

Abstract
Streams, riparian areas, floodplains, alluvial aquifers and downstream waters (e.g., large rivers, lakes, oceans) are interconnected by longitudinal, lateral, and vertical fluxes of water, other materials and energy. Collectively, these interconnected waters are called fluvial hydrosystems. Physical and chemical connectivity within fluvial hydrosystems is created by the transport of nonliving materials (e.g., water, sediment, nutrients, contaminants) which either do or do not chemically change (chemical and physical connections, respectively). A substantial body of evidence unequivocally demonstrates physical and chemical connectivity between streams and riparian wetlands and downstream waters. Streams and riparian wetlands are structurally connected to downstream waters through the network of continuous channels and floodplain form that make these systems physically contiguous, and the very existence of these structures provides strong geomorphologic evidence for connectivity. Functional connections between streams and riparian wetlands and their downstream waters vary geographically and over time, based on proximity, relative size, environmental setting, material disparity, and intervening units. Because of the complexity and dynamic nature of connections among fluvial hydrosystem units, a complete accounting of the physical and chemical connections and their consequences to downstream waters should aggregate over multiple years to decades.
Keywords: hydrologic connectivity, functional connectivity, river networks, streams, riparian, floodplain wetlands, fluvial hydrosystems
INTRODUCTION
Streams are flowing water bodies that occur as hierarchical, interconnected networks draining surface and subsurface water from watersheds to downstream waters (e.g., large rivers, lakes, estuaries, and oceans). Stream channels, riparian areas, floodplains, underlying alluvial aquifers, and downstream waters have been collectively described as the fluvial hydrosystem (Petts and Amoros, 1996). Most (98%) of fluvial hydrosystems in North America drain to oceans through surface and subsurface flow paths (Vörösmarty et al., 2000). Connectivity is a fundamental property of fluvial hydrosystems, which are characterized by longitudinal, lateral, and vertical fluxes of energy, materials and biota within and among hydrosystem units (Figure 1). The types of connections formed by fluxes through fluvial hydrosystems can be broadly classified as physical, chemical, or biological based on the type of material and energy conveyed (Lamberti et al., 2010).
Figure 1.
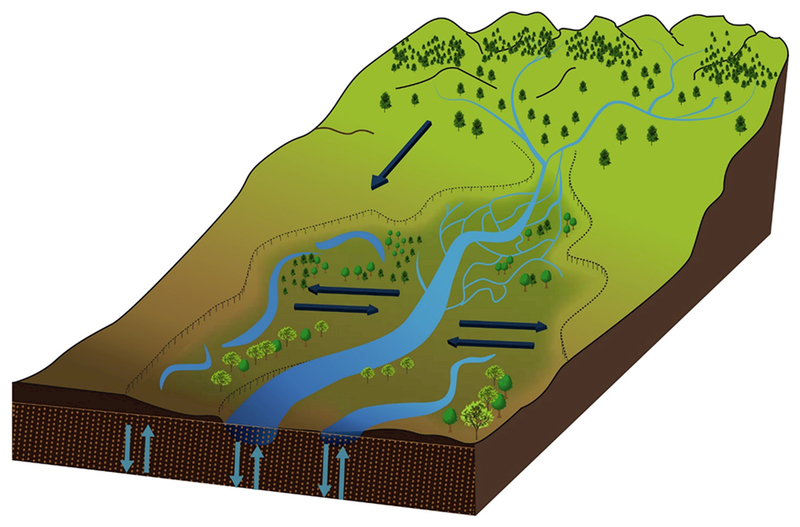
The fluvial hydrosystem with arrows showing the longitudinal, lateral, and vertical transfers among units (Adapted after Petts and Amoros 1996).
The objective of this paper is to review and synthesize existing evidence of the physical and chemical connections by which streams and associated riparian and floodplain wetlands influence the structure and function of downstream waters. The synthesis includes identifying nine key factors governing how downstream waters are connected to and then respond to the fluxes from streams and wetlands. Hereafter, we use the term riparian wetland to collectively describe both floodplain wetlands and other wetlands in riparian areas. Riparian wetlands are distinguished from wetlands situated outside of floodplains and riparian areas by having at least occasional bidirectional, lateral hydrologic flows with the adjacent stream or river segment (Alexander et al., this issue; Lane et al., this issue). Bidirectional flows can be through overbank and hyporheic flows during the lateral expansion and subsequent contraction associated with periodic flooding and other water level oscillations (e.g., seiches, tides).
Physical connections refer to the transport of nonliving materials that do not chemically change en route from streams and riparian wetlands to downstream waters, whereas chemical connections refer to the transport of nonliving materials that can chemically change. Although we recognize that streams and wetlands also exchange water and other materials with nearby terrestrial and deep ground-water systems, this review only considers surface and shallow subsurface flow paths that establish physical and chemical connections from streams and riparian wetlands to larger surface waters, such as rivers, lakes, and coastal bays (Alexander et al., this issue). Evidence of the numerous biological connections between fluvial hydrosystem units by the active and passive movements of living organisms is reviewed by Schofield et al. (this issue). However, this paper does consider biological activities of microbes, primary producers, invertebrates, and vertebrates, in terms of their importance in mediating fluxes of nonliving material between fluvial hydrosystem units.
SYNTHESIZING CONCEPTUAL FRAMEWORKS OF CONNECTIVITY
Connectivity is defined by Leibowitz et al. (this issue) as the degree to which different fluvial hydrosystem units, including streams, riparian wetlands, and downstream waters, are joined and interact, both structurally and functionally. Structural connectivity (or connectedness sensu Baudry and Merriam, 1988) describes the spatial configuration and contiguity of units, whereas functional connectivity describes the spatiotemporally-variable processes linking units (Amoros et al., 1996). Connectivity, therefore, reflects not only the composition and spatial configuration of fluvial hydrosystem units but also how the movement, transformation, and other processes of materials and energy respond to those structural aspects.
The connections that integrate units of environmental systems can be described based on their physical structure, transmission mechanisms, materials conveyed, temporal dynamics, and influences on other units. The functional connectivity framework (Leibowitz et al., 2008; Leibowitz et al., this issue) classifies connectivity according to the functions by which streams and wetlands affect material and energy fluxes to downstream waters. Central to the functional connectivity framework is the idea that fluxes from watershed units to downstream waters can be mediated by 1) functions occurring within units that affect material and energy fluxes and 2) alteration of connectivity or isolation between units that allows or prevents the transport of materials and energy. The distinction between functions occurring within a unit and the transport of materials from a unit to downstream waters varies with the unit. Within-unit functions and downstream transport can occur simultaneously when and where streams are flowing or when rivers and riparian wetlands are connected by high water conditions. Within-unit functions and downstream transport become more distinct when streams and riparian wetlands are temporarily disconnected from downstream waters by drying or other factors that halt or delay transport between units. Most within-unit functions continue under such conditions (although some may be enhanced or suppressed), but the fluvial transport of materials and energy downstream is dramatically reduced or temporarily ceases.
Characterizing connectivity in fluvial hydrosystems that span large, heterogeneous areas can be daunting (Lamberti et al., 2010), even for the relatively narrow set of flow paths considered in this review. It may be helpful to consider the downstream connections of streams and wetlands in terms of the interacting structural and functional elements of flow paths that propagate ecological influence from one hydrosystem unit to another. Reiners and Driese (2001; 2003) proposed a framework for documenting propagation events – that is, how conditions in one location can lead to ecological effects elsewhere. This framework divides propagation events into four elements: 1) the causal agent that initiates event or chronic condition; 2) the conveyance mechanism or vector; 3) the conveyed entity; and 4) the locus of deposition or consequence (Figure 2). In terms of propagation events, streams and their associated riparian wetlands can be considered the causal agents that result in effects at a downstream water (i.e., the locus of consequence).
Figure 2.
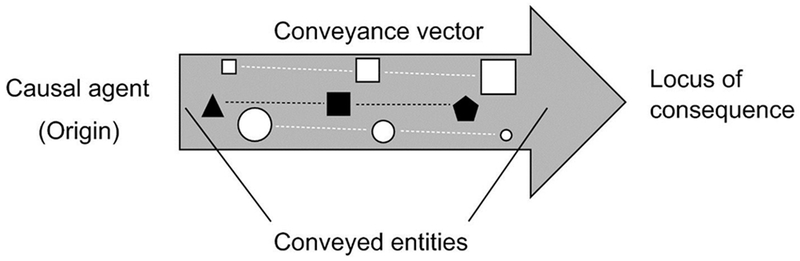
Diagram illustrating the four elements of propagation events (modified from Reiners and Driese, 2003). Different conveyed entities (materials or energy) are portrayed by different colors and shapes. These entities are propagated (dashed lines) along the conveyance vector and describe different functional connections. Entity changes in magnitude (portrayed by size) and form (portrayed by shape) are illustrated as examples of source (white squares), transformation (black symbols), lag and sink (white circles) functions that can occur en route to the locus of consequence (i.e., downstream water).
There are parallels between how the functional connectivity framework classifies connectivity between fluvial hydrosystem units and how others have classified interfaces or boundaries (Amoros et al., 1996; Strayer et al., 2003). For example, conveyance mechanisms and boundaries can be further classified into exchanges or boundary functions that transfer materials and energy (can be neutral to or amplify the transfer) and those that filter, which absorb, reflect, slow or accelerate movement of material energy through obstruction, storage and transformation (Amoros et al., 1996; Strayer et al., 2003). These classifications describe changes in the form and quantities of conveyed entities or materials as they are transported by the conveyance vector from the causal agent to the locus of consequence (Figure 2). Water flow (this paper) and the movement of aquatic fauna (Schofield et al., this issue) are the dominant transport or conveyance mechanisms for cause and effect in fluvial hydrosystems. Table 1 uses examples of stream and riparian wetland connections to downstream waters to align the functional connectivity framework with other frameworks that describe the interactions between fluvial hydrosystem units.
Table 1.
Examples of different functions of streams and floodplain wetlands and their downstream effects.
| Function1 | Exchange2 | Boundary3 | Propagation4 | Downstream Effect | Supporting reference | |||
|---|---|---|---|---|---|---|---|---|
| Function | Origin | Conveyance mechanism | Conveyed entity | Locus of consequence | ||||
| Source | Transfer | Neutral, Amplification | Stream | Fluvial transport | Water | River | More flow, volume, and wetted area downstream | Alexander et al., 2007 |
| Source | Transfer | Neutral, Amplification | Coastal wetland | Seiche | Organic matter | Lake Erie | Subsidy of unique carbon to support life | Bouchard, 2007 |
| Lag | Filter | Transmission | Stream | Overbank flooding | Sediment | Floodplain wetland | Less sedimentation, turbidity | Noe and Hupp, 2009 |
| Transformation | Filter | Transformation | Stream | Hyporheic exchange | Manganese | Hyporheic zone | Reduced contaminant exposure | Fuller and Harvey, 2000 |
| Sink | Filter | Absorption | Stream or wetland | Fluvial transport, overbank flooding | Nitrogen | Atmosphere | Reduced downstream nitrogen loading. | Mulholland et al., 2008; Forshay and Stanley, 2005 |
After Leibowitz et al., this issue;
After Amoros et al., 1996;
After Strayer et al., 2003;
After Reiners and Driese 2001
PHYSICAL AND CHEMICAL CONNECTIONS AS FUNCTIONS
The functional connectivity framework classifies connectivity based on the means by which streams and wetlands influence material fluxes into downstream waters. Five functions (source, sink, refuge, lag and transformation) are used to describe net differences in terms of the quantity, form, and timing of material or energy fluxes from streams and wetlands to downstream waters for a given unit of time (Leibowitz et al., 2008; Leibowitz et al., this issue). Under the functional connectivity framework, the function ascribed to the relationship between a fluvial hydrosystem unit and a downstream water for a given material at within a time and space is mutually exclusive (Leibowitz et al., this issue). However, the function of a fluvial hydrosystem unit may vary with the type of material or energy and vary over space and time for a given material or energy. In the following sections, we present examples that describe these functions (source, sink, lag, and transformation) in terms of the different physical and chemical connections between fluvial hydrosystem units. The refuge function of a stream or wetland enables biota to avoid mortality when conditions are inhospitable in downstream waters. Examples of this function are discussed in Schofield et al. (this issue). Under this framework, functions are dynamic such that a fluvial unit may serve multiple functions simultaneously for different materials or under different environmental conditions for the same materials (Leibowitz et al., this issue).
Source Functions
Streams and riparian wetlands commonly serve as sources of materials or energy to downstream waters (Amoros et al., 1996; Leibowitz et al., this issue). A fluvial hydrosystem unit functions as a source if the quantity of material or energy exported from the fluvial unit is higher than the quantity imported into the fluvial unit (Figure 2).
Hydrologic Source Function.
Water is the key medium transferring energy and other materials between streams and riparian wetlands to downstream waters. Therefore, the physical connection of water flowing through fluvial hydrosystems is the foundation for most other connections (but see Schofield et al., this issue).
Rivers receive more of their water from tributaries than from direct precipitation on or ground-water input into river segments (Bukaveckas, 2009). Rivers are fundamentally cumulative in their formation and maintenance. Flows of materials and energy from headwater channels combine to form larger channels, incrementally accumulating volume and impact. For example, first-order tributaries (designated on 1:100,000-scale National Hydrography Dataset; 57% of total drainage area) in the northeastern United States were estimated to provide ~70% of mean annual water volume in second-order streams and ~55% of mean annual water volume in fourth- and higher order rivers (Alexander et al., 2007). Cumulatively, first-order streams were estimated to be the source of ~60% of mean annual flow to all northeastern streams. First-order stream catchments in the Upper Colorado River basin represent 61% of the total drainage area, but first-order streams produced only 41% of the total annual flow of the Upper Colorado River basin, in part because 84% of the first-order streams were intermittent (i.e., they ceased to flow for part of the year) (Caruso and Haynes, 2011). Although conducted in different hydroclimatic regions of the United States, both studies indicate tributary streams supply much of the water to downstream rivers.
Water in riparian wetlands can originate from direct precipitation, surficial water transported from the river or adjacent hillslopes, groundwater from river infiltration, and groundwater from the hillslope (Amoros and Bornette, 2002). For example, Keizer et al. (2014) concluded that spatial water chemistry patterns in the inundated floodplain show that its water originated from multiple sources, including groundwater mixed with river water during inundation. Multiple riparian wetland water sources were also identified to contribute to inundation patterns prior to the onset of river overbank flow, and following overbank flow there were mixing zones that combined river and riparian wetland source (Mertes, 1997).
Storm flows, particularly when generated by localized storms over a headwater portion of the river network, provide opportunities to observe flow propagation from tributaries to a downstream river. For example, a monsoonal storm event dropped 18-25% of the annual rainfall on a tributary catchment that is ~14% of the Río Grande drainage area (above Ft. Quitman, NM), over just a 2-day period (Vivoni et al., 2006). Storm-flow contributions from an ephemeral tributary stream accounted for 76% of flood volume in the Río Grande despite the flood return interval being only 1.11 to 1.84 years across the gage network. A combination of high antecedent wetness, changes to watershed land cover and river regulation factored into the large contributions from the ephemeral tributary during that moderate-return interval flood.
Sediment Source Function.
Flow from streams and riparian wetlands to downstream waters can also entrain other physical and chemical materials. Sediments are transported through river networks, deposited to form floodplains and other channel features, and thereby influence hydrodynamics and chemical processes (Church, 2006). Sediment entering streams derives from different locations and sources, including hillslopes via overland flow, bank and channel erosion, and infrequent mass wasting (Grimshaw and Lewin, 1980; Miller et al., 2003; Florsheim et al., 2008).
The dynamic balance between sediment supply and transport capacity is a principal paradigm of fluvial geomorphology (Lane, 1955). Sediment load and grain size are dynamically balanced against factors that influence transport capacity (i.e., discharge, channel slope, channel dimensions). If any of these variables change, there is a compensatory response in at least one of the other variables. This balance is particularly relevant to connectivity in river networks, because variables controlling sediment supply and transport capacity change along the river network (Ferguson and Hoey, 2008). Typically, slope and grain size decrease in the downstream direction, whereas channel dimensions and discharge increase (Vannote et al., 1980). Therefore, the cumulative sediment source function of streams to downstream waters mirrors the hydrologic source function, and is reflected in the physical contiguity of river networks, their continuous bed and bank structure, and adjustments in channel dimensions and floodplain form.
Longitudinal discontinuities along downstream waters also illustrate the transfer of material and energy from streams and riparian wetlands (Poole, 2002). Studies have linked abrupt changes in sediment grain size and channel morphology in rivers with tributary confluences with symmetry ratios (ratio of tributary drainage area to the main stem drainage area) falling in the range 0.01 to 0.1 (Rice, 2017). Tributaries with smaller symmetry ratios are less likely to be associated with abrupt mainstem changes because sediment fluxes from such tributaries are likely overwhelmed by sediment fluxes in the mainstem. Tributaries with larger ratios are likely to have sediment fluxes with similar grain size as their mainstems and other conditions (e.g., broad valley width, similar terminal channel slopes) that discourage confluence effects resulting in longitudinal discontinuities (Benda et al., 2004; Rice, 2017). Therefore, tributaries of intermediate size (1/100 to 1/10) relative to mainstem channels will have the highest probability of being associated with channel discontinuities at confluences.
Although surface flows infrequently connect ephemeral streams to downstream waters at any given point in time, the flow magnitudes and sediment loads transported by ephemeral streams can still make them significant sources (e.g., Laronne and Reid, 1993; Coppus and Imeson, 2002). For example, the ephemeral Santa Clara River carries plumes of suspended sediment ~10 km off the southern California coast during 2-y recurrence floods and ~30 km offshore during ~10-y recurrence floods (Warrick and Milliman, 2003). The heavy, sediment-laden flows, referred to as hyperpycnal flows, are caused when the density of water entering the ocean is greater than the receiving seawater. Hyperpycnal plumes are significant because they transport sediment farther offshore than other flows to the continental slope and beyond. These pulsed sediment-laden discharges represent only 0.15% of the loading time period for the Santa Clara River but carry 75% of ephemeral river’s cumulative load offshore of region’s beaches, and thus do not directly contribute to long-term beach maintenance (Warrick and Milliman, 2003).
Chemical Source Function.
Streams and riparian wetlands can also be sources of organic carbon, nutrients, and contaminants to downstream waters. Dissolved and particulate organic carbon support heterotrophic biological activity in fluvial hydrosystems. Nutrients are elements (e.g., nitrogen, phosphorus) required by biota for growth, are often in short supply, and can limit primary production and heterotrophic activity. Contaminants are chemicals that can adversely affect biota when present at sufficient concentrations.
Studies have documented nutrient-based chemical source functions provided by streams and riparian wetlands to downstream waters. For instance, Alexander et al. (2007) investigated how stream size affected nitrogen transport in a northeastern U.S. river network. Approximately 65% of the nitrogen mass in second-order streams, and approximately 40% of the nitrogen mass in fourth- and higher-order streams had been transported from first-order headwater streams (Alexander et al., 2007). Long-term discharge and nutrient concentration data were also used to assess how three Midwestern tributaries functioned as nutrient sources to the Mississippi River (Royer et al., 2006). Nearly all the nitrate-nitrogen and dissolved reactive phosphorus exported by the tributaries occurred when tributary flow was equal to or greater than their median discharge. Over 50% and 84% of the nitrate and phosphorus, respectively, were exported during the largest (> 90th percentile) floods, such that the nutrient source function of these tributaries was largely restricted to periods of high discharge (mid-January through June) (Royer et al., 2006). In another study, investigators used spatial analyses to assess the degree of spatial dependence of water chemistry measurements taken throughout a portion of the Gallatin River network in southwest MT; their results indicated that tributary streams influence downstream nitrate-nitrogen concentrations over channel distances up to 5.5 km downstream (Gardner and McGlynn, 2009).
Longitudinal discontinuities for water chemistry and organic matter distribution along rivers have been identified and attributed to inputs from tributaries (Bruns et al., 1984; Johnson et al., 2010). For example, the location and water chemistry of seeps (small groundwater discharging wetlands) flowing into nearby stream channels in a New Hampshire watershed caused distinct changes in water chemistry throughout the stream network (Zimmer et al., 2013). Broad spatial gradients with water chemistry discontinuities across fluvial hydrosystems likely reflect processes simultaneously operating at different scales (e.g., stream network vs. landscape) and will vary with hydrologic connectivity (McGuire et al., 2014). Spatial patterns of stream chemistry across an entire fifth-order stream network reflected similarities over moderate longitudinal distances (1-2 km) associated with channel-mediated transport and uniform groundwater contributions; discontinuities over short distances were associated with groundwater seeps and the confluences of streams draining contrasting geology (McGuire et al., 2014). However, the lack of chemical discontinuities in fluvial hydrosystems may not always indicate that streams or wetlands do not function as chemical sources. Weak or no spatial patterns can be associated with sources when there is strong biological demand and chemical transformations (see below) of some materials over very short distances (McGuire et al., 2014).
Riparian wetlands are hot spots of productivity (e.g., Nixon, 1980; Cooper et al., 2013) and they can create productivity discontinuities in nearby rivers and lakes. Although riparian wetlands represent 0.7% of the total Laurentian Great Lakes area (pelagic+littoral+wetland), 7-14% of Great Lakes primary productivity was estimated to originate from riparian wetlands (Brazner et al., 2000). Wind-driven water level oscillations (seiches) have strong local ecological effects but are also important in transferring materials from riparian wetlands to nearby lakes (Trebitz, 2006). For example, net export of dissolved and particulate organic carbon (DOC and POC, respectively) from Metzger Marsh, a riparian wetland along the southwestern shore of Lake Erie, to the lake during seiche events was measured at 7.3 mg C L−1 and 3.4 mg C L−1, respectively (Bouchard, 2007). This equates to 4700 Mg of DOC and 2200 Mg of POC exported from the marsh to Lake Erie annually (Bouchard, 2007).
Streams and riparian wetlands also can transfer dissolved and material-bound contaminants to downstream waters. Headwater streams impacted by past mining and smelting activities have been demonstrated to be sources of metal contaminants bound to river sediments hundreds of kilometers downstream (e.g., Hornberger et al., 2009). Chemical spills entering fluvial hydrosystems are among the most publicly recognized disasters and can pose immediate and long-term human health and environmental impacts (e.g., Ruhl et al., 2010; Dollhopf et al., 2014). Because of the widespread nature of streams and wetlands and their hydrologic connectivity, many contaminant spills are transferred to downstream waters if they are not rapidly contained. Often the spatial scale of exposure from such spills can be expansive, so fate and transport models are used for early warning of downstream risks. For example, this approach was used to forecast travel time and downstream concentration of a 37,854-L spill of coal processing organic solvents that leaked from a storage container into the Elk River near Charleston, WV (Bahadur and Samuels, 2014). The peak concentration 402 km downstream on the Ohio River near Cincinnati, OH was modeled to be 14 ppb at 150 hours following the spill, which matched well with observed concentrations from water samples (Bahadur and Samuels, 2014).
Water Temperature Source Function.
Water temperature is an important control of physical (e.g., water density, oxygen solubility), biochemical (e.g., nutrient mineralization), and biological (e.g., organism growth and behavior) characteristics of fluvial hydrosystems. The heat flux within water from streams and riparian wetlands into downstream waters is a source function. The average channel distance over which water temperatures were correlated in a Catskill Mountains, NY river network was approximately 20 km (Gardner and Sullivan, 2004). Water temperature in two contiguous networks in the Central Appalachian Mountains, KY, one largely draining intact forest and the other draining a mixture of forest and surface coal mines, were correlated over an average channel distance of 5 km (Johnson et al., 2010). These findings suggest that water temperature in small streams influences water temperature downstream and that the extent of influence varies geographically as a function of channel length, riparian shading, and extent of groundwater exchange.
Water temperature maps and longitudinal profile plots show some tributary confluences coincide with distinct peaks and troughs in river temperature and, at times, can be useful in predicting the distribution of biota (Torgersen et al., 2001). For example, confluences with spring-fed streams originating on floodplains were identified as the coldest patches along a northeastern Oregon river that otherwise had summer water temperatures exceeding the tolerance limit of native salmonids (Ebersole et al., 2003). A subsequent study determined that tributary confluences commonly provided coldwater patches (≥ 3 °C colder than mainstem temperatures) during the summer (Ebersole et al., 2015). Spring snowmelt can be a significant contributor to summer base flow in the region. Interestingly, 39% of these confluences were with tributaries that contributed cold hyporheic water even when they lacked surface water—that is, they were ephemeral and intermittent tributaries that were significantly connected to downstream waters even when they lacked surface flow (Ebersole et al., 2015). Whether or not a tributary confluence would be a cold patch was not predicted by tributary size, surface flow presence, or flow-path length but was largely explained by amount of available water at the end of the snowmelt season (Ebersole et al., 2015).
As these examples illustrate, a substantial body of evidence demonstrates that streams and riparian wetlands are connected to downstream waters as sources. Longitudinal and lateral transport of surface water (e.g., via stream flow, seiches) are key physical mechanisms for transferring sediment, organic matter, nutrients, contaminants, and heat energy from streams and riparian wetlands to downstream waters. The physical form and contiguity of streams and floodplains with downstream waters is in itself a manifestation of this source function, via the dynamic balance of sediment supply and transport capacity. The source functions provided by these units vary over time and space, often alternating with other connectivity functions (i.e., sink, lag, and transformation functions) that counteract their role as sources of materials and energy to downstream waters.
Sink, lag & transformation functions
In addition to serving as sources of materials and energy for downstream waters, streams and riparian wetlands can mediate physical and chemical connections to downstream waters by decreasing or halting transport (sink function), slowing or storing (lag function), and converting (transformation function) materials and energy (Leibowitz et al., 2008; Leibowitz et al., this issue). The degree to which streams and riparian wetlands fuel downstream waters with material and energy inputs depend on the extent to which streams and riparian wetlands transform, halt, and/or store that material and energy (Covino, 2017). The framework presented in Leibowitz et al. (this issue) categorizes functional connectivity types based on the net change, timing, and compositional shits that result from connections. However, in many cases the mechanisms associated with sink, lag, and transformation functions are not mutually exclusive within and between fluvial system units, and these functions do not operate in isolation. Even though these functions may not always be easily delineated, recognition of the differences among functions is useful in understanding how individual fluvial units influence downstream waters.
Hydrologic Sink Function.
Streams and riparian wetlands can function as sinks, over varying spatial and temporal scales, by preventing surface flow from reaching downstream waters and thereby minimizing downstream flooding. Transmission losses (i.e., losses of surface flow volume by infiltration into underlying channel alluvium), evaporation,and evapotranspiration are ways that streams and wetlands can function as hydrologic sinks. Seepage through channel beds and banks commonly dominates these losses, although evaporation and evapotranspiration losses can be significant in stream reaches with prolonged surface flows (Dahm et al., 2002; Hamilton et al., 2005;). For example, cumulative transmission losses over 54 km of channel reduced peak flow volumes by 57% from a storm in an arid southwestern United States watershed (Renard and Keppel, 1966). Similarly, up to half the flow volume in three ephemeral channels in CA and NM was lost to channel transmission (Constantz et al., 2002). The Río Grande’s mainstem, which has a multi-threaded channel and an extensive floodplain area, is conducive to transmission losses via infiltration and groundwater recharge. Along a 127-km distance downstream of a dense riparian forest on the mainstem, these transmission losses accounted for a 49% decrease in flood wave volume (at rates of 39 to 46 m3/m losses to the underlying aquifer) before reaching a downstream reservoir. As a result of these rapid transmission loss rates during peak flood conditions, the groundwater table response declined from a 0.6-m rise to a <0.3-m rise in the lowest floodplain transect (Vivoni et al., 2006). These examples clearly demonstrate how streams and riparian wetlands can function as hydrologic sinks to downstream waters.
Chemical Sink Function.
The diversion of water by channel transmission from surface waters can also be a sink mechanism for contaminants. Field and laboratory experiments have shown that beds of streams and ditches are preferential areas for pesticide infiltration into underlying groundwater, but that they also retain pesticides through sorption to bed sediments and deeper soils (Burkart et al., 1999; Dages et al., 2015). Streams and riparian wetlands can also be sinks for excess nutrients. For example, first- to third-order and first- to fifth-order streams are estimated to remove 8% and 16% of nitrogen, respectively, from river networks to the atmosphere via denitrification (Alexander et al., 2007; Wollheim et al., 2008). Headwater streams draining natural land cover were estimated to reduce downstream delivery of nitrogen by 20-40% (Mulholland et al., 2008). Based on a combination of field and experimental data, floodplain wetlands along the Wisconsin River, a 7th-order tributary to the Mississippi River, are a sink for 37% of the nitrate load entering a 4-km reach of the river during a 2-year return interval flood (Forshay and Stanley, 2005). Riparian wetlands incidentally created during the restoration of a Maryland urban stream were sinks for 23% to 87% of the nitrate load entering the wetlands during four storm events (Harrison et al., 2014).
Streams and riparian wetlands also function as carbon sinks through sequestration in floodplain sediments, photo-oxidation, and outgassing to the atmosphere (Cole et al., 2007; Battin et al., 2009). Freshwater systems are increasingly recognized as important contributors of carbon dioxide and methane to the atmosphere; relative to land, streams in particular are large contributors of carbon dioxide to the atmosphere because of their high partial pressure of dissolved carbon dioxide and gas transfer velocity (Saarnio et al., 2009; Butman and Raymond, 2011; Raymond et al., 2013). Ephemeral and intermittent streambeds and wetlands function as sinks by redirecting materials from downstream transfer to the atmosphere, even when they are dry and especially immediately after rewetting (Fenner and Freeman, 2001; Gallo et al., 2013). Because of their predominance in arid regions, ephemeral and intermittent channels were estimated to account for at least half of the carbon dioxide emissions from all rivers and streams (Von Schiller et al., 2014).
Sediment Sink Function.
Streams and riparian wetlands can prevent sediment and associated contaminants from being transported downstream. Most headwater streams have low competence to transport sediment during base flow (Gooderham et al., 2007), and contain structures (e.g., boulders, large wood) that retain and store colluvial sediments between episodic storm flows when sediment is primarily transferred downstream (Gomi and Sidle, 2003). Because of their abundance and distribution, headwater streams and associated riparian wetlands can have a substantial cumulative sink effect by intercepting and trapping sediment transported from hillslopes. For instance, Sediment accretion and accumulation rates were measured on forested riparian wetlands along seven Atlantic Coastal Plain rivers draining into Chesapeake Bay (Noe and Hupp, 2009). Sediment accumulation rates in riparian wetlands ranged from 303 – 4,600 g m−2 y−1, averaging 976 g m−2 y−1. Scaling up to the entire wetland area, sediment retention in riparian wetlands ranged from 30 – 690% of a river’s annual sediment load, with a mean of 119% of annual sediment load across the seven rivers (Noe and Hupp, 2009). Floodplain wetlands trap and store sediment carried by flooding rivers as overbank water slows down over floodplains and floodwaters recede. For example, Kleiss (1996) measured a 16% decrease in sediment load between upstream and downstream points in the Cache River, AR that were contiguous with a bottomland hardwood wetland. Deposited sediment that remains in riparian wetlands shape the habitat mosaics that contribute to the chemical and biological patterns and diversity of fluvial hydrosystems (Amoros and Bornette, 2002; Ward et al., 2002). The associated organic matter and nutrients also fuel the high level of plant productivity in riparian areas (Tabacchi et al., 1998).
Hydrologic Lag Function.
Streams and riparian wetlands can temporarily store water, thereby delaying transfer to downstream waters. This lag function of fluvial hydrosystem units on hydrologic response can be characterized as dispersion or the spreading of water output across different spatial scales over its residence time (Saco and Kumar, 2002). Hydrologic dispersion is subdivided to describe lag at different scales. Hydrodynamic dispersion is the local, temporary storage that makes some of the channel’s water volume flow downstream faster than other parts of its volume, rather than the entire volume being transported as a single, discrete pulse. Local transient storage in the hyporheic zone, along channel margins, in off-channel sloughs, and in eddies behind obstructions all contribute to the temporary storage and slow downstream transport (Harvey et al., 2005; Briggs et al., 2009). Lag at the network scale is described by geomorphologic and kinematic dispersion. Geomorphologic dispersion is the cumulative effect of different travel distances throughout a river network to its outlet (Rodríguez-Iturbe and Valdes, 1979). Kinematic dispersion takes into account how river network structure contributes to hydrologic response by incorporating the cumulative effect of spatially variable water velocity (or hydraulic geometry) as water flows through the network (Saco and Kumar, 2002). The lag described by the three scales of hydrologic dispersion is therefore fundamental to the spread of water flowing through fluvial hydrosystems, integrating the cumulative effects of local structure and the hierarchical structure of river networks.
Downstream transport of peak flows following storms can be delayed via bank storage or the transfer of stream water to an alluvial aquifer for temporary storage. Bank storage is gradually released after flooding subsides, augmenting base flow (Whiting and Pomeranets, 1997; Menichino and Hester, 2015). Kondolf et al. (1987) investigated the role of bank storage on streamflow in the Carmel River in central CA. They identified three key conditions for substantial bank storage: 1) stream stage must increase during floods; 2) bank material must be permeable; and 3) there must be sufficiently high volume within the permeable bank material for storage relative to the base-flow stage. These conditions appear to favor individual, large channels, but this effect diminishes when cumulative effects of headwater streams are considered. The lower 24 km of the Carmel River flows through an alluvial valley with alluvium (15-30 m deep) that increases from <1 to 50 m wide near the river mouth. Typically discharge increases proportionally with drainage area, but 60% of the first storm flows of the wet season (Dec-Feb) each year had discharges that decreased with drainage area. This decline reflected storm-flow losses to bank storage and shallow aquifers. One month after peak storm flow, water released from bank storage was estimated to contribute ~20% of the recession flow along the lower 24 km of the river. The lag continued for another month before bank storage was depleted and discharge declined downstream due to transmission losses (Kondolf et al., 1987).
Temporary storage and subsequent release of water by riparian wetlands can also attenuate flooding in downstream waters. For example, peak discharges between upstream and downstream segments of the Cache River, AR were reduced 10-20% mainly through riparian wetland storage and subsequent release (Walton et al., 1996). A review of published studies on wetland hydrology reported that 23 of 28 studies on riparian wetlands from all regions of the world concluded riparian wetlands reduce or delay floods (Bullock and Acreman, 2003).
Chemical Lag Function.
Streams and riparian wetlands delay and disperse the transport of pollutants over time, increasing opportunity for degradation and thereby reducing exposures in downstream waters. Radioactive chemical releases provided unintended, tracer experiments that revealed the chemical lag function within fluvial hydrosystems. From 1942-1952, untreated plutonium was discharged from Los Alamos National Laboratory, NM into intermittent headwater tributaries of the Río Grande, and also entered the watershed from nuclear weapons testing fallout (Graf, 1994). Although the catchment area of the intermittent headwater tributaries represented only 0.4% of the Río Grande drainage area at their confluence with the Río Grande, the mean annual bed load of plutonium-bound sediment from the tributaries was ~sevenfold greater than that from the mainstem. Most bed-load transport occurred sporadically during intense storms. Only 10% of the plutonium directly discharged into the river network, and less than 2% of the fallout were exported to the Río Grande; most of it was bound to sediment and soil that had either not yet been transported to the river network or was stored on the floodplain or in tributary channels (Graf, 1994). Approximately 50% of the plutonium that entered the Río Grande was stored in the river and its floodplain, and the remainder was transported downriver to Elephant Butte reservoir (Graf, 1994).
Transformation Functions.
Transformation of materials and energy is fundamental to the form and function of fluvial hydrosystems. The transformation of potential energy (gravitational) to kinetic energy results in work that contributes to turbulence, bed friction, erosion, and material transport (Knighton, 1998). Energy is also transformed or dissipated from the system as heat. The geometry of channels and river networks has been proposed to be governed by the minimization of energy transformation necessary for transporting materials across landscapes (e.g., Molnár and Ramírez, 1998; Huang et al., 2004).
By mediating the form of materials and energy, either in temporary storage or flux, streams and riparian wetlands alter the mobility of those materials. Materials cycle through various forms or ecosystem compartments, as they are taken up or assimilated and excreted by organisms or otherwise regenerated for reuse. Downstream water chemistry reflects the combined processes of biogeochemical cycling and transport (Webster and Patten, 1979) that occur throughout the fluvial hydrosystem but function most intensively in headwater streams (e.g., Peterson et al., 2001; Aufdenkampe et al., 2011). For example, Ensign and Doyle (2006) compiled and normalized the results of 404 phosphate, ammonium, and nitrate uptake measurements from streams and rivers ranging from first- to fifth-order. Nitrogen cycled between approximately 8 and 90 times within the respective lengths of first- to fourth-order streams (Ensign and Doyle, 2006). In another review, phosphorus uptake lengths (distance traveled by dissolved nutrients before biological acquisition) were reported to be shortest (2-530 m) in first-order streams, longer (26-3,460 m) in second- to fourth-order streams, and longest (4,140-367,000 m) in fifth-order rivers (Withers and Jarvie, 2008). Because roughly half of the water reaching larger tributaries and rivers originates from headwater streams, it is clear that most of the phosphorus and nitrogen arriving at downstream waters has already been transformed and recycled many times in upstream channels.
Overbank flooding can export large quantities of nutrients from rivers to riparian wetlands for transformation and storage. For example, a nutrient amendment study was conducted to examine the allocation of nutrients in a riparian wetland along the Tar River, NC (Brinson et al., 1984). The nutrient budget indicated that 53% and 49% of the amended nitrogen and phosphorus, respectively, were transformed by denitrification, microbial assimilation, plant uptake, or adsorption, and thus, in part, stored in the riparian wetland (Brinson et al., 1984).
As materials are cycled during longitudinal, lateral, and vertical transport through the fluvial hydrosystem, they are exposed to different physicochemical conditions over varying residence times (Covino, 2017). These physicochemical gradients regulate, in part, material transformations that subsequently affect the type of materials transported downstream (e.g., Neubauer et al., 2013). For example, physicochemical conditions and residence time associated with hyporheic exchange enabled stream microbial activity to reduce the dissolved manganese load by 20% in a stream contaminated by copper mining (Harvey and Fuller, 1998). The oxidation of manganese enhanced the adsorption of other trace metals and thereby decreased cobalt, nickel, and zinc export by 12-68% (Fuller and Harvey, 2000). Managing the connectivity between fluvial hydrosystem units is becoming more widely recognized as an important tool in the protection and remediation of chemical contaminants in water resources (e.g., Newcomer-Johnson et al., 2016).
Headwater streams support downstream biological activity by supplying downstream waters with dissolved and particulate organic carbon. Almost all organic carbon exported to downstream waters has been physically and chemically transformed in headwater streams and riparian wetlands. On average, leaf litter represents half of the organic matter imported into forested headwater streams (Benfield, 1997) but accounts for ≤ 2% of organic matter exported from these streams (Naiman and Sedell, 1979; Minshall et al., 1983). Terrestrial leaves are transformed into dissolved organic matter, fine particles, and living biomass through interacting organic matter breakdown processes (leaching, microbial assimilation, abrasion, and macroinvertebrate shredding and consumption) in stream channels and riparian wetlands. Organic turnover length is the ratio of downstream organic carbon flux to ecosystem respiration per length of stream, and approximates the average distance that organic carbon travels before being consumed and mineralized (Newbold et al., 1982). Carbon turnover length for first-order streams ranges from 1 to 10 km (Newbold et al., 1982; Minshall et al., 1983), indicating that organic carbon exported from headwater streams is likely to be exported to and used in larger (second- or third-order) streams. However, carbon turnover length is a weighted average of widely varying turnover lengths associated with a broad range of particulate and dissolved forms. For example, turnover lengths for whole leaves and fine organic particles in a North Carolina mountain headwater stream were estimated to be 108 m and 40 km, respectively. Newbold et al. (2005) similarly estimated 38 km and 59 km for the turnover lengths of two different size fractions of fine organic particles in an Idaho headwater stream. Dissolved organic carbon in a southern Pennsylvania stream consisted of a labile fraction with turnover length of 240 m, a semilabile fraction with turnover length of 4500 m, and a refractory fraction with an indefinitely long turnover length sufficient to carry the carbon to coastal waters (Kaplan et al., 2008).
Streams and riparian wetlands are connected to downstream waters through their sink, lag, and transformation functions which can alternate or occur simultaneously with material transport to downstream waters. Just as the channel continuity between streams and downstream waters reflects the hydrologic source function of streams, the geomorphic setting of floodplains and their associated wetlands reflects deposition or storage of sediment and other materials. Sink, lag, and transformation functions can occur at particular times and places associated with particular physicochemical conditions or organisms. However, some connectivity functions are described cumulatively over space and time, such as hydrologic lag through river networks. The function by which streams and riparian wetlands affect material and energy fluxes to downstream waters will vary over time and space. This variation can be governed by a suite of factors that influence the degree of connectivity or isolation between hydrosystem units.
Biological Mediation of Physical and Chemical Connectivity
The activities (e.g., habitat creation, bioturbation, bioconsolidation) of aquatic organisms can influence the storage, transfer and transformation of materials and energy (Gutiérrez and Jones, 2006; Moore, 2006). These activities range from diurnal (Beck et al., 2009; Richardson et al., 2009) to annual (Hassan et al., 2008) cycles occurring over variable to stable spatial scales depending on organism behavior and life history (Zanetell and Peckarsky, 1996; Fritz et al., 2004). For example, headwater streams and their valleys in old-growth Colorado forests, which represent <1% of the watershed by area, store ~23% of the organic carbon in the watershed in large part by the beaver dams that enhance overbank flooding, floodplain organic matter deposition and retention, and elevated groundwater levels (Wohl et al., 2012). Because much of this carbon is in the form of floodplain sediments and coarse wood, this storage is estimated to last from 100 to 1000 years (Wohl et al., 2012).
Small organisms can cumulatively have similar or more expansive effects on sediment transport than larger, less frequently occurring organisms like beavers (Albertson and Allen, 2015). For example, the discharge associated with initiating bed sediment transport is typically a bankfull (i.e., 1.5-yr return interval) flood, but bioturbation activity of crayfish initiates equivalent sediment movement at discharges that occur every day (Statzner, 2012). In contrast, the bioconsolidation activity of net-spinning caddisflies increases the discharge required to initiate transport equivalent to that of a 100-yr flood (Statzner, 2012).
FACTORS AFFECTING CONNECTIVITY AND ISOLATION
Many factors affect the connectivity between streams and riparian wetlands and their downstream waters and thus the functions these streams and riparian wetlands serve (Table 2). Within a given fluvial hydrosystem, the degree of connectivity between streams and riparian wetlands and downstream waters varies over space and time. The expansion and contraction of river networks in response to storm events, seasonality, and multi-annual phenomena (e.g., El Niño) results in different degrees of hydrologic connectivity along the longitudinal, lateral, and vertical dimensions. Although longitudinal flow is unidirectionally downstream, lateral and vertical exchanges are bidirectional and their flow direction varies with local physical context and hydrologic conditions. For example, water and materials move from the river to adjacent floodplain wetlands during overland flooding but move from floodplain wetlands to the river during recession (Amoros and Bornette, 2002; Jung et al., 2004). Recognizing the key factors governing spatial and temporal variation in physical and chemical connectivity is fundamental to understanding how streams and riparian wetlands affect downstream waters.
Table 2.
Key factors (bold) affecting connectivity from streams and riparian wetlands to downstream waters and its resulting effects.
| Factors | Connectivity | Effect |
|---|---|---|
| Proximity | Closer → Stronger, frequent, longer Farther → Weaker, less frequent, shorter |
Closer → Stronger Farther → Weaker |
| Individual size | Larger → Stronger, more likely Smaller → Weaker, less likely |
Larger → Stronger Smaller → Weaker |
| Number | More → Stronger, more likely Fewer → Weaker, less likely |
More → Stronger, stable Fewer → Weaker, variable |
| Climate | Wetter → Continuous Drier → Episodic |
Wetter → Stable Drier → Variable |
| Geology | Impermeable → More surface Permeable → More subsurface |
Stable, Indistinct → Permeable Variable, Distinct → Impermeable |
| Terrain/Slope | Steep → Rapid Gradual → Slow |
Steep → Variable, indistinct Gradual → Stable, distinct |
| Land use/cover | Homogeneous → Variable Heterogenous → Stable |
Homogeneous → Variable Heterogenous → Stable |
| Distinctiveness | May be related to timing and characteristics of the transported material | Dissimilar → Detectable Similar → Undetectable |
| Intervening units | Facilitate → Stronger Resistant → Weaker |
Facilitate, congruent → Stronger Resistant, contradictory → Weaker |
Weak connectivity = episodic (low frequency, low duration) and/or slow (low rate, low magnitude)
Strong connectivity = continuous (high frequency, high duration), and/or rapid (high rate, high magnitude)
Proximity
Proximity is one of the more straightforward factors affecting connectivity. Typically, the closer a tributary or wetland is to a downstream water, the more likely that tributary or wetland is to have strong or frequent connections to the downstream water. A study of two oxbow lakes that vary in their distances to the nearby river provides such an example (Negrel et al., 2003). The more recently formed oxbow lake was closer to the river channel, more frequently connected to the river through surface flows, and thus had water geochemistry more similar to the river than the more distant oxbow lake (Negrel et al., 2003). The older, more distant oxbow lake was more dependent on subsurface flow.
The hydrologic connectivity of flood pulses to water bodies within the floodplain can also vary. The lake levels of two oxbow lakes, one recently formed and one older, along the lower Guadalupe River, TX differed in their relationship to river discharge (Hudson et al., 2012). Lake level of the recently formed oxbow lake had a strong linear relationship with river discharge in which lake level peaks and recessions were nearly synchronous with river discharge, indicating tightly connected hydrology. In contrast, the older oxbow lake had more complex hydrologic connectivity with the river, reflected in a nonlinear relationship between river discharge and lake level (Hudson et al., 2012).
Although the presence of a continuous channel is itself a physical reflection of hydrologic connection (Wolman and Gerson, 1978), channel distances between tributaries and downstream waters are among the factors that determine the degree of connectivity. The distribution of channel distances to a downstream river for tributaries of a given size is a function of basin shape and network configuration. Compact basins tend to have dendritic networks; elongate basins tend to have trellis networks. Most headwater streams in dendritic networks are more distant from downstream waters than headwater streams of trellis networks. Locations along rivers can be described as positively autocorrelated: locations are not independent, and those closer to one another are more strongly correlated with one another than those separated by greater distances. Numerous studies have demonstrated spatial dependence of physical and chemical parameters measured throughout stream networks (e.g., Gardner and McGlynn, 2009; McGuire et al., 2014). The decline in relationship strength with increasing distance occurs in large part because of the sink, lag, and transformation functions performed by upstream waters, which counteract the source function as materials are transported downstream. In addition, the likelihood of gradual or sudden changes associated with other interacting factors (e.g., geology, barrier, effluent discharge) increases with increasing distance between locations.
However, proximity defined simply by two-dimensional distance may not accurately reflect connectivity due to spatial differences in landscape resistances to water and material transport. For example, riparian wetlands farther from the main river channel may be more frequently connected for longer periods than wetlands contiguous to the main river channel because of flow through side channels, permeable alluvium, or other preferential flow paths (Poole et al., 2002; Phillips, 2013).
Size and density
The size and density of streams and riparian wetlands are other factors that affect connectivity. Generally, larger streams—and sometimes larger riparian wetlands—will have closer proximity to downstream waters and so will likely maintain more frequent and longer connections to downstream waters. The likelihood of an individual connection, and the combined strength of cumulative connections to downstream waters, increases with increasing stream and wetland density. Headwater tributaries and riparian wetlands are the exterior interface between uplands and the fluvial hydrosystem. Baker et al. (2007) compared how this interface changes with stream density using different stream map resolutions. The proportion of the watershed area occurring near a stream tripled from low resolution (median = 0.6 km−1) to high resolution (2.2 km−1) maps (Baker et al., 2007). Increasing stream and wetland densities increase travel times by increasing channel length and creating more opportunities for lag and transformation functions. For example, model simulations predicted that watersheds with high headwater drainage densities would have longer lag times for mercury delivery to downstream waters compared to watersheds with low headwater drainage densities (Knightes et al., 2009).
Climate and landscape characteristics
Physical and chemical connectivity in fluvial hydrosystems vary with climate and landscape characteristics. Climate governs form, distribution, rate, magnitude, and timing of water supply. A higher proportion of streams in more mesic regions of the United States have year-round flow, whereas in semiarid and arid regions a higher proportion have ephemeral flow (Nadeau and Rains, 2007). Temperature and moisture levels are also fundamental physical factors that govern ecosystem processes affecting material transport. For example, transformations tend to be enhanced and storage reduced with warmer temperatures and increased moisture, although saturated conditions can limit aerobic respiration (e.g., Strauss et al., 2002; Sutfin et al., 2016).
Landscape characteristics including geology, soils, and terrain govern local drainage patterns and functions and structural connectivity throughout fluvial hydrosystems. River segments flowing through steeper terrain have narrower floodplains and smaller riparian wetlands than river segments flowing through more gradual topography (Karim et al., 2012). Most riparian wetlands along rivers draining confined valleys are inundated during even small floods. Rivers flowing through gradual topography may frequently overtop their banks and inundate adjacent wetlands, but less frequent, high magnitude floods are typically required to inundate all their floodplain wetlands, unless more distant wetlands have other sources of water and the wetlands can coalesce (Karim et al., 2012; Keizer et al., 2014). Changes in landscape characteristics (e.g., geology, groundwater contribution) influence spatial patterns of hydrological connectivity in fluvial hydrosystems which are reflected in the physical, chemical, and biological gradients seen across connected units (e.g., Katz et al., 1997; Fergus et al., 2017). The spatial extent of base-flow discharge patterns was assessed across nested gages distributed in the Catskill Mountains river network, NY (Shaman et al., 2004). Because base flow in headwater streams (i.e., those with watersheds <8 km2) was more weakly correlated with mainstem flow than base flow in larger streams, the authors concluded that deep groundwater contributed more to mainstem discharge as drainage area increased.
Land cover and use
Human activities and structures alter connectivity between streams and riparian wetlands and downstream waters. Modifications of hydrologic connectivity are a ubiquitous source of impairment to U.S. rivers (e.g., Meador, 1996; Gregory, 2006; Gordon et al., 2015). Activities (e.g., water abstraction) and structures (e.g., dams) can reduce longitudinal connectivity. Channelization, dredging, bank stabilization, and levee construction enhance longitudinal connectivity but decrease vertical and lateral connectivity (Valett et al., 2005), ultimately reducing sink, lag, and transformation functions, but possibly enhancing source function. Most temperate floodplains—and a growing number in other regions—have been modified by land use and disconnected from their streams and rivers. River flows are modified and contained within artificial levees so that floodplains and their wetlands can be converted and maintained for other land uses. The recurrent conveyance of floodwaters of varying magnitudes and durations onto floodplains, which contributed to the conditions ideal for such land uses (e.g., productive soils, flat topography), are reduced or eliminated, thereby affecting connectivity to and functions within floodplain wetlands (Opperman et al., 2010).
Many streams in human-dominated watersheds, particularly in arid and semiarid regions, receive the bulk of their summer low flow from effluent discharges (Rice et al., 2013). Streams that would be dry in the absence of these discharges are called effluent-dependent streams; streams that receive most, but not all, of their flow from effluent are called effluent-dominated streams (Brooks et al., 2006). About 25% of permitted effluent discharges in the United States enter streams with mean annual discharges incapable of diluting effluents by more than 10-fold; this percentage increases to 60% when low-flow discharges are considered (Brooks et al., 2006). However, treated wastewater discharge has been recognized as a way to augment stream connectivity that would otherwise be limited (Luthy et al., 2015). Streams draining human-dominated areas also can derive base flow from groundwater recharged by over-irrigation and leaky infrastructure (Lerner, 1986; Townsend-Small et al., 2013).
Land cover and land use changes also alter connectivity between streams, riparian wetlands and downstream waters. Urban development has a strong effect on hydrologic connectivity by drastically altering the frequency, duration, magnitude, timing, and rate of change of storm flows and base flows. Connectivity can be enhanced or fragmented by these alterations. Impervious cover, burial of natural channels, and storm drains rapidly deliver stormwater away from buildings and roads to downstream channels which increase hydrologic connectivity. However, by shortening storm-flow durations, these alterations reduce infiltration and watershed storage of water, reduce base-flow discharge and can reduce hydrologic connectivity (but see Bhaskar et al., 2016).
Ditch drainage is a particular form of hydromodification that elevates peak flows and facilitates the transport of runoff and associated pollutants to rivers. Ditches concentrate and direct surface runoff, capturing and converting shallow subsurface flow to more rapid channel flow and lowering soil moisture downgradient of roads. A study determined that 94% of road ditches were connected to streams, more than doubling the channel length in a central New York watershed. These ditches contributed 22% and 29% of total flow at the watershed outlet during wet and dry season storms, respectively (Buchanan et al., 2013). The presence of ditches was calculated to increase the maximum and total storm flow downstream by 78% and 57%, respectively. Given that 75% of ditch channel length was contiguous to unbuffered agricultural areas, it is likely that ditches were also conduits of non-point source pollutants to downstream waters (Buchanan et al., 2013).
Land use can also affect chemical connectivity by changing the distribution of chemical sources, the conditions conducive for chemical transformations, and hydrologic conveyances. For example, in some Illinois watersheds with tile drains and channelized headwaters, stream nitrate concentration is positively correlated with stream discharge, suggesting that these altered streams are rapidly transporting nitrate inputs downstream with little retention or processing (Royer et al., 2004). Model simulations predicted that total mercury fluxes from a mixed (forest, pasture, and developed) landscape to downstream waters would increase by 95% under an increased suburbanization scenario, but decrease by 7% in total and methylmercury export under a reforestation scenario (Golden and Knightes, 2011).
FACTORS INFLUENCING EFFECTS ON DOWNSTREAM WATERS
Multiple key factors govern how physical and chemical connectivity affect downstream waters (see Table 2, which summarizes the factors using a binary approach). We recognize that many of these relationships are likely more complex and that factors are likely to interact, but we have separated them here for clarity.
Size and density
The relative size of the stream or wetland connected to the downstream water is often a useful predictor of downstream effect. Symmetry ratios characterize tributary size (in terms of discharge, drainage areas or channel widths) relative to mainstem size at river confluences (Benda, 2008). Other characteristics such as geology, topography, and land use associated with tributaries or wetlands can influence the effect of symmetry ratios. For example, steeper tributaries will tend to have a more discernible effect on mainstem sediment grain size at confluences, potentially disrupting downstream fining along mainstems and increasing habitat heterogeneity (Rice, 1998). The individual effect or discontinuity associated with a tributary confluence, if discernible, is typically present for a short segment of river channel (e.g., Rice and Church, 1998; Torgersen et al., 2001), often because of the overwhelming effect of the mainstem relative to the minor transport capacity of a single tributary.
However, the cumulative or aggregate effect of many tributaries and their associated riparian wetlands over time is reflected in the more gradual downstream changes in physical and chemical characteristics of the mainstem (Vannote et al., 1980; Knighton, 1998). The network structure of fluvial hydrosystems, including longitudinal, lateral and vertical connections, homogenizes solute concentrations downstream. Convergent spatial distribution patterns of water chemistry provide evidence for these aggregate effects and have been documented in natural and human-dominated systems (e.g., Asano et al., 2009; Hoellein et al., 2011). Size and number of streams and wetlands are important factors governing how downstream waters connect to tributaries and riparian wetlands and then respond to their contributions.
Cumulative contributions associated with the spatial structure of river networks affect how downstream waters vary over time. Flowing water within fluvial hydrosystems expands and contracts in response to meteorology (e.g., short-term precipitation events, seasonal weather shifts, multi-annual oscillations). The watershed stability hypothesis states that the structure and connectivity of fluvial hydrosystems dampen fluctuation magnitudes in downstream waters by integrating the asynchronous dynamics of contributing streams and riparian wetlands (Moore et al., 2015). This aggregation of contributions is akin to the portfolio effect, where having a diverse set of investments buffers against market volatility and has a stabilizing effect on overall investment (Doak et al., 1998). The hypothesis proposes that stability should increase with downstream direction in river networks because hierarchical connectivity of river networks increasingly integrates more diverse conditions with size and connectivity largely in the downstream direction.
To test the hypothesis Moore et al. (2015) compiled spatially explicit discharge and summer water temperature data from 142 sites (19.8 - 229,684 km2) throughout the network of the second largest unimpounded river in North America, the Fraser River in BC, Canada. Sites with highest variability were sites with small drainage areas; small sites that did have stable discharge and temperature were usually downstream of lakes. Flows in the largest portions of the network were 2.2 times more stable than those in the smallest catchments. Extreme flows occurred on average once every seven days in the smaller tributaries, but only once in 30 years in the downstream river. On average, variation in daily summer water temperature decreased with increasing drainage area and the frequency of extremely warm water temperatures declined exponentially with increasing drainage area (Moore et al., 2015).
In contrast to the snowmelt-driven and mountainous environment of the Fraser River, the Gila River, NM drains a semiarid landscape where a monsoon primarily controls the relatively short rainy season and subsequent flow patterns. The temporal variability of water chemistry increased with drainage area among Gila River sites (Acuña and Dahm, 2007); the authors attributed this pattern to shifts in the relative contribution of different water sources as routing changed seasonally with the expansion-contraction of the river network (Acuña and Dahm, 2007).
Intervening units
Within fluvial hydrosystems, transport of materials from a given stream or riparian wetland to a downstream water typically must pass through other intervening units (other stream segments, wetlands, lakes), which also can function as sources, sinks, lags, and transformers of materials and energy. Streams and riparian wetlands can have individual and/or aggregate effects on these intervening units and thereby indirectly affect connected-downstream waters. The degree to which intervening units mediate the effects that an upstream water ultimately has on a downstream water depends largely on the relative sizes of the upstream and the intervening units and their turnover of water and materials. (Jones, 2010; Mallard et al., 2014). For example, a large, deep lake with a long residence time will have a stronger mediating effect on downstream contributions from a small tributary than a small, shallow lake with a short residence time receiving inflow from a large tributary.
A number of other interacting factors at varying spatial scales, such as drainage basin shape, number of intervening lakes, and the position of lakes in the network, can augment or dampen the effects of upstream contributing waters (Jones, 2010). Depending upon the characteristics of the intervening unit and meteorological conditions at the time, the downstream signal from the upstream water can be amplified, dampened, or muted by the intervening unit (Reiners and Driese, 2001; Cardille et al., 2007). Intervening segments between headwater tributaries and a downstream river that flow through unconstrained valleys will interact more with adjacent lateral and subsurface areas than intervening segments that flow through constrained valleys where connectivity will be more longitudinally focused (Montgomery, 1999; Poole, 2002).
Distinctiveness
Differences in the character of the physical or chemical material being transported from a stream or riparian wetland relative to that in the mainstem are also important in determining whether there are discernible effects. It is more difficult to identify an effect when the material or parameter is uniform between streams or wetlands and the mainstem, versus when there are distinct material or parameter difference between fluvial hydrosystem units. For example, researchers were able to determine, based on distinct geochemical signatures across the river network, that streams draining the upper 54% of the River Dee watershed in Scotland contributed 71% of the downstream base flow. Despite the upper watershed receiving only 58% of total annual precipitation, researchers concluded that long residence time ground-water flow paths from the headwater watersheds were also important in maintaining the downstream base flows (Tetzlaff and Soulsby, 2008). In the Mokelumne River in California’s central Sierra Nevada Mountains, differences in the underlying geology allowed researchers to characterize the relative contributions of nitrate to downstream reservoirs (Holloway et al., 1998). Researchers conducted a paired watershed comparison with two ephemeral streams in adjacent watersheds underlain by different rock types (diorite vs. biotite schist) but with similar land use, vegetation, topography, and watershed area. Many samples from the diorite watershed had nitrate concentrations below detection limits (<4 μM), with a median concentration of 3.3 μM; concentrations were not strongly associated with the start or end of the high precipitation period. In the biotite schist watershed, maximum stream concentrations of nitrate (>300 μM) occurred at the start of the high precipitation period, and concentrations decreased over time. A nearby perennial stream, also in a biotite schist watershed, displayed this same temporal trend, with highest nitrate concentrations at the beginning of the rainy season and decreasing concentrations during the spring. Holloway et al. (1998) concluded that biotite schist streams contributed a disproportionately large amount of total nitrate to downstream reservoirs, despite draining only a small area of the entire watershed. This example also illustrates that the effect on downstream waters can vary over time.
Timing
Because connectivity between fluvial hydrosystem units and functions within units can vary over time, effects on downstream waters can also exhibit temporal variability. Storm flows in a tropical stream network that are common during the rainy season result in a greater than 40% increase in longitudinal connectivity (flow in ephemeral streams) and an exponential increase in the downstream transport of sediment and phosphorus (Zimmermann et al., 2014). Another example of the role of timing on downstream effects is the Santa Clara River, an ephemeral river in southern California. The river flows into the Pacific Ocean at the Santa Barbara Channel, which receives water from the cooler, nutrient-rich California Current; the warmer, nutrient-poor Southern California Countercurrent; and discharge from ephemeral rivers. Water chemistry data (nitrate, phosphorus and silicate) from the river and river plume in the Santa Barbara Channel during base flow and storm events indicated that the ephemeral river is an important source of nutrients to coastal waters, extending at least 20 km offshore (Warrick et al., 2005). Although ocean upwelling is 2-4 orders of magnitude larger of a nutrient source to Santa Barbara Channel than the Santa Clara River, the timing, quality, and specific location of the river discharge make it significant. The molar ratio of river nutrients differed from that of ocean upwelling, as did the timing of delivery: nutrients from the river arrived in winter, whereas the upwelling nutrients arrived primarily in summer (Warrick et al., 2005).
The timing of landscape disturbances can also factor into the effect streams and riparian wetlands have on downstream waters. For example, the Las Conchas fire burned over 63,000 ha in the central New Mexico portion of the Río Grande drainage in the summer of 2011. A network of water quality sensors provided measures of the downstream propagation of water quality impacts to the Río Grande (Dahm et al., 2015). Water quality responses included turbidity peaks, dissolved oxygen and pH sags, and shifts in specific conductance with monsoonal storm flow that entrained high concentrations of ash and black carbon. The pulse of water quality changes associated with transport of material from the wildfire was clearly discernible >100 km downstream in the Río Grande (Dahm, et al., 2015). Floodwater storage and controlled releases from an intervening reservoir dampened some downstream effects, but flash floods coming from an ephemeral tributary that entered the Río Grande downriver from the reservoir was the major contributor (Dahm et al., 2015). Similar propagation events have been documented following storm flows preceded by wildfires and droughts (e.g., Hladyz et al., 2011; Harris et al., 2015).
SUMMARY AND IMPLICATIONS
Downstream waters, including the water and materials they carry and the organisms living in them, represent the time-integrated, cumulative longitudinal, lateral, and vertical connections with their upstream hydrosystem units, namely streams and riparian wetlands (Leibowitz et al., this issue). A substantial body of evidence unequivocally demonstrates physical and chemical connectivity between streams and riparian wetlands and their downstream waters via both structural and functional connectivity. The network of continuous channels (bed and banks) structurally connect most streams to downstream waters. The very existence of a continuous bed and bank structure makes these fluvial units physically contiguous, and provides strong geomorphologic evidence for connectivity. A stream must be linked to a larger, downstream water body by a channel for the two to have a surface water connection. Streams lacking a surface water connection to large inland or coastal waters (i.e., small endorheic basins) are the exception, as most streams are connected to larger water bodies through networks of continuous bed and banks. Floodplains and associated riparian wetlands are maintained by the recurrent inundation and deposition of materials during peak and recession flows. This structure of fluvial hydrosystems reflects the aggregate and cumulative nature of the connections between streams, riparian wetlands, and their downstream waters.
Physical and chemical connectivity is dynamic, changing with immediate, seasonal, and interannual or interdecadal (e.g., climate oscillations) conditions that affect the availability and distribution of water, materials, and mediating biota. Given this dynamism, a complete understanding of a stream’s or riparian wetland’s connections to and effects on downstream waters should aggregate connections over relatively long time scales (multiple years to decades). Although distances between streams and riparian wetlands and downstream waters vary, other factors such as relative size, density, environmental setting, distinctiveness, biology, and intervening units also influence the degree of physical and chemical connectivity with and level of consequence for downstream waters (Table 2). For example, despite being distant from downstream waters, headwater streams make up the majority of stream channels in most river networks and cumulatively supply most of the water in rivers.
Physical and chemical connectivity not only reflects the spatial configuration of hydrosystem units but also how materials move, accumulate, and transform in response to that spatial structure. The functional connectivity framework (Leibowitz et al., this issue) has strong parallels with other frameworks that describe ecological connectivity and boundaries. This review highlights the scientific body of evidence demonstrating that streams and riparian wetlands are functionally connected to downstream waters by storing, mixing, transforming, and transporting water and other materials. Streams and riparian wetlands are connected to downstream water through more than one of these functions. For example, streams simultaneously transport and transform nutrients and carbon as they cycle between mineral and organic forms while being carried downstream with stream flow. The functions by which different fluvial hydrosystem units most strongly influence their downstream waters may also differ. For example, a watershed model integrating the size and location of riparian wetlands within a north Florida drainage basin suggested that large, floodplain wetlands were most effective as hydrologic lags, mid-order wetlands as phosphorus sinks, and headwater wetlands as sediment sinks (Cohen and Brown, 2007).
Human alterations affect nearly all the factors that control the downstream effects of streams and riparian wetlands. Effect on downstream waters can be amplified, dampened, or averted by alteration of functions within and transport between units, as well as the properties of materials that are introduced and subsequently transported through the fluvial hydrosystem. For example, shifts in land practices, have resulted in higher mean annual rates of sediment, organic carbon, nitrogen and phosphorus accumulation in riparian wetlands over the last 100 years compared to the last 30 years (Craft and Casey, 2000). Coastal eutrophication and hypoxia are well-known downstream effects that have been linked to the downstream delivery of excess nutrients (Rabalais et al., 2002). In some cases, connectivity and its effects on downstream waters become more discernable with human alteration (e.g., Chin and Gregory, 2001; Wigmosta and Perkins, 2001); however, when human alterations are widespread and relatively uniform (e.g., Blann et al., 2009) attributing downstream effects to particular tributaries or parts of the river network can be more challenging.
Physical and chemical connectivity in fluvial hydrosystems is important to society. There is growing interest to better understand how changes in connectivity across landscapes affect the flow of specific goods and services used by society (Mitchell et al., 2015). Headwater streams draining the over 2 million headwater catchments (those areas in which first- and second-order streams drain portrayed by 1:100,000 scale maps) represent 74-80% of total catchment stream length across 9 different ecoregions in the United States. The combined potential economic value of the services provided by headwater catchments was estimated as $14,000 ha−1 yr−1 or $30 million yr−1 per headwater catchment (Hill et al., 2014). Water movement affects a range of services, including the provision of fresh water, regulation of water quality, and flood control. Decreasing discharge in rivers by implementing riparian buffers increases the lag, transformation, and sink of various pollutants that enhance water quality, but may decrease the quantity of water transported downriver. Likewise, reducing lateral flows to floodplain wetlands may decrease water quality and flood control but increase the quantity of water provided downstream. There is a need to understand how connectivity (and disconnectivity) of fluvial hydrosystems affects tradeoffs among various services and spatiotemporal relationships between producers and users for the management of sustainable ecosystem goods and services production (Brauman et al., 2007).
ACKNOWLEDGEMENTS
We thank the authors and reviewers of the U.S. EPA report Connectivity of Streams & Wetlands to Downstream Waters: A Review & Synthesis of the Scientific Evidence for their contributions to that report, from which this paper originates. We thank Joe Ebersole, Rose Kwok, Tammy Newcomer-Johnson, and three anonymous reviewers for helpful feedback on our manuscript. Special thanks to Katherine Loizos for preparing Figure 1. Although this work was reviewed by USEPA and approved for publication, it might not necessarily reflect official Agency policy.
LITERATURE CITED
- Acuña V and Dahm CN, 2007. Impact of Monsoonal Rains on Spatial Scaling Patterns in Water Chemistry of a Semiarid River Network. Journal of Geophysical Research 112:G04009 DOI: 10.1029/2007JG000493. [DOI] [Google Scholar]
- Albertson LK and Allen DC, 2015. Meta-analysis: Abundance, Behavior, and Hydraulic Energy Shape Biotic Effects on Sediment Transport in Streams. Ecology 96(5):1329–1339. DOI: 10.1890/13-2138.1. [DOI] [PubMed] [Google Scholar]
- Alexander LC, Fritz KM, Schofield KA, Autrey BA, DeMeester J, Golden HE, Goodrich DC, Kepner WG, Lane CR, LeDuc SD, Leibowitz SG, McManus MG, Pollard AI, Raanan Kiperwas H, Ridley CE, Vanderhoof MK, and Wigington PJ Jr., submitted Connectivity of Streams and Wetlands to Downstream Waters: An Overview. Journal of the American Water Resources Association (this issue). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander RB, Boyer EW, Smith RA, Schwarz GE, and Moore RB, 2007. The Role of Headwater Streams in Downstream Water Quality. Journal of the American Water Resources Association 43(1):41–59. DOI: 10.1111/j.1752-1688.2007.00005.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoros C and Bornette G, 2002. Connectivity and Biocomplexity in Waterbodies of Riverine Floodplains. Freshwater Biology 47(4):761–776. DOI: 10.1046/j.1365-2427.2002.00905.x. [DOI] [Google Scholar]
- Amoros C, Gilbert J, and Greenwood MT, 1996. Interactions between Units of the Fluvial Hydrosystem In: Fluvial Hydrosystems, Petts GE and Amoros C (Editors). Chapman & Hall, London, United Kingdom, pp. 184–210. DOI: 10.1007/978-94-009-1491-9_9. [DOI] [Google Scholar]
- Asano Y, Uchida T, Mimasu Y, and Ohte N, 2009. Spatial Pattern of Stream Solute Concentrations in a Steep Mountainous Catchment with a Homogeneous Landscape. Water Resources Research 45(10):W10432 DOI: 10.1029/2008WR007466. [DOI] [Google Scholar]
- Aufdenkampe AK, Mayorga E, Raymond PA, Melack JM, Doney SC, Alin SR, Aalto RE, and Yoo K, 2011. Riverine Coupling of Biogeochemical Cycles between Land, Oceans, and Atmosphere. Frontiers in Ecology and the Environment 9(1):53–60. DOI: 10.1890/100014. [DOI] [Google Scholar]
- Baker ME, Weller DE, and Jordan TE, 2007. Effects of Stream Map Resolution on Measures of Riparian Buffer Distribution and Nutrient Retention Potential. Landscape Ecology 22:973–992. DOI: 10.1007/s10980-007-9080-z. [DOI] [Google Scholar]
- Bahadur R and Samuels WB, 2015. Modeling the fate and transport of a chemical spill in the Elk River, West Virginia. Journal of Environmental Engineering 141(7):05014007 DOI: 10.1061/(ASCE)EE.1943-7870.0000930. [DOI] [Google Scholar]
- Baudry J, and Merriam HG. 1988 Connectivity and Connectedness: Functional Versus Structural Patterns in Landscapes. In: Connectivity in Landscape Ecology, Proceedings of the 2nd International Seminar of the International Association for Landscape Ecology Schreiber K-F (Editor). Müstersche Geographische Arbeiten 29:23–28. [Google Scholar]
- Bhaskar AS, Beesley L, Burns MJ, Fletcher TD, Hamel P, Oldham CE, and Roy AH, 2016. Will It Rise or Will It Fall? Managing the Complex Effects of Urbanization on Base Flow. Freshwater Science 35(1): 293–310. DOI: 10.1086/685084. [DOI] [Google Scholar]
- Battin TJ, Kaplan LA, Findlay S, Hopkinson CS, Marti E, Packman AI, Newbold JD, and Sabater F, 2009. Biophysical Controls on Organic Carbon Fluxes in Fluvial Networks. Nature Geoscience 1(2):95–100. DOI: 10.1038/ngeo101. [DOI] [Google Scholar]
- Beck AJ, Janssen F, Polerecky L, Herlory O, and de Beer D, 2009. Phototrophic Biofilm Activity and Dynamics of Diurnal Cd Cycling in a Freshwater Stream. Environmental Science & Technology 43(19):7245–7251. DOI: 10.1021/es900069y. [DOI] [PubMed] [Google Scholar]
- Benda L, Andras K, Miller D, and Bigelow P, 2004. Confluence Effects in Rivers: Interactions of Basin Scale, Network Geometry, and Disturbance Regimes. Water Resources Research 40(5):W05402 DOI: 10.1029/2003WR002583. [DOI] [Google Scholar]
- Benda L, 2008. Confluence Environments at the Scale of River Networks In: River Confluences, Tributaries and the Fluvial Network. Rice SP, Roy AG, and Rhoads BL (Editors). John Wiley & Sons, Chichester, United Kingdom, pp. 271–300. DOI: 10.1002/9780470760383.ch13. [DOI] [Google Scholar]
- Benfield EF, 1997. Comparison of Litterfall Input to Streams. Journal of the North American Benthological Society 16(1):104–108. DOI: 10.2307/1468242. [DOI] [Google Scholar]
- Blann KL, Anderson JL, Sands GR, and Vondracek B, 2009. Effects of Agricultural Drainage on Aquatic Ecosystems: A Review. Critical Reviews in Environmental Science and Technology 39(11):909–1001. DOI: 10.1080/10643380801977966. [DOI] [Google Scholar]
- Bouchard V, 2007. Export of Organic Matter from a Coastal Freshwater Wetland to Lake Erie: An Extension of the Outwelling Hypothesis. 41(1):1–7. DOI 10.1007/s10452-006-9044-4. [DOI] [Google Scholar]
- Brazner JC, Sierszen ME, Keough JR, and Tanner DK, 2000. Assessing the Ecological Importance of Coastal Wetlands in a Large Lake Context. Verhandlungen der Internationale Vereinigung für Theoretische und Angewandte Limnologie 27(4):1950–1961. DOI: 0368-0770/00/0027-01950. [Google Scholar]
- Brauman KA, Daily GC, Duarte TK, and Mooney HA, 2007. The Nature and Value of Ecosystem Services: An Overview Highlighting Hydrologic Services. Annual Review of Environment and Resources 32:67–98. DOI: 10.1146/annurev.energy.32.031306.102758. [DOI] [Google Scholar]
- Briggs MA, Gooseff MN, Arp CD, and Baker MA, 2009. A Method for Estimating Surface Transient Storage Parameters for Streams with Concurrent Hyporheic Storage. Water Resources Research 45(4):W00D27 DOI: 10.1029/2008WR006959. [DOI] [Google Scholar]
- Brinson MM, Bradshaw HD, and Kane ES, 1984. Nutrient Assimilative Capacity of an Alluvial Floodplain Swamp. Journal of Applied Ecology 21(3):1041–1057. DOI: 10.2307/2405066. [DOI] [Google Scholar]
- Brooks BW, Riley TM, and Taylor RD, 2006. Water Quality of Effluent-Dominated Ecosystems: Ecotoxicological, Hydrological, and Management Considerations. Hydrobiologia 556(1):365–379. DOI 10.1007/s10750-004-0189-7. [DOI] [Google Scholar]
- Bruns DA, Minshall GW, Cushing CE, Cummins KW, Brock JT, and Vannote RL, 1984. Tributaries as Modifiers of the River Continuum Concept: Analysis by Polar Ordination and Regression Models. Archivs für Hydrobiologie 99(2):208–220. [Google Scholar]
- Buchanan BP, Falbo K, Schneider RL, Easton ZM, and Walter MT, 2013. Hydrological Impact of Roadside Ditches in an Agricultural Watershed in Central New York: Implications for Non-point Source Pollutant Transport. Hydrological Processes 27(17):2422–2437. DOI: 10.1002/hyp.9305. [DOI] [Google Scholar]
- Bukaveckas PA, 2009. Rivers In: Encyclopedia of Inland Waters, Volume 1, Likens GE (Editor). Elsevier, Oxford, United Kingdom, pp. 721–732. DOI: 10.1016/B978-012370626-3.00248-9. [DOI] [Google Scholar]
- Bullock A, and Acreman M, 2003. The Role of Wetlands in the Hydrological Cycle. Hydrology and Earth System Sciences 7(3):358–389. [Google Scholar]
- Burkart MR, Simpkins WW, Squillace PJ, and Helmke M. 1999. Tributary Stream Infiltration as a Source of Herbicides in an Alluvial Aquifer. Journal of Environmental Quality 28(1):69–74. DOI: 10.2134/jeq1999.00472425002800010007x. [DOI] [Google Scholar]
- Butman D and Raymond PA, 2011. Significant Efflux of Carbon Dioxide from Streams and Rivers in the United States. Nature Geoscience 4(12): 839–842. DOI: 10.1038/ngeo1294. [DOI] [Google Scholar]
- Cardille JA, Carpenter SR, Coe MT, Foley JA, Hanson PC, Turner MG, and Vano JA, Carbon and Water Cycling in Lake-Rich Landscapes: Landscape Connections, Lake Hydrology, and Biogeochemistry. Journal of Geophysical Research 12:G02031 DOI: 10.1029/2006JG000200. [DOI] [Google Scholar]
- Caruso BS and Haynes J, 2011. Biophysical-regulatory Classification and Profiling of Streams across Management Units and Ecoregions. Journal of the American Water Resources Association 47(2):386–407. DOI: 10.1111/j.1752-1688.2010.00522.x. [DOI] [Google Scholar]
- Chin A, and Gregory KJ. 2001. Urbanization and Adjustment of Ephemeral Stream Channels. Annals of the Association of American Geographers 91(4):595–608. DOI: 10.1111/0004-5608.00260. [DOI] [Google Scholar]
- Church M, 2006. Bed Material Transport and the Morphology of Alluvial River Channels. Annual Review of Earth and Planetary Sciences 34:325–354. DOI: 10.1146/annurev.earth.33.092203.122721. [DOI] [Google Scholar]
- Cohen MJ and Brown MT, 2007. A Model Examining Hierarchical Wetland Networks for Watershed Stormwater Management. Ecological Modelling 201(2):179–193. DOI: 10.1016/j.ecolmodel.2006.09.029. [DOI] [Google Scholar]
- Cole JJ, Prairie YT, Caraco NF, McDowell WH, Tranvik LJ, Striegl RG, Duarte CM, Kortelainen P, Downing JA, Middelburg JJ, and Melack J, 2007. Plumbing the Global Carbon Cycle: Integrating Inland Waters into the Terrestrial Carbon Budget. Ecosystems 10(1):171–184. DOI: 10.1007/s10021-006-9013-8. [DOI] [Google Scholar]
- Constantz J, Stewart AE, Niswonger R, and Sarma L, 2002. Analysis of Temperature Profiles for Investigating Stream Losses beneath Ephemeral Channels. Water Resources Research 38(12):1316 DOI: 10.1029/2001WR001221. [DOI] [Google Scholar]
- Cooper MJ, Steinman AD, and Uzarski DG, 2013. Influence of Geomorphic Setting on the Metabolism of Lake Huron Fringing Wetlands. Limnology and Oceanography 58(2):452–464. DOI: 10.4319/lo.2013.58.2.0452. [DOI] [Google Scholar]
- Coppus R and Imeson AC. 2002. Extreme Events Controlling Erosion and Sediment Transport in a Semi-Arid Sub-Andean Valley. Earth Surface Processes and Landforms 27(13):1365–1375. DOI: 10.1002/esp.435. [DOI] [Google Scholar]
- Covino T 2017. Hydrologic Connectivity as a Framework for Understanding Biogeochemical Flux Through Watersheds and Along Fluvial Networks. Geomorphology 277:133–144. DOI: 10.1016/j.geomorph.2016.09.030. [DOI] [Google Scholar]
- Craft C and Casey W, 2000. Sediment and Nutrient Accumulation in Floodplain and Depressional Freshwater Wetlands of Georgia, USA. Wetlands 20(2):323–332. DOI: 10.1672/0277-5212(2000)020[0323:SANAIF]2.0.CO;2. [DOI] [Google Scholar]
- Dages C, Samouëlian A, Negro S, Huttel O, and Volz M, 2015. Seepage Patterns of Diuron in a Ditch Bed during a Sequence of Flood Events. Science of the Total Environment 537:120–128. DOI: 10.1016/j.scitotenv.2015.07.130. [DOI] [PubMed] [Google Scholar]
- Dahm CN, Cleaverly JR, Allred Coonrod JE, Thibault JR, McDonnell DE, and Gilroy DJ. 2002. Evapotranspiration at the Land/Water Interface in a Semi-Arid Drainage Basin. Freshwater Biology 47(4):831–843. DOI: 10.1046/j.1365-2427.2002.00917.x. [DOI] [Google Scholar]
- Dahm CN, Candelaria-Ley RI, Reale CS, Reale JK, and Van Horn DJ, 2015. Extreme Water Quality Degradation Following a Catastrophic Forest Fire. Freshwater Biology 60(12):2584–2599. DOI: 10.1111/fwb.12548. [DOI] [Google Scholar]
- Doak DF, Bigger D, Harding EK, Marvier MA, Malley REO, and Thomson D, 1998. The Statistical Inevitability of Stability-Diversity Relationships in Community Ecology. The American Naturalist 151(3):264–276. DOI: 10.1086/286117. [DOI] [PubMed] [Google Scholar]
- Dollhopf RH, Fitzpatrick FA, Kimble JW, Capone DM, Graan TP, Zelt RB, and Johnson R, 2014. Response to Heavy, Non-floating Oil Spilled in a Great Lakes River Environment—A Multiple-lines-of-evidence Approach for Submerged Oil Assessment and Recovery. In: Proceedings, 2014 International Oil Spill Conference, Savannah, Georgia, May 7–9, 2014, American Petroleum Institute, Washington, DC, pp. 434–448. DOI: 10.7901/2169-3358-2014.1.434. [DOI] [Google Scholar]
- Ebersole JL, Liss WJ, and Frissell CA, 2003. Cold Water Patches in Warm Streams: Physicochemical Characteristics and the Influence of Shading. Journal of the American Water Resources Association 39(2):355–368. DOI: 10.1111/j.1752-1688.2003.tb04390.x. [DOI] [Google Scholar]
- Ebersole JL, Wigington PJ, Leibowitz SG, Comeleo RL, and Sickle JV. 2015. Predicting the Occurrence of Cold-water Patches at Intermittent and Ephemeral Tributary Confluences with Warm Rivers. Freshwater Science 34(1):111–124. DOI: 10.1086/678127. [DOI] [Google Scholar]
- Ensign SH and Doyle MW, 2006. Nutrient Spiraling in Streams and River Networks. Journal of Geophysical Research: Biogeosciences 111:G04009 DOI: 10.1029/2005JG000114. [DOI] [Google Scholar]
- Fenner N and Freeman C, 2011. Drought-induced Carbon Loss in Peatlands. Nature Geoscience 4(12):895–900. DOI: 10.1038/NGEO1323. [DOI] [Google Scholar]
- Fergus CE, Lapierre J-F, Oliver SK, Skaff NK, Cheruvelil KS, Webster K, Scott C, and Soranno P. 2017. The Freshwater Landscape: Lake, Wetland, and Stream Abundance and Connectivity at Macroscales. Ecosphere 8(8):e01911 DOI: 10.1002/ecs2.1911. [DOI] [Google Scholar]
- Ferguson R and Hoey T. 2008. Effects of Tributaries on Main-channel Geomorphology In: River Confluences, Tributaries and the Fluvial Network, Rice SP, Roy AG, and Rhoads BL (Editors). John Wiley & Sons, Chichester, United Kingdom, pp. 183–208. DOI: 10.1002/9780470760383.ch10. [DOI] [Google Scholar]
- Florsheim JL, Mount JF, and Chin A, 2008. Bank Erosion as a Desirable Attribute of Rivers. BioScience 58(6):519–529. DOI: 10.1641/B580608. [DOI] [Google Scholar]
- Forshay KJ and Stanley EH, 2005. Rapid Nitrate Loss and Denitrification in a Temperate River Floodplain. Biogeochemistry 75(1):43–64. DOI 10.1007/s10533-004-6016-4. [DOI] [Google Scholar]
- Fritz KM, Gangloff MM, and Feminella JW, 2004. Habitat Modification by the Stream Macrophyte Justicia americana and Its Effects on Biota. Oecologia 140(3):388–397. DOI 10.1007/s00442-004-1594-3. [DOI] [PubMed] [Google Scholar]
- Fuller CC and Harvey JW, 2000. Reactive Uptake of Trace Metals in the Hyporheic Zone of a Mining-contaminated Stream, Pinal Creek, Arizona. Environmental Science & Technology 34(7):1150–1155. DOI: 10.1021/es990714d. [DOI] [Google Scholar]
- Gallo EL, Lohse KA, Ferlin CM, Meixner T and Brooks PD, 2014. Physical and Biological Controls on Trace Gas Fluxes in Semi-arid Urban Ephemeral Waterways. Biogeochemistry 121(1):189–207. DOI 10.1007/s10533-013-9927-0. [DOI] [Google Scholar]
- Gardner B and Sullivan PJ, 2004. Spatial and Temporal Stream Temperature Prediction: Modeling Nonstationary Temporal Covariance Structures. Water Resources Research 40(1):1–9. DOI: 10.1029/2003WR002511. [DOI] [Google Scholar]
- Gardner KK and McGlynn BL, 2009. Seasonality in Spatial Variability and Influence of Land Use/Land Cover and Watershed Characteristics on Stream Water Nitrate Concentrations in a Developing Watershed in the Rocky Mountain West. Water Resources Research 45(8):W08411 DOI: 10.1029/2008WR007029. [DOI] [Google Scholar]
- Golden HE and Knightes CD, 2011. Simulated Watershed Mercury and Nitrate Flux Responses to Multiple Land Cover Conversion Scenarios. Environmental Toxicology and Chemistry 30(4):773–786. DOI: 10.1002/etc.449. [DOI] [PubMed] [Google Scholar]
- Gomi T and Sidle RC, 2003. Bed Load Transport in Managed Steep-gradient Headwater Streams of Southeastern Alaska. Water Resources Research 39(12):1336 DOI: 10.1029/2003WR002440. [DOI] [Google Scholar]
- Gooderham JPR, Barmuta LA, and Davis PE. 2007. Upstream Heterogeneous Zones: Small Stream Systems Structured by a Lack of Competence? Journal of the North American Benthological Society 26(3):365–374. DOI: 10.1899/06-067.1. [DOI] [Google Scholar]
- Gordon J, Arbeider M, Scott D, Wilson SM, and Moore JW, 2015. When the Tides Don’t Turn: Floodgates and Hypoxic Zones in the Lower Fraser River, British Columbia, Canada. Estuaries and Coasts 38(6):2337–2344. DOI: 10.1007/s12237-014-9938-7. [DOI] [Google Scholar]
- Graf WL, 1994. Plutonium and the Rio Grande: Environmental Change and Contamination in the Nuclear Age. Oxford University Press, New York, ISBN-13: 978-0195089332. [Google Scholar]
- Gregory KJ, 2006. The Human Role in Changing River Channels. Geomorphology 79(3-4):172–191. DOI: 10.1016/j.geomorph.2006.06.018. [DOI] [Google Scholar]
- Grimshaw DL, and Lewin J, 1980. Source Identification for Suspended Sediments. Journal of Hydrology 47(1-2):151–162. DOI: 10.1016/0022-1694(80)90053-0. [DOI] [Google Scholar]
- Gutiérrez JL and Jones CG, 2006. Physical Ecosystem Engineers as Agents of Biogeochemical Heterogeneity. BioScience 56(3):227–236. DOI: 10.1641/0006-3568(2006)056[0227:PEEAAO]2.0.CO;2. [DOI] [Google Scholar]
- Hamilton SK, Bunn SE, Thoms MC, and Marshall JC, 2005. Persistence of Aquatic Refugia between Flow Pulses in a Dryland River System (Cooper Creek, Australia). Limnology and Oceanography 50(3):743–754. DOI: 10.4319/lo.2005.50.3.0743. [DOI] [Google Scholar]
- Harris HE, Baxter CV, and Davis JM, 2015. Debris Flows Amplify Effects of Wildfire on Magnitude and Composition of Tributary Subsidies to Mainstem Habitats. Freshwater Science 34(4):1457–1467. DOI: 10.1086/684015. [DOI] [Google Scholar]
- Harrison MD, Miller AJ, Groffman PM, Mayer PM, and Kaushal SS, 2014. Hydrologic Controls on Nitrogen and Phosphorous Dynamics in Relict Oxbow Wetlands Adjacent to an Urban Restored Stream. Journal of the American Water Resources Association 50(6):1365–1382. DOI: 10.1111/jawr.12193. [DOI] [Google Scholar]
- Harvey JW, and Fuller CC. 1998. Effect of Enhanced Manganese Oxidation in the Hyporheic Zone on Basin-scale Geochemical Mass Balance. Water Resources Research 34(4):623–636. DOI: 10.1029/97WR03606. [DOI] [Google Scholar]
- Harvey JW, Saiers JE, and Newlin JT, 2005. Solute Transport and Storage Mechanisms in Wetlands of the Everglades, South Florida. Water Resources Research 41(5):W05009 DOI: 10.1029/2004WR003507. [DOI] [Google Scholar]
- Hassan MA, Gottesfeld AS, Montgomery DR, Tunnicliffe JF, Clarke GKC, Wynn G, Jones-Cox H, Poirier R, MacIsaac E, Herunter H, and Macdonald SJ, 2008. Salmon-driven Bed Load Transport and Bed Morphology in Mountain Streams. Geophysical Research Letters 35(4):L04405 DOI: 10.1029/2007GL032997. [DOI] [Google Scholar]
- Hill BH, Kolka RK, McCormick FH, and Starry MA, 2014. A Synoptic Survey of Ecosystem Services from Headwater Catchments in the United States. Ecosystem Services 7:106–115. DOI: 10.1016/j.ecoser.2013.12.004. [DOI] [Google Scholar]
- Hladyz S, Watkins SC, Whitworth KL, and Baldwin DS, 2011. Flows and Hypoxic Blackwater Events in Managed Ephemeral River Channels. Journal of Hydrology 401(1-2):117–125. DOI: 10.1016/j.jhydrol.2011.02.014. [DOI] [Google Scholar]
- Hoellein TJ, Arango CP, and Zak Y, 2011. Spatial Variability in Nutrient Concentration and Biofilm Nutrient Limitation in an Urban Watershed. Biogeochemistry 106(2):265–280. DOI: 10.1007/s10533-011-9631-x. [DOI] [Google Scholar]
- Holloway JM, Dahlgren RA, Hansen B, and Casey WH, 1998. Contribution of Bedrock Nitrogen to High Nitrate Concentrations in Stream Water. Nature 395(6704):785–788. DOI: 10.1038/27410. [DOI] [Google Scholar]
- Hornberger MI, Luoma SN, Johnson ML, and Holyoak M. 2009. Influence of Remediation in a Mine-impacted River: Metal Trends over Large Spatial and Temporal Scales. Ecological Applications 19(6):1522–1535. DOI: 10.1890/08-1529.1. [DOI] [PubMed] [Google Scholar]
- Huang HQ, Chang HH, and Nanson GC, 2004. Minimum Energy as the General Form of Critical Flow and Maximum Flow Efficiency and for Explaining Variations in River Channel Pattern. Water Resources Research 40(4):W04502 DOI: 10.1029/2003WR002539. [DOI] [Google Scholar]
- Hudson PF, Heitmuller FT, and Leitch MB, 2012. Hydrologic Connectivity of Oxbow Lakes along the Guadualupe River, Texas: The Influence of Geomorphic and Climatic Controls on the “Flood Pulse Concept.” Journal of Hydrology 414-415:174–183. DOI: 10.1016/j.jhydrol.2011.10.029. [DOI] [Google Scholar]
- Johnson BR, Haas A, and Fritz KM, 2010. Use of Spatially Explicit Physicochemical Data to Measure Downstream Impacts of Headwater Stream Disturbance. Water Resources Research 46(9):W09526 DOI: 10.1029/2009WR008417. [DOI] [Google Scholar]
- Jones NE, 2010. Incorporating Lakes within the River Discontinuum: Longitudinal Changes in Ecological Characteristics in Stream–Lake Networks. Canadian Journal of Fisheries and Aquatic Sciences 67(8):1350–1362. DOI: 10.1139/F10-069. [DOI] [Google Scholar]
- Jung M, Burt TP, and Bates PD. 2004. Toward a Conceptual Model of Floodplain Water Table Response. Water Resources Research 40(12):W12409 DOI: 10.1029/2003WR002619. [DOI] [Google Scholar]
- Kaplan LA, Wiegner TN, Newbold JD, Ostrom PH, and Gandhi H, 2008. Untangling the Complex Issue of Dissolved Organic Carbon Uptake: A Stable Isotope Approach. Freshwater Biology 53(5):855–864. DOI: 10.1111/j.1365-2427.2007.01941.x. [DOI] [Google Scholar]
- Karim F, Kinsey-Henderson A, Wallace J, Arthington AH, and Pearson RG, 2012. Modelling Wetland Connectivity during Overbank Flooding in a Tropical Floodplain in North Queensland, Australia. Hydrological Processes 26(18):2710–2723. DOI: 10.1002/hyp.8364. [DOI] [Google Scholar]
- Katz TK, Webster KE, Bowser CJ, Magnuson JJ, and Benson BJ. 1997. The Influence of Landscape Position on Lakes in Northern Wisconsin. Freshwater Biology 37(1):209–217. DOI: 10.1046/j.1365-2427.1997.00149.x [DOI] [Google Scholar]
- Keizer FM, Schot PP, Okruszko T, Chormański J, Kardel I, and Wassen MJ, 2014. A New Look at the Flood Pulse Concept: The (Ir)relevance of the Moving Littoral in Temperate Zone Rivers. Ecological Engineering 64:85–99. DOI: 10.1016/j.ecoleng.2013.12.031. [DOI] [Google Scholar]
- Kleiss BA, 1996. Sediment Retention in a Bottomland Hardwood Wetland in Eastern Arkansas. Wetlands 16(3):321–33. DOI: 10.1007/BF03161323. [DOI] [Google Scholar]
- Knightes CD, Sunderland EM, Craig Barber M, Johnston JM, and Ambrose RB, 2009. Application of Ecosystem-scale Fate and Bioaccumulation Models to Predict Fish Mercury Response Times to Changes in Atmospheric Deposition. Environmental Toxicology and Chemistry 28(4):881–893. DOI: 10.1897/08-242R.1. [DOI] [PubMed] [Google Scholar]
- Knighton AD, 1998. Fluvial Forms and Processes: A New Perspective. Hodder Arnold, London, United Kingdom: ISBN-13: 978-0340663134. [Google Scholar]
- Kondolf GM, Maloney LM, and Williams JG, 1987. Effects of Bank Storage and Well Pumping on Base Flow, Carmel River, Monterey County, California. Journal of Hydrology 91(3-4):351–369. DOI: 10.1016/0022-1694(87)90211-3. [DOI] [Google Scholar]
- Lamberti GA, Chaloner DT, and Hershey AE, 2010. Linkages Among Aquatic Ecosystems. Journal of the North American Benthological Society 29(1):245–263. DOI: 10.1899/08-166.1. [DOI] [Google Scholar]
- Lane CR, Leibowitz SG, Autrey BC, LeDuc SD, and Alexander LC. Submitted Hydrological, Physical, and Chemical Functions and Connectivity of Non-floodplain Wetlands to Downstream Waters: A Review. Journal of the American Water Resources Association (this issue). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lane EW, 1955. The Importance of Fluvial Morphology in Hydraulic Engineering. Proceedings of the American Society of Civil Engineers 81(745):1–17. [Google Scholar]
- Laronne JB and Reid I, 1993. Very High Rates of Bedload Sediment Transport by Ephemeral Desert Rivers. Nature 366(6451):148–150. DOI: 10.1038/366148a0. [DOI] [Google Scholar]
- Leibowitz SG, Wigington PJ, Rains MC, and Downing DM, 2008. Non-navigable Streams and Adjacent Wetlands: Addressing Science Needs Following the Supreme Court’s Rapanos Decision. Frontiers in Ecology and the Environment 6(7):364–371. DOI: 10.1890/070068. [DOI] [Google Scholar]
- Leibowitz SG, Wigington PJ Jr., Schofield KA, Alexander LC, Vanderhoof MK, and Golden HE, Submitted Connectivity of Streams and Wetlands to Downstream Waters: An Integrated Systems Framework. Journal of the American Water Resources Association (this issue). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerner DN, 1986. Leaking Pipes Recharge Ground Water. Ground Water 24(5):654–662. DOI: 10.1111/j.1745-6584.1986.tb03714.x. [DOI] [Google Scholar]
- Luthy RG, Sedlak DL, Plumlee MH, Austin D, and Resh VH, 2015. Wastewater-Effluent-Dominated Streams as Ecosystem-Management Tools in a Drier Climate. Frontiers in Ecology and the Environment 13(9):477–485. DOI: 10.1890/150038. [DOI] [Google Scholar]
- Mallard J, McGlynn B, and Covino T, 2014. Lateral Inflows, Stream-Groundwater Exchange, and Network Geometry Influence Stream Water Composition. Water Resources Research 50(6):4603–4623. DOI: 10.1002/2013WR014944. [DOI] [Google Scholar]
- McGuire KJ, Torgersen CE, Likens GE, Buso DC, Lowe WH, and Bailey SW, 2014. Network Analysis Reveals Multiscale Controls on Streamwater Chemistry. Proceedings of the National Academy of Sciences 111(19):7030–7035. DOI: 10.1073/pnas.1404820111 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meador MR, 1996. Water Transfer Projects and the Role of Fisheries Biologists. Fisheries 21(9):18–23. DOI: 10.1577/1548-8446(1996)021<0018:WTPATR>2.0.CO;2. [DOI] [Google Scholar]
- Menichino GT and Hester ET. 2015. The Effect of Macropores on Bi-directional Hydrologic Exchange between a Stream Channel and Riparian Groundwater. Journal of Hydrology 529(3):830–842. DOI: 10.1016/j.jhydrol.2015.09.005. [DOI] [Google Scholar]
- Mertes LAK, 1997. Documentation and Significance of the Perirheic Zone on Inundated Floodplains. Water Resources Research 33(7):1749–1762. DOI: 10.1029/97WR00658. [DOI] [Google Scholar]
- Miller D, Luce C, and Benda L, 2003. Time, Space, and Episodicity of Physical Disturbance in Streams. Forest Ecology and Management 178(1-2):121–140. DOI: 10.1016/S0378-1127(03)00057-4. [DOI] [Google Scholar]
- Minshall GW, Petersen RC, Cummins KW, Bott TL, Sedell JR, Cushing CE, and Vannote RL, 1983. Interbiome Comparison of Stream Ecosystem Dynamics. Ecological Monographs 53(1):1–25. DOI: 10.2307/1942585. [DOI] [Google Scholar]
- Mitchell MGE, Suarez-Castro AF, Martinez-Harms M, Maron M, McAlpine C, Gaston KJ, Johansen K, and Rhodes JR, 2015. Reframing Landscape Fragmentation’s Effects on Ecosystem Services Trends in Ecology and Evolution 30(4):190–198. DOI: 10.1016/j.tree.2015.01.011. [DOI] [PubMed] [Google Scholar]
- Molnár P and Ramírez JA, 1998. Energy Dissipation Theories and Optimal Channel Characteristics of River Networks. Water Resources Research 34(7):1809–1818. DOI: 10.1029/98WR00983. [DOI] [Google Scholar]
- Montgomery DR, 1999. Process Domains and the River Continuum. Journal of the American Water Resources Association 35(2):397–410. DOI: 10.1111/j.1752-1688.1999.tb03598.x. [DOI] [Google Scholar]
- Moore JW, 2006. Animal Ecosystem Engineers in Streams. BioScience 56(3):237–246. DOI: 10.1641/0006-3568(2006)056[0237:AEEIS]2.0.CO;2. [DOI] [Google Scholar]
- Moore JW, Beakes MP, Nesbitt HK, Yeakel JD, Patterson DA, Thompson LA, Phillis CC, Braun DC, Favaro C, Scott D, Carr-Harris C, and Atlas WI, 2015. Emergent Stability in a Large, Free-flowing Watershed. Ecology 96(2):340–347. DOI: 10.1890/14-0326.1. [DOI] [PubMed] [Google Scholar]
- Mulholland PJ, Helton AM, Poole GC, Hall RO, Hamilton SK, Peterson BJ, Tank JL, Ashkenas LR, Cooper LW, Dahm CN, Dodds WK, Findlay SEG, Gregory SV, Grimm NB, Johnson SL, McDowell WH, Meyer JL, Valett HM, Webster JR, Arango CP, Beaulieu JJ, Bernot MJ, Burgin AJ, Crenshaw CL, Johnson LT, Niederlehner BR, O’Brien JM, Potter JD, Sheibley RW, Sobota DJ, and Thomas SM, 2008. Stream Denitrification across Biomes and Its Response to Anthropogenic Nitrate Loading. Nature 452(7184):202–205. DOI: 10.1038/nature06686. [DOI] [PubMed] [Google Scholar]
- Nadeau TL and Rains MC, 2007. Hydrological Connectivity between Headwater Streams and Downstream Waters: How Science Can Inform Policy. Journal of the American Water Resources Association 43(1):118–133. DOI: 10.1111/j.1752-1688.2007.00010.x. [DOI] [Google Scholar]
- Naiman RJ and Sedell JR, 1979. Characterization of Particulate Organic Matter Transported by Some Cascade Mountain Streams. Journal of the Fisheries Research Board of Canada 36(1):17–31. DOI: 10.1139/f79-003. [DOI] [Google Scholar]
- Negrel P, Petelet-Giraud E, Barbier J, and Gautier E, 2003. Surface Water-Groundwater Interactions in an Alluvial Plain: Chemical and Isotopic Systematics. Journal of Hydrology 277(3-4):248–267. DOI: 10.1016/S0022-1694(03)00125-2. [DOI] [Google Scholar]
- Neubauer E, Köhler SJ, von der Kammer F, Laudon H, and Hofmann T. 2013. Effect of pH and Stream Order on Iron and Arsenic Speciation in Boreal Catchments. Environmental Science & Technology 47(13):7120–7128. DOI: 10.1021/es401193j. [DOI] [PubMed] [Google Scholar]
- Newbold JD, Mulholland PJ, Elwood JW, and O’Neill RV, 1982. Organic Carbon Spiralling in Stream Ecosystems. Oikos 38(3):266–272. DOI: 10.2307/3544663. [DOI] [Google Scholar]
- Newbold JD, Thomas SA, Minshall GW, Cushing CE, and Georgian T, 2005. Deposition, Benthic Residence, and Resuspension of Fine Organic Particles in a Mountain Stream. Limnology and Oceanography 50(5):1571–1580. DOI: 10.4319/lo.2005.50.5.1571. [DOI] [Google Scholar]
- Newcomer-Johnson TA, Kaushal SS, Mayer PM, Smith RM, and Sivirichi GM. 2016. Nutrient Retention in Restored Streams and Rivers: A Global Review and Synthesis. Water 8(4):116 DOI: 10.3390/w8040116. [DOI] [Google Scholar]
- Nixon SW, 1980. Between Coastal Marshes and Coastal Waters: A Review of Twenty Years of Speculation and Research on the Role of Salt Marshes in Estuarine Productivity and Water Chemistry In: Estuarine and Wetland Processes, Hamilton P, MacDonald KB (Editors). Plenum Press, New York, pp. 437–525. DOI: 10.1007/978-1-4757-5177-2_20. [DOI] [Google Scholar]
- Noe GB and Hupp CR, 2009. Retention of Riverine Sediment and Nutrient Loads by Coastal Plain Floodplains. Ecosystems 12(5):728–746. DOI: 10.1007/s10021-009-9253-5 [DOI] [Google Scholar]
- Opperman JJ, Luster R, McKenney BA, Roberts M, and Wrona Meadows A, 2010. Ecologically Functional Floodplains: Connectivity, Flow Regime, and Scale. Journal of the American Water Resources Association. 46(2):211–226. DOI: 10.1111/j.1752-1688.2010.00426.x. [DOI] [Google Scholar]
- Peterson BJ, Wollheim WM, Mulholland PJ, Webster JR, Meyer JL, Tank JL, Martí E, Bowden WB, Valett HM, Hershey AE, McDowell WH, Dodds WK, Hamilton SK, Gregory S, and Morrall DD, 2001. Control of Nitrogen Export from Watersheds by Headwater Streams. Science 292(5514):86–90. DOI: 10.1126/science.1056874. [DOI] [PubMed] [Google Scholar]
- Petts GE and Amoros C, 1996. The Fluvial Hydrosystem In: Fluvial Hydrosystems Petts GE Amoros C (Editors). Chapman & Hall, London, United Kingdom, pp. 1–12. DOI: 10.1007/978-94-009-1491-9_1. [DOI] [Google Scholar]
- Phillips JD, 2013. Hydrological Connectivity of Abandoned Channel Water Bodies on a Coastal Plain River. River Research and Applications 29(2):149–160. DOI: 10.1002/rra.1586. [DOI] [Google Scholar]
- Poole GC, 2002. Fluvial Landscape Ecology: Addressing Uniqueness within the River Discontinuum. Freshwater Biology 47(4):641–660. DOI: 10.1046/j.1365-2427.2002.00922.x. [DOI] [Google Scholar]
- Poole GC, Stanford JA, Frissell CA, and Running SW, 2002. Three-dimensional Mapping of Geomorphic Controls on Floodplain Hydrology and Connectivity from Aerial Photos. Geomorphology 48(4):329–347. DOI: 10.1016/S0169-555X(02)00078-8. [DOI] [Google Scholar]
- Rabalais NN, Turner RE, and Wiseman WJ, 2002. Gulf of Mexico Hypoxia, A.K.A. “The Dead Zone”. Annual Review of Ecology and Systematics 33:235–263. DOI: 10.1146/annurev.ecolsys.33.010802.150513. [DOI] [Google Scholar]
- Raymond PA, Hartmann J, Lauerwald R, Sobek S, McDonald C, Hoover M, Butman D, Striegl R, Mayorga E, Humborg C, Kortelainen P, Dürr H, Meybeck M, Ciais P, and Guth P, 2013. Global Carbon Dioxide Emissions from Inland Waters. Nature 503(7476):355–359. DOI: 10.1038/nature12760. [DOI] [PubMed] [Google Scholar]
- Reiners WA and Driese KL, 2001. The Propagation of Ecological Influences through Heterogeneous Environmental Space. BioScience 51(11):939–950. DOI: 10.1641/0006-3568(2001)051[0939:TPOEIT]2.0.CO;2. [DOI] [Google Scholar]
- Reiners WA and Driese KL, 2003. Transport of Energy, Information, and Material through the Biosphere. Annual Review of Environment and Resources 28:107–135. DOI: 10.1146/annurev.energy.28.050302.105452. [DOI] [Google Scholar]
- Renard KG and Keppel RV, 1966. Hydrographs of Ephemeral Streams in the Southwest. Journal of the Hydraulics Division Proceedings of the American Society of Civil Engineers 92(HY2):33–52. [Google Scholar]
- Rice J, Wuitch A, and Westerhoff P, 2013. Assessment of De Facto Wastewater Reuse across the U.S.: Trends between 1980 and 2008. Environmental Science & Technology 47(19):11099–11105. DOI: 10.1021/es402792s. [DOI] [PubMed] [Google Scholar]
- Rice S, 1998. Which Tributaries Disrupt Downstream Fining Along Gravel-Bed Rivers? Geomorphology 22(1):39–56. DOI: 10.1016/S0169-555X(97)00052-4. [DOI] [Google Scholar]
- Rice SP, 2017. Tributary Connectivity, Confluence Aggradation and Network Biodiversity. Geomorphology 277:6–16. DOI: 10.1016/j.geomorph.2016.03.027. [DOI] [Google Scholar]
- Rice S and Church M, 1998. Grain Size Along Two Gravel-bed Rivers: Statistical Variation, Spatial Pattern and Sedimentary Links. Earth Surface Processes and Landforms 23(4):345–363. DOI: 10.1002/(SICI)1096-9837(199804)23:4<345::AID-ESP850>3.0.CO;2-B. [DOI] [Google Scholar]
- Richardson DC, Kaplan LA, Newbold JD, and Aufdenkampe AK, 2009. Temporal Dynamics of Seston: A Recurring Nighttime Peak and Seasonal Shifts in Composition in a Stream Ecosystem. Limnology and Oceanography 54(1):344–354. DOI: 10.4319/lo.2009.54.1.0344. [DOI] [Google Scholar]
- Rodríguez-Iturbe I and Valdes JB, 1979. The Geomorphologic Structure of Hydrologic Response. Water Resources Research 15(6):1409–1420. DOI: 10.1029/WR015i006p01409. [DOI] [Google Scholar]
- Royer TV, David MB, and Gentry LE, 2006. Timing of Riverine Export of Nitrate and Phosphorus from Agricultural Watersheds in Illinois: Implications for Reducing Nutrient Loading to the Mississippi River. Environmental Science & Technology 40(13):4126–4131. DOI: 10.1021/es052573n. [DOI] [PubMed] [Google Scholar]
- Royer TV, Tank JL, and David MB, 2004. Transport and Fate of Nitrate in Headwater Agricultural Streams in Illinois. Journal of Environmental Quality 33(4):1296–1304. DOI: 10.2134/jeq2004.1296. [DOI] [PubMed] [Google Scholar]
- Ruhl L, Vengosh A, Dwyer GS, Hsu-Kim H, and Deonarine A, 2010. Environmental Impacts of the Coal Ash Spill in Kingston, Tennessee: An 18-Month Survey. Environmental Science & Technology 44(24):9272–9278. DOI: 10.1021/es1026739. [DOI] [PubMed] [Google Scholar]
- Saarnio S, Winiwarter W, and Leitão J, 2009. Methane Release from Wetlands and Watercourses in Europe. Atmospheric Environment 43(7):1421–1429. DOI: 10.1016/j.atmosenv.2008.04.007. [DOI] [Google Scholar]
- Saco PM and Kumar P, 2002. Kinematic Dispersion in Stream Networks 1. Coupling Hydraulics and Network Geometry. Water Resources Research 38(11):1244 DOI: 10.1029/2001WR000695. [DOI] [Google Scholar]
- Schofield KA, Alexander LC, Ridley CE, Vanderhoof MK, Fritz KM, Autrey BA, DeMeester J, Kepner WG, Lane CR, Leibowitz SG, and Pollard AI, Submitted Biota Connect Aquatic Habitats throughout Freshwater Ecosystem Mosaics. Journal of the American Water Resources Association (this issue). [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shaman J, Stieglitz M, and Burns D, 2004. Are Big Basins Just the Sum of Small Catchments? Hydrological Processes 18(16):3195–3206. DOI: 10.1002/hyp.5739. [DOI] [Google Scholar]
- Statzner B, 2012. Geomorphological Implications of Engineering Bed Sediments by Lotic Animals. Geomorphology 157–158:49–65. 10.1016/j.geomorph.2011.03.022. [DOI] [Google Scholar]
- Strauss EA, Mitchell NL, and Lamberti GA, 2002. Factors Regulating Nitrification in Aquatic Sediments: Effects of Organic Carbon, Nitrogen Availability, and pH. Canadian Journal of Fisheries and Aquatic Sciences 59(3):554–563. DOI: 10.1139/f02-032. [DOI] [Google Scholar]
- Strayer DL, Power ME, Fagan WF, Pickett STA, and Belnap J, 2003. A Classification of Ecological Boundaries. BioScience 53(8):723–729. DOI: 10.1641/0006-3568(2003)053[0723:ACOEB]2.0.CO;2. [DOI] [Google Scholar]
- Sutfin NA, Wohl EE, and Dwire KA, 2016. Banking Carbon: A Review of Organic Carbon Storage and Physical Factors Influencing Retention in Floodplains and Riparian Ecosystems. Earth Surface Processes and Landforms 41(1):38–60. DOI: 10.1002/esp.3857. [DOI] [Google Scholar]
- Tabacchi E, Correll DL, Hauer R, Pinay G, Planty-Tabacchi A-M, and Wissmar RC. 1998. Development, Maintenance and Role of Riparian Vegetation in the River Landscape. Freshwater Biology 40(3):497–516. DOI: 10.1046/j.1365-2427.1998.00381.x. [DOI] [Google Scholar]
- Tetzlaff D and Soulsby C, 2008. Sources of Baseflow in Larger Catchments– Using Tracers to Develop a Holistic Understanding of Runoff Generation. Journal of Hydrology 359(3–4):287–302. DOI: 10.1016/j.jhydrol.2008.07.008. [DOI] [Google Scholar]
- Torgersen CE, Faux RN, McIntosh BA, Poage NJ, and Norton DJ, 2001. Airborne Thermal Remote Sensing for Water Temperature Assessment in Rivers and Streams. Remote Sensing of Environment 76(3):386–398. DOI: 10.1016/S0034-4257(01)00186-9.. [DOI] [Google Scholar]
- Townsend-Small A, Pataki DE, Liu H, Li Z, Wu Q, and Thomas B, 2013. Increasing Summer River Discharge in Southern California, USA, Linked to Urbanization. Geophysical Research Letters 40(17):4643–4647. DOI: 10.1002/grl.50921. [DOI] [Google Scholar]
- Trebitz AS, 2006. Characterizing Seiche and Tide-driven Daily Water Level Fluctuations Affecting Coastal Ecosystems of the Great Lakes. Journal of Great Lakes Research 32(1):102–116. DOI: 10.3394/0380-1330(2006)32[102:CSATDW]2.0.CO;2. [DOI] [Google Scholar]
- Valett HM, Baker MA, Morrice JA, Crawford CS, Molles MC, Dahm CN, Moyer DL, Thibault JR, and Ellis LM, 2005. Biogeochemical and Metabolic Responses to the Flood Pulse in a Semiarid Floodplain. Ecology 86(1):220–234. DOI: 10.1890/03-4091. [DOI] [Google Scholar]
- Vannote RL, Minshall GW, Cummins KW, Sedell JR, and Cushing CE, 1980. The River Continuum Concept. Canadian Journal of Fisheries and Aquatic Sciences 37(1):130–137. DOI: 10.1139/f80-017. [DOI] [Google Scholar]
- Vivoni E, Bowman RS, Wyckoff RL, Jakubowski RT, and Richards KE, 2006. Analysis of a Monsoon Flood Event in an Ephemeral Tributary and Its Downstream Hydrologic Effects. Water Resources Research 42(3):W03404 DOI: 10.1029/2005WR004036. [DOI] [Google Scholar]
- Von Schiller D, Marcé R, Obrador B, Gómez-Gener L, Casas-Ruiz JP, Acuña V, Koschorreck M, 2014. Carbon Dioxide Emissions from Dry Watercourses. Inland Waters 4(4):377–382. DOI: 10.5268/IW-4.4.746. [DOI] [Google Scholar]
- Vörösmarty CJ, Fekete BM, Meybeck M, and Lammers RB, 2000. Global System of Rivers: Its Role in Organizing Continental Land Mass and Defining Land-to-Ocean Linkages. Global Biogeochemical Cycles 14(2):599–621. DOI: 10.1029/1999GB900092. [DOI] [Google Scholar]
- Walton R, Davis JE, Martin TH, and Chapman RS, 1996. Hydrology of the Black Swamp Wetlands on the Cache River, Arkansas. Wetlands 16(3):279–287. DOI: 10.1007/BF03161319. [DOI] [Google Scholar]
- Ward JV, Tockner K, Arscott DB, and Claret C. 2002. Riverine Landscape Diversity. Freshwater Biology 47(4): 517–539. DOI: 10.1046/j.1365-2427.2002.00893.x [DOI] [Google Scholar]
- Warrick JA and Milliman JD, 2003. Hyperpycnal Sediment Discharge from Semiarid Southern California Rivers: Implications for Coastal Sediment Budgets. Geology 31(9):781–784. DOI: 10.1130/G19671.1. [DOI] [Google Scholar]
- Warrick JA, Washburn L, Brzezinski MA, and Siegel DA, 2005. Nutrient Contributions to the Santa Barbara Channel, from the Ephemeral Santa Clara River. Estuarine, Coastal and Shelf Science 62(4):559–574. DOI: 10.1016/j.ecss.2004.09.033. [DOI] [Google Scholar]
- Webster JR, and Patten BC, 1979. Effects of Watershed Perturbation on Stream Potassium and Calcium Dynamics. Ecological Monographs 49(1):51–72. DOI: 10.2307/1942572. [DOI] [Google Scholar]
- Whiting PJ and Pomeranets M, 1997. A Numerical Study of Bank Storage and Its Contribution to Streamflow. Journal of Hydrology 202(1-4):121–136. DOI: 10.1016/S0022-1694(97)00064-4. [DOI] [Google Scholar]
- Wigmosta MS and Perkins WA, 2001. Simulating the Effects of Forest Roads on Watershed Hydrology In: Land Use and Watersheds: Human Influence on Hydrology and Geomorphology in Urban and Forest Areas, Wigmosta MS and Burges SJ (Editors). American Geophysical Union, Washington, DC, pp. 127–143. DOI: 10.1029/WS002p0127. [DOI] [Google Scholar]
- Withers PJA and Jarvie HP, 2008. Delivery and Cycling of Phosphorus in Rivers: A Review. Science of the Total Environment 400(1-3):379–395. DOI: 10.1016/j.scitotenv.2008.08.002. [DOI] [PubMed] [Google Scholar]
- Wohl E, Dwire K, Sutfin N, Polvi L, and Bazan R, 2012. Mechanisms of Carbon Storage in Mountainous Headwater Rivers. Nature Communications 3:1263 DOI: 10.1038/ncomms2274. [DOI] [PubMed] [Google Scholar]
- Wollheim WM, Peterson BJ, Thomas SM, Hopkinson CH, and Vorosmarty CJ, 2008. Dynamics of N Removal Over Annual Time Periods in a Suburban River Network. Journal of Geophysical Research 113:G03038 DOI: 10.1029/2007JG000660. [DOI] [Google Scholar]
- Wolman MG and Gerson R, 1978. Relative Scales of Time and Effectiveness of Climate in Watershed Geomorphology. Earth Surface Processes 3(2):189–208. DOI: 10.1002/esp.3290030207. [DOI] [Google Scholar]
- Zanetell B and Peckarsky BL, 1996. Stoneflies as Ecological Engineers - Hungry Predators Reduce Fine Sediments in Stream Beds. Freshwater Biology 36(3):569–577. DOI: 10.1046/j.1365-2427.1996.00132.x. [DOI] [Google Scholar]
- Zimmer MA, Bailey SW, McGuire KJ, and Bullen TD, 2013. Fine Scale Variations of Surface Water Chemistry in an Ephemeral to Perennial Drainage Network. Hydrological Processes 27(24):3438–3451. DOI: 10.1002/hyp.9449. [DOI] [Google Scholar]
- Zimmermann B, Zimmermann A, Turner BL, Francke T, and Elsenbeer H, 2014. Connectivity of Overland Flow by Drainage Network Expansion in a Rain Forest Catchment. Water Resources Research 50(2):1457–1473. DOI: 10.1002/2012WR012660. [DOI] [Google Scholar]