

Abstract
Protection of ecosystem services is increasingly emphasized as a risk-assessment goal, but there are wide gaps between current ecological risk-assessment endpoints and potential effects on services provided by ecosystems. The authors present a framework that links common ecotoxicological endpoints to chemical impacts on populations and communities and the ecosystem services that they provide. This framework builds on considerable advances in mechanistic effects models designed to span multiple levels of biological organization and account for various types of biological interactions and feedbacks. For illustration, the authors introduce 2 case studies that employ well-developed and validated mechanistic effects models: the inSTREAM individual-based model for fish populations and the AQUATOX ecosystem model. They also show how dynamic energy budget theory can provide a common currency for interpreting organism-level toxicity. They suggest that a framework based on mechanistic models that predict impacts on ecosystem services resulting from chemical exposure, combined with economic valuation, can provide a useful approach for informing environmental management. The authors highlight the potential benefits of using this framework as well as the challenges that will need to be addressed in future work.
Keywords: Ecological production function, Ecological risk assessment, Ecosystem service, Environmental management, Mechanistic effects model
Challenges for Ecological Risk Assessment and Management
The primary goal of ecological risk assessment (ERA) of chemicals is to provide defensible science-based support for environmental management decisions. This involves making explicit connections between impacts on the benefits derived by people from ecosystems (so-called ecosystem services 1) and the costs of managing the causes of those impacts. At the core of this approach is the need for relevant chemical exposure–response relationships. However, current ERA approaches often fall short in these regards because methods for estimating and integrating exposure and effects are often based on overly simplistic assumptions 2, 3. For example, measures of organism-level toxicity (e.g., 50% effect concentrations) are used as indicators of population-level impacts of chemicals. A primary concern is that the kinds of information collected to support ERAs are far removed from the kinds of ecological entities (e.g., species or habitats) that are the targets of protection, which themselves are often only vaguely defined in legislation (e.g., European pesticides legislation refers to “no unacceptable effects on the environment”). In practice, protection goals for ecological systems are (implicitly or explicitly) often at the population, community, or ecosystem level (e.g., persistence or abundance of a particular species, maintenance of biodiversity, and protection of ecosystem properties such as water quality), but data available for ERA typically include toxicity data from a few standard test species on organism-level endpoints (e.g., survival, reproductive output, growth). Clearly, relating results from laboratory toxicity tests to ecosystem-level consequences highlights the magnitude of the challenge for the ecological risk assessor. Although significant scientific advances are occurring in predictive ecotoxicology, such as the development of mechanistic effects models 4 and quantitative adverse outcome pathways (AOPs) 5, the principal challenge still lies in linking organismal (or suborganismal) data to ecosystem properties and processes that people care about.
Part of the problem is that ecological systems are complex and difficult to study. Ecosystems and their components (populations, communities) display nonlinear dynamics that can change in time and space. Functional redundancy at the species level means that species may be lost from a system without any obvious impacts on other ecosystem components or processes 6. Alternatively, impacts to keystone or foundation species can lead to disproportionately large changes in system properties 7. Ecosystems may also exhibit threshold behavior where incremental changes or disturbances push the system beyond a threshold and result in dramatic changes that are not easily reversible 8.
Even if we consider one of the simplest ecosystem components, that is, a population of a single species, the dynamics of such a population exposed to chemicals in natural systems can be challenging to predict. Although individual-level responses in terms of survival, reproduction, and growth are the focus of toxicity tests, changes in these responses are not directly proportional to changes in population size or growth rate because of feedback between these 2 levels of organization 9. And when other important and variable influences on population dynamics (e.g., rainfall, nutrients, temperature) are added to the mix, it should come as no surprise that simplistic estimates of chemical exposure and organism-level toxicity for a few species are not likely to provide robust predictors of risk to ecological systems. Instead, we need approaches that can incorporate these complexities, can connect across spatial and temporal scales, are based on mechanistic understanding of the relationships among system components, and provide quantitative predictions relevant to risk-management decisions 10.
The framework we introduce in the present Focus article represents an ongoing initiative supported by the National Institute of Mathematical and Biological Synthesis. The present study grew out of an investigative workshop held in 2014, “Predictive Systems Models for the Ecological Risk Assessment of Chemicals” 11. This workshop led to the formation of 2 National Institute of Mathematical and Biological Synthesis working groups 12, 13. Our working group is focused on developing dynamic models to link organism performance to ecosystem service delivery for ecological risk assessment of chemicals. It brings together population, community, and ecosystem ecologists, ecotoxicologists, and mathematicians with interest and expertise in developing dynamic, mechanistic models of complex systems to predict impacts on ecosystem processes and service delivery from data typically collected to support ERAs. Our aim is to develop example models as well as a general framework for model development, evaluation, and communication that can be applied across different ecosystem services and ecotoxicological endpoints. To leverage the increasing amounts of data being generated from high-throughput molecular (omics) technologies and their incorporation into AOPs, our working group is coordinating with a parallel group focused on developing quantitative models that link responses of organisms to chemicals at the molecular level to the organism level. Through this joint effort, we hope to create a seamless interface at the level of the organism.
In the present article, we focus on ecosystem services as assessment endpoints for ERA. We begin by describing the different kinds of models that can mechanistically link effects on organism-level endpoints typically evaluated in ERAs to impacts on ecosystem service delivery. Next, we put forward a methodological framework by which we construct explicit quantitative relationships between impacts on valued ecosystem services and the causes of those effects (i.e., impacts on service-providing units). In addition, we outline 2 case studies that are being developed as proof of concept. Finally, we summarize the challenges that will need to be addressed for the framework to be useful for informing risk-management decisions.
Ecosystem Services as ERA Protection Goals
An essential step in any ERA is the identification of an environmental value to be protected. This environmental value is defined as an assessment endpoint 14 or a specific protection goal 15. The more explicitly that this value can be expressed, and the more robustly the assessment endpoint can be related to something that can be measured, the more informative the resulting ERA will be for guiding management decisions. In the following, we argue for using ecosystem services as the entities to be protected in ERA and show how they can be quantified.
The ecosystem services definitions are from 21, 27 (not including ecological outputs, which according to 21 are those outputs from ecological production functions that can be used as ecosystem services).
Beneficiaries represent people benefitting from a final ecosystem service, e.g. anglers benefit from trout of a certain size being present in the stream, nature enthusiasts benefit from a diversity of birds around a lake, and the general public benefits from good water quality.
Ecological production functions are analogous to economic production functions and represent types, quantities and interactions of natural properties of ecological systems that generate measurable ecological outputs. Ecological production functions may represent outcomes of ecological processes that can be used for human well-being. They are the focus of environmental and management efforts and they form control points we can use to achieve environmental goals. A specific ecological production function can represent the dynamics and production of a trout population in a lake that delivers a certain number of harvestable sized fish as the final ecosystem service. The trout population is defined as a service-providing unit.
Economic production functions are models of the relationship between the input of human capital and labor and production outputs.
Final ecosystem goods and services are outputs of ecological processes that contribute to human well-being, but not all outputs are valued explicitly. Intermediate ecosystem services contribute indirectly to social welfare, whereas the final ecosystem goods and services are directly enjoyed, used or consumed. For instance, a viable macroinvertebrate community in a lake may not directly be enjoyed by the beneficiaries, but it provides a food base for recreational activities such as trout fishing or birding. Here the macroinvertebrate community represents an intermediate service, and fishing or birding is viewed as final services. Final services are more amenable to valuation, because taking both intermediate and final services into account could result in double-counting.
Total economic value is the value of the output from an economic production function (i.e., product) that includes the ecological inputs (i.e., the ecological production functions) that capture the values of beneficiaries, using stated or revealed preferences, and the inputs of labor and capital goods.
The Millennium Ecosystem Assessment 16 defined ecosystem services, broadly, as the benefits that people derive from ecosystems, including goods, services, and cultural benefits. These services are provided by abiotic entities (e.g., groundwater providing drinking water), single-species populations, and complex assemblages of species, in some cases including their interactions with the abiotic environment, although most ERAs focus on the impacts of chemicals on biota. An ecosystem services framework for ERA can be used to quantify and evaluate the effects of chemical stressors on ecological processes and entities, and there are many potential advantages to doing so. Because ecosystem services are appreciated by the public and decision makers, they are a helpful communication tool when interpreting legislative mandates by translating vague goals (e.g., protection of the environment) into quantities that can be both measured and valued by appropriate economic methods 17. An ecosystem services framework also emphasizes the importance of assessing a multiplicity of services 1 and trade-offs among them 18 that must be resolved as part of selecting a management approach.
Boyd and Banzhaf 19 and several others 20-22 have emphasized the importance of distinguishing between “final” and “intermediate” services. “Final” ecosystem goods and services are defined as components of nature, directly enjoyed, consumed, or used to yield human well-being. “Intermediate” services are ecological assets or processes that contribute to the production of final ecosystem goods and services but whose benefits to human well-being are indirect. For example, riparian ecosystems provide the intermediate services of fish habitat provision (e.g., via stream bank stabilization and stream temperature moderation) and removal of pollutants from runoff, which may contribute to final ecosystem goods and services provided by streams, such as fish desired by anglers and water suitable for human uses including aesthetic and recreational enjoyment. Both final and intermediate services are relevant to ERA, though in different ways. When assessment endpoints align with final ecosystem goods and services, social relevance of the assessment is enhanced. When models employed in risk analysis simulate intermediate services, mechanisms for risk reduction or mitigation and ecosystem restoration (e.g., riparian ecosystem restoration) become explicit.
International interest in incorporating ecosystem services into the ERA process is growing, based on the perceived benefits for regulatory decision-making. A recent White House directive to US agencies aimed “to develop and institutionalize policies to promote consideration of ecosystem services” 23. The directive highlights the preservation of benefits provided by ecosystems, the reduction of unintended negative consequences, and the promotion of cost efficiencies and investment returns. As another example, the European Food Safety Authority has developed guidance on defining environmental protection goals in relation to ecosystem services. This guidance emphasizes ecosystem services as a way to translate generalized aspirations for protection into operational and actionable goals 15. Also, the US Environmental Protection Agency (USEPA) has released 2 reports 24, 25 that describe and support the use of ecosystem services as assessment endpoints for ERA.
Nevertheless, there are significant scientific challenges to overcome when considering ecosystem services in ERA. One such challenge is in linking the data that we typically collect for ERA to the ecosystem services we want to protect. The need to test more chemicals while at the same time reducing the use of animals in ERA has spurred the development of a range of high-throughput testing methods 26. As a result, there is an even larger disconnect between the data we gather to conduct ERAs and the ecosystem services that we aim to protect. Without an appropriate assessment framework, it is unclear whether, and to what extent, exposure to chemicals that alter suborganismal processes (e.g., cell proliferation or gene expression) has effects at the whole-organism level, much less the ecosystem services that they contribute to society. In addition, the benefits that humans derive from ecosystems may be nontangible, including aesthetic, cultural, and spiritual dimensions. Even if the task of the risk assessor is confined to estimating impacts on the service-providing units and risk estimation is separated from valuation, the determination of what ecosystem services are valued, and by whom, is a substantial and occasionally controversial undertaking.
Landers and Nahlik 27 have proposed a final ecosystem goods and services classification system that aims to rigorously and systematically categorize final ecosystem goods and services and their associated beneficiaries. According to this classification system, final ecosystem goods and services are grouped according to specific environments (e.g., “rivers and streams”) and specific beneficiaries (e.g., anglers). We have adopted this classification scheme in our framework and build further on it by developing concrete metrics for each final ecosystem goods and services–beneficiary combination, which is critical to develop quantitative relationships from toxicological responses of individual organisms to impacts on populations or communities, (in some cases) intermediate services, and ultimately final ecosystem goods and services. Determining the best metrics for a particular service requires understanding of those aspects of the service that are most highly valued by the beneficiaries. The choice of metrics is also critical to account for multiple beneficiaries with different expectations. The metrics may have important space–time dimensions that are not well matched to available data, and this may be an area in which further developments are needed. The models necessary for relating changes in final ecosystem goods and services to stressors or management actions may span multiple ecosystems (such as when riparian processes impact downstream condition) and levels of biological organization (because much of the data available for assessing toxicity of chemicals is collected at organizational levels well below those needed to ensure delivery of final ecosystem goods and services).
In Table 1 we illustrate some potential metrics for final ecosystem goods and services delivered by 2 freshwater ecosystem types to different groups of beneficiaries that will be considered in our case studies. As a way of ensuring that these metrics apply to aspects of the environment that are “directly enjoyed” by human beneficiaries 19, we selected metrics previously employed in stated-preference surveys of individuals’ willingness-to-pay for or willingness-to-accept changes in environmental quality.
Table 1.
Illustration of beneficiaries and potential metrics for final ecosystem goods and services for freshwater ecosystems to be considered in our case studiesa
| Environment | Case study 1: River or stream | Case study 2: Lake |
|---|---|---|
| Selected Beneficiariesb |
Recreational anglers; people who care | Property owners; waders, swimmers, and divers; boaters; recreational anglers |
| Final ecosystem services for the selected beneficiaries |
Provisioning of recreational fishing opportunities |
Recreational experience (e.g., opportunity and conditions for wading, swimming, and/or boating); provisioning of recreational fish |
| Examples of potential metrics for the final ecosystem service, previously used in stated- preference surveys |
Abundance of game fish Abundance of game fish of catchable size Numbers of species of abundant game fish Ratio of game fish to rough fish Safety of game fish for consumption Estimated sustainability of game fish population under different use assumptions (number of days fishery is open; type of fishing equipment allowed) |
Water clarity Frequency of algal blooms Abundance of game fish Abundance of game fish of catchable size Numbers of species of abundant game fish |
Examples are based on descriptions used in published economic surveys: Some metrics may be easy to estimate using existing models; others may require development of new quantitative relationships.
”Waders, swimmers, and divers” and “Anglers” are 2 example beneficiary categories for freshwater environments as identified in the Final Ecosystem Goods and Services Classification System 27. The full list of beneficiaries for these environments also includes municipal drinking water plant operators, wastewater-treatment plant operators, residential property owners, water subsisters, food subsisters, experiencers and viewers, boaters, and “people who care” (existence and option values). References for uses of metrics: 85-93.
Next, we propose a general framework that formally incorporates ecosystem services into the ERA process. A key goal of the framework is to link data that are commonly provided in support of ERAs to the delivery of intermediate and final goods and services. This framework builds on considerable advances in mechanistic effects models that bridge levels of biological organization and account for various biological interactions and feedbacks 28, 29. An important goal of the present Focus article is to provide a framework for translating exposure and effects measured in common ecotoxicological tests into risks to ecosystem services. The ultimate goal is to provide the kind of relevant exposure–response relationships needed to inform management decisions.
An Organism-to-Ecosystem Services Conceptual Framework for ERA
In Figure 1, we depict our generalized framework for an ecosystem potentially influenced by toxicants and other drivers (natural or anthropogenic). Because the context in which ERAs are performed varies widely, because data may be available at only some levels of biological organization, and because ecosystem services may be provided by different components of ecosystems, it may not be necessary to use the entire framework in the ERA process. For example, if the ecosystem service of concern is delivered by a fish population, it may not be necessary to include a community or ecosystem model to link fish toxicity data to ecosystem service delivery. Alternatively, if delivery of an ecosystem service involves interactions of multiple species or trophic groups, it may not be necessary to develop detailed population models of each species but, rather, implement the toxicity data in an ecosystem or food-web model. Likewise, it may not always be necessary or possible to model linkages from the molecular level through the organism level to predict impacts on an ecosystem service at a higher level of biological organization.
Figure 1.
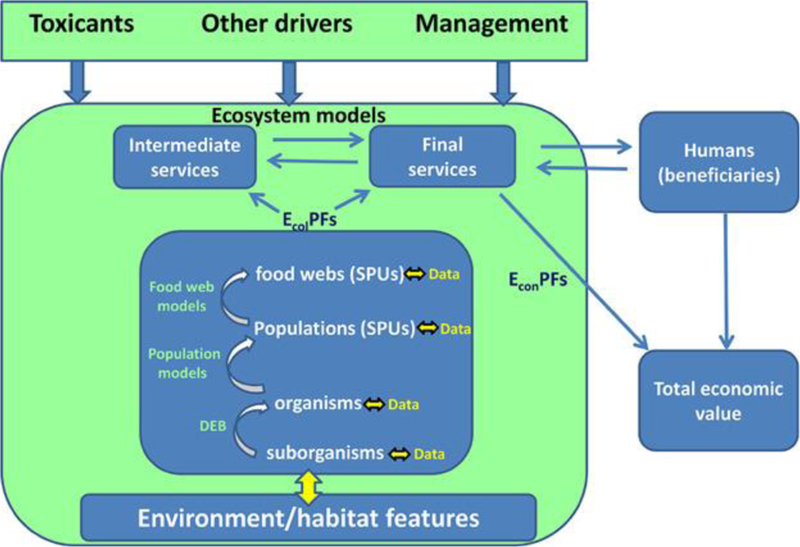
General framework for linking data that are commonly provided in support of ecological risk assessments to ecosystem services. The double arrows connecting data to each level of organization indicate that an iterative process, involving feedback between data and modeling, is employed. See text for details. DEB = dynamic energy budget; EcolPF = ecological production function; EconPF = economic production function; SPU = service providing unit.
Because our primary aim is to show how chemical effects included in ERAs can be linked to final ecosystem goods and services, we start by considering the human beneficiaries and the final ecosystem goods and services that are of concern (Figure 1, upper right). Because final ecosystem goods and services often depend on intermediate services, we indicate that relationship as well. Once the final and intermediate services are defined, it is possible to identify the service-providing units (i.e., those parts of ecosystems involved in service provision) which are typically single-species populations or multiple species interacting in food webs and sometimes interacting with the physical environment. Because our emphasis is on predicting risks of chemicals to ecological systems, we do not explicitly consider potential impacts on the physical environment but focus, rather, on the living components of the environment (i.e., living organisms and the processes in which they are involved). Quantitative relationships between the properties of service-providing units and service delivery can be modeled through ecological production functions. As defined by Bruins et al. 30, ecological production functions are quantitative or semiquantitative models of the processes by which ecosystems produce ecosystem services. Following Landers and Nahlik27, we distinguish ecological production functions from economic production functions that relate economic outputs of a production process to physical inputs relating to human capital. Total economic value is the value of the output from an economic production function (i.e., product) that includes the ecological inputs (i.e., the ecological production function) and the inputs of labor and capital goods.
In the middle box of Figure 1, depicting an idealized ecosystem, we indicate that data on the effects of toxic chemicals may be collected at different levels of biological organization and that models are needed to link responses across levels. The different types of models used to link toxicant effects from (sub)organismal levels to higher levels are described further below. Traditionally, the most common types of toxicity data used for ERAs are collected at the organismal level, and it is less common to have empirical measurements of toxicant effects on populations or food webs. Methodological advances are increasingly providing data at suborganismal levels (e.g., omics), and new modeling frameworks are being developed to link responses from the molecular to the organismal level (C.A. Murphy et al., Michigan State University, East Lansing, MI, USA, unpublished data). Validation of these various models is an important consideration and a topic of lively debate (which is beyond the scope of the present Focus article). However, using an iterative process that involves feedback between data collection and modeling in a so-called modeling cycle 31, 32 as well as efforts to increase systematic testing, documentation, and evaluation of model outputs 33-36 will help to address validation issues.
The Key Role of Mechanistic Effects Models
Mechanistic effects models include organism-level models, population models, and multispecies models (food-web and ecosystem models). The utility of mechanistic effects models in ERA was recognized in the mid-1980s and early 1990s 37, but this type of model was not adopted by risk assessors in toxic chemical regulatory programs at that time. Since then, various models describing different levels of biological organization have proved to be powerful tools for extrapolating impacts of chemicals across levels and scales 3, 38-41. Recently, key scientific and regulatory organizations have advocated the use of mechanistic effects models in ERA 42-45. An important advantage of mechanistic effects models is that, once adequately validated, they allow extrapolation to novel conditions, in contrast to most current empirical approaches.
In an ecosystem services framework, models can provide explicit mechanistic links between impacts on services and the causes of those impacts. These exposure–response relationships are essential in carrying out cost–benefit analyses and hence in choosing among management alternatives. Mechanistic effects modeling makes quantitative evaluation of the “ecological significance” (sensu; see USEPA 46) of risks possible while reducing the large uncertainty in ERAs that arises from qualitative extrapolation of laboratory toxicity test results to ecologically relevant impacts 2, 40. It also describes the causal basis of these changes in terms of chemical exposure. In the following we distinguish 3 categories of models that link suborganismal processes to organism responses, organism responses to population responses, and single-species responses to multispecies (ecosystem) responses.
Models to link physiological modes of toxicant action to whole-organism effects
Mechanistic models of organisms seek to describe the relationship between exposure (toxicokinetics) and effects on the physiology of the organism (toxicodynamics). The toxicokinetics part of the model translates the external concentration or dose of the chemical into an internal concentration at a target site, as a function of time. The toxicodynamics part then relates the internal concentration at the target site with physiological damages that may eventually lead to sublethal effects and/or death of the organism. Reviews on toxicokinetics–toxicodynamics models used in aquatic ecotoxicology describe this approach 47. Several models describe uptake/elimination processes and the physiology of the organisms with different levels of detail. For example, physiologically based toxicokinetics models focus on describing absorption, distribution, metabolism, and excretion in detail, whereas dynamic energy budget models put emphasis on describing effects of the toxicant on the major physiological functions of the organism. Overall, toxicokinetics–toxicodynamics models offer several advantages for our purpose because they allow 1) optimal extraction of information from toxicity test data; 2) extrapolation to nontested concentrations, exposure durations, and species; 3) consideration of realistic exposure scenarios and organism or population effect/recovery mechanisms; and 4) consideration of additional information available on the chemical (e.g., bioaccumulation or metabolism) and species (e.g., life history, recovery potential), thus bringing biological realism to their predictions 47.
Among existing toxicokinetics–toxicodynamics models, dynamic energy budget models have a number of attractive features. Dynamic energy budget theory was originally developed with the aim of understanding how energy allocation is modified in response to chemicals 48, 49. Dynamic energy budget theory provides a coherent scientific framework and a set of mechanistic models describing energy acquisition and allocation within organisms. It describes the full life-history characteristics (survival, growth, sexual development, reproduction, and aging) of individuals with general rules describing energy assimilation from food and energy allocation to growth, development, reproduction, and maintenance50, 51. Generic energy-allocation rules obey basic thermodynamics and rules of conservation of mass and energy and are generic across species. Expressed as a set of differential equations, representing metabolic and physiological processes in organisms, dynamic energy budget models quantify biological responses of organisms to changes in their environment, including resource availability, temperature, and exposure to chemicals52, 53.
A particularly attractive feature of dynamic energy budget models is that they facilitate interspecies extrapolation of toxicity data. Although large amounts of data are collected for standard test species, chemical toxicity data for service-providing units of interest are typically not available. Lack of methods to extrapolate toxicological and ecological data from test species to service-providing units remains 1 of the major obstacles in achieving an ecologically grounded ERA. Historically, interspecies extrapolation has been attempted using standard extrapolation or assessment factors or statistical methods (i.e., interspecies correlation models, e.g., Web-ICE 54). General rules for metabolic organization in dynamic energy budget theory and model structure are conserved across most species, and interspecies differences lie in parameter values. As a result, use of dynamic energy budget principles and models provides a more mechanistically justified approach to extrapolation of chemical effects across species and service-providing units.
Another reason for using dynamic energy budget models is to facilitate mechanistic linking of molecular or cellular effects of chemicals to higher levels of biological organization. In this context, it is helpful to identify core state variables with relevance across scales. State variables such as mass and energy reflect the most important features at each level (e.g., health status and productivity) and are conserved across levels of biological and ecological complexity. Energy production and use also underlie AOPs 55, 56. At the suborganismal level, exposure to chemicals induces changes in energy acquisition and allocation; for example, less energy is allocated to growth/reproduction, or energy is shifted to maintenance of tissues, which ensures survival. These changes involve molecular initiating events which can cause a cascade of physiological reactions and are measurable using omics approaches. These responses can affect organism performance as measured in toxicity tests via life-history traits, including survival, growth, development, and reproduction. Measurable changes in organism performance may have substantial impacts on important population-level features, such as population density, structure, and biomass. Indeed, population features are consequences of organism performance (e.g., reduced reproductive output can impact population density and biomass), but they are also a consequence of interactions between organisms and their environment. Intraspecific interactions regulate populations through competition for resources (density-dependent regulation), whereas interspecific interactions are the main channels for energy and mass transfer in multispecies systems. Both types of interactions directly or indirectly constrain individual organisms through resource availability and energy assimilation. Thus, considering energy and mass flows among organisms and across levels of biological organization has the potential to mechanistically translate impacts of chemicals at the suborganismal level to consequences for ecosystem services 57.
Models to link effects on organisms to impacts on populations
Single-species population models can provide insight into how stresses on components of individual fitness (e.g., survival, growth, reproduction) combine to affect overall individual fitness and manifest in the demography of the population as metrics such as the per capita population growth rate. Population models are classified based on how much variation (or structure) is assumed to be present in the population. For instance, scalar models treat individuals in the population as identical. Matrix models represent populations as size, age, or stage classes that differ in their vital rates; but within classes all individuals are assumed to have identical properties. They are the most-used model type in conservation biology, as evidenced by the widespread use of population viability analyses for estimating extinction risks to endangered species 9. Matrix models have often been used in ecotoxicological research because it is relatively straightforward to parameterize them with data from life-table response experiments and obtain population growth rates under a range of chemical concentrations 2, 58. However, they are not very amenable to projecting impacts of chemicals under novel environmental conditions, such as changes in organism behavior, effects of temporally or spatially heterogeneous chemical exposure, or impacts of environmental drivers such as temperature or precipitation.
Individual-based models treat every individual as unique. Individual-based models offer several advantages for our purposes in that they 1) are well suited to represent animal movements and can therefore account for spatiotemporal dynamics in resources and stressors and individual variation in exposures to chemicals; 2) can represent local interactions among individuals and adaptive behavior; 3) predict outcomes as properties that emerge from individual decisions, interactions, and responses to the environment; 4) are often designed to reproduce not just 1 but multiple patterns observed in real systems at different scales and levels of organization; and 5) are mechanistic and therefore capable of prediction under new conditions 59, 60.
For example, inSTREAM is an individual-based model for trout populations 61, 62. The model represents trout population dynamics as emerging from how individual trout select habitat, compete for food and feeding habitat, grow, survive predation and other sources of mortality, and reproduce, all in a simulated environment that varies in space and time. The model also represents several elements of the larger ecosystem: how sympatric trout species compete, the consumption of lower trophic levels (drifting invertebrates) by trout, and the consumption of trout by terrestrial predators. Key environmental drivers of inSTREAM’s simulated trout population are daily river flow and temperature and availability of invertebrate food. The model includes several important ways that fish physiology—and, hence, contaminants with sublethal effects on physiology—affect population and ecosystem dynamics. Trout behavior (selection of feeding habitat each day) and competition determine food intake, while an energy budget converts food intake, temperature, and swimming speed to daily growth. Other physiological processes in the model include fecundity as a function of fish size and energy status, egg development rates, and how sex of offspring is determined. Because inSTREAM can simulate the size (weight or length) profile of the population, the sustainability of different catch restrictions can be simulated, per Table 1.
Models to link effects on single-species to multispecies food webs and ecosystem services
Multispecies models include mathematical models of 1 or more biological communities of interacting species populations (e.g., predator–prey or food-web models with or without bioenergetics 63), process-based ecosystem models that include food-web or community models embedded within models of their physical–chemical environment (e.g., aquatic or terrestrial ecosystem models 64), and multimodels (e.g., ATLSS 65), which are a series of linked physical–chemical habitat models and multiple population models.
Whereas all of these modeling approaches could potentially be used to develop ecological production functions that link ecosystem characteristics to final ecosystem goods and services, process-based ecosystem models are a good choice because they explicitly describe relevant biological and physical–chemical processes. Consequently, they can be used to project responses under new conditions, for example, under a warmer climate. Depending on the level of complexity in a given model, ecosystem models provide a relatively complete description of a community and encompass not only the interactions within and among species but also responses to physicochemical habitat. Species populations, age/stage classes, or functional groups are most often represented as biomass compartments; and temporal dynamics are described by difference or differential equations.
Because multispecies models represent species interactions that provide intermediate ecosystem services and indirect effects of stressors, they can provide useful predictions and diagnostic mechanistic information not available from a single-population model. For example, the decrease in biomass of a Daphnia population in a lake because of a chemical stressor may result in changes in higher–trophic level fish species, which would be revealed in multispecies model simulations. An exposure–response relationship for a given chemical and population may change depending on species interactions within a food web because increasing or decreasing competition for food may interact positively or negatively with the impact of the chemical. For example, experimental removal of largemouth bass from Long Lake (Michigan, USA) had a dramatic effect on the appearance of the lake (Figure 2, top panel). Bass indirectly reduce phytoplankton (thereby increasing water clarity) by limiting smaller zooplanktivorous fish, thus causing zooplankton to increase and phytoplankton to decline. In another example, pools in Brier Creek, a prairie margin stream in south-central Oklahoma, USA, differ dramatically depending on whether largemouth and spotted bass are present (Figure 2, bottom panel). The predatory bass extirpate herbivorous minnows, promoting the growth of benthic algae. Ecosystem services depend on environmental context (habitat, food web, and ecosystem properties), which can be represented in multispecies models. Ecosystem models provide additional perspectives regarding physical-chemical habitat features because a population, or a competing population from another species, may modify the physical–chemical environment, resulting in feedbacks.
Figure 2.
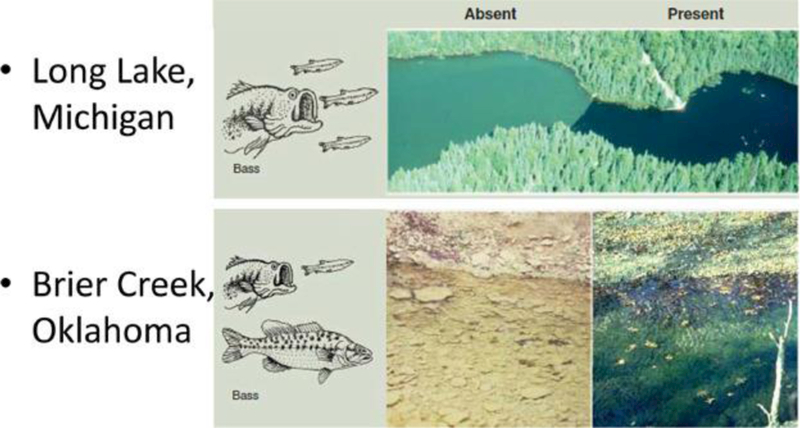
Top: Long Lake (Michigan) with largemouth bass experimentally removed (left) and present (right). Bass indirectly reduce phytoplankton (thereby increasing water clarity) by limiting smaller zooplanktivorous fish, thus causing zooplankton to increase and phytoplankton to decline 94. Bottom: Pools in Brier Creek, a prairie margin stream in south-central Oklahoma with (right) and lacking (left) largemouth and spotted bass. The predatory bass extirpate herbivorous minnows, promoting the growth of benthic algae 95. Adapted from Estes et al. 96.
Ecosystem models have a long history, particularly for lakes, streams, and other aquatic habitats 66; and many include an ecotoxicological component 40. For example, AQUATOX is a comprehensive aquatic ecosystem modeling system combining a multispecies food web, the aquatic physical–chemical habitat (stream, lake, or estuary), chemical fate processes, and a basic ecotoxicological effects submodel 67. Figure 3 illustrates a generalized food web in AQUATOX. Key outputs from AQUATOX, such as biomass of valued species, harvestable biomass, and water quality indicators like Secchi depth, chlorophyll a, total phosphorus, and total nitrogen, can be related to intermediate or final ecosystem services including several listed in Table 1.
Figure 3.
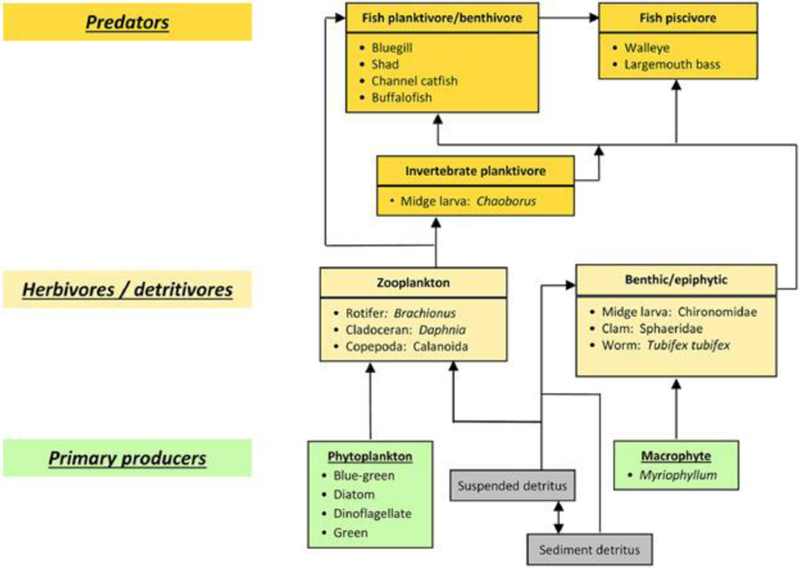
Generalized food web as represented in the AQUATOX model. Although some species feed at multiple trophic levels, which will be accounted for in the food preference matrices of the AQUATOX model, only the primary functional feeding role is shown in this figure. All biotic groups contribute to the suspended and sediment detritus through death and decomposition (those arrows are omitted for the sake of simplicity).
Valuing Costs and Benefits to Beneficiaries of Ecosystem Services
An important rationale for the approach outlined in the present Focus article is that it can make an explicit link between the benefits derived from mitigating an impact on ecosystem services with the costs of the management options to optimize delivery of increased welfare to those affected. There are 3 elements to these analyses—assessing costs, assessing benefits, and identifying the people affected—and each of these raises challenges and uncertainties (for a good general review, see Hanley and Barbier 68). The costs of reducing exposures relate to the costs of reducing the concentration of chemicals in the environment, for example, through better treatment of effluents and/or restricting releases. Being technological, the costs of treatment are straightforward but should include capital investment and running costs. The costs of chemical restrictions are more difficult to quantify and should take into account both the administrative costs of any intervention as well as the opportunity cost of loss of service or product or incomplete replacement in the event of substitution. The monetary values of ecosystem services are more problematic. For those services that lead to goods that are traded in markets, such as fish, a market valuation is obvious. But most are not traded (e.g., recreational fisheries), and then values are assessed from surveys of willingness to pay (stated preferences) or observations of behavior such as travel to see a natural resource (revealed preferences). An important distinction is made between use values that capture benefits that are directly used by the people concerned and nonuse, existence, values that reflect the desire to preserve ecological entities (e.g., species) even if they are not used. Existence values are more difficult to assess than use values 69. All of these kinds of surveys are time-consuming, so there is a temptation to use literature values; but this raises additional uncertainties because the values that people put on ecosystems are sensitive to socioeconomic circumstances. Where costs and benefits are incurred at different times, discounts have to be considered to take account of the time sensitivity of preferences. Finally, of considerable importance to calculating total benefit is identifying the people who represent the socioeconomic group that benefits from an ecosystem service as well as the size of that group.
Introduction of Case Studies
In the following section we introduce 2 case studies to demonstrate use of a mechanistic modeling approach to predict whether and how effects of chemicals at individual (or subindividual) organism levels (i.e., what we measure) translate into impacts on ecosystem service delivery (what we care about). The case studies are expected to show how commonly collected toxicity test data, for fish and Daphnia, respectively, can be mechanistically linked to population and multispecies effects and how such effects can be translated (via ecological production functions) into impacts on ecosystem service delivery with corresponding economic consequences. We have chosen fish and Daphnia because there is a wealth of toxicity data for these taxa as well as a variety of models that could be adapted for our purposes. Although there is some overlap between the case studies (e.g., both focus on freshwater ecosystems and both include provision of recreational fish as one of the final ecosystem goods and services considered), we believe that it will be illuminating to explore the problem using different models, different environments (lentic vs lotic), and different chemical stressors.
Case study 1: Predicting impacts of an endocrine disruptor on valued fish populations
Conceptual model
The conceptual model for this case study is shown in Figure 4. The case study considers a mountain stream system with 2 trout species. Although the case is hypothetical, the ecosystem is based on streams supporting greenback cutthroat trout, Oncorhynchus clarkii stomias. The greenback cutthroat trout was officially declared extinct in 1937. Additional populations were later found, but the range of this species is currently limited to the upper Arkansas and South Platte basins of Colorado, USA. In the present study system, based on the Arkansas River (Figure 5), greenback cutthroat trout coexist with introduced brown trout, Salmo trutta. Both species are fished recreationally; therefore, the estimated final ecosystem goods and services will be provision of catchable fish. We recognize that the monetized value of blue-ribbon trout fisheries in this region is largely determined by out-of-state, rather than residential, fishers 70, 71. In addition, because the greenback cutthroat trout is the state fish of Colorado, the presence of this species as a final ecosystem good will be considered, with the citizens of Colorado as the beneficiary.
Figure 4.
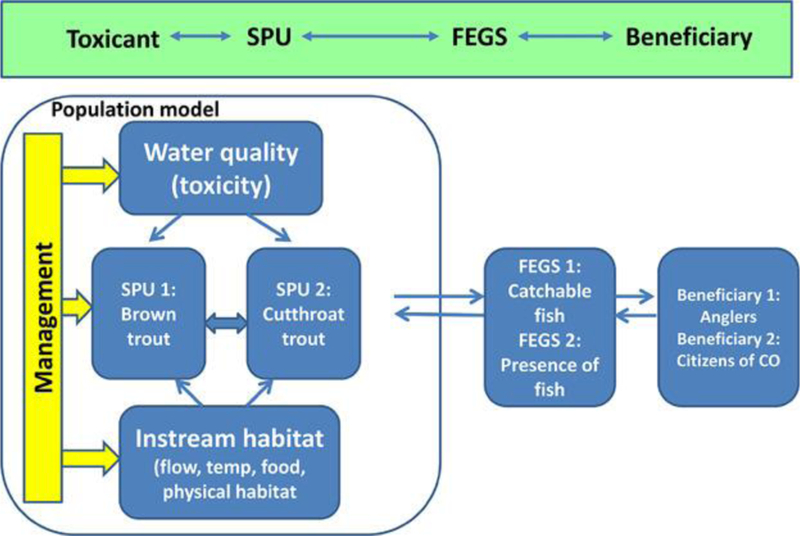
Conceptual model for case study 1—impacts of an endocrine disruptor on valued fish populations. CO = Colorado; FEGS = final ecosystem good or service; SPU = service providing unit.
Figure 5.

Geographical location and stream reaches simulated in case study 1.
We will develop this case study to explore the potential population-level impacts of exposure to 17α-ethinylestradiol (EE2). A derivative of 17β-estradiol is EE2, which is the major endogenous estrogen in humans and the bioactive estrogen in many oral contraceptives. Exposure of fish to environmental estrogens is known to affect sexual development and spawning 72. We will assume that EE2 enters the Arkansas River via sewage-treatment outfalls and that the concentration of EE2 in the sewage-treatment effluent is constant. Recognizing that concentrations in the stream will vary with flow, for simplicity we will begin with a constant exposure at realistic measured environmental levels (based on available environmental monitoring data). Because we assume secondary treatment of sewage (i.e., removal of solids and dissolved and suspended biological matter, followed by chemical or physical disinfection), we do not consider organic enrichment effects on prey of trout.
Mechanistic effects modeling
We will represent the habitat as a network of 5 linked stream reaches of 3 sizes (mainstem, large, and small tributaries 73). Although the physical habitat of each reach is copied from other study sites, we produce input representing greenback cutthroat trout habitat in the upper Arkansas River basin (Figure 5). Daily flows and temperatures can be obtained from US Geological Survey gages on the Arkansas and Eagle Rivers for the last 22 yr (1994–2015). Flow input will be scaled to match the watershed areas represented by each simulated reach. In this case study the final ecosystem goods and services of concern are delivered by populations of trout, and we need to mechanistically link EE2 toxicity data to impacts at the population level. We will adapt the inSTREAM model for trout populations 61, 62 to simulate trout population dynamics under the influence of a chemical contaminant. The simulations will represent greenback cutthroat trout and brown trout, with the only differences between the species being 1) when they spawn (brown trout spawn in late fall, with eggs incubating until early spring and hence being vulnerable to winter floods; greenback cutthroat trout spawn in the spring, so their eggs are vulnerable to spring floods), 2) the relationships between temperature and egg development and survival rates, and 3) small differences in how length varies with weight.
Cost–benefit analyses
These analyses will involve comparing the increased amounts of catchable fish against the costs of constructing and running technology for removing EE2 and its derivatives. Because the fishery is recreational, it has no market value. Values will therefore be obtained from the increase in travel to sites by those who fish as catches increase and the costs they incur as a result 74. This can be computed from licensing data and/or surveys. Willingness-to-pay surveys have also been used for assessing the values that those who fish put on good catches 75. In either case, using revealed or stated preferences, benefits to local populations will be considered in terms of impacts on their general economy through commerce and property values. The costs of the treatment technology will involve installation and running of sophisticated systems involving, for example, ultrafiltration and ozonation as tertiary treatment. The costs will be technology-specific but are likely to be considerable 76. The management decision on how much extra treatment to deploy will depend on the extent to which costs of the technology make sense against the returns from the increased catch, all discounted to a common time. Willingness-to-pay surveys can also be used to assess the value that people put on the existence of a species (e.g., the state fish); however, in this case the relevant beneficiaries would be at least statewide, and designing an appropriate survey will therefore be challenging 69, 70.
Issues to address
Developing this case study will require that we collect relevant toxicity data for EE2, extrapolate these data to the present study species, and translate them mechanistically into dose–response functions for trout survival, growth, and/or reproduction. These dose–response functions will be integrated into inSTREAM to provide impacts on trout population-level impacts. The primary final ecosystem goods and services metric for this case study will be abundance of catchable-sized trout. As a first approximation, we will assume constant EE2 exposure concentrations; however, investigating consequences of fluctuating concentrations caused by changes in river flow could also be of interest. Whereas inSTREAM incorporates bioenergetics in several key processes, one of our aims is to explore whether revising the model to use dynamic energy budget theory (see Einarsson et al. 77 for a dynamic energy budget model of a related species) offers any advantages in terms of mechanistic insight, predictability, or the ability to connect with responses at lower levels of biological organization. Outputs of inSTREAM will be expressed as (or translated into) metrics that are valued by beneficiaries (Table 1) and subjected to valuation using appropriate economic models. The valuation step will facilitate an assessment of the costs and benefits (e.g., of additional sewage treatment, tourism revenue) of different management alternatives.
Case study 2: Predicting impacts of a pesticide on lake water clarity and recreational fishing
Conceptual model
The conceptual model for this case study is shown in Figure 6. This case study is based on Coralville Reservoir, Iowa (USA), which is a eutrophic run-of-the river lake formed in 1958 by impoundment of the Iowa River. Although the dimensions of the lake vary depending on use as water storage, the lake has a mean surface area of 22 km2, a maximum depth of 9.1 m, and a mean depth of 2.5 m (5.8 m when water is at spillway level). According to the US Department of Agriculture, 57% of the watershed was identified as cropland in 2011 78.
Figure 6.
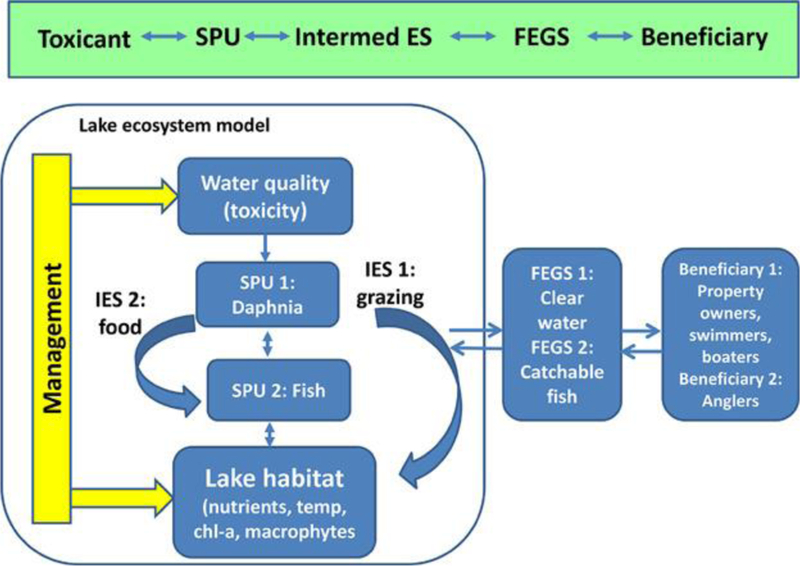
Conceptual model for case study 2—impacts of a pesticide on a lake for recreational use. chl-a = chlorophyll a; Intermed ES/IES = intermediate ecosystem service; FEGS = final ecosystem good or service; SPU = service providing unit.
The reservoir is closely monitored, and water quality reports are published periodically. The reservoir has a relatively simple fish community for a lake its size. Water-level fluctuations limit rooted macrophytes in the littoral zone, limiting the size and diversity of game fish populations. Nevertheless, anglers appear to especially target species such as crappie, bass, and channel catfish.
This case study will examine impacts on 2 final ecosystem goods and services: 1) provisioning of clear water for recreation, with 3 groups of beneficiaries—property owners; waders, swimmers, and divers; and boaters—and 2) provisioning of a fishery with recreational anglers as the primary beneficiary. Although beneficiaries in this case study are those affected via impacts on the aquatic food web, for other sites it would be possible to include additional beneficiaries such as municipal drinking water plant operators (who are responsible for supplying safe drinking water to the public).
We explore the effects of an organophosphate insecticide on zooplankton and fish and the potential implications for ecosystem services. We selected this group of pesticides for the case study because of the extensive toxicity data and continued use of these chemicals. We assume that the pesticide enters the Coralville Reservoir via drift and runoff after application for agricultural uses, leading to temporally varying exposure of the aquatic (nontarget) species. Exposure of daphnids to peaks of different duration and pesticide concentrations has shown that chemicals of this kind could reduce daphnid survival for some of the relevant exposure patterns 79. A reduction in zooplankton could lead to an increase in algae, thereby reducing water clarity. In addition, we expect fish to be much less sensitive to the insecticide than their invertebrate prey but likely to suffer indirect effects of the insecticide because of reduced food.
Mechanistic effects modeling
In this case study, the final ecosystem goods and services of concern involve interactions among several species, and therefore we need a model that links standard toxicity data (likely to be available for Daphnia and fish) to the ecosystem level. AQUATOX is selected as the mechanistic effects model for this case study because it can simulate temporal trends in the biomass of daphnids, harvestable fish, and other species within a lake food web. The ecosystem model constructed for this case study includes phytoplankton, periphyton, macrophytes, zooplankton, benthic/epiphytic invertebrates, and fish within a lake habitat. Habitat compartments include surface water, sediments, and atmosphere, with temperature, nutrient regimes, and incoming solar radiation, as well as the flow of nutrients, detritus, and insecticide, between various environmental compartments. In addition, AQUATOX allows evaluation of indirect effects through food-web interactions; has been well tested and validated for the Coralville Reservoir and other aquatic systems throughout the United States, Europe, and Asia; and is readily available and supported by the USEPA.
Cost–benefit analyses
These analyses will involve comparing the value that people accord to clear water and the associated improved recreational opportunities with the costs of achieving the cleaner conditions either by deploying technology that removes pesticide to the extent required or by controlling pesticide inputs to the water body through label restrictions, other agricultural practices, or mitigation measures such as buffer strips. Stated preference techniques can be used to assess the value that people ascribe to lake clarity 80, but they are time-consuming. Keeler et al. 81 have developed a novel, revealed preference technique that promises a more cost-effective and generalizable approach that will be relevant for the present study. They used geotagged photographs uploaded to a photo-sharing public website by lake visitors across a range of lakes in Minnesota (USA) and Iowa (USA) to gauge the number of visitors and the distances they traveled to the lakes. From these data, multiple regression techniques showed that users traveled almost 1 h more for every 1-m increase in clarity, and at the time this amounted to US$22, taking account of both the value of personal time used (as fraction of salaries) and transportation costs. This technique counts all visitors irrespective of use but again will not account for changes to the local economy. For making a management decision, the monetary benefits have to be calibrated against the cost of achieving each meter of clarity from restrictions on pesticide applications and/or mitigation measures. Reduced pesticide application can result in costs associated with the loss of crop yield to insects, competing weeds, and disease 82. Introduction of buffer zones involves the costs from reduced growing area for crops and, in the event of planted zones, the costs of initial establishment and subsequent maintenance 83. Again, discounting to a common time may be important. Valuation related to provision of harvestable fish will be estimated as in case study 1.
Issues to address
Because nonlinear dynamics arise from population processes and trophic interactions, these services are not directly proportional to our intermediate service-providing unit, the zooplankton food resource that is represented by Daphnia. Therefore, a multispecies model is needed to translate changes in abundance (or other metrics) of zooplankton resulting from toxic chemical effects to metrics related to final ecosystem goods and services. We will consider 2 metrics for ecosystem services: 1) water clarity (e.g., as measured by Secchi depth), and 2) biomass/abundance of recreationally harvested fish (e.g., bass, channel catfish, and forage fish larger than a specified size). To explore different realistic scenarios of pesticide exposure, we will use the USEPA exposure model that simulates runoff and drift transport 84.
Although AQUATOX does have the capability to model toxicant effects on food-web components, one goal of this case study is to integrate dynamic energy budget theory with AQUATOX. This integration will facilitate extrapolation of toxicity data from standard test species to other species included in the food web and will provide a mechanistic interface with toxic responses of test species at lower levels of organization. Dynamic energy budget models that describe the direct effects of the chemical on fish and daphnids may be coupled to AQUATOX, which describes the biomass of fish and daphnid populations. The AQUATOX model will provide the environmental context where the daphnids, fish, and other species (e.g., algae prey species for the daphnids) live; and the dynamic energy budget model will allow estimation of the reduction in biomass in fish and daphnids as a function of the chemical concentration.
As for case study 1, outputs of the mechanistic effects model (AQUATOX) will be expressed as (or translated into) metrics that are valued by beneficiaries (Table 1) and subjected to valuation using appropriate economic models. The valuation step will facilitate an assessment of the costs and benefits of different management alternatives. Because this case study considers multiple services (intermediate and final ecosystem goods and services) and beneficiaries, the cost–benefit analysis will be more complex.
Conclusions and Future Work
Ecological risk assessment should inform risk-management decisions. However, the data typically collected for ERAs are frequently at the level of the organism or below and do not correspond directly with ecological protection goals, which themselves may be only loosely defined in the relevant environmental legislation. Using ecosystem services for identifying and articulating environmental protection goals has a number of benefits as outlined in the present Focus article. We present a framework that uses mechanistic effects models to quantitatively link the data collected for ERAs to the delivery of final ecosystem goods and services and in so doing provides the kind of relevant exposure–response relationships that are directly and quantitatively related to things people value. In the present Focus article, we also provide a conceptual overview of our approach and introduce 2 case studies that are under development. By coordinating with a parallel National Institute of Mathematical and Biological Synthesis working group that is developing models to link high-throughput molecular data to organism-level effects using quantitative AOPs, we hope to create a single, unified modeling approach. At a minimum, these efforts will identify key data and methodological gaps that prevent robust extrapolation of chemical impacts across different levels of biological organization and highlight priorities for future research in this area. If successful, this coordinated research program will provide a comprehensive and mechanistically based framework for predicting effects of chemicals on ecologically relevant protection goals from modern high-throughput toxicological data.
Acknowledgment
The present study was conducted as part of the Organisms to Ecosystems Working Group at the National Institute for Mathematical and Biological Synthesis, sponsored by the National Science Foundation through award DBI-1300426, with additional support from The University of Tennessee, Knoxville. We thank members of our sister working group (Molecules to Organisms) and in particular its cochairs, C. Murphy and R. Nisbet, for helpful discussion and input.
Footnotes
Publisher's Disclaimer: Disclaimer
The views expressed in the present Focus article are those of the authors and do not necessarily reflect the views or policies of the US Environmental Protection Agency (USEPA). Mention of trade names, products, or services does not convey and should not be interpreted as conveying official USEPA approval, endorsement, or recommendation.
Data Availability
The present study was coauthored by UT-Battelle under contract DE-AC05-00OR22725 with the US Department of Energy. The US government retains and the publisher, by accepting the Focus article for publication, acknowledges that the US government retains a nonexclusive, paid-up, irrevocable, worldwide license to publish or reproduce the published form of this manuscript, or allow others to do so, for US government purposes. The Department of Energy will provide public access to these results of federally sponsored research in accordance with its Public Access Plan (http://energy.gov/downloads/doe-public-access-plan).
References
- 1.van Wensem J, Calow P, Dooacker A, Maltby L, Olander L, Tuvendal M, van Houtven G. 2016Identifying and assessing the application of ecosystem services approaches in environmental policies and decision-making. Integr Environ Assess Manage 13:41–51. [DOI] [PubMed] [Google Scholar]
- 2.Stark JD, Banks JE, Vargas R. 2004. How risky is risk assessment: The role that life history strategies play in susceptibility of species to stress. Proc Natl Acad Sci USA 101:732–736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Forbes VE, Calow P, Grimm V, Hayashi TI, Jager T, Katholm A, Palmqvist A, Pastorok R, Salvito D, Sibly RM, Spromberg JA, Stark J, Stillman RA. 2011. Adding value to ecological risk assessment with population modelling. Hum Ecol Risk Assess 17:287–299. [Google Scholar]
- 4.Hommen U, Forbes VE, Grimm V, Preuss T, Thorbek P, Ducrot V. 2015. How to use mechanistic effect models in risk assessment of pesticides: Case studies and recommendations from the SETAC workshop MODELINK. Integr Environ Assess Manage 12:21–31. [DOI] [PubMed] [Google Scholar]
- 5.Gust KA, Collier ZA, Mayo ML, Stanley JK, Gong P, Chappell MA. 2015. Limitations of toxicity characterization in life cycle assessment: Can adverse outcome pathways provide a new foundation? Integr Environ Assess Manage 12:580–590. [DOI] [PubMed] [Google Scholar]
- 6.Walker BH. 1992. Biodiversity and ecological redundancy. Conserv Biol 6:18–23. [Google Scholar]
- 7.Paine R 1966. Food web complexity and species diversity. Am Nat 100:65–75. [Google Scholar]
- 8.Lenton TM. 2013. Environmental tipping points. Annu Rev Environ Res 38:1–29. [Google Scholar]
- 9.Forbes VE, Galic N, Schmolke A, Vavra J, Pastorok R, Thorbek P. 2016. Assessing the risks of pesticides to threatened and endangered species using population modeling: A review and recommendations for future work. Environ Toxicol Chem 35:1904–1913. [DOI] [PubMed] [Google Scholar]
- 10.Forbes VE, Galic N. 2016. Next generation ecological risk assessment: Predicting risk from molecular initiation to ecosystem service delivery. Environ Int 91:215–219. [DOI] [PubMed] [Google Scholar]
- 11.National Institute of Mathematical and Biological Synthesis. NIMBioS Investigative Workshop: Predictive Models for ERA. 2016 Nov 28; [cited. ]. Available from: http://www.nimbios.org/workshops/WS_era.
- 12.National Institute of Mathematical and Biological Synthesis. NIMBioS Working Group: Modeling Organisms-to-Ecosystems. 2016 Nov 28; [cited. ]. Available from: http://www.nimbios.org/workinggroups/WG_o2e.
- 13.National Institute of Mathematical and Biological Synthesis. NIMBioS Working Group: Modeling Molecules-to-Organisms. 2016 Nov 28; [cited. ]. Available from: http://www.nimbios.org/workinggroups/WG_m2o.
- 14.US Environmental Protection Agency. 1992. Framework for ecological risk assessment EPA/630/R-92/001. Washington, DC. [Google Scholar]
- 15.Nienstedt K, Brock T, van Wensem J, Montforts M, Hart A, Hardy A, Aagaard A, Alix A, Boesten J, Bopp SK, Brown C, Capri E, Forbes VE, Köpp H, Liess M, Luttik R, Maltby L, Sousa P, Streissl F.2012. Development of a framework based on an ecosystem services approach for deriving specific protection goals for environmental risk assessment of pesticides. Sci Total Environ 415:31–38. [DOI] [PubMed] [Google Scholar]
- 16.Millennium Ecosystem Assessment. 2005. Ecosystems and Human Well-Being: Synthesis Island, Washington, DC. [Google Scholar]
- 17.Forbes VE, Calow P. 2012. Promises and problems for the new paradigm for risk assessment and an alternative approach involving predictive systems models. Environ Toxicol Chem 31:2663–2671. [DOI] [PubMed] [Google Scholar]
- 18.Holt AR, Alix A, Thompson A, Maltby L. 2016. Food production ecosystem services and biodiversity: We can’t have it all everywhere. Sci Total Environ 573:1422–1429. [DOI] [PubMed] [Google Scholar]
- 19.Boyd J, Banzhaf S. 2007. What are ecosystem services? The need for standardized environmental accounting units. Ecol Econ 63:616–626. [Google Scholar]
- 20.Nahlik AM, Landers DH, Ringold PL, Weber MA. 2012. Protecting our environmental wealth: Connecting ecosystem goods and services to human well-being. National Wetlands Newsletter 34:14–18. [Google Scholar]
- 21.Munns WR, Rea AW, Mazzotta MJ, Wainger LA, Saterson K. 2015. Toward a standard lexicon for ecosystem services. Integr Environ Assess Manage 11:666–673. [DOI] [PubMed] [Google Scholar]
- 22.Munns WR, Rea AW, Suter GW, Martin L, Blake-Hedges L, Crk T, Davis C, Ferreira G, Jordan S, Mahoney M, Barron MG. 2015. Ecosystem services as assessment endpoints for ecological risk assessment. Integr Environ Assess Manage 12:522–528. [DOI] [PubMed] [Google Scholar]
- 23.Donovan S, Goldfuss C, Holdren J. 2015. Incorporating ecosystem services into federal decision making. Memorandum for executive departments and agencies M-16-01. [cited 2016. April 7]. Available from: https://www.whitehouse.gov/sites/default/files/omb/memoranda/2016/m-16-01.pdf
- 24.US Environmental Protection Agency. 2016. Generic ecological assessment endpoints (GEAEs) for ecological risk assessment: Second edition with generic ecosystem services endpoints added. EPA/100/F15/005. Washington, DC. [Google Scholar]
- 25.US Environmental Protection Agency. 2016. Ecosystem services as assessment endpoints in ecological risk assessment EPA/100/F-15/004. Washington, DC. [DOI] [PubMed] [Google Scholar]
- 26.Schroeder AL, Ankley GT, Houck KA, Villeneuve DL. 2016. Environmental surveillance and monitoring—The next frontiers for high-throughput toxicology. Environ Toxicol Chem 35:513–525. [DOI] [PubMed] [Google Scholar]
- 27.Landers DH, Nahlik AM. 2013. Final ecosystem goods and services classification system (FEGS-CS) EPA/600/R-13/ORD-004914. Washington, DC. [Google Scholar]
- 28.Grimm V, Thorbek P. 2014. Population models for ecological risk assessment of chemicals: Short introduction and summary of a special issue. Ecol Model 280:1–4. [Google Scholar]
- 29.Galic N, Forbes VE. 2014. Ecological models in ecotoxicology and ecological risk assessment: An introduction to the special section. Environ Toxicol Chem 33:1446–1448. [DOI] [PubMed] [Google Scholar]
- 30.Bruins RJF, Canfield TJ, Duke C, Kapustka L, Nahlik AM, Schäfer RB. 2016. Using ecological production functions to link ecological processes to ecosystem services. Integr Environ Assess Manage 13:52–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jager HI, Hargrove WW, Brandt CC, King AW, Olson RJ, Scurlock JMO, Rose KA. 2000Constructive contrasts between modeled and measured climate responses over a regional scale. Ecosystems 3:396–411. [Google Scholar]
- 32.Railsback SF, Grimm V. 2011. Agent-Based and Individual-Based Modeling: A Practical Introduction Princeton University Press, Princeton, NJ, USA. [Google Scholar]
- 33.Grimm V, Berger U, DeAngelis DL, Polhill JG, Giske J, Railsback SF. 2010. The ODD protocol: A review and first update. Ecol Model 221:2760–2768. [Google Scholar]
- 34.Grimm V, Augusiak J, Focks A, Frank BM, Gabsi F, Johnston ASA, Liu C, Martin BT, Meli M, Radchuk V, Thorbek P, Railsback S. 2014. Towards better modelling and decision support: Documenting model development, testing, and analysis using trace. Ecol Model 280:129–139. [Google Scholar]
- 35.Grimm V, Railsback SF. 2012. Pattern-oriented modelling: A “multiscope” for predictive systems ecology. Philos Trans R Soc Lond B Biol Sci 367:298–310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Augusiak J, van den Brink PJ, Grimm V. 2014. Merging validation and evaluation of ecological models to “evaluation”: A review of terminology and a practical approach. Ecol Model 280:117–128. [Google Scholar]
- 37.Bartell SM, Gardner RH, O’Neill RV. 1992. Ecological Risk Estimation Lewis, Chelsea, MI, USA. [Google Scholar]
- 38.Pastorok RA, Bartell SM, Ferson S, Ginzburg LR. 2002. Ecological Modeling in Risk Assessment: Chemical Effects on Populations, Ecosystems, and Landscapes CRC, Boca Raton, FL, USA. [Google Scholar]
- 39.Thorbek P, Forbes V, Heimbach F, Hommen U, Thulke H-H, Van den Brink PJ, Wogram J, Grimm V, eds. 2010. Ecological Models for Regulatory Risk Assessments of Pesticides: Developing a Strategy for the Future CRC, Boca Raton, FL, USA. [DOI] [PubMed] [Google Scholar]
- 40.Galic N, Hommen U, Baveco JM, van den Brink PJ. 2010. Potential application of population models in the European ecological risk assessment of chemicals. II. Review of models and their potential to address environmental protection aims. Integr Environ Assess Manage 6:338–360. [DOI] [PubMed] [Google Scholar]
- 41.Jager T, Albert C, Preuss TG, Ashauer R. 2011. General unified threshold model of survival—A toxicokinetic–toxicodynamic framework for ecotoxicology. Environ Sci Technol 45:2529–2540. [DOI] [PubMed] [Google Scholar]
- 42.European Commission. 2013. Making risk assessment more relevant for risk management. Opinion of the Scientific Committee on Health and Environmental Risks (SCHER) and the Scientific Committee on Emerging and Newly Identified Health Risks (SCENIHR) [cited 2013 January]. Available from: http://ec.europa.eu/health/scientific_committees/consumer_safety/docs/sccs_o_130.pdf
- 43.European Commission. 2013. Addressing the new challenges for risk assessment. Joint Opinion of SCHER (Scientific Committee on Health and Environmental Risks), SCENIHR (Scientific Committee on Emerging and Newly Identified Health Risks), SCCS (Scientific Committee on Consumer Safety), Addressing the new challenges for risk assessment [cited 2013 January]. Available from: http://ec.europa.eu/health/scientific_committees/consumer_safety/docs/sccs_o_131.pdf
- 44.National Research Council. 2013. Assessing Risks to Endangered and Threatened Species from Pesticides National Academies, Washington, DC. [Google Scholar]
- 45.European Food Safety Authority. 2014. Scientific opinion on good modelling practice in the context of effect models for risk assessment of plant protection products. EFSA Panel on Plant Protection Products and their Residues. EFSA Journal 12:3589. [Google Scholar]
- 46.US Environmental Protection Agency. 1998. Guidelines for ecological risk assessment EPA/630/R-95/002F. Washington, DC: Available from: https://www.epa.gov/sites/production/files/2014-11/documents/eco_risk_assessment1998.pdf [Google Scholar]
- 47.Ashauer R, Escher BI. 2010. Advantages of toxicokinetic and toxicodynamic modelling in aquatic ecotoxicology and risk assessment. J Environ Monitor 12:2056–2061. [DOI] [PubMed] [Google Scholar]
- 48.Kooijman SALM, Metz JA. 1984. On the dynamics of chemically stressed populations—The deduction of population consequences from effects on individuals. Ecotoxicol Environ Saf 8:254–274. [DOI] [PubMed] [Google Scholar]
- 49.Kooijman SALM. 2010. Dynamic Energy Budget Theory for Metabolic Organisation Cambridge University Press, Cambridge, UK. [Google Scholar]
- 50.Jager T, Heugens EHV, Kooijman SALM. 2006. Making sense of ecotoxicological test results: Towards application of process-based models. Ecotoxicology 15:305–314. [DOI] [PubMed] [Google Scholar]
- 51.Sousa T, Domingos T, Poggiale JC, Kooijman SALM. 2010. Dynamic energy budget theory restores coherence in biology. Philos Trans R Soc Lond B Biol Sci 365:3413–3428. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jager T 2012. Making Sense of Chemical Stress. Applications of Dynamic Energy Budget Theory in Ecotoxicology and Stress Ecology, Ver 9. [cited 2016 November 28]. Available from: http://www.debtox.info/book.php
- 53.Jager T, Barsi A, Hamda NT, Martin BT, Zimmer EI, Ducrot V. 2014. Dynamic energy budgets in population ecotoxicology: Applications and outlook. Ecol Model 280:140–147. [Google Scholar]
- 54.Raimondo S, Mineau P, Barron MG. 2007. Estimation of chemical toxicity to wildlife species using interspecies correlation models. Environ Sci Technol 41:5888–5894. [DOI] [PubMed] [Google Scholar]
- 55.Ankley GT, Bennett RS, Erickson RJ, Hoff DJ, Hornung MW, Johnson RD, Mount DR, Nichols JW, Russom CL, Schmieder PK, Serrrano JA, Tietge JE, Villeneuve DL. 2010. Adverse outcome pathways: A conceptual framework to support ecotoxicology research and risk assessment. Environ Toxicol Chem 29:730–741. [DOI] [PubMed] [Google Scholar]
- 56.Groh KJ, Carvalho RN, Chipman JK, Denslow ND, Halder M, Murphy CA, Roelofs D, Rolaki A, Schirmer K, Watanabe KH. 2015. Development and application of the adverse outcome pathway framework for understanding and predicting chronic toxicity: I. Challenges and research needs in ecotoxicology. Chemosphere 120:764–777. [DOI] [PubMed] [Google Scholar]
- 57.Daily GC, Polasky S, Goldstein J, Kareiva PM, Mooney HA, Pejchar L, Ricketts TH, Salzman J, Shallenberger R. 2009. Ecosystem services in decision making: Time to deliver. Front Ecol Environ 7:21–28. [Google Scholar]
- 58.Salice CJ, Miller TJ. 2003. Population-level responses to long-term cadmium exposure in two strains of the freshwater gastropod Biomphalaria glabrata: Results from a life-table response experiment. Environ Toxicol Chem 22:678–688. [PubMed] [Google Scholar]
- 59.Stillman RA, Railsback SF, Giske J, Berger U, Grimm V. 2015. Making predictions in a changing world: The benefits of individual-based ecology. Bioscience 65:140–150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Grimm V, Berger U. 2016. Structural realism, emergence, and predictions in next-generation ecological modelling: Synthesis from a special issue. Ecol Model 326:177–187. [Google Scholar]
- 61.Railsback SF, Harvey BC. 2002. Analysis of habitat selection rules using an individual-based model. Ecology 83:1817–1830. [Google Scholar]
- 62.Railsback SF, Harvey BC, Jackson SK, Lamberson RH. 2009. InSTREAM: The individual-based stream trout research and environmental assessment model. Report PSW-GTR-218, US Department of Agriculture Forest Service, Pacific Southwest Research Station, Albany, CA [cited 2016 November 28]. Available from: www.fs.fed.us/psw/publications/documents/psw_gtr218/psw_gtr218.pdf
- 63.Layman CA, Giery ST, Buhler S, Rossi R, Penland T, Henson MN, Bogdanoff AK, Cove MV, Irizarry AD, Schalk CM. 2015. A primer on the history of food web ecology: Fundamental contributions of fourteen researchers. Food Webs 4:14–24. [Google Scholar]
- 64.Jørgensen SE. 2011. Handbook of Ecological Models Used in Ecosystem and Environmental Management Copenhagen University, Copenhagen, Denmark. [Google Scholar]
- 65.DeAngelis DL, Gross LJ, Huston MA, Wolff WF, Fleming DM, Comiskey EJ, Sylvester SM. 1998Landscape modeling for Everglades ecosystem restoration. Ecosystems 1:64–75. [Google Scholar]
- 66.Janssen ABG, Arhonditsis GB, Beusen A, Bolding K, Bruce L, Bruggeman J, Couture R-M, Downing AS, Elliott JA, Frassl MA, Gal G, Gerla DJ, Hipsey MR, Hu F, Ives SC, Janse JH, Jeppesen E, Jöhnk KD, Kneis D, Kong X, Kuiper JJ, Lehmann MK, Lemmen C, Özkundakci D, Petzoldt T, Rinke K, Robson BJ, Sachse R, Schep SA, Schmid, Scholten H, Teurlincx S, Trolle D, Troost TA, Van Dam AA, Van Gerven LPA, Weijerman M, Wells SA, Mooij WM. 2015. Exploring, exploiting and evolving diversity of aquatic ecosystem models: A community perspective. Aquat Ecol 49:513–548. [Google Scholar]
- 67.Park RA, Clough JS, Wellman MC. 2008. AQUATOX: Modeling environmental fate and ecological effects in aquatic ecosystems. Ecol Model 213:1–15. [Google Scholar]
- 68.Hanley N, Barbier EB. 2009. Pricing Nature: Cost–Benefit Analysis and Environmental Policy Edward Elgar, Cheltenham, UK, and Northampton, MA, USA. [Google Scholar]
- 69.Frank EG, Schlenker W. 2016. Balancing economic and ecological goals. Science 353:651–652. [DOI] [PubMed] [Google Scholar]
- 70.Loomis JB, White DS, 1996. Economic benefits of rare and endangered species: Summary and meta analysis. Am J Agric Econ 78:1407–1407. [Google Scholar]
- 71.Layman RC, Boyce JR, Criddle KR. 1996. Economic valuation of the chinook salmon sport fishery of the Gulkana River, Alaska, under current and alternate management plans. Land Econ 72:113–128. [Google Scholar]
- 72.Brown KH, Schultz IR, Nagler JJ. 2007. Reduced embryonic survival in rainbow trout resulting from paternal exposure to the environmental estrogen 17α-ethynylestradiol during late sexual maturation. Reproduction 134:659–666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Harvey BC, Railsback SF. 2012. Effects of passage barriers on demographics and stability properties of a virtual trout population. River Research and Applications 28:479–489. [Google Scholar]
- 74.Shrestha RK, Seidl AF, Moraes AS. 2002. Value of recreational fishing in the Brazilian Pantanal: A travel cost analysis using count data models. Ecol Econ 42:289–299. [Google Scholar]
- 75.Connelly NA, Brown TL. 1991. Net economic value of the freshwater recreational fisheries of New York. Trans Am Fish Soc 120:770–775. [Google Scholar]
- 76.Margota J, Kienleb C, Magnetc A, Weild M, Rossia L, de Alencastroa LP, Abegglene C, Thonneyc D, Chèvref N, Schärerg M, Barrya DA. 2013. Treatment of micropollutants in municipal wastewater: Ozone or powdered activated carbon? Sci Total Environ 461–462:480–495. [DOI] [PubMed] [Google Scholar]
- 77.Einarsson B, SigurÐsson SÞ, Birnir B. 2011. A dynamic energy budget (DEB) model for the energy usage and reproduction of the Icelandic capelin (Mallotus villosus). J Theor Biol 281:1–8. [DOI] [PubMed] [Google Scholar]
- 78.Natural Resources Conservation Service. Iowa River, middle—07080208. [cited 2016 November 28]. Available from: http://www.nrcs.usda.gov/Internet/FSE_DOCUMENTS/nrcs142p2_006352.pdf.
- 79.Naddy RB, Johnson KA, Klaine SJ. 2000. Response of Daphnia magna to pulsed exposures of chlorpyrifos. Environ Toxicol Chem 19:423–431. [Google Scholar]
- 80.Carson RT, Mitchell RC. 1993. The value of clean water: The public’s willingness to pay for boatable, fishable and swimmable water quality. Water Resour Res 29:245–254. [Google Scholar]
- 81.Keeler BL, Wood SA, Polasky S, Kling C, Filstrup CT, Downing JA. 2015. Recreational demand for clean water: Evidence from geotagged photographs by visitors to lakes. Front Ecol Environ 13:76–81. [Google Scholar]
- 82.Popp J, Pető K, Nagy J. 2013. Pesticide productivity and food security. Agron Sustain Dev 33:243–255. [Google Scholar]
- 83.Roberts DC, Clark CD, English C, Park WM, Roberts RK. 2009. Estimating annualized riparian buffer costs for the Harpeth River watershed. Review of Agricultural Economics 31:894–913. [Google Scholar]
- 84.US Environmental Protection Agency. Pesticide in Water Calculator. [cited 2016 November 28]. Available from: https://www.epa.gov/pesticide-science-and-assessing-pesticide-risks/models-pesticide-risk-assessment#PWC. [Google Scholar]
- 85.Christie M, Azevedo CD. 2009. Testing the consistency between standard contingent valuation, repeated contingent valuation and choice experiments. J Agric Econ 60:154–170. [Google Scholar]
- 86.Corrigan JR, Kling CL, Zhao J. 2008. Willingness to pay and the cost of commitment: An empirical specification and test. Resour Econ (Dordr) 40:285–298. [Google Scholar]
- 87.van Houtven G, Mansfield C, Phaneuf DJ, von Haefen R, Milstead B, Kenney MA, Reckhow KH.2014. Combining expert elicitation and stated preference methods to value ecosystem services from improved lake water quality. Ecol Econ 99:40–52. [Google Scholar]
- 88.Nelson NM, Loomis JB, Jakus PM, Kealy MJ, von Stackelburg N, Ostermiller J. 2015. Linking ecological data and economics to estimate the total economic value of improving water quality by reducing nutrients. Ecol Econ 118:1–9. [Google Scholar]
- 89.Collins A, Rosenberger R, Fletcher J. 2005. The economic value of stream restoration. Water Resour Res 41: 10.1029/2004WR003353. [DOI] [Google Scholar]
- 90.Holmes TP, Bergstrom JC, Huszar E, Kask SB, Orr F. 2004. Contingent valuation, net marginal benefits, and the scale of riparian ecosystem restoration. Ecol Econ 49:19–30. [Google Scholar]
- 91.Johnston RJ, Schultz ET, Segerson K, Besedin EY, Ramachandran M. 2013. Stated preferences for intermediate versus final ecosystem services: Disentangling willingness to pay for omitted outcomes. Agric Resour Econ Rev 42:98–118. [Google Scholar]
- 92.Owens N, Simon NB. 2004. The value of improvements to California’s coastal waters: Results from a stated-preference survey. Presented at Valuation of Ecological Benefits: Improving the Science behind Policy Decisions, Washington DC, Oct-27 US Environmental Protection Agency, National Center for Environmental Research; [cited 2016 August 8]. Available from: https://yosemite.epa.gov/ee/epa/eerm.nsf/vwAN/EE-0489-03.pdf/$file/EE-0489-03.pdf [Google Scholar]
- 93.Roe B, Boyle KJ, Teisl MF. 1996. Using conjoint analysis to derive estimates of compensating variation. J Environ Econ Manage 31:145–159. [Google Scholar]
- 94.Carpenter SR, Cole JJ, Hodgson JR, Kitchell JF, Pace ML, Bade D, Cottingham KL, Essington TE, Houser JN, Schindler DE. 2001. Trophic cascades, nutrients and lake productivity: Whole-lake experiments. Ecol Monogr 71:163–186. [Google Scholar]
- 95.Power ME, Matthews WJ, Stewart AJ. 1985. Grazing minnows, piscivorous bass and stream algae: Dynamics of a strong interaction. Ecology 66:1448–1456. [Google Scholar]
- 96.Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, Carpenter SR, Essington TE, Holt RD, Jackson JB, Marquis RJ, Oksanen L, Oksanen T, Paine RT, Pikitch EK, Ripple WJ, Sandin SA, Scheffer M, Schoener TW, Shurin JB, Sinclair AR, Soulé ME, Virtanen R, Wardle DA. 2011. Trophic downgrading of planet earth. Science 333:301–306. [DOI] [PubMed] [Google Scholar]