

Abstract
It has often been suggested in the popular and academic literature that the psychedelic state qualifies as a higher state of consciousness relative to the state of normal waking awareness. This article subjects this proposal to critical scrutiny, focusing on the question of what it would mean for a state of consciousness to be ‘higher’. We begin by considering the contrast between conscious contents and conscious global states. We then review the changes in conscious global state associated with psychedelic drug use, focusing on the effects of two serotonergic hallucinogens: psilocybin and lysergic acid diethylamide. Limiting our review to findings obtained from lab-based experiments and reported in peer-reviewed journals, we prioritize the more common and reliably induced effects obtained through subjective questionnaires and psychophysical measures. The findings are grouped into three broad categories (sensory perception, cognitive function, and experiences of unity) and demonstrate that although certain aspects of consciousness are improved or enhanced in the psychedelic state, many of the functional capacities that are associated with consciousness are seriously compromised. Psychedelic-induced states of consciousness are indeed remarkable in many ways, but it is inappropriate to regard them as ‘higher’ states of consciousness. The fact that psychedelics affect different aspects of consciousness in fundamentally different ways provides evidence against the unidimensional (or ‘level-based’) view of consciousness, and instead provides strong support for a multidimensional conception of conscious states. The final section of the article considers the implications of this analysis for two prominent theories of consciousness: the Global Workspace Theory and Integrated Information Theory.
Keywords: psychedelics, psilocybin, lysergic acid diethylamide (LSD), states of consciousness, levels of consciousness
Introduction
In the science of consciousness a clear distinction is typically drawn between the ‘contents of consciousness’ on the one hand and ‘states of consciousness’ on the other. Even researchers who endorse very different theories of consciousness (e.g. Global Workspace Theory versus Integrated Information Theory) assume that consciousness involves these two dimensions of analysis (Baars 1988; Dehaene and Changeux 2011; Tononi and Koch 2015). The distinction between contents and states is also increasingly used to frame attempts to measure consciousness (e.g. Casali et al. 2013), and to investigate consciousness in non-human animals and infants (e.g. Boly et al. 2013).
Although there are many unresolved questions about what precisely the contents of consciousness are, the notion itself is relatively well-understood. Particular contents differ from each other in terms of ‘what it is like’ for them to characterize one’s consciousness (Nagel 1974; Chalmers 2004). For example, what it’s like to have a certain type of visual content (e.g. seeing motion) is different from what it’s like to have a certain type of auditory content (e.g. hearing a trumpet), and each of these contents differs from what it’s like to have a certain type of cognitive content (e.g. realizing the solution to a crossword puzzle). In contrast, the notion of a conscious state is not particularly well-understood.
Roughly speaking, conscious states can be thought of as global dimensions of consciousness that modulate both the kinds of contents that can enter consciousness and the way in which those contents can be used by the organism for cognitive and behavioural control (Bayne and Hohwy 2016; Bayne et al. 2016). For example, in the state of ordinary waking awareness, a wide variety of contents can enter consciousness, and a wide variety of cognitive and behavioural capacities can be exercised. In other conscious states, however, both the range of conscious contents and the range of cognitive and behavioural capacities may be curtailed. Conditions that are often associated with changes in conscious state include post-comatose disorders of consciousness (Bruno et al. 2011; Casarotto et al. 2016), sleep and drug-induced sedation (Sarasso et al. 2015), and certain pathologies of consciousness, such as epileptic absence seizures (Gloor 1986; Blumenfeld 2005; Bayne 2011). For example, Minimally Conscious State patients can track certain features of their environment (such as the presence of motion or the semantic content of simple instructions), but they lack the capacity to engage in complex forms of cognition or behaviour, and they seem unable to entertain complex thoughts or ideas (Giacino et al. 2002).
In the literature, conscious states are usually understood in terms of distinct ‘levels of consciousness’ (Laureys 2005; Koch et al. 2016; Amico et al. 2017). However, we avoid this terminology here for it assumes that conscious states can be ordered along a single dimension, and that for any pair of conscious states, one member of that pair must be higher than the other. (As a parallel, consider temperature: If two objects have different temperatures, then one object must be hotter than the other.) However, that implication is open to various kinds of challenges. Although it is certainly true that some conscious states are intuitively lower than others (e.g. the state associated with mild sedation is intuitively lower than the state associated with ordinary waking awareness), it is doubtful whether all distinctions between conscious states can be captured in terms of a single dimension of analysis (Bayne et al. 2016; Bayne et al. 2017). Instead of viewing the relationship between conscious states in unidimensional terms (as the level-based approach does), we will argue that it ought to be construed in multidimensional terms, and that conscious states can differ from each other along multiple dimensions. Indeed, one of the central claims of this article is that although the psychedelic state is distinct from the state of ordinary waking awareness, it is neither ‘higher’ nor ‘lower’ than the state of ordinary waking awareness. In other words, we will argue that an analysis of the psychedelic state counts against the plausibility of the unidimensional view of conscious states and in favour of the multidimensional approach.
Charting the Psychedelic State of Consciousness
We turn now to the nature of the psychedelic state of consciousness, focusing on two of the most commonly used serotonergic hallucinogens: lysergic acid diethylamide (LSD) and psilocybin (found in ‘magic mushrooms’). Although there are differences between the states of consciousness associated with the ingestion of these two substances, and there are a range of other drugs known to induce altered states of consciousness, we will ignore these points here and will use ‘psychedelic state’ as a general term to refer to the paradigmatic states of consciousness associated with the consumption of psilocybin and LSD.
In this theoretical analysis, we will concentrate on reported subjective changes to an individual’s conscious experience (as measured by questionnaires), and psychophysical measures of basic perceptual and cognitive functioning. We will not consider the neural or physiological changes associated with psychedelics (Gouzoulis-Mayfrank et al. 1999; Vollenweider and Geyer 2001; Schartner et al. 2017). This decision should not be seen as questioning the value of that work. Rather our focus on the subjective reports and behavioural measures follows a long tradition of consciousness research that is motivated by the quest to understand ‘what it is like’ to be in a specific state of consciousness. Although we have sampled across a broad range of relevant psychophysical and behavioural studies, for the subjective reports we have limited ourselves to studies using the English or German (OAV) versions of the Altered States of Consciousness (ASC) rating scales adapted from Dittrich’s APZ (Abnormal Mental States) questionnaire (Dittrich 1996, 1998; Dittrich et al. 2006). We have limited our analysis to studies that employ this scale as it is well validated and has been used by multiple research groups to capture the effects of both psilocybin and LSD. Importantly, this questionnaire has also been the focus of a rigorous clustering analysis of 327 individual psilocybin sessions drawing from multiple psilocybin studies conducted over many years (Studerus et al. 2010).
There are a number of ways in which one might structure a discussion of changes in consciousness that are associated with the psychedelic state. Here, we focus on three broad aspects of consciousness for which psychedelic-induced changes are reported in the literature: (i) sensory and perceptual experience; (ii) cognitive capacities; and (iii) experiences relating to time, self and space. Let us consider these in turn and ask when it might be appropriate (or inappropriate) to label these different aspects of consciousness ‘higher’ or ‘lower’ relative to normal waking awareness.
Psychedelics and perception
The reputation of psychedelics as ‘mind-expanding’ suggests that they bring with them an expansion in the range of contents that enter consciousness. Here, we consider whether this claim is supported by the self-report and objective psychophysical findings relating to various aspects of perception.
Visual imagery and perceptual meaning
We begin by considering the impact of psychedelics on imagery. In the cluster analysis of the 327 psilocybin sessions performed by Studerus et al (2010), the two factors from the ASC questionnaire that receive the highest scores both related to visual imagery. The first was ‘Elementary Imagery’ (including items such as ‘I saw colours before me in total darkness or with closed eyes’) and the second was ‘Complex Imagery’ (with items like ‘I could see pictures from my past or fantasy extremely clearly’). Scores for these same factors were also increased after LSD consumption (Kraehenmann et al. 2017). In respect to qualitative self-report data, one study found that autobiographical memories were judged to be both ‘more visual’ and ‘more vivid’ after taking psilocybin (Carhart-Harris et al. 2012). LSD was also found to enhance dream-like imagery in a lab-based setting (Kraehenmann et al. 2017), and self-reports of hallucinogen experiences showed high semantic similarity with dream reports across large community-based self-report repositories (Sanz and Tagliazucchi 2018).
The factor ‘Changed Meaning of Percepts’ captures reports of objects in an individual’s environment as appearing more salient and personally significant than they ordinarily do. Here, participants who have taken psychedelics are more likely to endorse items such as ‘Objects around me engaged me emotionally much more than usual’ (Studerus et al. 2010). Psilocybin and LSD have also been found to enhance the subjective experience of colour (Hartman and Hollister 1963). For example, after-images are described as containing more colours, and the flicker-based generation of colour experience is said to be enhanced. These reports mirror those that are commonly found in popular writing, such as those found in Aldous Huxley's book ‘Heaven and Hell: First and most important is the experience of light. Everything seen by those who visit the mind’s antipodes is brilliantly illuminated and seems to shine from within. All colours are intensified to a pitch far beyond anything seen in the normal state, and at the same time the mind’s capacity for recognizing fine distinctions of tone and hue is notably heightened’. (1956, 89).
However, it is unclear to what extent these reports reflect objective improvements in colour perception, for laboratory-based studies have failed to find any evidence of visual improvements as the result of ingesting psychedelics. In one study, both psilocybin and LSD were found to impair objective measures of hue discrimination despite the participants’ subjective reports of enhanced colour perception (Hartman and Hollister 1963). Another study found no evidence that psilocybin was associated with increased sensitivity to stimulus contrast or brightness (Carter et al. 2004). It is an open question how the subjective increase in the richness and vividness of colour experience might be reconciled with the fact that psychedelics are not associated with any objective improvements in colour or brightness perception.
The bandwidth of consciousness
There is also evidence that the psychedelic state might be associated with an increase in the amount of sensory content that can enter consciousness at any given time. One of the most frequently studied paradigms used to measure psychedelic effects on sensory processing involves pre-pulse inhibition (PPI). PPI is defined in terms of a reduction in the natural startle reflex (measured as the magnitude of an eye-blink response in humans) that typically occurs if a startle tone stimulus is preceded by another tone (the pre-pulse). The relative reduction in this inhibitory response is widely regarded as a behavioural measure of ‘sensory gating’ (Graham 1975; Braff et al. 2001). PPI has been used extensively to assess the impact of psychedelic drugs (Gouzoulis-Mayfrank et al. 1998a; Vollenweider and Geyer 2001; Vollenweider et al. 2007; Quednow et al. 2012) as well as altered sensory processing associated with psychiatric conditions such as schizophrenia (Nuechterlein and Dawson 1984; Geyer and Braff 1987; Kumari et al. 2000). Psilocybin has been found to cause reduced PPI, which is taken as evidence of lower levels of sensory gating and an increase in the amount of sensory information that is ‘making it through’ to conscious awareness (Vollenweider and Geyer 2001; Vollenweider et al. 2007).
Another line of research supporting the idea of reduced perceptual suppression (or an ‘increased permeability’ of consciousness) comes from a series of studies involving binocular rivalry with observers reporting a greater proportion of time experiencing mixed or incomplete perceptual suppression (Carter et al. 2007). Using a very different measure of sensory sampling (saccadic eye movements), psilocybin and LSD were also both found to increase the frequency of saccadic eye-movements (Hebbard and Fischer 1966). While this study did not measure changes in sensory performance, it is clearly possible that such increases in saccade frequency could allow for a higher ‘sampling rate’ as a person views their environment. In respect to the ASC questionnaire, one of the few items relevant to this point—‘Everything around me was happening so fast that I no longer could follow what was going on’—was more likely to be endorsed after psilocybin (Studerus et al. 2010). Although these studies do not allow us to quantify the amount of sensory information that makes it into consciousness, they do provide consistent and compelling support for the claim that psychedelic drugs may increase the ‘bandwidth’ of perceptual experience.
In summary, with respect to the subjective experience of the psychedelic state, there is compelling evidence for an increase in the intensity and volume of sensory information experienced at any given time. When we consider objective measures of perceptual function, there is also good support for reduced perceptual suppression leading to a real increase in the permeability or bandwidth of consciousness. However, there is less objective support for improvements in either brightness or colour sensitivity, despite repeated self-reports suggesting that psychedelics enhance brightness and colour perception.
Psychedelics and cognition
We turn now to issues of cognitive and behavioural control. In the state of ordinary alert wakefulness, conscious contents are able to guide and control a wide-range of cognitive and behavioural operations, such as those that are implicated in working memory, attention, communication, decision-making, creativity and abstract thought. As we have noted, these capacities are invariably compromised in states of consciousness associated with brain trauma or epileptic seizures, thus undermining the ability of individuals to pursue their goals in an intelligent and flexible manner. What impact do psychedelics have on these capacities?
In general, the psychedelic state appears to be strongly associated with impairments in cognitive and behavioural control. Indeed, one of the largest clusters identified in the analysis by Studerus et al. (2010) is labelled ‘Impaired Control and Cognition’, and includes statements such as ‘I had difficulty making even the smallest decision’. However, as outlined below there are also suggestions that psychedelics can increase creativity and other aspects of cognitive function.
Memory, attention and communication
In regard to memory, one study found that after consumption of LSD participants were unimpaired on tests of declarative memory (e.g. ‘How old are you?’ ‘Who is the president?’ ‘Where are you?’) and learned associations (e.g. metal, iron, hard etc.) (Silverstein and Klee 1958). However, the same participants did show impairments in tasks involving mental control and the manipulation of new items (e.g. counting backwards, serial addition, reproducing brief passages of text and visual reproduction through drawing). Interestingly, measures of working memory—such as repeating a series of digits forwards or backwards—were unimpaired (Silverstein and Klee 1958). This sparing of working memory function is consistent with studies involving psilocybin showing slower reaction times but no reduction in accuracy during spatial working memory tasks involving the recall of spatial locations (Vollenweider et al. 1998; Carter et al. 2005).
In contrast with the variable effects of psychedelics on memory, the impact of psychedelics on attention seems to be more consistent. Using psychophysical measures, psilocybin has been found to impair measures of sustained (Quednow et al. 2012), divided (Carter et al. 2005) and covert orienting of attention (Gouzoulis-Mayfrank et al. 2002). Deshon et al. (1952) also describe difficulties in concentration as one of the disturbances that is most frequently seen in LSD.
Relatively little is known about the impact of psychedelics on communication, although early reports indicate clear communication impairments during the psychedelic state. For example, a study of the effects of LSD by Deshon et al. (1952) highlighted marked impairments in speech production seen across all subjects. One participant experienced complete blocking of speech for around 3 h. The subject was able to read when requested to do so but was unable to generate spontaneous utterances or answer questions except by gestures. This participant subsequently reported an inability to put into words the thoughts he wished to express (Deshon et al. 1952, 43). It is unclear to what extent these difficulties in comprehension are related to the deficits in memory, attention, and mental control that we have reviewed.
Creativity, insight, and abstract thinking
One cognitive domain in which psychedelics have been said to augment functionality involves creativity. Certainly, individuals in the psychedelic state often claim to have had novel insights into conceptual problems. One of the 11 clusters identified by Studerus et al. (2010) has been termed ‘Insightfulness’ and includes statements such as ‘I had very original thoughts’ and ‘I gained clarity into connections that puzzled me before’. Carhart-Harris et al. (2014) have referred to this dimension of the psychedelic state in terms of an ‘unconstrained style of thinking’. The idea that psychedelics might foster creativity is supported by an earlier line of research that examined the effects of psilocybin on semantic associations using word-priming. Comparing direct (e.g. ‘black-white’) and indirect (e.g. ‘lemon-sweet’) word pairs, psilocybin was found to significantly increase indirect but not direct semantic priming (Spitzer et al. 1996). This was interpreted as evidence that psilocybin increased the availability of remote associations, which in turn made a wider array of thoughts available to the agent. However, it should also be noted that the authors of this study suggest that the indirect priming effect might be associated with cognitive impairments, insofar as it reflects a decreased capacity to use contextual information for semantic processing (Spitzer et al. 1996, 1057).
These findings suggest that psychedelics enhance the capacity to formulate novel thoughts, but this capacity is only one aspect of creativity. A second—and no less important—aspect of creativity is the capacity to distinguish novels thoughts that are genuinely insightful from those that merely seem to have those properties (Boden 1996). We know of no evidence that psychedelics enhance this capacity, and the self-report data suggest that they actually impair it. Consider the following three items from the ASC, each of which received high ratings: ‘Everything around me was happening so fast that I no longer could follow what was going on; I had difficulty in distinguishing important from unimportant things’; and ‘Conflicts and contradictions seemed to dissolve’ received high ratings (Studerus et al. 2010). An impairment in the capacity to detect contradictions could hardly foster abstract thought given the importance that this capacity plays in the evaluation of evidence and argument. Further evidence in favour of the idea that psychedelics impair the evaluative dimension of thought is provided by Deshon et al. (1952), who describe LSD as giving rise to a ‘flight of ideas with rhyming and punning, preoccupation, irrelevant, and pedantic imitation’. (p. 37), and as leading to an ‘overgeneralized and tangential’ style of thought (p. 47). They also found that LSD impaired the ability of subjects to understand proverbs, a finding that was subsequently replicated by Silverstein and Klee (1958). Findings that psychedelics impair the evaluative dimensions of abstract thought are in line with the parallels that previous researchers have drawn between certain aspects of the psychedelic state and the psychotic state (see for review, Fischman 1983; Gouzoulis-Mayfrank et al. 1998b). Indeed, psychedelics are sometimes referred to as ‘psychotomimetics’ (psychosis-mimicking drugs), a term that reflects the view that the psychedelic state can be usefully viewed as a partial model of psychosis (Geyer and Vollenweider 2008; Halberstadt and Geyer 2013).
In summary, psychedelics seem to impair many of the central elements of cognition: decision-making, memory, attention and abstract thought. Although psychedelics appear to enhance the capacity to generate novel ideas, they also seem to impair the capacity to evaluate these ideas and distinguish those that represent genuine cognitive advances from those that do not. Based on the evidence available, it seems plausible that the increase in salience and/or amount sensory information experience (real or imagery-based) may itself be sub-optimal for higher-level functions (Spitzer et al. (1996). Finally, the deep commonalities between the ‘unconstrained’ style of thought seen in psychedelics on the one hand and that seen in psychosis, delusions and dream states on the other is itself an indication that psychedelics do not lead to a ‘cognitively optimal’ state.
Psychedelics and the experience of unity (time, space and self)
The cluster analysis performed by Studerus et al. (2010) identified a single factor—labelled ‘Experience of Unity’—that accounted for a shared increased experience of unity (dissolving of boundaries) relevant to both time and self. In light of the fact that behavioural investigations have explored the sense of unity for time and self independently, we too will begin by considering these two dimensions of unity independently before considering the eroding of boundaries more generally.
Time
One key feature of the psychedelic state is a distorted experience of time, with subjects typically reporting that time has stopped or slowed (Ludwig 1966). In respect to items in the ASC questionnaire (Dittrich 1996, 1998; Dittrich et al. 2006), participants reported high levels of agreement with statements such as ‘I experienced a touch of eternity’ and ‘I experienced past, present and future as an oneness’ (Studerus et al. 2010). Using objective measures of time perception based on interval matching or reproduction, a number of studies have shown significant impairments in both humans (Wittmann et al. 2007; Wackermann et al. 2008) and mice (Halberstadt et al. 2016) in the psychedelic state. For example, psilocybin was found to significantly shorten subjects’ reproduction of temporal intervals longer than 2.5 s and impair their ability to synchronize to inter-beat intervals longer than 2 s (Wittmann et al. 2007).
Self and space
Another consistent finding is that psychedelics alter the experience of the self (Nour et al. 2016). More specifically, subjects often experience a breakdown in the perceived boundary between themselves and their environment, a phenomenon that is often termed ‘ego-dissolution’ (Lebedev et al. 2015; Millière 2017). In the analysis by Studerus et al. (2010), ‘the experience of unity’ cluster clearly identified aspects of dissolving self-boundaries, with statements such as ‘The boundaries between myself and my surroundings seemed to blur’ and ‘It seemed to me that my environment and I were one’. Although objective measures of self-perception are yet to be used in lab-based experiments, two recent studies have replicated earlier reports of the experience of ego-dissolution after administration of psilocybin (Lebedev et al. 2015) and LSD (Tagliazucchi et al. 2016).
Interestingly, data from the ASC questionnaire appears to identify a distinction between the perception of the self/other boundary dissolving when compared with the sensation of ‘disembodiment’, which is linked to statements such as ‘It seemed as though I did not have a body anymore’ and ‘I had a feeling of being outside my body’. In contrast to the relatively strong impact of psychedelics on the experience of unity and dissolution of self-boundaries, Studerus et al. (2010) found that psilocybin was not as clearly associated with a sense of disembodiment typical of dissociative anaesthetics such as ketamine. This distinction might reflect the fact that the experience of disembodiment requires a robust sense of the boundary between oneself and one’s environment.
General boundaries
The fact that the items relating to self and time cluster together suggests that the underlying feature being captured here is a generalized weakening or dissolution of the natural boundaries and segmentation that structure perceptual experience. Indeed, certain items in the ASC questionnaire reflect a very generalized experience of unification, such as ‘Everything seemed to unify into a oneness’. At a much more basic perceptual level, Carter et al. (2004) found that psilocybin impaired the integration and grouping of low-level sensory signals required for the detection of coherent motion patterns. Using Kanizsa figures, psilocybin was also found to reduce/impair amodal completion, a process that is important for identifying the boundaries of objects and segmenting scenes within the visual environment (Kometer et al. 2011). The prevalence of synaesthesia in the psychedelic state could also be regarded as an example of inappropriate binding. One cluster identified within the ASC questionnaire included such items as ‘The shape of things seemed to change by sounds and noises’ and ‘Noises seemed to influence what I saw’ (Studerus et al. 2010).
Whether these alterations in the experience of unity involve improvements or impairments in conscious processing partially turns on contested issues concerning the nature of reality. Consider, for example, the question of how the phenomenon of ego-dissolution should be understood. Those who ascribe to a ‘no-self’ view of ultimate reality might argue that the experience of ego-dissolution isn’t an illusory experience, but instead involves a new and important insight into reality. However, even if the self is ultimately a fiction (e.g. Dennett 1992; Albahari 2006; Metzinger 2009; Bayne 2010; Letheby and Gerrans 2017), a well-functioning organism must be able to distinguish changes in its environment that are due to its own activity from changes that are due to the activity of other agents. Thus, it seems likely that a blurring of the boundaries between future and past or oneself and others will compromise cognitive and behavioural function.
Implication for Consciousness Science
We turn now from surveying the general impact of psychedelics on consciousness to the question of what lessons these data have for accounts of consciousness. In our view, the lessons to be drawn are both negative and positive.
Negatively, it is clear that the analysis of the psychedelic state puts further pressure on the idea that conscious states can be understood in terms of levels of consciousness. As we noted earlier, a central implication of the levels-based account is that distinct conscious states (S1 and S2), can be given a total ordering, such that either S1 is higher than S1 or vice-versa. However, it is clear from the previous sections that the psychedelic state is neither higher nor lower than the state of ordinary waking awareness. Although there are certain respects in which the psychedelic state is arguably ‘higher’ than the state of ordinary waking awareness (e.g. it is associated with increases in the richness and vividness of mental imagery), there are other respects in which the psychedelic state is arguably ‘lower’ than the state of ordinary waking awareness (e.g. it is associated with impairments in cognitive capacities such as attention, memory, communication and abstract reasoning). In short, although the conscious state associated with psychedelics is clearly distinct from the conscious state that characterizes ordinary waking awareness, there is no determinate sense in which one of these two states is ‘higher’ or ‘lower’ than the other (Fig. 1).
Figure 1.
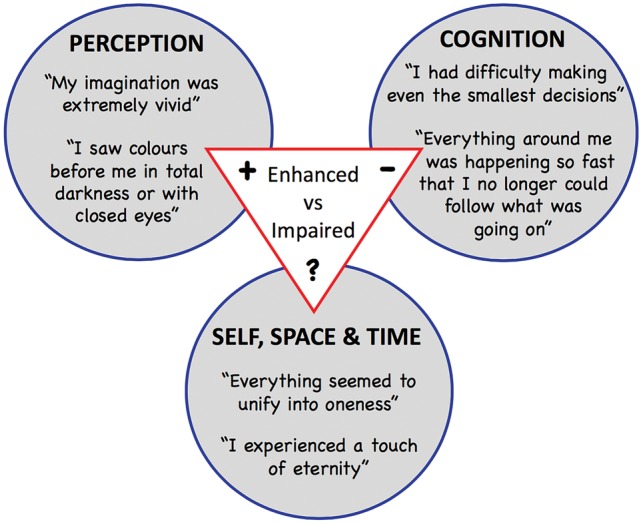
A schematic summary of the key aspects of consciousness discussed in this paper, which illustrates the inappropriateness of a unidimensional account of consciousness.
This is important for a number of reasons. Firstly, it raises questions about the recent suggestion that psychedelics might be useful in treating patients with disorders of consciousness (DoC) such as the vegetative state and the minimally conscious state (Carhart-Harris 2018). This proposal looks attractive if one assumes that the psychedelic state is ‘higher’ than the state of ordinary waking awareness in precisely the same sense that disorders of consciousness are ‘lower’ than the state of ordinary waking awareness. However, the considerations provided in this article undermine that assumption. This point, together with the fact that psychedelics impair certain aspects of cognition and have the potential to disrupt the perception of space, time and the self, raises serious questions about their therapeutic value in the context of treating DoC patients.
Second, the rejection of the levels-based account suggests that some of the most influential theories of consciousness need to be significantly revised. Consider, for example, the Global Workspace Theory (GWT) of consciousness (Baars 1988; Dehaene and Changeux 2011). Although advocates of the GWT have said little about precisely how their account captures changes in conscious state, presentations of the theory typically assume a rather simplistic conception of conscious states. For example, Dehaene et al (2017) suggest that changes in conscious state involve nothing more than ‘vigilance’ and ‘wakefulness’, and that although elucidating the brain mechanisms of vigilance and wakefulness ‘is an essential scientific goal’, ‘its computational impact [on theories of consciousness] seems minimal’ (2017, 486). These statements are puzzling, for it is clear that the changes in a creature’s conscious state have a fundamental impact on the functional role of consciousness. (Consider, for example, the way in which psychedelics alter attention, decision-making and working memory.) Dehaene et al. overlook this fact because they regard the global availability of content as an ‘essential’ feature of consciousness—that is, a property that is present when, and only when, an organism is conscious. But the multidimensional account of conscious states suggests that ‘global availability’ is not an all-or-none phenomenon, for there are conscious states in which the conscious contents are available for only some forms of high-level control. In order to be a viable theory of consciousness, GWT will need to be developed so as to accommodate the multidimensional nature of consciousness.
The data that we have reviewed also pose a challenge to the Integrated Information Theory (IIT) of consciousness, an influential complexity-based theory of consciousness. Advocates of IIT are explicitly committed to the unidimensional view of conscious states, for they equate a creature’s conscious state with its level of consciousness, and degrees of consciousness, according to IIT, are in turn understood in terms of the amount of integrated information (Φ) associated with the relevant system (e.g. Oizumi et al. 2014; Tononi et al. 2016). The considerations advanced in this paper raise questions about the plausibility of this view, for we have seen that global states cannot be ordered along a single dimension. Of course, there are algorithms that can transform highly non-linear dynamic states into values that can be ordered along a single dimension. However, this is conceptually problematic, for it is far from clear that it is coherent to describe one individual as more or less conscious than another. The psychedelic state involves a conscious state that certainly differs from the conscious state associated with ordinary waking awareness, but there is no reason to think that individuals in the psychedelic state are more conscious (or, for that matter, less conscious) than individuals who are not in it. If it is to be a viable theory of consciousness, IIT too will need to be developed so as to accommodate the multidimensional nature of consciousness.
Positively, the analysis of the psychedelic state provides an important source of data for attempts to develop a multidimensional account of conscious states. By identifying the patterns of association and dissociation that characterize the psychedelic state we can determine the state space in terms of which conscious states are structured. Why do psychedelics increase the vividness, complexity and possibly also the bandwidth of sensory experience? Why do they impair an individual’s capacities for various forms of high-level cognitive and behavioural control? What explains the systematic effects that psychedelics have on the experience of unity across a wide range of domains (e.g. time, space and the self)? Addressing these questions will help us to identify the various dimensions that structure consciousness. This task should be pursued in tandem with the investigation of other conscious states, such as those that are associated with sleep, sedation, and the post-comatose disorders of consciousness (e.g. Sergent et al. 2017). For example, we might ask whether attention qualifies as a separate dimension within this state-space, or whether changes in conscious state are better modelled by distinguishing between different aspects of attention (for example, sustained attention versus selective attention).
The study of psychedelics also provides an important—and neglected—source of data for understanding the neural basis of consciousness. We know a lot about the neurobiology of psychedelics (for a detailed review see Nichols 2016). For example, we know that psychedelics act predominantly through 5-HT2A receptor activation, and we know where these receptors are typically located in the brain. Further, through the use of animal models (Aghajanian and Marek 1999; Halberstadt and Geyer 2013) and human neuroimaging studies (Dos Santos et al. 2016; Carhart-Harris 2018), we are increasingly able to manipulate these receptors to understand the biological impact of their activation. These techniques provide us with a powerful and under-utilized avenue for understanding the neural basis of consciousness.
Although we have identified some of the more prominent changes in consciousness associated with psychedelic states, we acknowledge the limitations associated with the data we have used. The ASC represents one of the best available self-report measures of psychedelic effects and has impressive reliability and drug selectivity, but it should not be regarded as a comprehensive survey of the impact of psychedelics on consciousness. Thus, although the data surveyed here are incompatible with a unidimensional account of consciousness, they are too limited to allow for a formal assessment of the number of dimensions that would best characterise consciousness. As we gain a better understanding of the ways in which the subjective and functional dimensions of consciousness can change, so too we will gain a better understanding of the dimensional nature of consciousness, and thus better grasp what it would be like to be in each of the various states of consciousness.
Acknowledgements
We are very grateful to an anonymous referee for his/her comments.
Funding
We gratefully acknowledge the support provided by Australian Research Council Future Fellowships to Bayne (FT150100266) and Carter (FT140100807), and the support of the Canadian Institute for Advanced Research (CIFAR).
References
- Aghajanian GK, Marek GJ.. Serotonin and hallucinogens. Neuropsychopharmacology 1999;21:16S–23S. [DOI] [PubMed] [Google Scholar]
- Albahari M. Analytical Buddhism: The Two-Tiered Illusion of Self. London: Palgrave-Macmillan, 2006. [Google Scholar]
- Amico E, Marinazzo D, Di Perri C. et al. Mapping the functional connectome traits of levels of consciousness. Neuroimage 2017;148:201–11. [DOI] [PubMed] [Google Scholar]
- Baars BA. A Cognitive Theory of Consciousness. Cambridge: Cambridge University Press, 1988. [Google Scholar]
- Bayne T. The Unity of Consciousness. Oxford: OUP, 2010. [Google Scholar]
- Bayne T. The presence of consciousness in absence seizures. Behav Neurol 2011;24:47–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayne T, Hohwy J.. Modes of consciousness In: Sinnott-Armstrong W. (ed.), Finding Consciousness: The Neuroscience, Ethics and Law of Severe Brain Damage. New York: Oxford University Press, 2016, 57–80. [Google Scholar]
- Bayne T, Hohwy J, Owen AM.. Are there levels of consciousness? Trends Cogn Sci 2016;20:405–13. [DOI] [PubMed] [Google Scholar]
- Bayne T, Hohwy J, Owen AM.. Reforming the taxonomy in disorders of consciousness. Ann Neurol 2017;82:866–72. [DOI] [PubMed] [Google Scholar]
- Blumenfeld H. Consciousness and epilepsy: why are patients with absence seizures absent? Prog Brain Res 2005;150:271–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boden M. Creativity. Artificial Intelligence. Cambridge, MA: Academic Press, 1996, 267–91. [Google Scholar]
- Boly M, Seth AK, Wilke M. et al. Consciousness in humans and non-human animals: recent advances and future directions. Front Psychol 2013;4:625.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Braff DL, Geyer MA, Swerdlow NR.. Human studies of prepulse inhibition of startle: normal subjects, patient groups, and pharmacological studies. Psychopharmacology (Berl) 2001;156:234–58. [DOI] [PubMed] [Google Scholar]
- Bruno MA, Vanhaudenhuyse A, Thibaut A. et al. From unresponsive wakefulness to minimally conscious PLUS and functional locked-in syndromes: recent advances in our understanding of disorders of consciousness. J Neurol 2011;258:1373–84. [DOI] [PubMed] [Google Scholar]
- Carhart-Harris RL. The entropic brain—Revisited. Neuropharmacology 2018; doi: 10.1016/j.neuropharm.2018.03.010. [DOI] [PubMed] [Google Scholar]
- Carhart-Harris RL, Leech R, Hellyer PJ. et al. The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs. Front Hum Neurosci 2014;8:20.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carhart-Harris RL, Leech R, Williams TM. et al. Implications for psychedelic-assisted psychotherapy: functional magnetic resonance imaging study with psilocybin. Br J Psychiatry 2012;200:238–44. [DOI] [PubMed] [Google Scholar]
- Carter OL, Burr DC, Pettigrew JD. et al. Using psilocybin to investigate the relationship between attention, working memory, and the serotonin 1A and 2A receptors. J Cogn Neurosci 2005;17:1497–508. [DOI] [PubMed] [Google Scholar]
- Carter OL, Hasler F, Pettigrew JD. et al. Psilocybin links binocular rivalry switch rate to attention and subjective arousal levels in humans. Psychopharmacology (Berl) 2007;195:415–24. [DOI] [PubMed] [Google Scholar]
- Carter OL, Pettigrew JD, Burr DC. et al. Psilocybin impairs high-level but not low-level motion perception. Neuroreport 2004;15:1947–51. [DOI] [PubMed] [Google Scholar]
- Casali AG, Gosseries O, Rosanova M. et al. A theoretically based index of consciousness independent of sensory processing and behavior. Sci Transl Med 2013;5:198ra105.. [DOI] [PubMed] [Google Scholar]
- Casarotto S, Comanducci A, Rosanova M. et al. Stratification of unresponsive patients by an independently validated index of brain complexity. Ann Neurol 2016;80:718–29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chalmers D. The representational character of experience In: Leiter B. (ed.), The Future for Philosophy. Oxford: Oxford University Press, 2004, 153–81. [Google Scholar]
- Deshon H, Rinkel M, Solomon HC.. Mental changes experimentally produced by d-lysergic acid diethylamide tartrate. Psychiatr Q 1952;26:33–53. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Changeux JP.. Experimental and theoretical approaches to conscious processing. Neuron 2011;70:200–27. [DOI] [PubMed] [Google Scholar]
- Dehaene S, Lau H, Kouider S.. What is consciousness, and could machines have it? Science 2017;358:486–92. [DOI] [PubMed] [Google Scholar]
- Dennett D. The self as a center of narrative gravity In: Kessel F, Cole P, Johnson D (eds.), Self and Consciousness: Multiple Perspectives. Hillsdale, NJ: Erlbaum, 1992, 275–88. [Google Scholar]
- Dittrich A. Ätiologie-unabhängige Strukturen veränderter Wachbewusstseinszustände. Berlin, Germany: VWB, 1996. [Google Scholar]
- Dittrich A. The standardized psychometric assessment of altered states of consciousness (ASCs) in humans. Pharmacopsychiatry 1998;31:80–4. [DOI] [PubMed] [Google Scholar]
- Dittrich A, Lamparter D, Maurer M.. 5D-ABZ: Fragebogen zur Erfassung Aussergewöhnlicher Bewusstseinszustände. Eine kurze Einführung. Zurich, Switzerland: PSIN PLUS, 2006. [Google Scholar]
- Dos Santos RG, Osorio FL, Crippa JAS. et al. Classical hallucinogens and neuroimaging: A systematic review of human studies: Hallucinogens and neuroimaging. Neurosci Biobehav Rev 2016;71:715–28. [DOI] [PubMed] [Google Scholar]
- Fischman LG. Dreams, hallucinogenic drug states, and schizophrenia: a psychological and biological comparison. Schizophr Bull 1983;9:73–94. [DOI] [PubMed] [Google Scholar]
- Geyer MA, Braff DL.. Startle habituation and sensorimotor gating in schizophrenia and related animal models. Schizophr Bull 1987;13:643–68. [DOI] [PubMed] [Google Scholar]
- Geyer MA, Vollenweider FX.. Serotonin research: contributions to understanding psychoses. Trends Pharmacol Sci 2008;29:445–53. [DOI] [PubMed] [Google Scholar]
- Giacino JT, Ashwal S, Childs N. et al. The minimally conscious state: definition and diagnostic criteria. Neurology 2002;58:349–53. [DOI] [PubMed] [Google Scholar]
- Gloor P. Consciousness as a neurological concept in epileptology: a critical review. Epilepsia 1986;27:S14–26. [DOI] [PubMed] [Google Scholar]
- Gouzoulis-Mayfrank E, Heekeren K, Thelen B. et al. Effects of the hallucinogen psilocybin on habituation and prepulse inhibition of the startle reflex in humans. Behav Pharmacol 1998a;9:561–6. [DOI] [PubMed] [Google Scholar]
- Gouzoulis-Mayfrank E, Hermle L, Thelen B. et al. History, rationale and potential of human experimental hallucinogenic drug research in psychiatry. Pharmacopsychiatry 1998b;31:63–8. [DOI] [PubMed] [Google Scholar]
- Gouzoulis-Mayfrank E, Schreckenberger M, Sabri O. et al. Neurometabolic effects of psilocybin, 3,4-methylenedioxyethylamphetamine (MDE) and d-methamphetamine in healthy volunteers. A double-blind, placebo-controlled PET study with [18F]FDG. Neuropsychopharmacology 1999;20:565–81. [DOI] [PubMed] [Google Scholar]
- Gouzoulis-Mayfrank E, Thelen B, Maier S. et al. Effects of the hallucinogen psilocybin on covert orienting of visual attention in humans. Neuropsychobiology 2002;45:205–12. [DOI] [PubMed] [Google Scholar]
- Graham FK. The more or less startling effects of weak prestimulation. Psychophysiology 1975;12:238–48. [DOI] [PubMed] [Google Scholar]
- Halberstadt AL, Geyer MA.. Serotonergic hallucinogens as translational models relevant to schizophrenia. Int J Neuropsychopharmacol 2013;16:2165–80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Halberstadt AL, Sindhunata IS, Scheffers K. et al. Effect of 5-HT2A and 5-HT2C receptors on temporal discrimination by mice. Neuropharmacology 2016;107:364–75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hartman AM, Hollister LE.. Effect of mescaline, lysergic acid diethylamide and psilocybin on color perception. Psychopharmacologia 1963;4:441–51. [DOI] [PubMed] [Google Scholar]
- Hebbard FW, Fischer R.. Effect of Psilocybin, LSD, and mescaline on small, involuntary eye movements. Psychopharmacologia 1966;9:146–56. [DOI] [PubMed] [Google Scholar]
- Huxley A. Heaven and Hell. UK: Harper Collins, 1956. [Google Scholar]
- Koch C, Massimini M, Boly M, Tononi G.. Neural correlates of consciousness: progress and problems. Nat Rev Neurosci 2016;17:307–21. [DOI] [PubMed] [Google Scholar]
- Kometer M, Cahn BR, Andel D. et al. The 5-HT2A/1A agonist psilocybin disrupts modal object completion associated with visual hallucinations. Biol Psychiatry 2011;69:399–406. [DOI] [PubMed] [Google Scholar]
- Kraehenmann R, Pokorny D, Vollenweider L. et al. Dreamlike effects of LSD on waking imagery in humans depend on serotonin 2A receptor activation. Psychopharmacology (Berl) 2017;234:2031–46. [DOI] [PubMed] [Google Scholar]
- Kumari V, Soni W, Mathew VM. et al. Prepulse inhibition of the startle response in men with schizophrenia: effects of age of onset of illness, symptoms, and medication. Arch Gen Psychiatry 2000;57:609–14.10839340 [Google Scholar]
- Laureys S. The neural correlate of (un)awareness: lessons from the vegetative state. Trends Cogn Sci 2005;9:556–9. [DOI] [PubMed] [Google Scholar]
- Lebedev AV, Lovden M, Rosenthal G. et al. Finding the self by losing the self: Neural correlates of ego-dissolution under psilocybin. Hum Brain Mapp 2015;36:3137–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letheby C, Gerrans P.. Self unbound: ego dissolution in psychedelic experience. Neurosci Conscious 2017;2017:1–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ludwig AM. Altered states of consciousness. Arch Gen Psychiatry 1966;15:225–34. [DOI] [PubMed] [Google Scholar]
- Metzinger T. The Ego Tunnel. New York: Basic Books, 2009. [Google Scholar]
- Millière R. Looking for the self: phenomenology, neurophysiology and philosophical significance of drug-induced ego dissolution. Front. Hum. Neurosci 2017; doi.org/10.3389/fnhum.2017.00245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagel T. What is it like to be a bat? Philos Rev 1974;83:435–50. [Google Scholar]
- Nichols DE. Psychedelics. Pharmacol Rev 2016;68:264–355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nour MM, Evans L, Nutt D. et al. Ego-dissolution and psychedelics: validation of the ego-dissolution inventory (EDI). Front Hum Neurosci 2016;10:269.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nuechterlein KH, Dawson ME.. Information processing and attentional functioning in the developmental course of schizophrenic disorders. Schizophr Bull 1984;10:160–203. [DOI] [PubMed] [Google Scholar]
- Oizumi M, Albantakis L, Tononi G.. From the phenomenology to the mechanisms of consciousness: Integrated Information Theory 3.0. PLoS Comput Biol 2014;10:e1003588.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quednow BB, Kometer M, Geyer MA. et al. Psilocybin-induced deficits in automatic and controlled inhibition are attenuated by ketanserin in healthy human volunteers. Neuropsychopharmacology 2012;37:630–40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanz C, Tagliazucchi E.. The experience elicited by hallucinogens presents the highest similarity to dreaming within a large database of psychoactive substance reports. Front Neurosci 2018;12:7.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarasso S, Boly M, Napolitani M. et al. Consciousness and complexity during unresponsiveness induced by propofol, xenon, and ketamine. Curr Biol 2015;25:3099–105. [DOI] [PubMed] [Google Scholar]
- Schartner MM, Carhart-Harris RL, Barrett AB. et al. Increased spontaneous MEG signal diversity for psychoactive doses of ketamine, LSD and psilocybin. Sci Rep 2017;7:46421.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sergent C, Faugeras F, Rohaut B. et al. Multidimensional cognitive evaluation of patients with disorders of consciousness using EEG: a proof of concept study. Neuroimage Clin 2017;13:455–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silverstein AB, Klee GD.. Effects of lysergic acid diethylamide (LSD-25) on intellectual functions. AMA Arch Neurol Psychiatry 1958;80:477–80. [DOI] [PubMed] [Google Scholar]
- Spitzer M, Thimm M, Hermle L. et al. Increased activation of indirect semantic associations under psilocybin. Biol Psychiatry 1996;39:1055–7. [DOI] [PubMed] [Google Scholar]
- Studerus E, Gamma A, Vollenweider FX.. Psychometric evaluation of the altered states of consciousness rating scale (OAV). PLoS One 2010;5:e12412.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tagliazucchi E, Roseman L, Kaelen M. et al. Increased global functional connectivity correlates with LSD-induced ego dissolution. Curr Biol 2016;26:1043–50. [DOI] [PubMed] [Google Scholar]
- Tononi G, Boly M, Massimini M. et al. Integrated information theory: from consciousness to its physical substrate. Nat Rev Neurosci 2016;17:450–61. [DOI] [PubMed] [Google Scholar]
- Tononi G, Koch C.. Consciousness: here, there and everywhere? Philos Trans R Soc Lond B Biol Sci 2015;370: [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollenweider FX, Csomor PA, Knappe B. et al. The effects of the preferential 5-HT2A agonist psilocybin on prepulse inhibition of startle in healthy human volunteers depend on interstimulus interval. Neuropsychopharmacology 2007;32:1876–87. [DOI] [PubMed] [Google Scholar]
- Vollenweider FX, Geyer MA.. A systems model of altered consciousness: integrating natural and drug-induced psychoses. Brain Res Bull 2001;56:495–507. [DOI] [PubMed] [Google Scholar]
- Vollenweider FX, Vollenweider-Scherpenhuyzen MF, Babler A. et al. Psilocybin induces schizophrenia-like psychosis in humans via a serotonin-2 agonist action. Neuroreport 1998;9:3897–902. [DOI] [PubMed] [Google Scholar]
- Wackermann J, Wittmann M, Hasler F. et al. Effects of varied doses of psilocybin on time interval reproduction in human subjects. Neurosci Lett 2008;435:51–5. [DOI] [PubMed] [Google Scholar]
- Wittmann M, Carter O, Hasler F. et al. Effects of psilocybin on time perception and temporal control of behaviour in humans. J Psychopharmacol 2007;21:50–64. [DOI] [PubMed] [Google Scholar]