

Abstract
Imaging of fatty acid (FA) trafficking revealed that FAs stored in lipid droplets were delivered to mitochondria when the cells were starved. This delivery required cytoplasmic lipases and mitochondrial fusion activity, whereas lipid droplets were replenished with FAs supplied by autophagy. These findings have important implications for cancer.
KEYWORDS: autophagy, fatty acid, lipase, lipid droplet, lipophagy, Mfn1, mitochondria, Opa1, tumor metabolism, tumor-stroma interactions
In order to survive and thrive, cells must continually adapt to the changing availability of nutrients. Cells store energy in the form of neutral lipids within lipid droplets (LDs). In response to nutrient deprivation, cells switch from reliance on glucose metabolism to oxidation of fatty acids (FAs), which occurs in mitochondria. How FAs are mobilized from LDs for transfer to mitochondria under these conditions remains unclear. To address this question, we developed a fluorescence pulse-chase assay that allowed us to visualize FA trafficking in live cells.1 Cells were pulsed with a fluorescent FA analog, which accumulated in neutral lipids within LDs under well-fed conditions. Upon subsequent nutrient deprivation, the fluorescent FA was released from LDs and trafficked to mitochondria. Analysis of this process allowed us to address several outstanding questions regarding FA mobilization in starved cells.
One question relates to how FAs are released from LDs. Two possible mechanisms have been described. Cytoplasmic neutral lipases can localize to LDs and directly hydrolyze triacylglycerol.2 Alternatively, autophagy of LDs (called lipophagy) can digest whole LDs or large portions of LDs, including proteins on the LD surface.3 Use of the FA pulse-chase assay allowed us to distinguish between these mechanisms. We found that in starved mouse embryonic fibroblast cells the ubiquitously expressed cytoplasmic lipase adipose triglyceride lipase was necessary for transfer of FAs from LDs to mitochondria, whereas autophagy was dispensable.1 Interestingly, depletion of serum, amino acids, and glucose did not cause upregulation of lipophagy, whereas depletion of serum alone induced lipophagy.1 Thus, the type of nutrient stress seems to affect the mechanism of FA release. Different tissues are also likely to differ in their response to nutrient stress. Although autophagy was dispensable for releasing FAs from LDs, we identified an alternative role for autophagy in the response to starvation: replenishing LDs with FAs recycled from cellular membranes. When cellular membranes were labeled with a fluorescent phospholipid followed by starvation of the labeled cells, fluorescence accumulated within LDs in an autophagy-dependent manner.1 Therefore, autophagy of cellular membranes releases FAs, which accumulate in LDs for continued transfer to mitochondria. The use of LDs as a central conduit for FA trafficking likely minimizes lipotoxicity caused by free FAs in the cytoplasm, which can generate damaging bioactive lipids and permeabilize cellular membranes.4
A second question that we addressed is how FAs released from LDs are trafficked to mitochondria. We observed that LDs in starved cells were closely apposed to mitochondria, implying that FA transfer may occur at interorganellar contact sites. Interestingly, in wild type (WT) cells the mitochondria were highly tubulated in response to starvation, as previously described.5 In addition, FAs were homogeneously distributed throughout the mitochondrial network.1 In contrast, in cells deficient for mitofusin 1 (Mfn1) or optic atrophy 1 (Opa1), which have fragmented mitochondria due to inhibition of mitochondrial membrane fusion, FAs did not become homogenously distributed across the mitochondrial population, resulting in either very low or exceedingly high FA concentrations within individual mitochondrial elements.1 Together, these data suggest that in response to starvation, FAs are transferred from LDs to mitochondria at sites of close contact, and that mitochondrial fusion is required to equilibrate FAs throughout the network and maximize FA availability for β-oxidation reactions. Indeed, cells with impaired mitochondrial fusion were unable to sustain increased β-oxidation in response to starvation, resulting in decreased mitochondrial oxygen consumption.1 This inability to efficiently metabolize FAs had drastic consequences: Mfn1- and Opa1-knockout (KO) cells both stored more FAs within LDs and effluxed FAs from the cell.1 A co-culture assay in which “donor” cells pulsed with fluorescent FA were incubated with labeled “acceptor” cells revealed that acceptor cells cultured with Mfn1 KO or Opa1 KO donor cells were exposed to higher levels of FAs than acceptor cells cultured with WT donor cells.1 Thus, dysregulation of mitochondrial morphology has metabolic consequences not only for the cell, but also for its neighbors (Fig. 1A).
Figure 1.
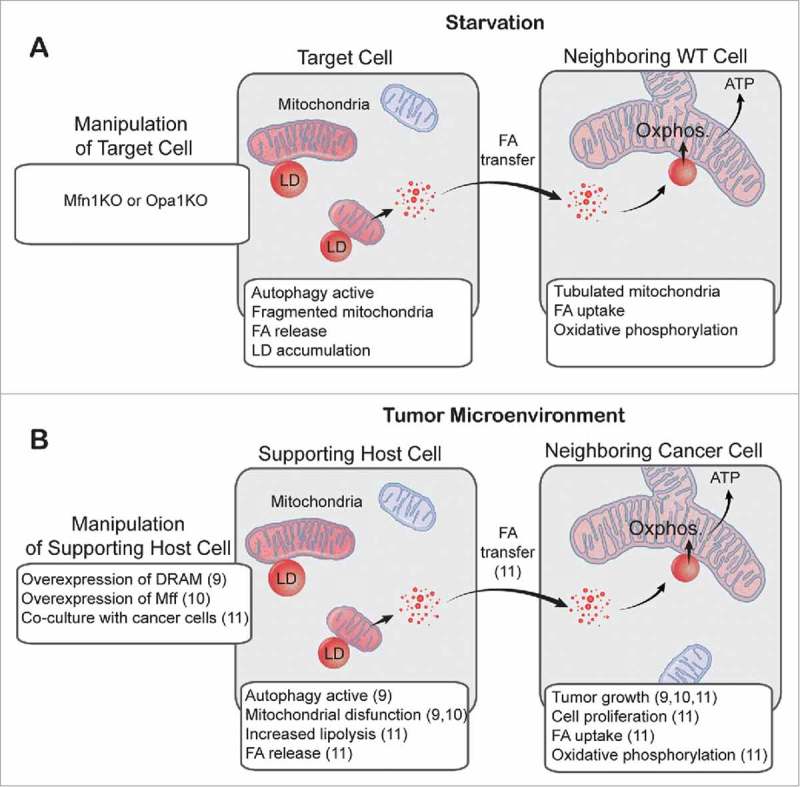
Intra- and intercellular fatty acid trafficking in starved cells and in the tumor microenvironment. (A) We showed that in starved cells with impaired mitochondrial fusion (mitofusin 1 knockout [Mfn1 KO] and optic atrophy 1 knockout [Opa1 KO]), fragmented mitochondria received highly variable amounts of fatty acids (FAs). These FAs were not efficiently metabolized, resulting in increased FA storage within lipid droplets (LDs) and transfer of FAs to neighboring wild-type (WT) cells. (B) This is analogous to a 2-compartment model of tumor metabolism, in which upregulated autophagy, fragmentation of mitochondria, or increased lipolysis in supporting host cells promotes tumor growth, presumably through intercellular transfer of FAs. Numbers in parentheses refer to references. Oxphos., oxidative phosphorylation; DRAM, damage-regulated autophagy modulator; Mff, mitochondrial fission factor.
FA trafficking is likely to be important in a variety of disease contexts, including cancer. The system we have described bears striking similarities to a recently proposed 2-compartment model of tumor metabolism (Fig. 1B). In this model, catabolism in supporting host cells results in transfer of metabolites (including FAs) to cancer cells, supporting the anabolic growth of tumors.6 Many of the features that we observed in starved Mfn1 KO and Opa1 KO cells—autophagy, fragmented mitochondria, and intercellular transfer of FAs—are likely to be important in the context of the tumor microenvironment. For example, one study showed that tumor growth was enhanced when cancer-associated fibroblasts (CAFs) with upregulated autophagy were co-injected with breast cancer cells,7 whereas another showed similar results when mitochondrial fission factor (Mff) was overexpressed in CAFs.8 In a different system, when ovarian cancer cells were co-cultured with supporting adipocytes, lipolysis was induced in the adipocytes, FAs were transferred from adipocytes to cancer cells, and β−oxidation was upregulated in the cancer cells, supporting cancer cell proliferation.9 Our results predict a build-up of LDs in catabolic supporting host cells, but this remains to be determined. In addition, in situations where cancer cells rely heavily on β−oxidation9, we expect mitochondria to be tubulated. However, this is likely to be context-specific; for example, it has recently been shown that metastatic breast cancer cells have more fragmented mitochondria than non-metastatic breast cancer cells.10 Clearly, in the future it will be important to investigate the co-ordination of multiple metabolic processes, including FA trafficking, autophagy, and mitochondrial fission/fusion dynamics, in both cancer cells and the tumor micro-environment.
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Funding Statement
HHS | NIH | National Institute of Child Health and Human Development (NICHD) 1ZIAHD00160921
References
- 1.Rambold AS, Cohen S, Lippincott-Schwartz J. Fatty acid trafficking in starved cells: regulation by lipid droplet lipolysis, autophagy, and mitochondrial fusion dynamics. Dev Cell 2015; 32(6):678-92; PMID:25752962 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zechner R, Zimmermann R, Eichmann TO, Kohlwein SD, Haemmerle G, Lass A, Madeo F. FAT SIGNALS–lipases and lipolysis in lipid metabolism and signaling. Cell Metab 2012; 15:279-91; PMID:22405066; https://doi.org/ 10.1016/j.cmet.2011.12.018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Singh R, Kaushik S, Wang Y, Xiang Y, Novak I, Komatsu M, Tanaka K, Cuervo AM, Czaja MJ. Autophagy regulates lipid metabolism. Nature 2009; 458:1131-5; PMID:19339967; https://doi.org/ 10.1038/nature07976 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Unger RH, Clark GO, Scherer PE, Orci L. Lipid homeostasis, lipotoxicity and the metabolic syndrome. Bioch Biophys Acta 2010; 1801:209-14; PMID:19948243; https://doi.org/ 10.1016/j.bbalip.2009.10.006 [DOI] [PubMed] [Google Scholar]
- 5.Rambold AS, Kostelecky B, Elia N, Lippincott-Schwartz J. Tubular network formation protects mitochondria from autophagosomal degradation during nutrient starvation. Proc Natl Acad Sci U S A 2011; 108:10190-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Martinez-Outschoorn UE, Sotgia F, Lisanti MP. Power surge: supporting cells “fuel” cancer cell mitochondria. Cell Metab 2012; 15:4-5; PMID:22225869; https://doi.org/ 10.1016/j.cmet.2011.12.011 [DOI] [PubMed] [Google Scholar]
- 7.Salem AF, Whitaker-Menezes D, Lin Z, Martinez-Outschoorn UE, Tanowitz HB, Al-Zoubi MS, Howell A, Pestell RG, Sotgia F, Lisanti MP. Two-compartment tumor metabolism: autophagy in the tumor microenvironment and oxidative mitochondrial metabolism (OXPHOS) in cancer cells. Cell Cycle 2012; 11:2545-56; PMID:22722266; https://doi.org/ 10.4161/cc.20920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Guido C, Whitaker-Menezes D, Lin Z, Pestell RG, Howell A, Zimmers TA, Casimiro MC, Aquila S, Ando S, Martinez-Outschoorn UE, et al.. Mitochondrial fission induces glycolytic reprogramming in cancer-associated myofibroblasts, driving stromal lactate production, and early tumor growth. Oncotarget 2012; 3:798-810; PMID:22878233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Nieman KM, Kenny HA, Penicka CV, Ladanyi A, Buell-Gutbrod R, Zillhardt MR, Romero IL, Carey MS, Mills GB, Hotamisligil GS, et al.. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nature Med 2011; 17:1498-503; PMID:22037646; https://doi.org/ 10.1038/nm.2492 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Zhao J, Zhang J, Yu M, Xie Y, Huang Y, Wolff DW, Abel PW, Tu Y. Mitochondrial dynamics regulates migration and invasion of breast cancer cells. Oncogene 2013; 32:4814-24; PMID:23128392; https://doi.org/ 10.1038/onc.2012.494 [DOI] [PMC free article] [PubMed] [Google Scholar]