disease. Allergy. 2000;55(suppl 64):7–16. doi: 10.1034/j.1398-9995.2000.00802.x. [DOI] [PubMed] [Google Scholar]
- 5.Yen HR, Liang KL, Huang TP, Fan JY, Chang TT, Sun MF. Characteristics of traditional Chinese medicine use for children with allergic rhinitis: a nationwide population-based study. Int J Pediatr Otorhino-laryngol. 2015;79(4):591–597. doi: 10.1016/j.ijporl.2015.02.002. [DOI] [PubMed] [Google Scholar]
- 6.Yang SH, Yu CL, Chen YL, Chiao SL, Chen ML. Traditional Chinese medicine, Xin-yi-san, reduces nasal symptoms of patients with perennial allergic rhinitis by its diverse immunomodulatory effects. Int Immunopharmacol. 2010;10(8):951–958. doi: 10.1016/j.intimp.2010.05.008. [DOI] [PubMed] [Google Scholar]
- 7.Shen Y, Li CG, Zhou SF, Pang EC, Story DF, Xue CC. Chemistry and bioactivity of Flos Magnoliae, a Chinese herb for rhinitis and sinusitis. Curr Med Chem. 2008;15(16):1616–1627. doi: 10.2174/092986708784911515. [DOI] [PubMed] [Google Scholar]
- 8.Yang SH, Yu CL, Yang YH, Yang YH. The immune-modulatory effects of a mixed herbal formula on dendritic cells and CD4+ T lymphocytes in the treatment of dust mite allergy asthma and perennial allergic rhinitis. J Asthma. 2016;53(4):446–451. doi: 10.3109/02770903.2015.1104692. [DOI] [PubMed] [Google Scholar]
- 9.Lim SS, Shin KH, Ban HS, et al. Effect of the essential oil from the flowers of Magnolia sieboldii on the lipopolysaccharide-induced production of nitric oxide and prostaglandin E2 by rat peritoneal macrophages. Planta Med. 2002;68(5):459–462. doi: 10.1055/s-2002-32085. [DOI] [PubMed] [Google Scholar]
- 10.Baek JA, Lee YD, Lee CB, et al. Extracts of Magnoliae flos inhibit inducible nitric oxide synthase via ERK in human respiratory epithelial cells. Nitric Oxide. 2009;20(2):122–128. doi: 10.1016/j.niox.2008.10.003. [DOI] [PubMed] [Google Scholar]
- 11.Carnesecchi S, Bras-Goncalves R, Bradaia A, et al. Geraniol, a component of plant essential oils, modulates DNA synthesis and potentiates 5-fluorouracil efficacy on human colon tumor xenografts. Cancer Lett. 2004;215(1):53–59. doi: 10.1016/j.canlet.2004.06.019. [DOI] [PubMed] [Google Scholar]
- 12.Chirumbolo S, Bjorklund G. The antinociceptive activity of geraniol. Basic Clin Pharmacol Toxicol. 2017;120(2):105–107. doi: 10.1111/bcpt.12683. [DOI] [PubMed] [Google Scholar]
- 13.Sharma Y, Khan LA, Manzoor N. Anti-Candida activity of geraniol involves disruption of cell membrane integrity and function. J Mycol Med. 2016;26(3):244–254. doi: 10.1016/j.mycmed.2016.04.004. [DOI] [PubMed] [Google Scholar]
- 14.Nam SY, Kim MH, Seo Y, et al. The (2′S,7′S)-O-(2-methylbutanoyl)-columbianetin as a novel allergic rhinitis-control agent. Life Sci. 2014;98(2):103–112. doi: 10.1016/j.lfs.2014.01.003. [DOI] [PubMed] [Google Scholar]
- 15.Cardozo MT, de Conti A, Ong TP, et al. Chemopreventive effects of beta-ionone and geraniol during rat hepatocarcinogenesis promotion: distinct actions on cell proliferation, apoptosis, HMGCoA reductase, and RhoA. J Nutr Biochem. 2011;22(2):130–135. doi: 10.1016/j.jnutbio.2009.12.007. [DOI] [PubMed] [Google Scholar]
- 16.Deng XY, Xue JS, Li HY, et al. Geraniol produces antidepressant-like effects in a chronic unpredictable mild stress mice model. Physiol Behav. 2015;152(pt A):264–271. doi: 10.1016/j.physbeh.2015.10.008. [DOI] [PubMed] [Google Scholar]
- 17.Bocsan CI, Bujor AI, Miron N, Vesa SC, Deleanu D, Buzoianu AD. In vivo anti-inflammatory effect of H1 antihistamines in allergic rhinitis: a randomized clinical trial. Balkan Med J. 2015;32(4):352–358. doi: 10.5152/balkanmedj.2015.15884. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Pawankar R, Mori S, Ozu C, Kimura S. Overview on the pathomechanisms of allergic rhinitis. Asia Pac Allergy. 2011;1(3):157–167. doi: 10.5415/apallergy.2011.1.3.157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Jia XJ, Li X, Wang F, Liu HQ, Zhang DJ, Chen Y. Berbamine exerts anti-inflammatory effects via inhibition of NF-kappaB and MAPK signaling pathways. Cell Physiol Biochem. 2017;41(6):2307–2318. doi: 10.1159/000475650. [DOI] [PubMed] [Google Scholar]