

Abstract
Background
Human perception of bitter substances is partially genetically determined. Previously we discovered a single nucleotide polymorphism (SNP) within the cluster of bitter taste receptor genes on chromosome 12 that accounts for 5.8% of the variance in the perceived intensity rating of quinine, and we strengthened the classic association between TAS2R38 genotype and the bitterness of propylthiouracil (PROP). Here we performed a genome-wide association study (GWAS) using a 40% larger sample (n = 1999) together with a bivariate approach to detect previously unidentified common variants with small effects on bitter perception.
Results
We identified two signals, both with small effects (< 2%), within the bitter taste receptor clusters on chromosomes 7 and 12, which influence the perceived bitterness of denatonium benzoate and sucrose octaacetate respectively. We also provided the first independent replication for an association of caffeine bitterness on chromosome 12. Furthermore, we provided evidence for pleiotropic effects on quinine, caffeine, sucrose octaacetate and denatonium benzoate for the three SNPs on chromosome 12 and the functional importance of the SNPs for denatonium benzoate bitterness.
Conclusions
These findings provide new insights into the genetic architecture of bitter taste and offer a useful starting point for determining the biological pathways linking perception of bitter substances.
Electronic supplementary material
The online version of this article (10.1186/s12864-018-5058-2) contains supplementary material, which is available to authorized users.
Keywords: Human, Taste, Perception, Bitter, Receptor, GWAS
Background
Bitterness is a taste sensation that arises when particular chemicals come into contact with receptors in specialized cells on the human tongue [1–3]. But not everyone perceives the same bitterness for a given stimulus; this individual variation is partially genetically determined and can affect food perception, preferences and intake [4–6]. Genetic effects for bitter taste perception, which are estimated by twin studies, range from 36 to 73% [7–9], with most of the known variation arising from inborn variation in the bitter receptor gene family (T2R) [10–12]. These bitter receptors are found in tissues beyond the tongue and oral cavity, including the airways, gut, thyroid, and brain [13] where they may function as toxin detectors or early-stage sentinel systems. Bitter taste responses may reflect how well the receptors detect ligands in other tissues [14]. Historically, the ability to taste one well-studied bitter compound, phenylthiocarbamide (PTC), has been related to many diseases [15]; more recently and more specifically, variation in the PTC taste receptor is shown to be involved in the immune system [16] and to predict surgical outcome for severe rhinosinusitis [17]. Thus, together with the better-known effects on food intake and nutrition, bitter taste perception is of increasing importance to the medical field.
Given the rising importance of taste genetics, studies have focused on determining the underlying genetic variation that leads to individual differences in bitter perception. Our earlier genome-wide association study [12] (GWAS), which included 1457 adolescents from 626 twin families, replicated the classic association between the bitter taste receptor gene TAS2R38 and the perception of propylthiouracil (PROP; a chemical relative of PTC) and revealed a single nucleotide polymorphism (SNP) within the bitter taste receptor gene TAS2R19, accounting for 5.8% of the variance in the perception of quinine. Whereas the quinine does not activate T2R19 (the protein product of TAS2R19) in vitro [18], the SNP was later shown to form a long-range haplotype with missense variants within a nearby bitter taste receptor gene TAS2R31 [19, 20] whose encoded protein T2R31 can be activated by quinine [18]. The GWAS study, however, could neither detect loci for the other compounds tested, such as caffeine and sucrose octaacetate (SOA), that are likely to be affected by a large number of small-effect alleles nor the previously proposed but yet to be identified second locus for thiourea-containing compounds like PTC and PROP [21], including the suggested loci on chromosomes 5 [22] and 16 [23].
Drawing on studies of complex traits such as body mass index [24] and schizophrenia [25], here we increased the overall sample size by 40% and used multivariate association analysis [26] to identify common genetic variants (minor allele frequency [MAF] ≥ 5%) with small effects. Multivariate GWAS has been used to detect SNP associations that did not reach genome-wide significance in univariate analyses, such as autism spectrum disorders [27] and bone mineral density [28]. This method can detect not only pleiotropic genetic variants but also variants associated with only one of the correlated phenotypes [29]. As shown by Stephens [29], bivariate analysis increases power when there is greater separation of genotype groups (0, 1 or 2 copies of the minor allele) in two- versus one-dimensional space. In Fig. 1a and b, we provide two illustrations of when a joint analysis of two correlated traits can provide greater separation of genotypes associated with the primary trait (trait 1). The first example (a) shows the case where only one trait (trait 1 on the y-axis) is associated with the variant (non-pleiotropic), with bivariate analysis providing better separation of the genotype groups in 2-dimensional space compared with the y-axis alone. A similar boost in signal would be found in a conditional analysis, where the non-associated trait is included as a covariate, as this removes the non-associated part of the variance in the associated trait (i.e. covariance between two traits) and, therefore, enhances the association. The second example (b) shows that maximum separation can be achieved when both traits (trait 1 on the y-axis, trait 2 on the x-axis) are associated with the variant and the effect of the minor allele on the two is in opposite direction. In the case where a variant has the same effect on both correlated traits (Fig. 1c), bivariate analysis provides minimum/no increase in power. The bivariate approach is especially well-justified for bitter taste traits because, with the exception of PROP, perception of these bitter substances are highly correlated (rp = ~0.6) [30] and their genetic variances largely overlap (rg = ~0.7) [7, 9].
Fig. 1.

Illustration of three scenarios in a bivariate analysis. Each dot represents an individual, colored according to their genotype (0, 1 or 2 copies of the minor allele). In (a) trait 1 and 2 are correlated but the variant is only associated with trait 1. When considering traits 1 and 2 jointly in testing for association, there is greater separation of the genotype groups for trait 1 in the two-dimensional space compared with the y-axis alone. For example, the blue and green dots would largely overlap in the one-dimensional space along the y-axis. In (b) the minor allele has opposite effects on traits 1 and 2 - increasing trait 1 and decreasing trait 2. The three genotype groups are better separated in the two-dimensional space than for either trait individually. In (c) the minor allele has a similar effect on traits 1 and 2 - increasing both traits. Separation of the three genotype groups in two-dimensional space is no greater than along the y-axis alone. The figures and text are adapted from Fig. 1 in Stephens (2013) [29]
Here we aimed to identify common genetic variants with small effects (i.e. 1 – 5%) on the perception of bitterness, building on our previous GWAS [12], which was under-powered to detect common genetic variants with small effects. We performed univariate GWAS for the perceived intensity of 5 bitter substances (PROP, quinine, caffeine, SOA, and denatonium benzoate [DB]) using our expanded sample, including 1999 individuals from 929 twin families. As these phenotypes were collected from the same individuals, to boost power we ran a series bivariate GWAS (6 in total) for the correlated phenotypes of quinine, caffeine, SOA and DB [9]. We looked for evidence of pleiotropy for each identified variant. When there was little evidence for pleiotropy, we tested the SNP association to the primary trait conditional on the second. For variants in linkage disequilibrium, we used bidirectional conditional analysis (i.e. including the genotype of one SNP as a covariate at a time to test the association with the other SNP) and plotted the SNP associations for one trait against the other. Finally, to help interpret the genotype-phenotype associations, we examined the potential function of the identified SNPs with bioinformatics tools.
Results
We confirmed two previously identified associations with large effects on PROP and quinine, provided the first independent replication of an association for caffeine, and revealed two new associations with small effects (< 2%) on SOA and DB (Table 1). In addition, we found evidence for pleiotropic effects on quinine, caffeine, SOA and DB.
Table 1.
Genetic variants associated with human bitter taste perception
| Trait 1 | SNP | Chr:Position | A1/A2 | MAF | β | SE | r2 | P | Trait 2 | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Quinine | Caffeine | SOA | DB | |||||||||
| P_bivariate | ||||||||||||
| Quinine | rs10772420 | 12:11174276 | G/A | 0.469 | -0.337 | 0.034 | 5.67% | 7.8e-23* | - | 4.8e-65* | 1.8e-24* | 6.4e-26* |
| Caffeine | rs2597979 † | 12:11189966 | G/C | 0.163 | 0.264 | 0.048 | 1.91% | 4.2e-8 | 8.4e-24* | - | 2.8e-10* | 4.5e-11* |
| SOA | rs67487380 | 12:11194384 | A/G | 0.275 | -0.202 | 0.040 | 1.63% | 3.8e-7 | 5.4e-13* | 4.5e-8 | - | 2.4e-6 |
| DB | rs10261515 | 7:141398707 | G/A | 0.491 | -0.136 | 0.037 | 0.93% | 2.5e-4 | 3.1e-8 | 4.0e-6 | 5.6e-4 | - |
| PROP solution | rs10246939 | 7:141672604 | C/T | 0.443 | 0.968 | 0.028 | 46.20% | 2.8e-199* | ||||
| PROP paper | rs10246939 | 7:141672604 | C/T | 0.441 | 0.534 | 0.032 | 14.08%a | 5.4e-59* | ||||
| PROP paper | rs6761655 ‡ | 2:218218646 | G/A | 0.186 | -0.246 | 0.044 | 1.83% | 2.7e-8 | ||||
We report the top SNP from the peak association. SNPs that were not identified in our previous GWAS are shown in italics. Allele frequency and effect sizes are reported with reference to allele A1. Base-pair position is based on GRCh37; A1/A2, minor/major allele; MAF, minor allele frequency; β, the effect size; SE, standard error of the β; r2, percent variance of the trait accounted for by the SNP; P, P-value from the univariate association analysis of trait 1; P_bivariate, P-value from the bivariate association analysis of traits 1 and 2; SOA, sucrose octaacetate; DB, denatonium benzoate; bold, P < genome-wide significance threshold of 5.0e-8; *, P < corrected significance threshold of 1.0e-8; †, an independent replication; ‡, no evidence of replication. See Additional files for the full list of SNPs (Additional file 1: Table S1, Additional file 2: Table S2, Additional file 3: Table S3, Additional file 4: Table S4, Additional file 5: Table S5, Additional file 6: Table S6, Additional file 7: Table S7)
ars10246939 accounted only a third of the variance in PROP paper compared to PROP solution. This was partly due to the lower heritability of PROP paper (h2 = 0.40) compared to PROP solution (h2 = 0.71, Additional file 8: Table S8)
Confirmation of the locus on chromosome 12 influencing quinine and pleiotropic effects
The peak association for quinine was a missense variant within the bitter taste receptor gene TAS2R19 on chromosome 12 (rs10772420, Figs. 2a and 3). As expected, with the increase in sample size the association was stronger (P = 7.8e-23) than that found in our initial GWAS (P = 1.8-e15) [12], and the peak SNP explained almost the same amount of variance (5.67%). Missense variants within TAS2R31, previously reported to form a haplotype with rs10772420 [20] and associate with the bitterness of quinine [19], were all highly correlated with rs10772420 (r2 ⩾ 0.97) in the present sample and showed strong associations with the perception of quinine (P = 9.4e-22 for rs10845295; P = 1.8e-22 for rs10845293; P = 9.4e-22 for rs10772423).
Fig. 2.

Common variants associated with the perception of (a) quinine, (b) caffeine, (c) SOA, (d) DB. Manhattan plots display the association P-value for each SNP in the genome (displayed as –log10 of the P-value). The red line indicates the genome-wide significance threshold of P = 5.0e-8. rs10772420 (labelled in red), rs2597979 (labelled in green), and rs67487380 (labelled in blue) are the most significant SNP within a putative or associated locus for quinine, caffeine, and sucrose octaacetate, respectively. rs10261515 is labelled in (d) because it reaches genome-wide significance in the bivariate analysis (Table 1 and Fig. 5)
Fig. 3.

Regional association plots for the perception of quinine, caffeine and SOA on chromosome 12 between 10900000 and 11400000 base pairs with gene model below. Plots are zoomed to highlight the genomic region that likely harbors the causal variant. Color scale for the linkage disequilibrium with the top SNPs (i.e. rs10772420 for quinine, rs2597979 for caffeine and rs67487380 for SOA) and their correlations are shown in the top left of the figure. Physical locations for the three SNPs are indicated with colored dashed lines (i.e. red for rs10772420, green for rs2597979 and blue for rs67487380) across the figure. *The dot representing the association between quinine and rs2597979 in the top panel is light blue (r2rs10772420-rs2597979 = 0.24) and hidden behind the green dot
In the bivariate analysis, which included caffeine, there was a further increase in the strength of the association (P = 4.8e-65, Table 1). This was due to the nominal association of caffeine with rs10772420 (P = 2.5e-3; Figs. 2b and 3) and the effect of the minor allele being in the opposite direction to quinine (i.e. decrease in caffeine versus increase in quinine perception), which provided greater separation of the rs10772420 genotypes in two-dimensional space (as illustrated in Fig. 1b). A much smaller increase in the quinine signal was found in the bivariate analysis with SOA (P = 1.8e-24) and DB (P = 6.4e-26). Both compounds (SOA: P = 1.0e-4; DB: P = 2.8e-3) were nominally associated with rs10772420 (Fig. 2c and d), but the effect of the minor allele was in the same direction as that for quinine, resulting in little/no further separation of the genotypes in two-dimensional space (as illustrated in Fig. 1c). Notably the size and direction of the effect of rs10772420 on the four bitter substances varied (Additional file 9: Figure S1; Additional file 10: Table S9): the strongest effect was on quinine (β = -0.337; 5.67% of the variance or 12.32% of the genetic variance), with a smaller fraction of the variance being explained for caffeine (β = 0.107; 0.57/1.24% of the total/genetic variance), SOA (β = -0.137; 0.94/2.04% of the total/genetic variance) and DB (β = -0.106; 0.56/1.22% of the total/genetic variance). In Fig. 4 we show that variants with the largest effect on quinine – a cluster of 263 SNPs – were also associated with SOA, caffeine and DB, and that this cluster was separate to the top SNPs for SOA (a cluster of 167 SNPs) and caffeine (a cluster of 116 SNPs).
Fig. 4.

Top SNP associations on chromosome 12 for perceived intensity of quinine, SOA, caffeine and DB. The red, blue and green clusters represent the top SNP associations with quinine, SOA and caffeine respectively. The top SNPs for these three bitter compounds are clustered separately from one another, even though the lead SNPs (rs10772420 for quinine; rs2597979 for caffeine; rs67487380 for SOA) of each cluster are correlated (r2rs10772420-rs2597979 = 0.24; r2rs10772420-rs67487380 = 0.43; r2rs2597979-rs67487380 = 0.08). The top SNPs for DB in this genomic region overlap with the tops SNPs for SOA, but the strengths of the associations with DB are weaker. In addition, there is evidence of pleiotropy. The red cluster is strongly associated with quinine, and more weakly associated with caffeine, SOA and DB; the blue cluster is associated with quinine, SOA and DB; the green cluster is associated with quinine and caffeine. A total of 1035 SNPs on chromosome 12 between 10950000 and 11350000 base pairs are plotted here
Independent replication of a SNP association on chromosome 12 for caffeine
For caffeine perception, we identified a peak association on chromosome 12 (rs2597979, P = 4.2e-8; Fig. 2b), which accounted for a maximum trait variance of 1.91%. This SNP was in high linkage disequilibrium with that identified in a previous GWAS for caffeine detection threshold [11] (r2 = 0.84 with rs2708377), and therefore we provide the first independent replication for this association. Further support was provided by our bivariate caffeine-quinine analysis (P = 8.4e-24). The enhancement in signal due to quinine also being associated with rs2597979 (P = 4.3e-3), with the effect in the opposite direction to caffeine (Additional file 9: Figure S1). Since the lead SNPs for caffeine (rs2597979) and quinine (rs10772420) were weakly correlated (r2 = 0.24), we tested whether the associations could be driven by the same SNP using conditional analysis, where each of the genotypes are included as a covariate. The caffeine-rs2597979 association remained (P = 4.4e-6; Table 2) after conditioning on the lead SNP for quinine, whereas the caffeine-rs10772420 association disappeared (P = 0.38) after conditioning on rs2597979, indicating that the caffeine-rs2597979 association was not driven by rs10772420. For quinine, the results of the conditional analysis were less clear. While the quinine-rs10772420 association remained highly significant after conditioning on the lead SNP for caffeine (P = 3.0e-19), a weak quinine-rs2597979 association remained after conditioning on rs10772420 (P = 0.044). Figure 4 shows that the top caffeine SNPs are weakly associated with quinine and largely independent from the top quinine SNPs.
Table 2.
Conditional analyses of correlated SNPs on chromosome 12 associated with the perception of quinine, caffeine and sucrose octaacetate (SOA)
| Trait | SNP | Chr:Position | Association (P-value) |
Association conditional on correlated SNP (P-value) | ||
|---|---|---|---|---|---|---|
| rs10772420 | rs2597979 | rs67487380 | ||||
| Quinine | rs10772420 | 12:11174276 | 7.8e-23 | - | 3.0e-19 | 1.5e-10 |
| rs2597979 | 12:11189966 | 4.3e-3 | 4.4e-2 | - | - | |
| rs67487380 | 12:11194384 | 1.5e-13 | 0.12 | - | - | |
| Caffeine | rs10772420 | 12:11174276 | 2.5e-3 | - | 0.38 | - |
| rs2597979 | 12:11189966 | 4.2e-8 | 4.4e-6 | - | 9.7e-8 | |
| rs67487380 | 12:11194384 | 0.11 | - | 0.47 | - | |
| SOA | rs10772420 | 12:11174276 | 1.0e-4 | - | - | 0.47 |
| rs2597979 | 12:11189966 | 0.38 | - | - | 0.63 | |
| rs67487380 | 12:11194384 | 3.8e-7 | 7.6e-4 | 5.3e-7 | - | |
r2 = 0.24 between rs10772420 and rs2597979; r2 = 0.43 between rs10772420 and rs67487380; r2 = 0.08 between rs2597979 and rs67487380. SNP in bold in the second column are index SNPs for the corresponding traits from Table 1. Base-pair position is based on GRCh37
In contrast to quinine, we found little evidence for an association of either SOA or DB with rs2597979 (SOA: P = 0.38; DB: P = 0.62). The small enhancement in the caffeine-rs2597979 association found in the bivariate analysis (caffeine and SOA: P = 2.8e-10; caffeine and DB: P = 4.5e-11) was likely due to the phenotypic correlation between the traits. This was supported by the enhancement in the caffeine-rs2597979 association when the intensity ratings for SOA (P = 5.9e-11) or DB (P = 7.9e-12) were included as a covariate as this removed the covariance which was not associated with rs2597979. Figure 4 shows that the caffeine-associated SNPs are largely independent from SOA/DB-associated SNPs in this genomic region of chromosome 12.
Putative novel associations identified in bivariate analyses influencing SOA and DB
The strongest association for SOA was found on chromosome 12 (rs67487380, P = 3.8e-7; Figs. 2c and 3). This SNP was also associated with quinine (P = 1.5e-13; Table 2, Fig. 2a) and DB (P = 8.5e-4), with the size and direction of the effect being similar to that for SOA (Additional file 9: Figure S1), so that the stronger signal found in the bivariate SOA-quinine analysis (P = 5.4e-13; Table 1) was likely due to quinine. Even so, we found that the SOA-rs67487380 association remained when we conditioned on the lead SNP for quinine (P = 7.6e-4, Table 2), which is moderately correlated with rs67487380 (r2 = 0.43), whereas the SOA-rs10772420 association was lost (P = 0.47) when rs67487380 was included as a covariate. Similarly, for quinine, the rs10772420 association remained after conditioning on the lead SOA SNP (P = 1.5e-10), but the quinine-rs67487380 association disappeared (P = 0.12, Table 2), after conditioning on the lead quinine SNP. These conditional analysis results indicated that each of lead SNPs for SOA and quinine represents the main signal for its corresponding taste. Figure 4 clearly shows that the top SNPs for SOA and quinine are clustered separately from each other, whereas the top SNPs for DB in the genomic region on chromosome 12 largely overlap with the top SNPs for SOA.
In contrast to quinine and DB, caffeine was not associated with the lead SOA SNP (P = 0.11; Table 2). A small enhancement in the association in the bivariate SOA-caffeine analysis (P = 4.5e-8) was largely due to the phenotypic correlation between SOA and caffeine. Further, the SOA-rs67487380 association remained after conditioning on the intensity rating for caffeine (P = 1.0e-8), indicating that the covariance between SOA and caffeine was not due to this SNP. Figure 4 shows that the top SNPs for SOA and caffeine are largely separated and this is because their lead SNPs are only subtly correlated (r2 = 0.08 between rs67487380 and rs2597979).
For DB, while all SNP associations had a P-value > 1.0e-6 (Fig. 2d), one association on chromosome 7 (rs10261515, P = 2.5e-4) became stronger in the bivariate DB-quinine analysis (P = 3.1e-8, Table 1, Fig. 5). The bivariate signal was mainly driven by the SNP association with DB, as there was no evidence for an association between quinine and rs10261515 (P = 0.15), and the strength of the SNP association with DB increased after conditioning on the intensity score for quinine (P-value changed from 2.5e-4 to 1.9e-8). There was no evidence that this DB-SNP was associated with caffeine (P = 0.81) or SOA (P = 0.15), and little evidence of a signal enhancement in either the DB-caffeine (P = 4.0e-6) or DB-SOA (P = 5.6e-4) bivariate analyses (Table 1).
Fig. 5.
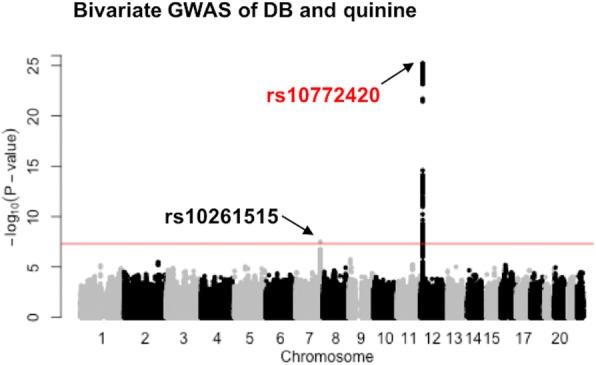
Bivariate GWAS showing a common variant on chromosome 7 associated with the perception of DB. The signal (rs10261515) on chromosome 7 is driven by DB (P = 2.5e-4 in the univariate analysis) not quinine (P = 0.15). The signal on chromosome 12 is mainly due to the association of rs10772420 with quinine rather than DB as shown in Fig. 2a and d. The red line indicates the genome-wide significance threshold of P = 5.0e-8
The SNP rs10261515 is located within KIAA1147 on chromosome 7, nearby three bitter taste receptor genes TAS2R3, TAS2R4 and TAS2R5 (Fig. 6), and is 274 kb upstream of the PROP-associated SNP rs10246939, with which it is weakly correlated (r2 = 0.23; Fig. 6). When we conditioned on the lead SNP for PROP, the DB-rs10261515 association remained (P = 9.0e-4), including after the additional adjustment for the quinine score (P = 1.7e-5).
Fig. 6.

Regional association plot for the perception of DB on chromosome 7 between 141200000 and 141700000 base pairs with gene model below. The top SNP for PROP (rs10246939) is also labelled due to its correlation with the top SNP rs10261515 (r2rs10261515-rs10246939 = 0.23)
Confirmation of previously identified locus on chromosome 7 influencing PROP
The peak association for PROP was the well-known missense variant rs10246939 within the bitter taste receptor gene TAS2R38 on chromosome 7 (Table 1, Additional file 11: Figure S2), confirming our previous findings [12]. However, we could not detect a signal on chromosomes 5 or 16 (Additional file 11: Figure S2) and all SNP associations had a P-value > 1.0e-5 on chromosomes 5p15 or 6, where previously suggested loci for PTC taste locate [22, 23].
For PROP paper, we identified a secondary locus within the DIRC3 gene on chromosome 2 (rs6761655 and its completely correlated SNP rs6736242 [r2 = 1.0], P = 2.7e-8, Additional file 11: Figure S2b). This SNP accounted for a maximum trait variance of 1.83% in PROP paper and showed a weaker but nominally significant association with the perception of PROP solution (P = 7.4e-4). We note that this signal was present in our previous GWAS [12] (Additional file 12: Figure S3), but was less obvious (i.e. not a solid peak – 4.4 million SNPs here vs 2.3million SNPs in our earlier GWAS) and therefore was not reported. However, we found no evidence for this association with PROP perception in one previously reported GWAS of 225 Brazilians [10], as well as two unpublished GWAS, one of ~500 individuals from the Silk Road population and one of ~2500 Italians (Additional file 13: Table S10).
Functional annotation of the identified SNPs
We performed functional analysis (i.e. the SNP effect on gene expression) for five of the six SNPs in Table 1 using the bioinformatics tool HaploReg v4.1 [31]. We did not include rs6761655 here due to lack of replication in the independent datasets. We also searched for bitter taste receptors that have been shown to respond to these bitter substances in human cell-based functional studies [18, 32]. The key results are presented in Table 3 and a summary of the functional analysis can be found in Additional file 14: Table S11.
Table 3.
Bioinformatics and cell-based functional studies of the genetic variants associated with bitter taste perception
| Trait | Index SNP | GENCODE genes | Non-synonymous SNPs in LD (r2 ⩾ 0.8) with index SNP | eQTLa | Cell-based functional analysisb |
|---|---|---|---|---|---|
| Quinine | rs10772420 | TAS2R19 | rs10772420 in TAS2R19; rs10845295, rs10845293 and rs10772423 in TAS2R31c | TAS2R10, TAS2R14, TAS2R19, TAS2R20, TAS2R31, TAS2R43, TAS2R50, TAS2R64P | T2R4, T2R7, T2R10, T2R14, T2R31, T2R39, T2R40, T2R43, T2R46 [18] |
| Caffeine | rs2597979 | PRR4, TAS2R31 | rs10743938 in TAS2R31 | TAS2R14, TAS2R15, TAS2R20, TAS2R31, TAS2R43, TAS2R45, TAS2R64P | T2R7, T2R10, T2R14, T2R43, T2R46 [18] |
| SOA | rs67487380 | PRR4 | TAS2R10, TAS2R12, TAS2R14, TAS2R15, TAS2R19, TAS2R20, TAS2R31, TAS2R43, TAS2R46, TAS2R64P | T2R46 [32] | |
| DB | rs10261515 | KIAA1147 | TAS2R4, TAS2R5 | T2R4, T2R8, T2R10, T2R13, T2R30, T2R39, T2R43, T2R46 [18] | |
| PROP | rs10246939 | MGAM, TAS2R38 | rs713598, rs1726866 and rs10246939 in TAS2R38 | TAS2R5, TAS2R38 | T2R38 [18] |
aExpression of these bitter taste receptor genes is associated with the genotype of the index SNP and/or its correlated SNPs (r2⩾0.8)
bBitter taste receptors shown to respond to bitter substances in cell-based functional analysis using human embryotic kidney cells [18, 32]
cThe 3 SNPs, previously reported to form a haplotype with rs10772420 [19, 20], in TAS2R31 did not show up in the HaploReg analysis when using the inclusion criteria of r2 ⩾ 0.8. They were included due to their strong correlations with rs10772420 (r2 ⩾ 0.97) in the present sample
The top SNP for quinine (rs10772420) and its correlated SNPs are missense variants within TAS2R19 and TAS2R31. The caffeine-associated SNP (rs2597979) is highly correlated with a missense variant rs10743938 (r2 = 0.92) within TAS2R31. This SNP has two possible allele changes of T>A and T>G, leading to residue changes of Leu162Met and Leu162Val respectively. In the present study, only rs10743938:T>A passed quality control and its association with caffeine had a P-value of 1.1e-7 (Additional file 2: Table S2). The top SNP for PROP (rs10246939) and its correlated SNPs are missense variants within TAS2R38.
Further, the SNPs for quinine, caffeine and SOA are common expression quantitative loci (eQTL) for five bitter taste receptor genes (TAS2R14, TAS2R20, TAS2R31, TAS2R43, TAS2R64P) on chromosome 12, and the expression of other bitter taste receptors in the same region is regulated by one or two of these three SNPs, e.g. the expression of TAS2R46 is only regulated by the SOA and quinine associated SNP rs67487380. The DB-associated SNP rs10261515 influences the expression of the bitter taste receptor genes, TAS2R4 and TAS2R5, on chromosome 7. T2R4 is more likely to be a receptor for DB because the allele (rs10261515 G allele) for weaker DB intensity rating is associated with a lower expression level of TAS2R4 and the opposite for TAS2R5. In addition, DB can activate T2R4 but T2R5 in cell-based functional analysis [18]. Results from the cell-based functional analysis do not necessarily agree with the results from the bioinformatics functional analysis. For example, the quinine-associated SNP rs10772420 is a missense variant within TAS2R19 and it regulates the gene expression of TAS2R19, but T2R19 does not respond to quinine [18]. In contrast, the association between the perception of quinine and TAS2R31 was supported by both bioinformatics and functional analyses (Table 3). We note that neither of these bioinformatics and cell-based functional analyses were based on taste tissues.
Discussion
In this study of bivariate GWAS on human taste perception, we identify two putative novel associations, including rs67487380 on chromosome 12 for SOA-elicited bitter taste and rs10261515 on chromosome 7 for DB-elicited bitter taste. In addition, we provide the first independent replication of an association on chromosome 12 for caffeine bitterness (rs2597979) and confirm our previously reported associations for quinine bitterness (rs10772420 on chromosome 12) and PROP bitterness (rs10246939 on chromosome 7). All variants are located within the bitter taste receptor clusters on chromosomes 7 and 12, highlighting the importance of these two regions in the genetics of bitter taste. Further, we show evidence of pleiotropy for those variants on chromosome 12 and the functional importance of the DB-associated SNP.
This is the first GWAS study to identify a SNP (rs67487380 on chromosome 12) association with human perception of SOA. In mice, a major locus for SOA perception (soa) was reported in the early 1990s [33]. Interestingly, the mouse soa locus also affects the perception of other bitter substances, including quinine, DB, PROP, but not caffeine [34, 35]. Here we provide evidence that rs67487380 is also associated with the perception of quinine and DB, but not caffeine or PROP (P > 0.05). SOA activates human T2R46 but no other T2Rs in heterologous expression assays [32]. It is possible that rs67487380 regulates the perception of SOA through its effect on mRNA expression because the G allele for weaker SOA intensity is also associated with a lower expression level of TAS2R46. Nevertheless, rs67487380 could still be a proxy for true causal variants.
The finding of the novel association between DB and the SNP rs10261515 suggests that there may be a second locus on chromosome 7 that affects human bitter taste perception (the first is the locus within TAS2R38 for PROP). Heterologous expression studies using human embryotic kidney (HEK) cells transfected with TAS2Rs have shown that DB activates T2R4 but no other bitter taste receptors in this region (e.g. T2R3, T2R5 and T2R38) [18]. In addition, the human T2R4 is the ortholog of mouse T2R8, which also responds to DB [1]. Our functional annotation results provide further support for T2R4 as a DB bitter taste receptor, since the allele (rs10261515 G allele) for a lower perceived intensity of DB is associated with lower expression level of TAS2R4 mRNA.
The SNP association for caffeine perception replicated a previous GWAS of 608 Brazilian adults [11]. In that study the lead SNP accounted for 8.9% of the variance in caffeine sensitivity, compared with our estimate of 1.9%. Similarly, the Brazilian study accounted for 23.2% of the variance of quinine taste with genetic mutations, which is four times the effect estimated here. This difference in effect sizes is likely due to two main factors. First, the taste scores in the Brazilian sample were corrected for overall-taste-sensitivity (an average score of the perception of sweet, umami, sour, salty and bitter compounds), which removed ~30% of the variance in the perception of caffeine and quinine. Without correction, rs10772420 accounted for 13.2% of the variance in quinine, and the caffeine association was not detected due to low power. Second, the Brazilian study used a detection threshold approach, which measures overall oral sensitivity, compared with our measure of bitter taste intensity. Regardless, both studies identified the same variants for caffeine, quinine as well as PROP, indicating that these are likely to be valid associations among human bitter taste perception and these T2R-rich regions of chromosomes 7 and 12.
The functional annotation of the caffeine-associated SNP showed that the highly correlated SNP (rs10743938) is a missense mutation that could affect the function of T2R31. This is the first evidence linking this bitter taste receptor to the perception of caffeine, while genetic variants in TAS2R31 have been shown to affect the perception of quinine [19] (confirmed in the present study), acesulfame potassium and saccharin [36] (the latter two are non-nutritive sweeteners with bitter aftertaste). Prior cell-based functional studies [18] reported that caffeine does not activate T2R31 in heterologous expression assays; rather, it activates T2R7, -10, -14, -43, and -46, and that the summed expression level of these activated T2Rs increases with the perceived intensity of caffeine [37]. However, comparing results from bioinformatics and cell-based analyses can be limited by two major factors. Here, we report associations for the index (lead) SNP with the lowest P-value, but since this SNP is in a linkage disequilibrium block, the association could be driven by any variant within the block. Second, these cell-based functional assays [18] were conducted in heterologous systems (i.e. HEK cells transfected with TAS2Rs), which may not always recapitulate human sensory experience faithfully [38]. We observed a similar difference for quinine, with the lead quinine-associated SNP rs10772420 constituting a missense mutation in TAS2R19. Yet T2R19 does not respond to quinine in functional expression assays [18]. Instead, rs10772420 is more likely to be a proxy of missense variants within TAS2R31 [19, 20], whose encoded protein T2R31 responds to quinine [18]. Therefore, a better method to identify causal SNPs for the foreseeable future is to tightly integrate genetic-perceptual association results with those of taste receptor cell-based assays using human taste tissues, such as taste buds or cultured human taste cells [39].
This study provides the first evidence for antagonistic genetic pleiotropy in bitter taste. The two SNPs rs10772420 and rs2597979 have opposite effects on the perceived intensity of quinine and caffeine (Additional file 9: Figure S1, Additional file 10: Table S9) and this largely enhances the strengths of their associations (P-value) in the bivariate analysis (Fig. 1b). As bitter-tasting substances (e.g. caffeine) can have both beneficial and detrimental effects, the antagonistic pleiotropy may be an evolutionary consequence that avoids over and under consumption.
The top SNPs for quinine, caffeine, and SOA were correlated (r2 = 0.08 – 0.43) and each could have various effects on one another. These correlations are due to the linkage disequilibrium between polymorphisms within bitter taste receptor genes on chromosome 12, which results in common haplotypes for nearby genes and long-range haplotypes for more distant ones [20, 36]. Previous studies have revealed a complex bitter substance – receptor relationship, with one bitter compound activating multiple T2Rs and one T2R responding to multiple bitter substances [18, 32, 40]. Taken together, it is likely that the perception of a bitter taste can be mediated by multiple T2Rs, and SNPs identified in the present study could represent haplotypes that regulate several T2Rs together. We have attempted to illustrate this in Fig. 7 by taking the perception of quinine and caffeine as an example. The lead SNP for quinine (rs10772420) is correlated with several SNPs (the regional association plot in Fig. 3) that regulate the T2Rs for quinine (cell-based functional analysis results in Table 3). Also, the lead SNP for caffeine (rs2597979) is correlated with SNPs that regulate T2Rs for caffeine (Table 3). In addition, the common T2Rs for the two tastes are regulated by SNPs that are in linkage disequilibrium with the two lead SNPs. We note that the real regulatory network can be more complex than this, such that one T2R can be regulated by multiple SNPs. Whereas we used conditional analysis (Table 2) and plotted the SNP associations against the three tastes (Fig. 4) to show that each of the lead SNPs represents the main signal in the linkage disequilibrium block, the clusters of nearby bitter receptors and many variants in high linkage disequilibrium create challenges in separating causal from non-causal variants.
Fig. 7.
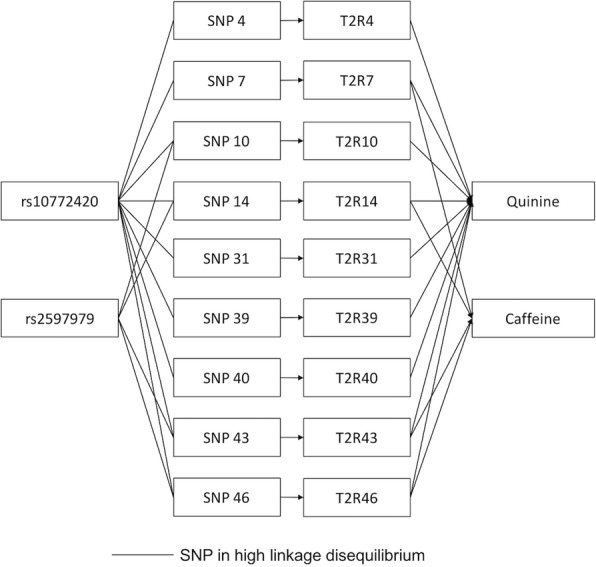
Potential model of the SNP regulation of human bitter taste perception. In vitro analysis has shown that quinine can be detected by bitter taste receptors T2R4, -7, -10, -14, -31, -39, -40, -43, and -46, and caffeine can be detected by the T2R7, -14, -43, and -46 (as summarized in Table 3), which overlap the T2Rs for quinine. Here we assume that each T2R is regulated by a major SNP with the corresponding number (e.g. SNP 4 for T2R4) and the top SNP for each taste is in linkage disequilibrium with the major SNPs for T2Rs that can detect the taste. Therefore, rs10772420 is associated with the perception of quinine via correlated SNPs, SNP 4, -7, -10, -14, -31, -39, -40, -43, and -46, and rs2597979 is associated with the perception of caffeine via correlated SNPs, SNP 7, -14, -43, and -46
Perceptual studies of bitter taste also have reported that individual differences in perceived bitterness from multiple compounds show positive correlations. Most relevant to the present work, past studies demonstrated a strong correlation of perceived bitter taste intensities among DB, SOA, and quinine [30]. This observation harkens to that reported in the present study for rs67487380 on chromosome 12. Furthermore, individual differences in bitterness from SOA, caffeine and quinine were also observed, suggesting a linkage between SOA receptor variants and caffeine receptor variants [30]. This too reflects associations observed in the present data set. Perhaps, a linkage disequilibrium block accounts both for the genetic architecture as well as the bitterness perception associations.
Prior work using pedigree segregation analysis has proposed that the perception of PTC (a structurally related chemical to PROP) is modulated by a 2-locus model [21], but the location of a second locus has been unclear for nearly 30 years. Here we found neither support for an association with TAS2R1 on chromosome 5 nor an association on chromosome 16, both of which was suggested by prior family-based linkage studies [22, 23], but identified a putative secondary locus within the DIRC3 gene on chromosome 2, which accounted for an additional 1.83% of the variance (4.58% of the genetic variance) in the perception of PROP paper. While we found no evidence for replication using three independent datasets – from one published study (i.e. the Brazilian sample) and two unpublished (the Silk Road and the Italian samples), we note that there are considerable differences across studies (e.g. sample age [all other studies used adults], ethnicity, and delivery method [the Brazilian study used PROP solution]), which may have influenced our ability to replicate their findings. Ideally, we need to test for this association using the same methods and materials (i.e. the perceived intensity of saturated PROP paper measured from adolescents with European ancestry), but at this stage the signal does not appear to be sufficiently robust to be detected with alternative methods.
The strengths of the present study include the use of the largest sample to date, and the collection of multiple taste phenotypes from the same individuals, which increases the statistical power via bivariate association analysis. We show that the association signals (P-value) for quinine and caffeine (rs10772420 and rs2597979 respectively) were stronger in the bivariate compared with the univariate analysis, but the estimated effect size remains the same. The signal boosts in these already established associations serve as a proof of principle for using bivariate GWAS. We also show that, through the discovery of the association of DB, a signal can be enhanced when only one of two correlated traits is associated. This is useful for identifying non-pleiotropic SNPs for correlated phenotypes. We used multiple levels of analysis (conditional on genotype and phenotype) as well as cluster plots to disentangle the pleiotropic nature of these SNPs with bitter tastes and provide additional support for the signals identified in the bivariate analyses. We attempted to obtain data to replicate every novel association. However, we were unable to test the association for SOA and DB because we are currently the only group in the world that has collected these two bitter taste phenotypes along with genomic data. Given the enhancement in the known signals for both quinine and caffeine in the bivariate analyses, together with the post-hoc bioinformatics analyses, as well as prior functional analyses, we believe the SOA and DB hits are unlikely to be false positives. Further, findings from multivariate GWAS of other phenotypes, e.g. levels of plasma lipids [41], have been replicated in independent studies. The variants for SOA and DB account for less than 10% of the genetic variance (< 2% of trait variance) of their associated traits, suggesting that there are more variants with smaller effects. The remaining genetic variance could be partly due to rare variants because SNPs with an MAF smaller than 5% were excluded here and rare variants can have a large effect on complex traits [42].
Conclusion
This study reveals the influence of multiple variants on bitter taste and demonstrates the benefits of multivariate analysis over the conventional univariate GWAS. Recent advancement in the methodology of multivariate GWAS (i.e. MTAG [43]) could make multivariate analysis easier to apply because it uses individual summary level results from different studies and does not require correlated phenotypes to be collected from the same sample. Whereas our previous twin analysis provided strong evidence of pleiotropy for the perception of several bitter compounds (except for PROP), there are numerous causal models that could underlie this shared genetic etiology. Identification of specific SNPs/genes involved offers a useful starting point for determining the biological pathways linking perception of bitter substances and for delineating of the mechanisms involved. Future studies integrating bioinformatics and functional analyses using human taste tissues will provide stronger evidence in identifying true causal variants, which could assist personalized nutrition and precision medicine.
Methods
Sample
Participants were 1999 adolescent and young adult Caucasian twins and their siblings from 929 families from the Brisbane Adolescent Twin Study [44], also referred to as the Brisbane Longitudinal Twin Study (BLTS), with data collected between August 2002 and July 2014. This sample consisted of 275 monozygotic (MZ) and 544 dizygotic (DZ) twin pairs, including 155 pairs with one to two singleton siblings, and 184 unpaired individuals (mean age of 16.0 ± 2.8 years [medium 14.5 years, range 11 – 25 years]; 1075 females, 924 males). It included all participants from our previous genome-wide association study [12], plus a 40% increase in sample size.
Taste Test
The taste test battery has been described in detail elsewhere [7]. Briefly, it included duplicated presentations of five bitter (6.0 x 10-4 M PROP, 2.0 x 10-4 M SOA, 1.81 x 10-4 M quinine, 0.05 M caffeine, and 4.99 x 10-6 M DB) solutions as well as a paper strip rinsed in a saturated PROP solution (0.059M). Participants were instructed to rate their perceived intensity for each solution and the PROP paper using a general Labelled Magnitude Scale (gLMS) [45] with labels of no sensation (0 mm), barely detectable (2 mm), weak (7 mm), moderate (20 mm), strong (40 mm), very strong (61 mm), and strongest imaginable (114 mm). Mean intensity ratings from duplicate presentations for each stimulus were used in all analyses. A total of 1757 participants completed the full test battery (solutions and PROP paper) with a further 242 providing an intensity rating for the PROP paper only.
Genotyping, Genetic Imputation and Quality Control
Genotyping was performed with the Illumina 610-Quad BeadChip (n = 1457 individuals) and the HumanCoreExome-12 v1.0 BeadChip (n = 542 individuals), with approximately 700 thousand SNPs passing standard quality control filters, as outlined previously [12]. These SNPs were then phased using ShapeIT [46] and imputed using Minimac3 [47] to extend the genomic coverage to 7,035,128 SNPs using the Haplotype Reference Consortium of Caucasian European ancestry (Release 1) [48]. Individuals who were > 6 standard deviations from the principal components 1 and 2 (PC1/PC2) centroid were excluded, so our sample was of exclusively European ancestry. To ensure SNPs were imputed with high data quality, we performed post-imputation QC. SNPs with a call rate < 90%, MAF < 0.05, imputation score < 0.3, and Hardy–Weinberg equilibrium score of P < 1.0e-6 were excluded, with a total of 4,381,914 SNPs remaining.
Genome-wide Association Analysis
Univariate and bivariate GWAS were conducted using a linear mixed model implemented in the software GEMMA (Genome-wide Efficient mixed-model association) [26]. This method of analysis is commonly used to analyze data from related individuals including twins [49–51]. Covariates (fixed effects) included age, sex, a history of ear infection, all of which were shown to be associated with taste intensity ratings [9], and the first five PCs calculated from the genotypes. Individual relatedness within families (i.e. twins and siblings) and between unrelated individuals were accounted for by the covariance matrix of the random effect in the model. The covariance matrix was an empirical genetic relatedness matrix, calculated from the genome-wide genotype data and representing genetic similarity across individuals. This model adjusts for the contribution of each individual to the SNP association and corrects for inflation so related individuals, including both members from monozygotic twin pairs [52], can be analyzed together without losing power. As requested by one reviewer, we also reported the association of the top SNP associations using the sample with one member of each MZ pair removed (Additional file 15: Table S12). Bivariate analysis essentially provides a complement to univariate analysis. It can enhance the strength of a SNP association, but the estimated effect on each of the two traits remains. For non-pleiotropic SNPs identified in bivariate analysis, we tested for their associations using conditional analysis of the associated trait conditional on the non-associated trait. When two identified SNPs were correlated, to test whether they were independent signals for the corresponding traits, we performed conditional analyses, by fitting each of the SNPs as an extra covariate. Prior to analyses, intensity ratings for each stimulus were square-root transformed to obtain a more normal distribution [9] and then converted to Z-scores. A genome-wide significance threshold was defined as P < 5.0e-8. As four of the phenotypes were correlated (rp between quinine, caffeine, SOA and DB = 0.58 – 0.64) [9] the number of independent tests was estimated using a matrix spectral decomposition algorithm [53] at 4.96 and accordingly a Bonferroni-corrected threshold was defined as P < 1.0e-8. The genomic inflation factor (λ) ranged between 0.99 and 1.02 (Additional file 16: Figure S4, Additional file 17: Figure S5), which indicates that potential technical or population stratification artifacts had a negligible impact on the results. As all association analyses were performed under an additive model and all phenotypes were converted to Z-scores, variance explained by a SNP was calculated as 2 x MAF x (1 – MAF) x β2. Manhattan and Q-Q plots were created using the “fastman” package [54] in R. Regional association plots were created using Locuszoom [55].
Functional annotation of the identified SNPs
To examine the potential role of the identified SNPs, we used HaploReg v4.1 [31] for functional annotation. Briefly, it annotates all index SNPs and their correlated SNPs (r2 was set to be ⩾ 0.8 [calculated based on 1000 Genome Phase 1 European population] for this study) by their associated chromatin states (e.g. conserved regions and DNAse hypersensitivity sites) from the Roadmap epigenomics project [56] and Encode project [57] and their effects on regulatory motifs. It also reports the effect of SNPs on gene expression in multiple tissues from eQTL studies, including results from the GTEx [58] project portal. Use of functional annotation may provide more information about the putative role of a specific gene as well as developing mechanistic hypotheses of the impact of the SNP on phenotypes (e.g. variation in taste perception). More details are provided in Additional file 14: Table S11.
Additional files
Table S1. Top 100 SNPs on chromosome 12 associated with the perceived intensity of quinine. (DOCX 177 kb)
Table S2. Top 100 SNPs on chromosome 12 associated with the perceived intensity of caffeine. (DOCX 178 kb)
Table S3. Top 100 SNPs on chromosome 12 associated with the perceived intensity of sucrose octaacetate (SOA). (DOCX 181 kb)
Table S4. Top 100 SNPs on chromosome 7 associated with the perceived intensity of denatonium benzoate (DB) from the bivariate analysis of DB and quinine. P_univaraite_DB is the P-value from the univariate analysis of DB. P_univariate DB_adjQ is the P-value from the univariate analysis of DB adjusted for the quinine score. P_bivariate DB_Q is the P-value from the bivariate analysis of DB and quinine. (DOCX 218 kb)
Table S5. Top 100 SNPs on chromosome 7 associated with the perceived intensity of PROP solution. (DOCX 175 kb)
Table S6. Top 100 SNPs on chromosome 7 associated with the perceived intensity of PROP paper. (DOCX 171 kb)
Table S7. Top 100 SNPs on chromosome 2 associated with the perceived intensity of PROP paper and their associations with PROP solution. (DOCX 183 kb)
Table S8. Mean, standard deviation, and heritability estimates for the perceived intensity of bitter tastes. (DOCX 47 kb)
Figure S1. Direction and size of the effects of SNP associations on the perceived intensities of quinine, caffeine, sucrose octaacetate (SOA) and denatonium benzoate (DB). (DOCX 240 kb)
Table S9. Phenotypic and genetic variance in the perceived intensity of quinine, caffeine, sucrose octaacetate (SOA) and denatonium benzoate (DB) explained by rs10772420, rs2597979, rs67487380 and rs10261515. (DOCX 51 kb)
Figure S2. Univariate GWAS for the perception of (a) PROP solution (n = 1757) and (b) PROP paper (n = 1999). Left part are Manhattan plots displaying the association P-value for each SNP in the genome (displayed as –log10 of the P-value). The red line indicates the genome-wide significance threshold of P = 5.0e-8. Right part are regional plots ±400kb from the top SNPs on chromosome 7 for PROP solution and chromosome 2 for PROP paper with the gene model below. (DOCX 444 kb)
Figure S3. Univariate GWAS for the perception of PROP paper from our previous GWAS of 1756 Australian adolescents. A Manhattan plot displays the association P-value for each SNP in the genome (displayed as –log10 of the P-value). This figure is the Fig. 1b from our previous published paper (PMID: 20675712). (DOCX 292 kb)
Table S10. Top PROP paper-associated SNPs on chromosome 2 in the current study (n = 1999) and their associations in GWAS of (1) 225 subjects from the general population of the São Paulo metropolitan area of Brazil, (2) 466 subjects from Silk Road and (3) 2588 subjects from three Italian cohorts. (DOCX 104 kb)
Table S11. Results of the annotation conducted with HaploReg v4.1. (DOCX 119 kb)
Table S12. Genetic variants associated with human bitter taste perception using the sample with one member of each MZ pair removed (see Table 1 for associations using the full sample). (DOCX 24 kb)
Figure S4. The Q-Q plots for each of the univariate analyses. PROP: propylthiouracil. (DOCX 2545 kb)
Figure S5. The Q-Q plots for each of the bivariate analyses. SOA: sucrose octaacetate. DB: denatonium benzoate. (DOCX 2547 kb)
Acknowledgements
The authors thank Kirsten J Mascioli, Christopher Tharp, Fujiko Duke, Deborah Lee and Corrine Mansfield from the Monell Chemical Senses Center for manufacturing the taste tests; and from the QIMR Berghofer, Marlene Grace, Ann Eldridge, Natalie Garden, and Kerrie McAloney for project co-ordination, data collection, and data entry, and David Smyth for computer support. We thank Robino Antonietta from Italian Ministry of Health for looking up SNPs in their unpublished GWAS of PROP. We thank the anonymous reviewers for providing constructive comments. In particular, thanks go to twins and their families for their participation.
Funding
This work was supported by the National Institute of Health [DC02995 to PASB and DC004698 to DRR], the Australian National Health and Medical Research Council [241944 and 1031119 to NGM], and the Australian Research Council [DP1093900 and DP0664638 to NGM and MJW]. The funding bodies did not participant in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- DB
Denatonium benzoate
- eQTL
Expression quantitative trait loci
- GWAS
Genome-wide association study
- MAF
Minor allele frequency
- PROP
Propylthiouracil
- SNP
Single nucleotide polymorphism
- SOA
Sucrose octaacetate
Authors’ contributions
Conceived and designed the experiments: PASB NGM DRR MJW. Processed data: LH SDG. Analyzed data: LH PG GZ. Drafted the first version of the manuscript: LH. Provided critical feedback on the manuscript: PG GZ PASB NGM DRR MJW. All authors read and approved the final manuscript.
Ethics approval and consent to participate
This study was approved by the QIMR Berghofer Human Research Ethics Committee (HREC reference number: P193). All study procedures were performed in accordance with the World Medical Association Declaration of Helsinki ethical principles for medical research. Informed written consent was obtained from both the participants and their parents (the latter not required for those 18 years and over).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Liang-Dar Hwang, Email: d.hwang@uq.edu.au.
Puya Gharahkhani, Email: Puya.Gharahkhani@qimrberghofer.edu.au.
Paul A. S. Breslin, Email: breslin@monell.org
Scott D. Gordon, Email: Scott.Gordon@qimrberghofer.edu.au
Gu Zhu, Email: Gu.Zhu@qimrberghofer.edu.au.
Nicholas G. Martin, Email: Nick.Martin@qimrberghofer.edu.au
Danielle R. Reed, Email: reed@monell.org
Margaret J. Wright, Email: margie.wright@uq.edu.au
References
- 1.Chandrashekar J, Mueller KL, Hoon MA, Adler E, Feng L, Guo W, Zuker CS, Ryba NJ. T2Rs function as bitter taste receptors. Cell. 2000;100(6):703–711. doi: 10.1016/S0092-8674(00)80706-0. [DOI] [PubMed] [Google Scholar]
- 2.Adler E, Hoon MA, Mueller KL, Chandrashekar J, Ryba NJ, Zuker CS. A novel family of mammalian taste receptors. Cell. 2000;100(6):693–702. doi: 10.1016/S0092-8674(00)80705-9. [DOI] [PubMed] [Google Scholar]
- 3.Matsunami H, Montmayeur JP, Buck LB. A family of candidate taste receptors in human and mouse. Nature. 2000;404(6778):601–604. doi: 10.1038/35007072. [DOI] [PubMed] [Google Scholar]
- 4.Reed DR, Tanaka T, McDaniel AH. Diverse tastes: Genetics of sweet and bitter perception. Physiol Behav. 2006;88(3):215–226. doi: 10.1016/j.physbeh.2006.05.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Reed DR, Knaapila A. Genetics of taste and smell: poisons and pleasures. Prog Mol Biol Transl Sci. 2010;94:213–240. doi: 10.1016/B978-0-12-375003-7.00008-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sandell MA, Breslin PA. Variability in a taste-receptor gene determines whether we taste toxins in food. Curr Biol. 2006;16(18):R792–R794. doi: 10.1016/j.cub.2006.08.049. [DOI] [PubMed] [Google Scholar]
- 7.Hansen JL, Reed DR, Wright MJ, Martin NG, Breslin PA. Heritability and genetic covariation of sensitivity to PROP, SOA, quinine HCl, and caffeine. Chem Senses. 2006;31(5):403–413. doi: 10.1093/chemse/bjj044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Knaapila A, Hwang LD, Lysenko A, Duke FF, Fesi B, Khoshnevisan A, James RS, Wysocki CJ, Rhyu M, Tordoff MG, et al. Genetic analysis of chemosensory traits in human twins. Chem Senses. 2012;37(9):869–881. doi: 10.1093/chemse/bjs070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hwang LD, Breslin PA, Reed DR, Zhu G, Martin NG, Wright MJ. Is the Association Between Sweet and Bitter Perception due to Genetic Variation? Chem Senses. 2016. [DOI] [PMC free article] [PubMed]
- 10.Genick UK, Kutalik Z, Ledda M, Destito MC, Souza MM, Cirillo CA, Godinot N, Martin N, Morya E, Sameshima K, et al. Sensitivity of genome-wide-association signals to phenotyping strategy: the PROP-TAS2R38 taste association as a benchmark. PLoS One. 2011;6(11):e27745. doi: 10.1371/journal.pone.0027745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ledda M, Kutalik Z, Souza Destito MC, Souza MM, Cirillo CA, Zamboni A, Martin N, Morya E, Sameshima K, Beckmann JS, et al. GWAS of human bitter taste perception identifies new loci and reveals additional complexity of bitter taste genetics. Hum Mol Genet. 2014;23(1):259–267. doi: 10.1093/hmg/ddt404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Reed DR, Zhu G, Breslin PA, Duke FF, Henders AK, Campbell MJ, Montgomery GW, Medland SE, Martin NG, Wright MJ. The perception of quinine taste intensity is associated with common genetic variants in a bitter receptor cluster on chromosome 12. Hum Mol Genet. 2010;19(21):4278–4285. doi: 10.1093/hmg/ddq324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Avau B, Depoortere I. The bitter truth about bitter taste receptors: beyond sensing bitter in the oral cavity. Acta physiologica. 2016;216(4):407–420. doi: 10.1111/apha.12621. [DOI] [PubMed] [Google Scholar]
- 14.Fischer R, Griffin F. Pharmacogenetic Aspects of Gustation. Arzneimittelforschung. 1964;14:673–686. [PubMed] [Google Scholar]
- 15.Guo SW, Reed DR. The genetics of phenylthiocarbamide perception. Ann Hum Biol. 2001;28(2):111–142. doi: 10.1080/03014460151056310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lee RJ, Xiong G, Kofonow JM, Chen B, Lysenko A, Jiang P, Abraham V, Doghramji L, Adappa ND, Palmer JN, et al. T2R38 taste receptor polymorphisms underlie susceptibility to upper respiratory infection. J Clin Invest. 2012;122(11):4145–4159. doi: 10.1172/JCI64240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Adappa ND, Farquhar D, Palmer JN, Kennedy DW, Doghramji L, Morris SA, Owens D, Mansfield C, Lysenko A, Lee RJ, et al. TAS2R38 genotype predicts surgical outcome in nonpolypoid chronic rhinosinusitis. Int Forum Allergy Rhinol. 2016;6(1):25–33. doi: 10.1002/alr.21666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Meyerhof W, Batram C, Kuhn C, Brockhoff A, Chudoba E, Bufe B, Appendino G, Behrens M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem Senses. 2010;35(2):157–170. doi: 10.1093/chemse/bjp092. [DOI] [PubMed] [Google Scholar]
- 19.Hayes JE, Feeney EL, Nolden AA, McGeary JE. Quinine Bitterness and Grapefruit Liking Associate with Allelic Variants in TAS2R31. Chem Senses. 2015;40(6):437–443. doi: 10.1093/chemse/bjv027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Roudnitzky N, Behrens M, Engel A, Kohl S, Thalmann S, Hubner S, Lossow K, Wooding SP, Meyerhof W. Receptor Polymorphism and Genomic Structure Interact to Shape Bitter Taste Perception. PLoS Genet. 2015;11(9):e1005530. doi: 10.1371/journal.pgen.1005530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Olson JM, Boehnke M, Neiswanger K, Roche AF, Siervogel RM. Alternative genetic models for the inheritance of the phenylthiocarbamide taste deficiency. Genet Epidemiol. 1989;6(3):423–434. doi: 10.1002/gepi.1370060305. [DOI] [PubMed] [Google Scholar]
- 22.Reed DR, Nanthakumar E, North M, Bell C, Bartoshuk LM, Price RA. Localization of a gene for bitter-taste perception to human chromosome 5p15. Am J Hum Genet. 1999;64(5):1478–1480. doi: 10.1086/302367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Drayna D, Coon H, Kim UK, Elsner T, Cromer K, Otterud B, Baird L, Peiffer AP, Leppert M, Utah Genetic Reference P Genetic analysis of a complex trait in the Utah Genetic Reference Project: a major locus for PTC taste ability on chromosome 7q and a secondary locus on chromosome 16p. Hum Genet. 2003;112(5-6):567–572. doi: 10.1007/s00439-003-0911-y. [DOI] [PubMed] [Google Scholar]
- 24.Locke AE, Kahali B, Berndt SI, Justice AE, Pers TH, Day FR, Powell C, Vedantam S, Buchkovich ML, Yang J, et al. Genetic studies of body mass index yield new insights for obesity biology. Nature. 2015;518(7538):197–206. doi: 10.1038/nature14177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Schizophrenia Working Group of the Psychiatric Genomics C Biological insights from 108 schizophrenia-associated genetic loci. Nature. 2014;511(7510):421–427. doi: 10.1038/nature13595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Zhou X, Stephens M. Efficient multivariate linear mixed model algorithms for genome-wide association studies. Nat Methods. 2014;11(4):407–409. doi: 10.1038/nmeth.2848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tao Y, Gao H, Ackerman B, Guo W, Saffen D, Shugart YY. Evidence for contribution of common genetic variants within chromosome 8p21.2-8p21.1 to restricted and repetitive behaviors in autism spectrum disorders. BMC Genomics. 2016;17:163. doi: 10.1186/s12864-016-2475-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Saint-Pierre A, Kaufman JM, Ostertag A, Cohen-Solal M, Boland A, Toye K, Zelenika D, Lathrop M, de Vernejoul MC, Martinez M. Bivariate association analysis in selected samples: application to a GWAS of two bone mineral density phenotypes in males with high or low BMD. Eur J Hum Genet. 2011;19(6):710–716. doi: 10.1038/ejhg.2011.22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Stephens M. A unified framework for association analysis with multiple related phenotypes. PLoS One. 2013;8(7):e65245. doi: 10.1371/journal.pone.0065245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Delwiche JF, Buletic Z, Breslin PA. Covariation in individuals' sensitivities to bitter compounds: evidence supporting multiple receptor/transduction mechanisms. Percept Psychophys. 2001;63(5):761–776. doi: 10.3758/BF03194436. [DOI] [PubMed] [Google Scholar]
- 31.Ward LD, Kellis M. HaploReg: a resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012;40(Database issue):D930–D934. doi: 10.1093/nar/gkr917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lossow K, Hubner S, Roudnitzky N, Slack JP, Pollastro F, Behrens M, Meyerhof W. Comprehensive Analysis of Mouse Bitter Taste Receptors Reveals Different Molecular Receptive Ranges for Orthologous Receptors in Mice and Humans. J Biol Chem. 2016;291(29):15358–15377. doi: 10.1074/jbc.M116.718544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Capeless CG, Whitney G, Azen EA. Chromosome mapping of Soa, a gene influencing gustatory sensitivity to sucrose octaacetate in mice. Behav Genet. 1992;22(6):655–663. doi: 10.1007/BF01066636. [DOI] [PubMed] [Google Scholar]
- 34.Boughter JD, Jr, Whitney G. Behavioral specificity of the bitter taste gene Soa. Physiol Behav. 1997;63(1):101–108. doi: 10.1016/S0031-9384(97)00398-3. [DOI] [PubMed] [Google Scholar]
- 35.Whitney G, Harder DB. Genetics of bitter perception in mice. Physiol Behav. 1994;56(6):1141–1147. doi: 10.1016/0031-9384(94)90358-1. [DOI] [PubMed] [Google Scholar]
- 36.Roudnitzky N, Bufe B, Thalmann S, Kuhn C, Gunn HC, Xing C, Crider BP, Behrens M, Meyerhof W, Wooding SP. Genomic, genetic and functional dissection of bitter taste responses to artificial sweeteners. Hum Mol Genet. 2011;20(17):3437–3449. doi: 10.1093/hmg/ddr252. [DOI] [PubMed] [Google Scholar]
- 37.Lipchock Sarah V., Spielman Andrew I., Mennella Julie A., Mansfield Corrine J., Hwang Liang-Dar, Douglas Jennifer E., Reed Danielle R. Caffeine Bitterness is Related to Daily Caffeine Intake and Bitter Receptor mRNA Abundance in Human Taste Tissue. Perception. 2017;46(3-4):245–256. doi: 10.1177/0301006616686098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Mennella JA, Pepino MY, Duke FF, Reed DR. Psychophysical dissection of genotype effects on human bitter perception. Chem Senses. 2011;36(2):161–167. doi: 10.1093/chemse/bjq106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Ozdener H, Spielman AI, Rawson NE. Isolation and culture of human fungiform taste papillae cells. J Vis Exp. 2012;63:e3730. doi: 10.3791/3730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Brockhoff A, Behrens M, Roudnitzky N, Appendino G, Avonto C, Meyerhof W. Receptor agonism and antagonism of dietary bitter compounds. J Neurosci. 2011;31(41):14775–14782. doi: 10.1523/JNEUROSCI.2923-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Shim H, Chasman DI, Smith JD, Mora S, Ridker PM, Nickerson DA, Krauss RM, Stephens M. A multivariate genome-wide association analysis of 10 LDL subfractions, and their response to statin treatment, in 1868 Caucasians. PLoS One. 2015;10(4):e0120758. doi: 10.1371/journal.pone.0120758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Manolio TA, Collins FS, Cox NJ, Goldstein DB, Hindorff LA, Hunter DJ, McCarthy MI, Ramos EM, Cardon LR, Chakravarti A, et al. Finding the missing heritability of complex diseases. Nature. 2009;461(7265):747–753. doi: 10.1038/nature08494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Turley P, Walters RK, Maghzian O, Okbay A, Lee JJ, Fontana MA, Nguyen-Viet TA, Wedow R, Zacher M, Furlotte NA, et al. Multi-trait analysis of genome-wide association summary statistics using MTAG. Nat Genet. 2018;50(2):229–237. doi: 10.1038/s41588-017-0009-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wright MJ, Martin NG. Brisbane adolescent twin study: outline of study methods and research projects. Aust J Psychol. 2004;56(2):65–78. doi: 10.1080/00049530410001734865. [DOI] [Google Scholar]
- 45.Green BG, Shaffer GS, Gilmore MM. Derivation and evaluation of a semantic scale of oral sensation magnitude with apparent ratio properties. Chem Senses. 1993;18(6):683–702. doi: 10.1093/chemse/18.6.683. [DOI] [Google Scholar]
- 46.O'Connell J, Gurdasani D, Delaneau O, Pirastu N, Ulivi S, Cocca M, Traglia M, Huang J, Huffman JE, Rudan I, et al. A general approach for haplotype phasing across the full spectrum of relatedness. PLoS Genet. 2014;10(4):e1004234. doi: 10.1371/journal.pgen.1004234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fuchsberger C, Abecasis GR, Hinds DA. minimac2: faster genotype imputation. Bioinformatics. 2015;31(5):782–784. doi: 10.1093/bioinformatics/btu704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Dachet C, Poirier O, Cambien F, Chapman J, Rouis M. New functional promoter polymorphism, CETP/-629, in cholesteryl ester transfer protein (CETP) gene related to CETP mass and high density lipoprotein cholesterol levels: role of Sp1/Sp3 in transcriptional regulation. Arterioscler Thromb Vasc Biol. 2000;20(2):507–515. doi: 10.1161/01.ATV.20.2.507. [DOI] [PubMed] [Google Scholar]
- 49.Davies MN, Verdi S, Burri A, Trzaskowski M, Lee M, Hettema JM, Jansen R, Boomsma DI, Spector TD. Generalised Anxiety Disorder--A Twin Study of Genetic Architecture, Genome-Wide Association and Differential Gene Expression. PLoS One. 2015;10(8):e0134865. doi: 10.1371/journal.pone.0134865. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Mullin BH, Walsh JP, Zheng HF, Brown SJ, Surdulescu GL, Curtis C, Breen G, Dudbridge F, Richards JB, Spector TD, et al. Genome-wide association study using family-based cohorts identifies the WLS and CCDC170/ESR1 loci as associated with bone mineral density. BMC Genomics. 2016;17:136. doi: 10.1186/s12864-016-2481-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ruth KS, Campbell PJ, Chew S, Lim EM, Hadlow N, Stuckey BG, Brown SJ, Feenstra B, Joseph J, Surdulescu GL, et al. Genome-wide association study with 1000 genomes imputation identifies signals for nine sex hormone-related phenotypes. Eur J Hum Genet. 2016;24(2):284–290. doi: 10.1038/ejhg.2015.102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Minica CC, Boomsma DI, Vink JM, Dolan CV. MZ twin pairs or MZ singletons in population family-based GWAS? More power in pairs. Mol Psychiatry. 2014;19(11):1154–1155. doi: 10.1038/mp.2014.121. [DOI] [PubMed] [Google Scholar]
- 53.Nyholt DR. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am J Hum Genet. 2004;74(4):765–769. doi: 10.1086/383251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Hwang LD, fastman: an R package that creates Manhattan and Q-Q plots 10 times faster than the classic way, GitHub repository. https://github.com/danielldhwang/fastman. Accessed 20 Dec 2016.
- 55.Pruim RJ, Welch RP, Sanna S, Teslovich TM, Chines PS, Gliedt TP, Boehnke M, Abecasis GR, Willer CJ. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 2010;26(18):2336–2337. doi: 10.1093/bioinformatics/btq419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jin F, Li Y, Dixon JR, Selvaraj S, Ye Z, Lee AY, Yen CA, Schmitt AD, Espinoza CA, Ren B. A high-resolution map of the three-dimensional chromatin interactome in human cells. Nature. 2013;503(7475):290–294. doi: 10.1038/nature12644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Consortium EP An integrated encyclopedia of DNA elements in the human genome. Nature. 2012;489(7414):57–74. doi: 10.1038/nature11247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Carithers LJ, Ardlie K, Barcus M, Branton PA, Britton A, Buia SA, Compton CC, DeLuca DS, Peter-Demchok J, Gelfand ET, et al. A Novel Approach to High-Quality Postmortem Tissue Procurement: The GTEx Project. Biopreserv Biobank. 2015;13(5):311–319. doi: 10.1089/bio.2015.0032. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Top 100 SNPs on chromosome 12 associated with the perceived intensity of quinine. (DOCX 177 kb)
Table S2. Top 100 SNPs on chromosome 12 associated with the perceived intensity of caffeine. (DOCX 178 kb)
Table S3. Top 100 SNPs on chromosome 12 associated with the perceived intensity of sucrose octaacetate (SOA). (DOCX 181 kb)
Table S4. Top 100 SNPs on chromosome 7 associated with the perceived intensity of denatonium benzoate (DB) from the bivariate analysis of DB and quinine. P_univaraite_DB is the P-value from the univariate analysis of DB. P_univariate DB_adjQ is the P-value from the univariate analysis of DB adjusted for the quinine score. P_bivariate DB_Q is the P-value from the bivariate analysis of DB and quinine. (DOCX 218 kb)
Table S5. Top 100 SNPs on chromosome 7 associated with the perceived intensity of PROP solution. (DOCX 175 kb)
Table S6. Top 100 SNPs on chromosome 7 associated with the perceived intensity of PROP paper. (DOCX 171 kb)
Table S7. Top 100 SNPs on chromosome 2 associated with the perceived intensity of PROP paper and their associations with PROP solution. (DOCX 183 kb)
Table S8. Mean, standard deviation, and heritability estimates for the perceived intensity of bitter tastes. (DOCX 47 kb)
Figure S1. Direction and size of the effects of SNP associations on the perceived intensities of quinine, caffeine, sucrose octaacetate (SOA) and denatonium benzoate (DB). (DOCX 240 kb)
Table S9. Phenotypic and genetic variance in the perceived intensity of quinine, caffeine, sucrose octaacetate (SOA) and denatonium benzoate (DB) explained by rs10772420, rs2597979, rs67487380 and rs10261515. (DOCX 51 kb)
Figure S2. Univariate GWAS for the perception of (a) PROP solution (n = 1757) and (b) PROP paper (n = 1999). Left part are Manhattan plots displaying the association P-value for each SNP in the genome (displayed as –log10 of the P-value). The red line indicates the genome-wide significance threshold of P = 5.0e-8. Right part are regional plots ±400kb from the top SNPs on chromosome 7 for PROP solution and chromosome 2 for PROP paper with the gene model below. (DOCX 444 kb)
Figure S3. Univariate GWAS for the perception of PROP paper from our previous GWAS of 1756 Australian adolescents. A Manhattan plot displays the association P-value for each SNP in the genome (displayed as –log10 of the P-value). This figure is the Fig. 1b from our previous published paper (PMID: 20675712). (DOCX 292 kb)
Table S10. Top PROP paper-associated SNPs on chromosome 2 in the current study (n = 1999) and their associations in GWAS of (1) 225 subjects from the general population of the São Paulo metropolitan area of Brazil, (2) 466 subjects from Silk Road and (3) 2588 subjects from three Italian cohorts. (DOCX 104 kb)
Table S11. Results of the annotation conducted with HaploReg v4.1. (DOCX 119 kb)
Table S12. Genetic variants associated with human bitter taste perception using the sample with one member of each MZ pair removed (see Table 1 for associations using the full sample). (DOCX 24 kb)
Figure S4. The Q-Q plots for each of the univariate analyses. PROP: propylthiouracil. (DOCX 2545 kb)
Figure S5. The Q-Q plots for each of the bivariate analyses. SOA: sucrose octaacetate. DB: denatonium benzoate. (DOCX 2547 kb)
Data Availability Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.