

Abstract
Background: Cerebral autoregulation (CA) is the brain’s ability to always maintain an adequate and relatively constant blood supply, which is often impaired in cerebrovascular diseases. Near-infrared spectroscopy (NIRS) examines oxygenated hemoglobin (OxyHb) in the cerebral cortex. Low- and very low-frequency oscillations ( and to 0.01 Hz) in OxyHb have been proposed to reflect CA.
Aim: To systematically review published results on OxyHb LFOs and VLFOs in cerebrovascular diseases and related conditions measured with NIRS.
Approach: A systematic search was performed in the MEDLINE database, which generated 36 studies relevant for inclusion.
Results: Healthy people have relatively stable LFOs. LFO amplitude seems to reflect myogenic CA being decreased by vasomotor paralysis in stroke, by smooth muscle damage or as compensatory action in other conditions but can also be influenced by the sympathetic tone. VLFO amplitude is believed to reflect neurogenic and metabolic CA and is lower in stroke, atherosclerosis, and with aging. Both LFO and VLFO synchronizations appear disturbed in stroke, while the former is also altered in internal carotid stenosis and hypertension.
Conclusion: We conclude that amplitudes of LFOs and VLFOs are relatively robust measures for evaluating mechanisms of CA and synchronization analyses can show temporal disruption of CA. Further research and more coherent methodologies are needed.
Keywords: cerebral autoregulation, near-infrared spectroscopy, low-frequency oscillations, cerebrovascular diseases, risk of stroke
1. Introduction
Cerebral autoregulation (CA) of cerebral blood flow (CBF) is the process in which the cerebral vasculature maintains a relatively constant blood flow despite changes in perfusion pressure.1 CA is impaired in several neurological diseases.2 The mechanisms and methods of investigating the CA have been a subject of research and discussion ever since CA as a concept was conceived.3 Both large arteries and small arterioles contribute significantly to vascular resistance in the brain and studies have shown that the large extracranial vessels (internal carotid and vertebral) and intracranial pial vessels contribute to around 50% of cerebral vascular resistance (CVR).4 In recent years, some of the most frequently used methods in studying CA in humans have been transcranial Doppler (TCD) and near-infrared spectroscopy (NIRS). TCD assesses blood velocity in major cerebral vessels, whereas NIRS detects cerebral cortical hemoglobin oxygenation and thereby changes in microcirculatory blood volume and flow. Both techniques can examine low-frequent hemodynamic parameters with high temporal resolution, but NIRS offers distinct advantages in being easy to apply, operator-independent and directly measuring hemodynamics in the cortical region of interest.
The aim here was to systematically review studies on low- frequency oscillations (LFOs) and very low-frequency oscillations (VLFOs) measured by NIRS in cerebrovascular diseases and related conditions to assess CA.
2. Background
2.1. Cerebral Autoregulation
The CA ensures that CBF is maintained at a relatively constant level within large variations of arterial blood pressure (ABP) and thus the cerebral perfusion pressure (CPP)1 (Fig. 1):
| (1) |
Fig. 1.

Near-infrared spectroscopy (NIRS)-derived oxygen saturation versus cerebral perfusion pressure (CPP), depicting the cerebral autoregulation curve. With permission from Moerman et al.5 under Creative Commons Attribution License. Original data from Brady et al.6
This serves as a protective measure against ischemia and failure could possibly result in pallor, sweating, confusion, dizziness, or syncope.7,8 In acute brain injury, where the tissue is particularly sensitive, hypoperfusion can lead to progression of the damages due to impaired CA7,8 and several neurologic disorders do exhibit impaired or changed CA.2 In the opposite end of the arterial blood pressure (ABP) spectrum, CA also protects against chronic hypertension and hypertensive encephalopathy.9,10
Several factors influence CA. The myogenic mechanism is a direct effect in smooth muscle due to changes in transmural pressure mostly active in the small vessels of the brain.1,11–13 The endothelium has a prominent role by the release of dilatory and constrictory substances in addition to the direct mechanical response.1,13,14 Though no one single chemical agent has yet been identified, a metabolic response is observed with neuronal activation in local or global areas.1,11,13,15,16 Autonomic nerve fibers richly innervate the cerebral vessels, especially the larger arteries and activation of the sympathetic nervous system (SNS) pushes the limits of the autoregulation plateau up higher and vice versa.1,11,13,17,18 The level of the partial pressure of carbon dioxide in arterial blood () also modifies CA considerably.1,19 Hypercapnia increases CBF by vasodilation, which also narrows the autoregulatory plateau with changes of both the upper and lower CA limit. Hypocapnia decreases CBF by vasoconstriction, but does not affect the lower limit of CA, while uncertainty remains concerning the upper limit. Furthermore, other physiological systems such as the renin–angiotensin–aldosterone system and pharmacologic substances can modify the complex nature of CA. The distribution of resistance within the microvasculature is a contentious area of research and especially the role of pericytes and capillary resistance.20,21
The research on CA has developed in two main fields. Static CA is the determination of the autoregulatory limits of ABP under different relatively steady circumstances. Dynamic CA includes investigations in both spontaneous physiological fluctuations in ABP and CBF, but also the CBF response to sudden changes in ABP, e.g., when changing body position or following deflation of a cuff in order to quickly pool or release blood from one or more extremities. The analysis of low-LFOs is a part of dynamic CA, which we will be reviewing in the following.
2.2. Mayer Waves and Low-Frequency Oscillations
Mayer waves (M-waves) is a phenomenon of slow spontaneous ABP oscillations observed in 1876 by Mayer in rabbits.22 Slower than ABP oscillations of cardiac () and respiratory rhythms ( to 0.5 Hz), these M-waves has different frequencies across species, but about 0.1 Hz when observed in humans (Fig. 2). M-waves are most often termed LFOs. The oscillations in this frequency are believed to reflect sympathetic nervous activity (SNA) as they are enhanced with activation of the SNS and SNA have, therefore, often been attached to the definition of LFOs.23 Their origin is thought to be the vasomotor tone of blood vessels synchronous throughout the body,23,24 but the driving mechanism behind the rhythmicity of the oscillations has not yet been fully established.
Fig. 2.

OxyHb oscillations at cardiac frequency (1 Hz) and at M-wave frequency (0.1 Hz) enhanced with deep breathing at 0.1 Hz. Unpublished data from healthy elderly woman.
2.2.1. Pacemaker theory and baroreceptor reflex theory
The pacemaker theory, primarily based on animal studies, suggests that the central nervous system contains a pacemaker that is responsible for the rhythmicity of LFOs.23 Studies have shown slow SNA and ABP rhythms in the LFO frequency despite the lack of sensory inputs from peripheral structures in animals that have undergone surgical and/or medical denervation.25–28 In tetraplegic humans with traumatic spinal cord lesions, the results have been conflicting.29–31 While one study showed consistent LFOs,31 other studies failed to detect LFOs in some29 or all subjects30 perhaps due to different levels of the spinal lesions or differences in tetraplegia durations as one study proved significantly increased LFOs 6 months after the initial examination.29
The baroreflex theory originates from the work of Guyton and Harris.32 The contribution of the baroreceptor reflex to the genesis of LFOs has since been confirmed in numerous studies of animals undergone surgical sinoaortic baroreceptor denervation33–35 showing either strong attenuation or abolishment of LFOs. Deactivation of the baroreceptor reflex with alpha-adreno blockers shows a similar trend in animals36–38 and humans.39,40 These studies support that the LFOs are caused by vasomotor tone, because alpha-adrenergic antagonists block the sympathetic effect on vasomotion.41
2.2.2. Amplitude
There are only a limited amount of studies focusing on the amplitude of the LFOs. In animal studies the amplitude follows the mean level of SNA when subjected to stimulations altering the sympathetic tone.42–45 Human studies show the same tendency46–50 though only in individuals and not across groups with different SNA levels51 perhaps due to age differences in the vasculature and regulation thereof. The reproducibility of LFO amplitudes is high over short-term, but low over long-term,52 though this could be influenced by several factors including overall stress level.49 It has been suggested that the LFO amplitudes indicates reflectory local myogenic activity53 in the terminal arteriole smooth muscle cells,54 and is influenced by sympathetic control mechanisms.55 It is thought that the local myogenic response deals with small changes in systemic pressure changes, while the SNS responds to larger changes,56 but the exact relationship remains uncertain.
2.3. Very Low-Frequency Oscillations
Oscillations at an even lower frequency have been observed in a range distinctly below LFOs (0.05 to 0.01 Hz). Originally observed in intracranial pressure by Lundberg,57 these oscillations were also seen in ABP and velocity of the medial cerebral artery ()58,59 and termed VLFOs. The oscillations in these different parameters have been shown to be connected and several autoregulatory mechanisms have been proposed to explain this connection.60–64
The origin of VLFOs is thought to be generated from a central pacemaker because of the relation between different oscillating parameters mentioned above, but also because of the interhemispheric synchronicity.59,65–68 Rhythmic changes in breathing and thereby changes in have also been shown to attribute to VLFOs as they occur at about the same frequency () and correlates well with blood-oxygenation-level-dependent (BOLD) signals for CBF both under steady state69,70 and with neural activity.71 CPP values are described to have influence on the amplitude and frequency of VLFOs,72 while they remain relatively independent of changes in ABP. Intaglietta et al.54 proposed that they stem from large arterioles under neurogenic innervation73 and thus sympathetic control.74 In some cases, the VLFOs are divided into two frequency ranges and Stefanovska et al.53 showed that while the upper VLFO range is endothelial independent and probably relies on the neurogenic activity in large arteries, the lower VLFO range is endothelial-related, and therefore, connected to metabolic changes in the microvessels (Table 1).
Table 1.
Physiological ABP oscillations.
| Oscillation | Frequency (Hz) | Factors considered to determine amplitude |
|---|---|---|
| Cardiac | Heart pumping | |
| Respiratory | Respiration changing intrathoracic pressure | |
| LFOs | Local myogenic activity in terminal arteriole | |
| Influenced by sympathetic tone and control mechanisms | ||
| Upper VLFOs | Neurogenic SNS activity in large arteries | |
| Spontaneous modulation of respiration frequency changing level | ||
| Independent of endothelium | ||
| Bottom VLFOs | Metabolic activity in microvessels | |
| Dependent on endothelium |
2.4. Physiological Function of Oscillations
The physiological purpose of the oscillations low in the frequency spectrum remains intangible. Theories of cyclic nitric oxide (NO) release from the endothelium beneficial to organ function75 and a possible nutritive function76 from animal studies have not been confirmed. The possibility remains that the oscillations are an observed epiphenomenon with no functional purpose.77
2.5. LFOs and VLFOs in Evaluating Cerebral Autoregulation
Similarities in existing knowledge of CA and origins of LFOs and VLFOs indicate an association. Giller originally proposed the method of coherence analysis between systemic and cerebral perfusion oscillations in the low-frequency spectrum to evaluate dynamic CA as the oscillations would be altered with intact CA and relatively unaltered with impaired CA.60 Diehl et al.78 contrived the model of dynamic CA being a biologic control system working as a high-pass filter transmitting high-frequent ABP-oscillation unfiltered to CBF, while LFOs are filtered and only passed through to CBF partially. Considering this, measures of gain and phase shift were suggested to quantify this transmission and thus CA.
Katura et al.79 demonstrated that while an substantial amount of the cerebral vessel LFOs can be attributed to systemic LFOs, more than half cannot, indicating that the origin of cerebral LFO may lie in the autoregulation of CBF rather than systemic cardiovascular regulation.
While the high-pass filter model is not perfect, it does imply two important features of LFOs that correlate with CA: the amplitude and the synchronization. Especially, the latter has been analyzed in different ways as healthy people exhibit highly synchronized oscillations as an expression of well-functioning central origin mechanisms and thereby intact CA. Feasibly, the amplitude on the other hand could in part be ascribed to the magnitude of autoregulatory mechanisms.
2.6. Investigating Cerebral Low-Frequency Oscillations
Spontaneous LFOs in cerebral vessels can be examined through methods such as ultrasonic TCD 60 and NIRS,80 but they can also be accentuated by stimulations of the same frequency.78,80 Both methods are equipped to examine the cerebral blood circulation over time with high temporal resolution and can be applied under various conditions and stimulations that cannot be examined by more confined imaging modalities such as BOLD-magnetic resonance imaging (MRI). While TCD estimates the velocity in larger cerebral arteries such as the MCA, NIRS investigates the cerebral tissue including all vascular compartments though quantitatively reflecting the microvasculature.81 The TCD method has dominated the CA research over NIRS, despite TCD being challenged with certain assumptions and limitations. To assess CBF from measurements of , it is assumed that the large arteries have a constant diameter while studies have shown that the opposite is the case.82,83 Additionally, 5% to 20% of the population cannot be assessed properly due to a poor acoustic window.84 Although NIRS has other limitations and assumptions, at least the method offers a direct measure of local CBF.
The method is simple to understand in general.81 Infrared light of certain wavelengths between 650 and 950 nm is passed from the skin through the extracranial layers and the cortex of the brain. The light passes through human tissue being absorbed by water, fat, melanin, and both oxygenated (OxyHb) and deoxygenated hemoglobin (DeoxyHb). On the skin, adjacent to the light source, detectors are that pick up the infrared signal emanating from the body. Since the levels of water, fat, and melanin remain constant, the signal can then be calculated into dynamic OxyHb and DeoxyHb concentrations by a modified version of the Beer–Lamberts law. To filter the extracranial signals from the intracranial signals, researchers often use multiple light detectors with different distances from the light source and separate the signals from each other to get the purest intracranial signal possible85 (Fig. 3).
Fig. 3.

NIRS with short separation.
In the current study, we will address the NIRS modality called continuous wave, which is not appropriate to calculate an exact hemoglobin concentration as this comes with certain assumptions. Rather, the signal is used to follow the dynamics of the hemoglobin concentration and thereby the CBF in the illumined portion of the cortex. NIRS cannot be used to examine deeper parts of the brain.86
2.7. Data Analysis
Spectral analysis is needed to convert the raw NIRS signal into oscillation parameters in the frequency domain. Only linear models will be considered here, though nonlinear models have shown comparable results.87 There are two common ways of doing the spectral analysis of the NIRS signals: the Fourier transform and the wavelet transform.
Time-domain analyses such as cerebral oximetry (Cox) index,6 mean velocity (Mx) index,88 and tissue oxygenation (Tox) index 89 will not be discussed in this paper as these analyses differ too greatly from frequency-domain analysis to be compared directly, despite being correlated to a certain degree.90
2.7.1. Fourier transform
The Fourier transform is commonly used to decompose a function of time, i.e., the NIRS signal, into a frequency domain. There are many different variations of the Fourier transform. We will address a couple of them here, but will not be discussing them further in this paper. One of the variations can transform the NIRS signals in the time domain, but this will not account for specific frequency intervals, and therefore, is not suitable for analyzing LFOs and VLFOs. The frequency domain analysis generates certain frequency intervals, in which the oscillations occur and the amplitudes of the oscillations in these frequency intervals. A variation of this that is often used is called the fast Fourier transform. It generates a measure called power spectral density (PSD), which is the amplitude within a given signal in relation to the frequency of the signal. The PSD and the amplitude do not, however, say anything about the synchronicity of the oscillations.
2.7.2. Transfer function analysis
The transfer function analysis (TFA) based on the Fourier transform is used to detect synchronization of LFOs and VFLOs and the method has been passed on from the TCD research paradigm.60,78,91 It is a black box input–output analysis, generating ratios of the input signal that is transferred to the output signal. In this context, the input can either be the ABP, mean arterial pressure, or , and the output being the NIRS signals. The analysis produces two certain measures, gain and phase shift. The gain is the amplitude of the input oscillations relative to the amplitude of the output NIRS oscillations. Equal amplitudes would, therefore, generate a gain of 1. The phase shift (or phase angle) is a measure of synchronization and if the oscillations occur at the exact same time the phase shift would be 0 deg. If phase shift is negative, the input oscillations occur before the output oscillations and vice versa, while a counterphase relationship would generate a phase shift of 180 deg.
The input in a TFA can also be the NIRS signal of a healthy hemisphere, which might serve as the best comparison if the individual subjects have a healthy hemisphere.
2.7.3. Wavelet transform
The wavelet transform differs from the Fourier in that it transforms data into a time-frequency domain, whereas Fourier can only transform data into either frequency or time domain one at a time. This gives the transform some slight mathematical benefits and some other possibilities. The downside is a trade-off between spectral and temporal resolution. The wavelet transform can, therefore, generate the amplitude of examined oscillations, but also an instantaneous phase.92
In the NIRS oscillation research, the most commonly used concepts are wavelet coherence (WCO) and wavelet phase coherence (WPCO). WCO is a coherence determination of both the amplitude and the relative phase shift at once. The WCO is difficult to understand in the context of LFOs and VLFOs as it is hard to dissociate the importance the amplitude and the phase shift to the WCO. Instead, WPCO determines how constant the phase difference is between to signals in a certain frequency range. A WPCO of 1 means that the phase difference between two oscillations constant over the entire time series, whereas a WPCO of 0 would only be generated from two totally independent oscillating signals.
Amplitude and phase of Fourier and wavelet transform can be compared without conversion unlike synchronization analysis (phase shift and WPCO).93 To our knowledge, the only studies using the two methods on the same group of patients are conducted by Tachtsidis et al.24 and Rowley et al.55 showing similar trends in PSD, while it is difficult to assess it quantitatively from the publicized results. Other methods are described thoroughly and compared in an excellent review by Thewissen et al.94 regarding NIRS investigations of CA in neonates.
3. Methods
3.1. Search Strategy
A systematic search of literature was conducted in MEDLINE database through May 1, 2018. The search strategy was a combination of the NIRS and oscillations concepts. Broad terms were used to minimize the risk of missing any articles. The full search strategy is depicted in Table 2.
Table 2.
Search strategy.
| AND | ||||
|---|---|---|---|---|
| NIRS concept | Oscillations concept | Filters | ||
| “Spectroscopy, NIR” [MeSH] OR NIRS OR “NIR spectroscopy” OR “Cerebral oximetry” OR “Cerebral oxygenation” OR “Cerebral oxygen saturation” |
Oscillation(s) OR Fluctuation(s) OR “M-wave(s)” OR M-wave(s) OR Vasomotion OR LFO(s) OR VLFO(s) |
OR | Fourier OR Wavelet OR “Transfer function” OR Transform AND Amplitude(s) OR Gain OR “PSD” OR PSD OR Phase(s) OR Phase shift OR WPCO |
Humans [MeSH terms] AND Adults [MeSH terms] AND English language [MeSH terms] |
The search resulted in 176 potentially relevant articles. In addition, the identified articles’ reference lists were reviewed to detect any relevant missing articles. This search resulted in two additional articles. All 178 potentially relevant articles were screened for relevance according to the following criteria.
3.2. Inclusion and Exclusion Criteria
To be included, studies had to use NIRS to examine cerebral LFOs or VLFOs with frequency domain analysis in cerebrovascular diseases or related conditions preceding them including aging. Studies of healthy populations were also included as an indicator of normal variations, though articles focusing on technical or analytical aspects were excluded. Traumatic brain injury patients and intraoperative monitoring were excluded. Only original peer-reviewed studies presenting own results were included leading to the exclusion of reviews. The process is depicted in Fig. 4.
Fig. 4.
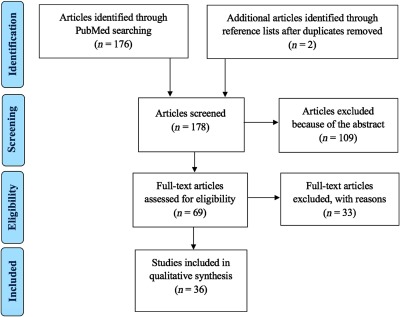
PRISMA flow diagram.
4. Results and Discussion
Thirty-six studies were included and reviewed. Studies were divided into four categories based on study population: healthy subjects, cerebral diseases with structural brain damage, symptomatic cerebrovascular disease with increased risk of further cerebrovascular damage, and asymptomatic populations with increased risk of cerebrovascular disease. Some studies overlapped these categories.
In the following, only OxyHb data will be included in the discussion. The DeoxyHb data conceivably have interesting findings, especially in functional activation studies and when comparing to BOLD-MRI measurements. However, the observed DeoxyHb-changes are mainly a reflection of the venous compartments, whereas OxyHb represents all vascular compartments.95 Additionally, the dynamic changes in OxyHb are closer related to the vascular compartments where CA is most active,6,96 and the OxyHb signals are more robust and also more reliable in the topical frequency spectrum97 making it the most suitable for evaluating CA. The transform parameters have been discussed previously.
Due to the heterogeneity of technical and analytical methods, no quantitative measures are listed in this section. Rather, results are presented as comparative measures as all studies in this area of research use some sort of control group. Most studies had a healthy control group, while others compared different stages of a disease or one hemisphere to the contralateral.
4.1. Healthy Subjects
In a seminal study, Reinhard et al.98 measured LFOs in ABP, and OxyHb in a healthy elderly population. The LFOs preceded the ABP LFOs, which were again slightly ahead of OxyHb LFOs. All LFOs were highly correlated with each other and so the phase shifts merely represent a temporal lag between which vessels the oscillations are measured from Fig. 5. This finding was later confirmed in healthy young people by Philip et al.,99 who additionally showed equal phase shifts across gender, between left and right hemispheres, and over time. Neither visual,80 respiratory,99,100 motoric,101 nor positional stimulations24 have caused any significant changes in the synchronization of LFOs. However, desynchronization has been shown with poor sleep quality in healthy elderly102 and with sleep deprivation in healthy young people.103
Fig. 5.

Physiological phase relationship between oscillations in ABP and different cerebral hemodynamic parameters. Schematic illustration of oscillations. . . With permission from Reinhard et al.98
In contrast, the amplitude of OxyHb LFOs is sensitive to stimulations and increases with sympathetic stimulations,24,100 deep breathing,100 and poor sleep,102 diminishes with hypercapnia,80 while visual stimulations did not affect them. No difference has been observed across gender or between hemispheres, but the day-to-day amplitude ratio seems to fluctuate considerably.99
Very little corresponding knowledge has been obtained about the normal variance of OxyHb VLFOs.
4.2. Cerebral Diseases with Structural Brain Damage
The studies in Table 3 were identified.
Table 3.
Included studies of cerebral diseases with structural brain damage.
| Authors | Population | Signals | Oscillations | Frequency range (Hz) | Analyses |
|---|---|---|---|---|---|
| Phillip et al.105 | Acute stroke | OxyHb | LFO | 0.09 to 0.11 | Amplitude and synchronization |
| Li et al.104 | Chronic stroke | OxyHb | LFO | 0.04 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.04 | ||||
| Han et al.112 | Chronic stroke | OxyHb | LFO | 0.052 to 0.145 | Synchronization |
| Upper VLFO | 0.021 to 0.052 | ||||
| Han et al.113 | Chronic stroke | OxyHb | LFO | 0.052 to 0.145 | Synchronization |
| Upper VLFO | 0.021 to 0.052 | ||||
| Tan et al.114 | Chronic stroke | OxyHb | LFO | 0.052 to 0.145 | Synchronization |
| Upper VLFO | 0.021 to 0.052 | ||||
| Bottom VLFO | 0.0095 to 0.021 | ||||
| Schroeter et al.107 | CMA | OxyHb | LFO | 0.07 to 0.12 | Amplitude |
| Upper VLFO | 0.01 to 0.05 | ||||
| van Beek et al.108 | AD | LFO | 0.07 to 0.13 | Amplitude and synchronization | |
| OxyHb | Upper VLFO | 0.02 to 0.07 |
4.2.1. Amplitude
In ischemic stroke occurring more than 12 months ago, the amplitude of both LFOs and VLFOs was shown to be lower than in healthy age-matched controls.104 Phillip et al.105 investigated LFOs of acute ischemic strokes within 5 days of ictus, but did not examine a healthy control group. Thrombolysed patients, conceivably having suffered less damage to their brain, exhibited amplitudes equal to that of nonthrombolysed patients. The absolute amplitude ratio was borderline different between the two group and amplitude did not correlate with NIHSS on admission. According to the prevailing oscillation theories, decreased amplitudes would indicate a lower mean SNA or less myogenic (LFO) as well as neurogenic and metabolic (VLFO) autoregulatory activity. The former does not correlate with the common understanding of the autonomic nervous system (ANS) after stroke, where the SNS is believed to be overactive and dominating over the relatively inert parasympathetic nervous system.106 However, the smooth muscle loses its tone after a stroke, which is an effect known as vasomotor paralysis.1 The ability to contract is a prerequisite for the SNS to raise LFO amplitude not being met due to the stroke. The decline in myogenic CA activity is an indication of the vasomotor paralysis rendering the brain vulnerable to further damage and could be permanent as it can be observed at least 12 months at minimum after a stroke.
Schroeter et al.107 found a decreased LFO amplitude in patients with cerebral microangiopathy (CMA), though only due to hypertension. The amplitude of VLFOs in this study increased with visual stimulation supporting the theory of VLFO amplitude as in indication of possible metabolic control mechanism (though the applied range was an aggregate of metabolic and neurogenic VLFO frequencies). As CMA patients have damaged small vessels, one would have expected altered VLFO amplitude compared to the control group as the metabolic control occurs in the small vessels, but this was not the case possibly due to the aggregate VLFO range applied.
Newly diagnosed Alzheimer’s (AD) patients exhibited a higher LFO amplitude at rest in a study by van Beek et al.108 The increased myogenic activity could be explained by a higher mean SNA level as shown in studies of the ANS in AD.109–111 The difference in LFO amplitude vanished when the groups performed a repetitive sit–stand maneuver possibly because of the SNS responding to large ABP-changes in healthy controls. VLFO amplitude was equivalent in the two groups under both conditions expressing equal neurogenic and metabolic CA activity.
A TFA of these patients showed that gain between and OxyHb VLFOs was increased compared to controls, which authors proposed as an indicator of reduced metabolic reserve or a reduced diffusion of oxygen as observed in positron emission tomography studies of AD. The LFO -OxyHb gain amplitude was similar across the two groups.
4.2.2. Synchronization
Several studies have explored the LFOs in resting patients with a chronic cerebral infarction and all found a lower interhemispheric WPCO in stroke patients suggesting a lesser stability of the phase difference between the hemispheres compared to healthy people.112–114 This effect was interpreted as a loss of the control on myogenic CA activity in the smooth muscle of resistance vessels. Such a desynchronization was also seen in acute stroke as evidenced by a higher absolute interhemispheric phase shift.105 Interestingly, the interhemispheric phase shift proved to be well correlated to the National Institutes of Health Stroke Scale (NIHSS) and so the impaired CA was due to stroke size and not to treatment (thrombolysis or not).
VLFOs in the upper range exhibited lower interhemispheric WPCO in chronic stroke than control groups in two studies,112,114 while another showed an equal WPCO though with a smaller sample size.113 Lower WPCO would indicate a disruption of spontaneous neurogenic CA activity. Tan et al.114 also inspected the oscillations in the bottom VLFO range, which also had a lower WPCO and thus a reduced synchronicity of the metabolic control mechanism.
AD patients displayed a higher -OxyHb phase shift in the VLFO range and authors explained this as differences in either active regulation mechanisms or in passive properties in the cerebral vasculature.108 They also found an equal phase shift in the LFO range suggesting intact coordination of myogenic CA.
4.3. Symptomatic Diseases with Increased Risk of Further Damage
The studies in Table 4 were identified.
Table 4.
Included studies of symptomatic diseases with increased risk of further damage.
| Authors | Population | Signals | Oscillations | Frequency range (Hz) | Analyses |
|---|---|---|---|---|---|
| Reinhard et al. 98 | Unilateral CS | ABP | LFO | Synchronization | |
| OxyHb | |||||
| Phillip et al. 115 | Symptomatic CS | ABP | LFO | 0.09 to 0.11 | Amplitude and synchronization |
| OxyHb | |||||
| Reinhard et al.124 | Unilateral CS | ABP | LFO | 0.095 to 0.105 | Synchronization |
| OxyHb | |||||
| Oldag et al. 126 | Severe unilateral MCA stenosis | ABP | LFO | 0.08 to 0.12 | Synchronization |
| OxyHb | |||||
| He et al. 117 | Moyamoya | OxyHb | LFO | 0.06 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.06 | ||||
| Bottom VLFO | 0.0095 to 0.02 | ||||
| Schytz et al. 119 | FHM ± common migraine | OxyHb | LFO | 0.09 to 0.11 | Amplitude |
| Schytz et al.120 | OSA | OxyHb | LFO | 0.05 to 0.15 | Amplitude |
| Kolyva et al.122 | Malaria | OxyHb | LFO | 0.04 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.04 |
Studies of migraine, obstructive sleep apnea (OSA), and malaria were included in the review, though the risk association to cerebrovascular disease is not strong enough to draw any definite conclusions. However, these diseases have cerebrovascular traits that make them interesting in this context.
4.3.1. Amplitude
Symptomatic carotid stenosis (CS) patients were examined by Phillip et al.,115 which revealed equal gain between ABP and OxyHb LFOs when comparing to both the contralateral hemisphere and the hemispheres of healthy controls. However, an interhemispheric TFA was performed and showed a higher amplitude ratio due to lower amplitude on the hypoperfusion side. This indicates lower myogenic activity in the microvasculature distal to the stenosis as an expression of compensatory dilation due to inadequate perfusion and oxygen delivery.
Moyamoya disease is characterized by intimal proliferation and stenosis of both the internal carotid arteries as well as its intracranial branches thereby creating the need for collateral angiogenesis looking like a dust cloud on angiography.116 The disease was inspected with NIRS, which showed lower amplitude in the LFO range and bottom VFLO range.117 In accordance with histological studies proving smooth muscle degeneration as the intima grows expansively,118 this result points to a decrease in spontaneous smooth muscle activity.
Schytz et al.119 examined both patients with familial hemiplegic migraine (FHM), common migraine, and OSA.120 Patients suffering from FHM combined with common migraine had induced attacks with infusion of glyceryl trinitrate, and this was accompanied by higher LFO amplitude possibly caused by a reflectory increase in SNA and smooth muscle tone. OSA patients were examined before and 2 months after initiation of continuous positive airway pressure (CPAP) treatment. Oddly, OSA patients did not exhibit any difference to controls in LFO amplitude before CPAP despite OSA usually raising the general sympathetic tone121 and thus the smooth muscle activity, but perhaps this was disrupted by endothelial damage also associated with OSA120. However, they displayed a significant decline after treatment, reflecting the expected modulation of sympathetic activity.
In malaria patients, no difference in LFO amplitude was detected between cerebral and noncerebral malaria.122 Upper VLFO amplitude, however, was lower in cerebral malaria perhaps demonstrating the changes in microvasculature flow prompted by the parasite.123 Authors state an increase in VLFO amplitude after recovery, but with no mention of which patients were included in the follow-up analysis.
4.3.2. Synchronization
CS has been under scrutiny by TFA due to the inherent nature of the disease. Reinhard et al.98 were first to investigate it with NIRS as the output measure. Compared to both healthy controls and the contralateral hemisphere, respiratory amplified LFOs in and OxyHb were delayed relative to ABP LFOs, but only the phase shift to the former was statistically significant. The normal counterphase relationship of OxyHb–DeoxyHb was also abrogated. These changes point to a desynchronization and thus impaired CA. Results from such patients must be carefully treated, as it is difficult to dissociate the effect of CS from other risk factors and infarction, though only a minor portion had any symptoms in this study. Authors later solidified their findings in an examination using a multichannel NIRS system, which also showed CA being mostly impaired in the MCA/ACA (anterior cerebral artery) border zone.124
Similarly, Philip et al.115 arrived at a borderline altered phase shift when comparing to the contralateral hemisphere in resting symptomatic CS patients also demonstrating an impaired CA, but no difference compared to healthy controls. Using interhemispheric OxyHb TFA, authors presented an altered absolute interhemispheric phase shift with borderline significance. Interhemispheric measures seem more intuitively correct as it would minimize anatomical variations125 despite the noteworthy limitation of needing a healthy contralateral hemisphere to compare with.
The trend also occurs in severe unilateral MCA stenosis although the examined patients were asymptomatic. Increased LFO ABP-OxyHb phase shift was observed in the affected hemisphere over the entire hemisphere and even higher in the core MCA distribution area.126 A subgroup was examined for CVR, which showed that diminished CVR led to higher phase shifts. The desynchronization of myogenic CA could thus be the consequence of the downstream vessels not being able to compensate for the MCA stenosis.
4.4. Asymptomatic Conditions with Increased Risk of Cerebrovascular Disease
The studies in Table 5 were identified.
Table 5.
Included studies of asymptomatic conditions with increased risk of further damage.
| Author | Population | Signals | Oscillations | Frequency range (Hz) | Analyses |
|---|---|---|---|---|---|
| Li et al.128 | Risk of stroke − elevated | OxyHb | LFO | 0.06 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.06 | ||||
| Bottom VLFO | 0.005 to 0.02 | ||||
| Li et al.129 | Hypertension ± elevated | OxyHb | LFO | 0.06 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.06 | ||||
| Bottom VLFO | 0.005 to 0.02 | ||||
| Li et al.138 | Hypertension | OxyHb | LFO | 0.05 to 0.15 | Synchronization |
| Upper VLFO | 0.02 to 0.05 | ||||
| Zeller et al.130 | Elderly ± MCI | OxyHb | LFO | 0.07 to 0.11 | Amplitude |
| Schroeter et al.131 | Elderly | OxyHb | LFO | 0.07 to 0.11 | Amplitude |
| Upper VLFO | 0.01 to 0.05 | ||||
| Peng et al.136 | Elderly | HR | LFO | 0.0625 to 0.125 | Amplitude and synchronization |
| ABP | |||||
| OxyHb | |||||
| Philip et al.99 | Elderly | ABP | LFO | 0.09 to 0.11 | Amplitude and synchronization |
| OxyHb | |||||
| Li et al.132 | Elderly | OxyHb | LFO | 0.06 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.06 | ||||
| Bottom VLFO | 0.0095 to 0.02 | ||||
| Vermeij et al.135 | Elderly | ABP | LFO | 0.07 to 0.2 | Amplitude and synchronization |
| OxyHb | Upper VLFO | 0.02 to 0.07 | |||
| Oudegeest-Sander et al.137 | Elderly | LFO | 0.07 to 0.2 | Amplitude and synchronization | |
| OxyHb | Upper VLFO | 0.02 to 0.07 | |||
| Cui et al.140 | Elderly | ABP | LFO | 0.05 to 0.15 | Synchronization |
| OxyHb | Upper VLFO | 0.02 to 0.05 | |||
| Bottom VLFO | 0.0095 to 0.02 | ||||
| Gao et al.139 | Elderly | ABP | LFO | 0.05 to 0.15 | Synchronization |
| OxyHb | Upper VLFO | 0.02 to 0.05 | |||
| Bottom VLFO | 0.0095 to 0.02 | ||||
| Song et al.133 | Elderly | OxyHb | LFO | 0.06 to 0.15 | Amplitude |
| Upper VLFO | 0.02 to 0.06 | ||||
| Bottom VLFO | 0.01 to 0.02 | ||||
| Tan et al. 134 | Elderly | OxyHb | LFO | 0.052 to 0.145 | Amplitude and synchronization |
| Upper VLFO | 0.021 to 0.052 | ||||
| Bottom VLFO | 0.0095 to 0.021 | ||||
| Wang et al.141 | Elderly | OxyHb | LFO | 0.052 to 0.145 | Synchronization |
| Upper VLFO | 0.021 to 0.052 | ||||
| Bottom VLFO | 0.0095 to 0.021 |
4.4.1. Amplitude
Elevated has been recognized as a measure of intracerebral atherosclerosis, and therefore, a risk factor for stroke.127 People with this condition displayed lower amplitude in LFO, upper and bottom VFLO ranges, though the former two were borderline significant.128 This provides indication of decreases in myogenic, neurogenic, and metabolic activity of the cerebral vasculature with elevated , which could all be attributed to the stiffening of vessel walls in atherosclerosis.
An important factor in the development of atherosclerosis is arterial hypertension, in which Li et al.129 examined with half of the hypertension subjects also exhibiting elevated . Hypertension increased the LFO amplitude as the result of myogenic autoregulatory mechanisms being activated to protect the brain in conform to general CA knowledge.1 The group with elevated had amplitudes between that of the hypertension group and the healthy controls. This dampening of amplitude consolidates the findings in the former study of elevated .128 Noteworthy is the opposite trend of LFO amplitude in CMA patients as the result of hypertension107 possibly elucidating that CMA patients do not have the ability to increase their myogenic CA in response to the challenge of hypertension. VLFO amplitudes did not show any definite trends in hypertension.
LFO amplitude has been shown to decrease with age in numerous studies with subjects both at rest130,131,132,133,134 and with stimulations of visual,131 cognitive,135 and positional character.136,133 Generally, this has been interpreted as an expression of vessel stiffening with age, and therefore, less microvascular smooth muscle activity. This trend was even more pronounced in elderly with mild cognitive impairment (MCI), though only in the parietal lobes.130 TFA showed no difference in LFO ABP-OxyHb gain across age groups during rest99,135 or cognitive memory task.135
VLFO amplitude in aging exhibited a similar decline132 although more apparent with stimulations131,135,133 displaying an age effect on neurogenic and metabolic CA. TFA of ABP-OxyHb VLFO during the cognitive test by Vermeij et al.135 showed no effect of age or cognitive load on gain.
During sit–stand maneuvers performed in a study by Oudegeest-Sander et al.,137 authors found no difference in -OxyHb gain across age groups, but a trend toward higher gain was conveyed in their regression analysis. Reduced distensibility and thereby less damping of oscillations in the elderly were proposed to account for this difference.
4.4.2. Synchronization
The interhemispheric WPCO approach was applied to hypertension patients and resulted in lower LFO WPCO and equal WPCO in the upper VLFO range.138 This loss of synchronicity was connected to a reduced control of the microvascular smooth muscle activity. The stroke study by Han et al.112 examined a subpopulation also suffering from hypertension and found an even lower WPCO in the upper VLFO range suggesting additional desynchronization of neurogenic CA activity, while hypertension did not desynchronize LFOs any further.
The synchronization of oscillations in the aging brain has been well examined. Peng et al.136 used a synchronicity analysis called wavelet cross correlation (WCC, higher values indicating stronger synchronicity). It showed that the WCC for HR-OxyHb, ABP-OxyHb, and -OxyHb was equal in the LFO range when subjects remained at supine rest but increased substantially more in young people with head-up tilt test and with active standing in another study.139 This implies an increase in synchronization between systemic and CBF under sympathetic challenges such as positional changes and that this mechanism is impaired in the elderly.
ABP-OxyHb LFO phase analyses have disclosed equal phase shifts in young and elderly under rest99,135 and during cognitive testing.135 Wavelet studies have come to the same conclusion, as the ABP-OxyHb WPCO was homogenous across age groups at rest.140,139 Meanwhile, interhemispheric analyses found lower WPCO with aging indicating desynchronization of myogenic CA at rest.134,141
Oudegeest-Sander et al.137 examined the VLFOs during a sit-stand maneuver and their regression analysis showed a trend toward higher -OxyHb phase shift with age explained by increased vascular tortuosity. The WPCO between VLFOs of ABP and OxyHb in the elderly has also been investigated. At rest synchronization of neurogenic CA activity was higher in the elderly perhaps to compensate for the decline in myogenic CA activity.140,139 During active standing, this difference evened out and both the neurogenic and metabolic CA synchronization was equivalent to the young.139 Interhemispheric WPCO analysis also showed intact synchronization of metabolic and neurogenic activity.134
5. Limitations
Before making any definitive conclusions based on the reviewed material, several limitations must be acknowledged. The penetration depth of infrared light prevents NIRS from ever examining deeper parts of the brain than the available cortex and can never be used to determine whether the observations are reflecting global hemispheric trends or local phenomena despite the use of multichannel systems. In general, the reviewed examinations have applied an interoptode distance of 3 to 5 cm to get the deepest penetration possible, but with no coherency across studies. Although signals from the skin and skull could pollute the cortical signal, the method of short separation optodes rectifies this shortcoming in most cases.
NIRS assumes that the infrared light passes through tissue with a constant spread and that only hemoglobin absorbs the light, which are both reasonable. The emanating signal is comprised of the entire tissue and all the different vascular compartments, which is hard to separate and only recently made conceivable with NIRS.142 The infrared light in the spectrum utilized has a wavelength between 650 and 950 nm, but there is no unified agreement, in which exact wavelengths are most appropriate. Because the relative absorption of OxyHb and DeoxyHb changes with the wavelength, there is a difference between the measured chromophores and thus the examined vascular compartment when using different wavelengths.95,143 NIRS also assumes that the measured hemoglobin concentration is the homogenous in the illuminated tissue, which is not necessarily the case, especially when local pathology is involved.
Processing NIRS signals can be done in a variety of ways to remove motion artefacts and while every process has its advantages, none have proved to be superior to others. The technique is more sensitive to motion artefacts than it is credited for, which will either limit the possible functional stimulations or raise the requirements for postprocessing.
The spectral analysis of NIRS signals can either be performed with Fourier or wavelet transforms and while the two methods are mathematically equivalent, the outcomes in the synchronization analysis cannot be compared directly. Also the differences in technical setup, postprocessing, and spectral analyses render any sort of metaanalysis impossible.
The oscillation analysis method is quite demanding in several ways. The nature of LFOs requires rather long recordings to build a significant data foundation as LFOs only occur 6 times per minute and VLFOs 1 to 3 times per minute. The analysis itself is quite complex and commands skilled personnel and time. Also, as shown in Tables 3–5, there is a need for coherency in the applied frequency ranges. These factors constitute a substantial obstacle of monitoring for immediate changes in CA and thus the clinical implementation.
In this paper, both spontaneous oscillations and oscillations enhanced with stimulations of appropriate frequency have been included. Debate remains between scientists as to which method is better.144 While spontaneous oscillations are claimed to have higher signal-to-noise ratio, this could be accounted for in the analysis and provide a reasonable method for evaluating CA in patients where stimulations can be challenging to perform, e.g., stroke or other cerebrovascular diseases.
Average population size in NIRS oscillation studies is not particularly large and the need for larger-scale investigations is obvious in order to solidify the current knowledge and showcase the possible clinical utilization. However, the technique suffers somewhat due to the perpetual technical and analytical development quickly causing results to be viewed as either outdated or inappropriate for comparison with other studies. It also makes the continuous examination of healthy control groups imperative. There have not yet been established any quantitative measure of intact or impaired CA, which also leaves power calculations impossible.
Caution should be advised in the oscillation research, as investigation of too many oscillating chromophores and NIRS parameters could potentially be more misleading than progressive. The absence of focus in both study design and outcome is dangerous when dealing with relatively small sample sizes. Additionally, the term “altered oscillations” is often applied in discussions of results across different parameters and should be avoided. Each parameter should be carefully analyzed separately as the current study has attempted.
More research is clearly needed both in experimental and clinical studies to improve our knowledge of the oscillations in general and expand the pathophysiological understanding of cerebrovascular diseases and conditions that lead to them. The method would benefit greatly from more coherent technical and analytical procedures, while advances in these areas could also provide the basis for implementation in clinical settings.
6. Conclusion
CA is a complex phenomenon ensuring adequate perfusion to the brain by myogenic, neural, metabolic, and possibly other mechanisms. The modulation of systemic oscillations in the low- and very low-frequency spectrum measured in cerebral vessels is thought to be an expression of the exerted autoregulation. Near-infrared spectroscopy can be used to examine regional cerebral perfusion and thus evaluate CA in cerebrovascular diseases and related conditions. The outcome of LFO amplitude is relatively robust in quantifying the myogenic CA from the smooth muscle cells in the microvasculature, which is lower in stroke, atherosclerosis, and with aging, but must be interpreted with great care as it can be affected by sympathetic activity as in hypertension. LFO synchronization analyses of oscillations have shown disruption of normally well-coordinated autoregulatory actions in stroke, internal carotid, and MCA stenosis as well as in hypertension in accordance with other measurement techniques, but the optimal comparison remains uncertain. VLFO amplitudes suggest lower metabolic and neurogenic CA in stroke, atherosclerosis, and aging, but are generally not as consistent as LFO amplitude, in part due to incoherent frequency ranges. VLFOs are desynchronized in stroke, intact in hypertension, and with aging. Although the outcomes can never stand alone, they seem able to enhance our knowledge of CA. Despite certain limitations, the oscillation analysis of NIRS data could be a valuable tool to both researchers and physicians in the future. Additional research and more coherent methodologies are needed.
Biographies
Adam Vittrup Andersen is a physician at the Department of Neurology, Rigshospitalet in Copenhagen and PhD student at the Unit of Clinical Stroke Research, Rigshospitalet and the University of Copenhagen. He has written his MS thesis on LFOs in cerebrovascular diseases and is currently performing clinical studies on stroke patients with NIRS. His major scientific fields of interest include cerebrovascular disease and hemodynamics, near-infrared spectroscopy, functional neuroimaging, and cerebral autoregulation.
Sofie Amalie Simonsen is a physician in the neurologist educational program at the Department of Neurology, Rigshospitalet in Copenhagen and PhD student at the University of Copenhagen. She has authored and coauthored several papers focusing on cerebrovascular disease. She has been doing stroke research since 2014, and in 2018 she will defend her PhD thesis on stroke classification, breathing-related sleep disorders, cognitive impairment, and depression in stroke.
Henrik Winther Schytz is a consultant at the Danish Headache Center, Rigshospitalet in Copenhagen and an associate professor at the University of Copenhagen. Has authored and coauthored more than 50 publications focusing on headache, cerebrovascular disease and cerebral hemodynamics. He has received the Harold Wolff–John Graham award as recognition for his substantial contribution to headache research. He has done numerous clinical studies with NIRS, which has been the basis of his doctoral dissertation.
Helle Klingenberg Iversen is a head of the stroke unit at Rigshospitalet in Copenhagen and associate professor at the University of Copenhagen. She has authored and coauthored more than 130 publications focusing on clinical and translational cerebrovascular research; stroke—subgroups, sleep disturbances in stroke, impact of naturalistic lighting on long-term hospitalized stroke patients, NIRS, organization of stroke service and patient involvement in stroke care planning. She is a chairman of the Danish Stroke Society.
Disclosures
The authors declare that there is no conflict of interest.
References
- 1.Paulson O. B., Strandgaard S., Edvinsson L., “Cerebral autoregulation,” Cerebrovasc. Brain Metab. Rev. 2(2), 161–92 (1990). [PubMed] [Google Scholar]
- 2.Donnelly J., Aries M. J., Czosnyka M., “Further understanding of cerebral autoregulation at the bedside: possible implications for future therapy,” Expert Rev. Neurother. 15, 169–185 (2015). 10.1586/14737175.2015.996552 [DOI] [PubMed] [Google Scholar]
- 3.Lassen N. A., “Cerebral blood flow and oxygen consumption in man,” Physiol. Rev. 39, 183–238 (1959). 10.1152/physrev.1959.39.2.183 [DOI] [PubMed] [Google Scholar]
- 4.Faraci F. M., Heistad D. D., “Regulation of large cerebral arteries and cerebral microvascular pressure,” Circ. Res. 66, 8–17 (1990). 10.1161/01.RES.66.1.8 [DOI] [PubMed] [Google Scholar]
- 5.Moerman A., De Hert S., “Recent advances in cerebral oximetry. Assessment of cerebral autoregulation with near-infrared spectroscopy: myth or reality?” F1000Res. 6, 1615 (2017). 10.12688/f1000research [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Brady K. M., et al. , “Continuous time-domain analysis of cerebrovascular autoregulation using near-infrared spectroscopy,” Stroke 38, 2818–2825 (2007). 10.1161/STROKEAHA.107.485706 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Chillon J. M., Baumbach G. L., “Autoregulation of cerebral blood flow,” in Cerebrovascular Diseases, 1st ed., Welch K., et al., Eds., pp. 51–54, Academic Press, San Diege, California: (1997). [Google Scholar]
- 8.Folino A. F., “Cerebral autoregulation and syncope,” Prog. Cardiovasc. Dis. 50, 49–80 (2007). 10.1016/j.pcad.2007.01.001 [DOI] [PubMed] [Google Scholar]
- 9.Lassen N. A., Agnoli A., “The upper limit of autoregulation of cerebral blood flow on the pathogenesis of hypertensive encephalopathy,” Scand. J. Clin. Lab. Invest. 30, 113–116 (1972). 10.3109/00365517209081099 [DOI] [PubMed] [Google Scholar]
- 10.Phillips S. J., Whisnant J. P., “Hypertension and the brain,” Arch. Intern. Med. 152, 938–945 (1992). 10.1001/archinte.1992.00400170028006 [DOI] [PubMed] [Google Scholar]
- 11.Donnelly J., et al. , “Regulation of the cerebral circulation: bedside assessment and clinical implications,” Crit. Care 20, 129 (2016). 10.1186/s13054-016-1293-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Folkow B., “Description of the myogenic hypothesis,” Circ. Res. 15(Suppl 1), 279–287 (1964). [PubMed] [Google Scholar]
- 13.Peterson E. C., Wang Z., Britz G., “Regulation of cerebral blood flow,” Int. J. Vasc. Med. 2011, 1–8 (2011). 10.1155/2011/823525 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rubanyi G. M., “Endothelium-derived relaxing and contracting factors,” J. Cell Biochem. 46, 27–36 (1991). 10.1002/(ISSN)1097-4644 [DOI] [PubMed] [Google Scholar]
- 15.Kontos H. A., Wei E. P., “Oxygen-dependent mechanisms in cerebral autoregulation,” Ann. Biomed. Eng. 13, 329–334 (1985). 10.1007/BF02584251 [DOI] [PubMed] [Google Scholar]
- 16.Aaslid R., et al. , “Cerebral autoregulation dynamics in humans,” Stroke 20, 45–52 (1989). 10.1161/01.STR.20.1.45 [DOI] [PubMed] [Google Scholar]
- 17.Hamner J. W., Tan C. O., “Relative contributions of sympathetic, cholinergic, and myogenic mechanisms to cerebral autoregulation,” Stroke 45, 1771–1777 (2014). 10.1161/STROKEAHA.114.005293 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Seifert T., Secher N. H., “Sympathetic influence on cerebral blood flow and metabolism during exercise in humans,” Prog. Neurobiol. 95, 406–426 (2011). 10.1016/j.pneurobio.2011.09.008 [DOI] [PubMed] [Google Scholar]
- 19.Meng L., Gelb A. W., “Regulation of cerebral autoregulation by carbon dioxide,” Anesthesiology 122, 196–205 (2015). 10.1097/ALN.0000000000000506 [DOI] [PubMed] [Google Scholar]
- 20.Gould I. G., et al. , “The capillary bed offers the largest hemodynamic resistance to the cortical blood supply,” J. Cereb. Blood Flow Metab. 37, 52–68 (2017). 10.1177/0271678X16671146 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hall C. N., et al. , “Capillary pericytes regulate cerebral blood flow in health and disease,” Nature 508, 55–60 (2014). 10.1038/nature13165 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Mayer S., “Studien zur physiologie des herzens und der blutgefässe,” Sitz Kaiser Akad Wiss 74, 281–307 (1876). [Google Scholar]
- 23.Julien C., “The enigma of Mayer waves: facts and models,” Cardiovasc. Res. 70, 12–21 (2006). 10.1016/j.cardiores.2005.11.008 [DOI] [PubMed] [Google Scholar]
- 24.Tachtsidis I., et al. , “Investigation of cerebral haemodynamics by near-infrared spectroscopy in young healthy volunteers reveals posture-dependent spontaneous oscillations,” Physiol. Meas. 25, 437–445 (2004). 10.1088/0967-3334/25/2/003 [DOI] [PubMed] [Google Scholar]
- 25.Grasso R., et al. , “Arterial baroreceptors are not essential for low frequency oscillation of arterial pressure,” J. Auton. Nerv. Syst. 50, 323–331 (1995). 10.1016/0165-1838(94)00103-Q [DOI] [PubMed] [Google Scholar]
- 26.Montano N., et al. , “Effects of spinal section and of positive-feedback excitatory reflex on sympathetic and heart rate variability,” Hypertension 36, 1029–1034 (2000). 10.1161/01.HYP.36.6.1029 [DOI] [PubMed] [Google Scholar]
- 27.Montano N., et al. , “Presence of vasomotor and respiratory rhythms in the discharge of single medullary neurons involved in the regulation of cardiovascular system,” J. Auton. Nerv. Syst. 57, 116–122 (1996). 10.1016/0165-1838(95)00113-1 [DOI] [PubMed] [Google Scholar]
- 28.Preiss G., Polosa C., “Patterns of sympathetic neuron activity associated with Mayer waves,” Am. J. Physiol. 226, 724–730 (1974). 10.1152/ajplegacy.1974.226.3.724 [DOI] [PubMed] [Google Scholar]
- 29.Guzzetti S., et al. , “Influences of neural mechanisms on heart period and arterial pressure variabilities in quadriplegic patients,” Am. J. Physiol. 266, H1112–H1120 (1994). 10.1152/ajpcell.1994.266.4.C1112 [DOI] [PubMed] [Google Scholar]
- 30.Inoue K., et al. , “Power spectral analysis of blood pressure variability in traumatic quadriplegic humans,” Am. J. Physiol. 260, H842–H847 (1991). 10.1152/ajpcell.1991.260.3.C658 [DOI] [PubMed] [Google Scholar]
- 31.Koh J., et al. , “Human autonomic rhythms: vagal cardiac mechanisms in tetraplegic subjects,” J. Physiol. 474, 483–495 (1994). 10.1113/jphysiol.1994.sp020039 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Guyton A. C., Harris J. W., “Pressoreceptor-autonomic oscillation; a probable cause of vasomotor waves,” Am. J. Physiol. 165, 158–166 (1951). 10.1152/ajplegacy.1951.165.1.158 [DOI] [PubMed] [Google Scholar]
- 33.Cerutti C., Barres C., Paultre C., “Baroreflex modulation of blood pressure and heart rate variabilities in rats: assessment by spectral analysis,” Am. J. Physiol. 266, H1993–H2000 (1994). 10.1152/ajpheart.1994.266.5.H1993 [DOI] [PubMed] [Google Scholar]
- 34.Jacob H. J., et al. , “Spectral analysis of arterial pressure lability in rats with sinoaortic deafferentation,” Am. J. Physiol. 269, R1481–R1488 (1995). 10.1152/ajpregu.1995.269.6.R1481 [DOI] [PubMed] [Google Scholar]
- 35.Julien C., et al. , “Hemodynamic analysis of arterial pressure oscillations in conscious rats,” J. Auton. Nerv. Syst. 50, 239–252 (1995). 10.1016/0165-1838(94)00095-2 [DOI] [PubMed] [Google Scholar]
- 36.Barres C., de Souza Neto E. P., Julien C., “Effect of alpha-adrenoceptor blockade on the 0.4 Hz sympathetic rhythm in conscious rats,” Clin. Exp. Pharmacol. Physiol. 28, 983–985 (2001). 10.1046/j.1440-1681.2001.03561.x [DOI] [PubMed] [Google Scholar]
- 37.Japundzic N., et al. , “Spectral analysis of blood pressure and heart rate in conscious rats: effects of autonomic blockers,” J. Auton. Nerv. Syst. 30, 91–100 (1990). 10.1016/0165-1838(90)90132-3 [DOI] [PubMed] [Google Scholar]
- 38.Rubini R., et al. , “Power spectrum analysis of cardiovascular variability monitored by telemetry in conscious unrestrained rats,” J. Auton. Nerv. Syst. 45, 181–190 (1993). 10.1016/0165-1838(93)90050-5 [DOI] [PubMed] [Google Scholar]
- 39.Cevese A., et al. , “Baroreflex and oscillation of heart period at 0.1 Hz studied by alpha-blockade and cross-spectral analysis in healthy humans,” J. Physiol. 531, 235–244 (2001). 10.1111/tjp.2001.531.issue-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.van de Borne P., et al. , “Contrasting effects of phentolamine and nitroprusside on neural and cardiovascular variability,” Am. J. Physiol. Heart Circ. Physiol. 281, H559–H565 (2001). 10.1152/ajpheart.2001.281.2.H559 [DOI] [PubMed] [Google Scholar]
- 41.Kimmerly D. S., et al. , “Circulating norepinephrine and cerebrovascular control in conscious humans,” Clin. Physiol. Funct. Imaging 23, 314–319 (2003). 10.1046/j.1475-0961.2003.00507.x [DOI] [PubMed] [Google Scholar]
- 42.Barres C., Cheng Y., Julien C., “Steady-state and dynamic responses of renal sympathetic nerve activity to air-jet stress in sinoaortic denervated rats,” Hypertension 43, 629–635 (2004). 10.1161/01.HYP.0000115384.01463.61 [DOI] [PubMed] [Google Scholar]
- 43.Bertram D., et al. , “Differential responses of frequency components of renal sympathetic nerve activity to arterial pressure changes in conscious rats,” Am. J. Physiol. Regul. Integr. Comp. Physiol. 289, R1074–R1082 (2005). 10.1152/ajpregu.00270.2005 [DOI] [PubMed] [Google Scholar]
- 44.Janssen B. J., et al. , “Frequency-dependent modulation of renal blood flow by renal nerve activity in conscious rabbits,” Am. J. Physiol. 273, R597–R608 (1997). 10.1152/ajpregu.1997.273.2.R597 [DOI] [PubMed] [Google Scholar]
- 45.Malpas S. C., et al. , “Contribution of renal nerves to renal blood flow variability during hemorrhage,” Am. J. Physiol. 274, R1283–R1294 (1998). 10.1152/ajpcell.1998.274.5.C1283 [DOI] [PubMed] [Google Scholar]
- 46.Castiglioni P., et al. , “Mechanisms of blood pressure and heart rate variability: an insight from low-level paraplegia,” Am. J. Physiol. Regul. Integr. Comp. Physiol. 292, R1502–R1509 (2007). 10.1152/ajpregu.00273.2006 [DOI] [PubMed] [Google Scholar]
- 47.Cooke W. H., et al. , “Human responses to upright tilt: a window on central autonomic integration,” J. Physiol. 517(Pt 2), 617–628 (1999). 10.1111/tjp.1999.517.issue-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Furlan R., et al. , “Oscillatory patterns in sympathetic neural discharge and cardiovascular variables during orthostatic stimulus,” Circulation 101, 886–892 (2000). 10.1161/01.CIR.101.8.886 [DOI] [PubMed] [Google Scholar]
- 49.Lucini D., et al. , “Hemodynamic and autonomic adjustments to real life stress conditions in humans,” Hypertension 39, 184–188 (2002). 10.1161/hy0102.100784 [DOI] [PubMed] [Google Scholar]
- 50.Pagani M., et al. , “Relationship between spectral components of cardiovascular variabilities and direct measures of muscle sympathetic nerve activity in humans,” Circulation 95, 1441–1448 (1997). 10.1161/01.CIR.95.6.1441 [DOI] [PubMed] [Google Scholar]
- 51.Taylor J. A., et al. , “Low-frequency arterial pressure fluctuations do not reflect sympathetic outflow: gender and age differences,” Am. J. Physiol. 274, H1194–H1201 (1998). [DOI] [PubMed] [Google Scholar]
- 52.van de Borne P., et al. , “Relationship between repeated measures of hemodynamics, muscle sympathetic nerve activity, and their spectral oscillations,” Circulation 96, 4326–4332 (1997). 10.1161/01.CIR.96.12.4326 [DOI] [PubMed] [Google Scholar]
- 53.Stefanovska A., Bracic M., Kvernmo H. D., “Wavelet analysis of oscillations in the peripheral blood circulation measured by laser Doppler technique,” IEEE Trans. Biomed. Eng. 46, 1230–1239 (1999). 10.1109/10.790500 [DOI] [PubMed] [Google Scholar]
- 54.Intaglietta M., “Vasomotion and flowmotion: physiological mechanisms and clinical evidence,” Vasc. Med. 1, 101–112 (1990). 10.1177/1358836x9000100202 [DOI] [Google Scholar]
- 55.Rowley A. B., et al. , “Synchronization between arterial blood pressure and cerebral oxyhaemoglobin concentration investigated by wavelet cross-correlation,” Physiol. Meas. 28, 161–173 (2007). 10.1088/0967-3334/28/2/005 [DOI] [PubMed] [Google Scholar]
- 56.Harper A. M., et al. , “The influence of sympathetic nervous activity on cerebral blood flow,” Arch. Neurol. 27, 1–6 (1972). 10.1001/archneur.1972.00490130003001 [DOI] [PubMed] [Google Scholar]
- 57.Lundberg N., “Continuous recording and control of ventricular fluid pressure in neurosurgical practice,” Acta Psychiatr. Scand. Suppl. 36, 1–193 (1960). [PubMed] [Google Scholar]
- 58.Mautner-Huppert D., et al. , “B-waves in healthy persons,” Neurol. Res. 11, 194–196 (1989). 10.1080/01616412.1989.11739891 [DOI] [PubMed] [Google Scholar]
- 59.Newell D. W., et al. , “The relationship of blood flow velocity fluctuations to intracranial pressure B waves,” J. Neurosurg. 76, 415–421 (1992). 10.3171/jns.1992.76.3.0415 [DOI] [PubMed] [Google Scholar]
- 60.Giller C. A., “The frequency-dependent behavior of cerebral autoregulation,” Neurosurgery 27, 362–368 (1990). 10.1227/00006123-199009000-00004 [DOI] [PubMed] [Google Scholar]
- 61.Magnaes B., “Body position and cerebrospinal fluid pressure. Part 1: clinical studies on the effect of rapid postural changes,” J. Neurosurg. 44, 687–697 (1976). 10.3171/jns.1976.44.6.0687 [DOI] [PubMed] [Google Scholar]
- 62.Piper I. R., et al. , “An experimental study of cerebrovascular resistance, pressure transmission, and craniospinal compliance,” Neurosurgery 32, 805–816, discussion 15–16 (1993). 10.1227/00006123-199305000-00014 [DOI] [PubMed] [Google Scholar]
- 63.Rosner M. J., Becker D. P., “Origin and evolution of plateau waves. Experimental observations and a theoretical model,” J. Neurosurg. 60, 312–324 (1984). 10.3171/jns.1984.60.2.0312 [DOI] [PubMed] [Google Scholar]
- 64.Steinmeier R., et al. , “Slow rhythmic oscillations of blood pressure, intracranial pressure, microcirculation, and cerebral oxygenation. Dynamic interrelation and time course in humans,” Stroke 27, 2236–2243 (1996). 10.1161/01.STR.27.12.2236 [DOI] [PubMed] [Google Scholar]
- 65.Einhäupl K. M., et al. , “Oscillations of ICP related to cardiovascular parameters,” in Intracranial Pressure VI, Miller J. D., Rowan J. O., Eds., pp. 290–297, Springer-Verlag, Berlin, Heidelberg, (1986). [Google Scholar]
- 66.Hashimoto M., et al. , “Respiratory and cardiovascular oscillations during B-waves,” in Intracranial Pressure VII, Hoff J. T., Betz A. L., Eds., pp. 217–279, Springer, Berlin, Heidelberg: (1989). [Google Scholar]
- 67.Higashi S., et al. , “The role of vasomotor center and adrenergic pathway in B-waves,” in Intracranial Pressure VII, Hoff J. T., Betz A. L., Eds., pp. 220–224, Springer, Berlin, Heidelberg: (1989). [Google Scholar]
- 68.Venes J. L., “B waves—a reflection of cardiorespiratory or cerebral nervous systems rhythm?” Childs Brain 5, 352–360 (1979). 10.1159/000119831 [DOI] [PubMed] [Google Scholar]
- 69.Birn R. M., et al. , “Separating respiratory-variation-related fluctuations from neuronal-activity-related fluctuations in fMRI,” Neuroimage 31, 1536–1548 (2006). 10.1016/j.neuroimage.2006.02.048 [DOI] [PubMed] [Google Scholar]
- 70.Wise R. G., et al. , “Resting fluctuations in arterial carbon dioxide induce significant low frequency variations in BOLD signal,” Neuroimage 21, 1652–1664 (2004). 10.1016/j.neuroimage.2003.11.025 [DOI] [PubMed] [Google Scholar]
- 71.Peng T., et al. , “The effects of respiratory fluctuations in the resting-state BOLD signal differ between eyes open and eyes closed,” Magn. Reson. Imaging 31, 336–345 (2013). 10.1016/j.mri.2012.06.013 [DOI] [PubMed] [Google Scholar]
- 72.Spiegelberg A., Preuß M., Kurtcuoglu V., “B-waves revisited,” Interdiscip. Neurosurg. 6, 13–17 (2016). 10.1016/j.inat.2016.03.004 [DOI] [Google Scholar]
- 73.Farkas E., Luiten P. G. M., “Cerebral microvascular pathology in aging and Alzheimer’s disease,” Prog. Neurobiol. 64, 575–611 (2001). 10.1016/S0301-0082(00)00068-X [DOI] [PubMed] [Google Scholar]
- 74.Zhang R., et al. , “Autonomic neural control of dynamic cerebral autoregulation in humans,” Circulation 106, 1814–1820 (2002). 10.1161/01.CIR.0000031798.07790.FE [DOI] [PubMed] [Google Scholar]
- 75.Nafz B., Wagner C. D., Persson P. B., “Endogenous nitric oxide buffers blood pressure variability between 0.2 and 0.6 Hz in the conscious rat,” Am. J. Physiol. 272(2), H632–H637 (1997). 10.1152/ajpheart.1997.272.2.H632 [DOI] [PubMed] [Google Scholar]
- 76.Rucker M., et al. , “Vasomotion in critically perfused muscle protects adjacent tissues from capillary perfusion failure,” Am. J. Physiol. Heart Circ. Physiol. 279(2), H550–H558 (2000). 10.1152/ajpheart.2000.279.2.H550 [DOI] [PubMed] [Google Scholar]
- 77.Chapuis B., et al. , “Linear modelling analysis of baroreflex control of arterial pressure variability in rats,” J. Physiol. 559, 639–649 (2004). 10.1113/jphysiol.2004.065474 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Diehl R. R., et al. , “Phase relationship between cerebral blood flow velocity and blood pressure. A clinical test of autoregulation,” Stroke 26, 1801–1804 (1995). 10.1161/01.STR.26.10.1801 [DOI] [PubMed] [Google Scholar]
- 79.Katura T., et al. , “Quantitative evaluation of interrelations between spontaneous low-frequency oscillations in cerebral hemodynamics and systemic cardiovascular dynamics,” Neuroimage 31(4), 1592–1600 (2006). 10.1016/j.neuroimage.2006.02.010 [DOI] [PubMed] [Google Scholar]
- 80.Obrig H., et al. , “Spontaneous low frequency oscillations of cerebral hemodynamics and metabolism in human adults,” Neuroimage 12, 623–639 (2000). 10.1006/nimg.2000.0657 [DOI] [PubMed] [Google Scholar]
- 81.Strangman G., Boas D. A., Sutton J. P., “Non-invasive neuroimaging using near-infrared light,” Biol. Psychiatry. 52(7), 679–693 (2002). 10.1016/S0006-3223(02)01550-0 [DOI] [PubMed] [Google Scholar]
- 82.Thomsen L. L., Iversen H. K., “Experimental and biological variation of three-dimensional transcranial Doppler measurements,” J. Appl. Physiol. 75, 2805–2810 (1993). 10.1152/jappl.1993.75.6.2805 [DOI] [PubMed] [Google Scholar]
- 83.Hoiland R. L., Ainslie P. N., “CrossTalk proposal: the middle cerebral artery diameter does change during alterations in arterial blood gases and blood pressure,” J. Physiol. 594, 4073–4075 (2016). 10.1113/tjp.2016.594.issue-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sarkar S., et al. , “Role of transcranial Doppler ultrasonography in stroke,” Postgrad. Med. J. 83(985), 683–689 (2007). 10.1136/pgmj.2007.058602 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Gagnon L., et al. , “Further improvement in reducing superficial contamination in NIRS using double short separation measurements,” Neuroimage 85(Pt1), 127–135 (2014). 10.1016/j.neuroimage.2013.01.073 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Obrig H., “NIRS in clinical neurology—a ‘promising’ tool?” Neuroimage 85(Pt1), 535–546 (2014). 10.1016/j.neuroimage.2013.03.045 [DOI] [PubMed] [Google Scholar]
- 87.Chacon M., et al. , “Comparing models of spontaneous variations, maneuvers and indexes to assess dynamic cerebral autoregulation,” Acta Neurochir. Suppl. 126, 159–162 (2018). 10.1007/978-3-319-65798-1 [DOI] [PubMed] [Google Scholar]
- 88.Czosnyka M., et al. , “Monitoring of cerebral autoregulation in head-injured patients,” Stroke 27, 1829–1834 (1996). 10.1161/01.STR.27.10.1829 [DOI] [PubMed] [Google Scholar]
- 89.Steiner L. A., et al. , “Near-infrared spectroscopy can monitor dynamic cerebral autoregulation in adults,” Neurocrit. Care 10, 122–128 (2009). 10.1007/s12028-008-9140-5 [DOI] [PubMed] [Google Scholar]
- 90.Liu X., et al. , “Comparison of frequency and time domain methods of assessment of cerebral autoregulation in traumatic brain injury,” J. Cereb. Blood Flow Metab. 35, 248–256 (2015). 10.1038/jcbfm.2014.192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Zhang R., et al. , “Transfer function analysis of dynamic cerebral autoregulation in humans,” Am. J. Physiol. 274, H233–H241 (1998). 10.1152/ajpheart.1998.274.1.H233 [DOI] [PubMed] [Google Scholar]
- 92.Shiogai Y., Stefanovska A., McClintock P. V., “Nonlinear dynamics of cardiovascular ageing,” Phys. Rep. 488, 51–110 (2010). 10.1016/j.physrep.2009.12.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Bruns A., “Fourier-, Hilbert- and wavelet-based signal analysis: are they really different approaches?” J. Neurosci. Methods 137, 321–332 (2004). 10.1016/j.jneumeth.2004.03.002 [DOI] [PubMed] [Google Scholar]
- 94.Thewissen L., et al. , “Measuring near-infrared spectroscopy derived cerebral autoregulation in neonates: from research tool toward bedside multimodal monitoring,” Front. Pediatr. 6, 117 (2018). 10.3389/fped.2018.00117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Strangman G., Franceschini M. A., Boas D. A., “Factors affecting the accuracy of near-infrared spectroscopy concentration calculations for focal changes in oxygenation parameters,” NeuroImage 18, 865–879 (2003). 10.1016/S1053-8119(03)00021-1 [DOI] [PubMed] [Google Scholar]
- 96.Smielewski P., et al. , “Can cerebrovascular reactivity be measured with near-infrared spectroscopy?,” Stroke 26(12), 2285–2292 (1995). 10.1161/01.STR.26.12.2285 [DOI] [PubMed] [Google Scholar]
- 97.Strangman G., et al. , “A quantitative comparison of simultaneous BOLD fMRI and NIRS recordings during functional brain activation,” Neuroimage 17, 719–731 (2002). 10.1006/nimg.2002.1227 [DOI] [PubMed] [Google Scholar]
- 98.Reinhard M., et al. , “Oscillatory cerebral hemodynamics—the macro- vs. microvascular level,” J. Neurol. Sci. 250, 103–109 (2006). 10.1016/j.jns.2006.07.011 [DOI] [PubMed] [Google Scholar]
- 99.Phillip D., et al. , “Low frequency oscillations in cephalic vessels assessed by near infrared spectroscopy,” Eur. J. Clin. Invest. 42, 1180–1188 (2012). 10.1111/eci.2012.42.issue-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Cheng R., et al. , “Noninvasive optical evaluation of spontaneous low frequency oscillations in cerebral hemodynamics,” Neuroimage 62, 1445–1454 (2012). 10.1016/j.neuroimage.2012.05.069 [DOI] [PubMed] [Google Scholar]
- 101.Pfurtscheller G., et al. , “About the stability of phase shifts between slow oscillations Around 0.1 Hz in cardiovascular and cerebral systems,” IEEE Trans. Biomed. Eng. 58, 2064–2071 (2011). 10.1109/TBME.2011.2134851 [DOI] [PubMed] [Google Scholar]
- 102.Bu L., et al. , “Effects of poor sleep quality on brain functional connectivity revealed by wavelet-based coherence analysis using NIRS methods in elderly subjects,” Neurosci. Lett. 668, 108–114 (2018). 10.1016/j.neulet.2018.01.026 [DOI] [PubMed] [Google Scholar]
- 103.Bu L., et al. , “Effects of sleep deprivation on phase synchronization as assessed by wavelet phase coherence analysis of prefrontal tissue oxyhemoglobin signals,” PLoS One 12, e0169279 (2017). 10.1371/journal.pone.0169279 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Li Z., et al. , “Wavelet analysis of cerebral oxygenation signal measured by near infrared spectroscopy in subjects with cerebral infarction,” Microvasc. Res. 80, 142–147 (2010). 10.1016/j.mvr.2010.02.004 [DOI] [PubMed] [Google Scholar]
- 105.Phillip D., et al. , “Spontaneous low frequency oscillations in acute ischemic stroke: a near infrared spectroscopy (NIRS) study,” J. Neurol. Neurophysiol. 5, 1000241 (2014). 10.4172/2155-9562 [DOI] [Google Scholar]
- 106.Dorrance A. M., Fink G., “Effects of stroke on the autonomic nervous system,” Compr. Physiol. 5, 1241–1263 (2015). 10.1002/cphy.c140016 [DOI] [PubMed] [Google Scholar]
- 107.Schroeter M. L., et al. , “Spontaneous slow hemodynamic oscillations are impaired in cerebral microangiopathy,” J. Cereb. Blood Flow Metab. 25, 1675–1684 (2005). 10.1038/sj.jcbfm.9600159 [DOI] [PubMed] [Google Scholar]
- 108.van Beek A. H., et al. , “Oscillations in cerebral blood flow and cortical oxygenation in Alzheimer’s disease,” Neurobiol. Aging 33, 428.e21–428.e31 (2012). 10.1016/j.neurobiolaging.2010.11.016 [DOI] [PubMed] [Google Scholar]
- 109.Aharon-Peretz J., et al. , “Increased sympathetic and decreased parasympathetic cardiac innervation in patients with Alzheimer’s disease,” Arch. Neurol. 49, 919–922 (1992). 10.1001/archneur.1992.00530330041013 [DOI] [PubMed] [Google Scholar]
- 110.de Vilhena Toledo M. A., Junqueira L. F., Jr., “Cardiac sympathovagal modulation evaluated by short-term heart interval variability is subtly impaired in Alzheimer’s disease,” Geriatr. Gerontol. Int. 8, 109–118 (2008). 10.1111/j.1447-0594.2008.00456.x [DOI] [PubMed] [Google Scholar]
- 111.Zakrzewska-Pniewska B., et al. , “Clinical and functional assessment of dysautonomia and its correlation in Alzheimer’s disease,” Am. J. Alzheimers Dis. Other Demen. 27, 592–599 (2012). 10.1177/1533317512459792 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Han Q., et al. , “Phase synchronization analysis of prefrontal tissue oxyhemoglobin oscillations in elderly subjects with cerebral infarction,” Med. Phys. 41, 102702 (2014). 10.1118/1.4896113 [DOI] [PubMed] [Google Scholar]
- 113.Han Q., et al. , “Wavelet coherence analysis of prefrontal tissue oxyhaemoglobin signals as measured using near-infrared spectroscopy in elderly subjects with cerebral infarction,” Microvasc. Res. 95, 108–115 (2014). 10.1016/j.mvr.2014.08.001 [DOI] [PubMed] [Google Scholar]
- 114.Tan Q., et al. , “Frequency-specific functional connectivity revealed by wavelet-based coherence analysis in elderly subjects with cerebral infarction using NIRS method,” Med. Phys. 42, 5391–5403 (2015). 10.1118/1.4928672 [DOI] [PubMed] [Google Scholar]
- 115.Phillip D., et al. , “Altered low frequency oscillations of cortical vessels in patients with cerebrovascular occlusive disease—a NIRS study,” Front. Neurol. 4, 204 (2013). 10.3389/fneur.2013.00204 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Kuroda S., Houkin K., “Moyamoya disease: current concepts and future perspectives,” Lancet Neurol. 7, 1056–1066 (2008). 10.1016/S1474-4422(08)70240-0 [DOI] [PubMed] [Google Scholar]
- 117.He Y., et al. , “Wavelet analysis of cerebral oxygenation oscillations in the screening of Moyamoya disease,” Biomed. Mater. Eng. 24, 3463–3469 (2014). 10.3233/BME-141171 [DOI] [PubMed] [Google Scholar]
- 118.Fukui M., et al. , “Moyamoya disease,” Neuropathology 20(Suppl 1), 61–64 (2000). 10.1046/j.1440-1789.2000.00300.x [DOI] [PubMed] [Google Scholar]
- 119.Schytz H. W., et al. , “Nitric oxide modulation of low-frequency oscillations in cortical vessels in FHM—a NIRS study,” Headache 52, 1146–1154 (2012). 10.1111/hed.2012.52.issue-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Schytz H. W., et al. , “Low-frequency oscillations and vasoreactivity of cortical vessels in obstructive sleep apnea during wakefulness: a near infrared spectroscopy study,” Sleep Med. 14, 416–421 (2013). 10.1016/j.sleep.2012.12.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Kohler M., Stradling J. R., “Mechanisms of vascular damage in obstructive sleep apnea,” Nat. Rev. Cardiol. 7(12), 677–685 (2010). 10.1038/nrcardio.2010.145 [DOI] [PubMed] [Google Scholar]
- 122.Kolyva C., et al. , “Oscillations in cerebral haemodynamics in patients with falciparum malaria,” Adv. Exp. Med. Biol. 765, 101–107 (2013). 10.1007/978-1-4614-4989-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Mishra S. K., Newton C. R., “Diagnosis and management of the neurological complications of falciparum malaria,” Nat. Rev. Neurol. 5, 189–198 (2009). 10.1038/nrneurol.2009.23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Reinhard M., et al. , “Spatial mapping of dynamic cerebral autoregulation by multichannel near-infrared spectroscopy in high-grade carotid artery disease,” J. Biomed. Opt. 19, 097005 (2014). 10.1117/1.JBO.19.9.097005 [DOI] [PubMed] [Google Scholar]
- 125.Muehlschlegel S., et al. , “Feasibility of NIRS in the neurointensive care unit: a pilot study in stroke using physiological oscillations,” Neurocrit. Care 11, 288–295 (2009). 10.1007/s12028-009-9254-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Oldag A., et al. , “Near-infrared spectroscopy and transcranial sonography to evaluate cerebral autoregulation in middle cerebral artery steno-occlusive disease,” J. Neurol. 263, 2296–2301 (2016). 10.1007/s00415-016-8262-5 [DOI] [PubMed] [Google Scholar]
- 127.Bos M. J., et al. “Transcranial Doppler hemodynamic parameters and risk of stroke: the Rotterdam study,” Stroke 38, 2453–2458 (2007). 10.1161/STROKEAHA.107.483073 [DOI] [PubMed] [Google Scholar]
- 128.Li Z., et al. , “Spectral analysis of near-infrared spectroscopy signals measured from prefrontal lobe in subjects at risk for stroke,” Med. Phys. 39, 2179–2185 (2012). 10.1118/1.3696363 [DOI] [PubMed] [Google Scholar]
- 129.Li Z., et al. , “Assessment of cerebral oxygenation oscillations in subjects with hypertension,” Microvasc. Res. 88, 32–41 (2013). 10.1016/j.mvr.2013.04.003 [DOI] [PubMed] [Google Scholar]
- 130.Zeller J. B. M., et al. , “Reduced spontaneous low frequency oscillations as measured with functional near-infrared spectroscopy in mild cognitive impairment,” Brain Imaging Behav. 1–10 (2018). 10.1007/s11682-018-9827-y [DOI] [PubMed] [Google Scholar]
- 131.Schroeter M. L., Schmiedel O., von Cramon D. Y., “Spontaneous low-frequency oscillations decline in the aging brain,” J. Cereb. Blood Flow Metab. 24, 1183–1191 (2004). 10.1097/01.WCB.0000135231.90164.40 [DOI] [PubMed] [Google Scholar]
- 132.Li Z., et al. , “Age-related changes in spontaneous oscillations assessed by wavelet transform of cerebral oxygenation and arterial blood pressure signals,” J. Cereb. Blood Flow Metab. 33, 692–699 (2013). 10.1038/jcbfm.2013.4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Song S., et al. , “Low-frequency oscillations in cerebrovascular and cardiovascular hemodynamics: their interrelationships and the effect of age, Microvasc. Res. 102, 46–53 (2015). 10.1016/j.mvr.2015.08.004 [DOI] [PubMed] [Google Scholar]
- 134.Tan Q., et al. , “Age-related alterations in phase synchronization of oxyhemoglobin concentration changes in prefrontal tissues as measured by near-infrared spectroscopy signals,” Microvasc. Res. 103, 19–25 (2016). 10.1016/j.mvr.2015.10.002 [DOI] [PubMed] [Google Scholar]
- 135.Vermeij A., et al. , “Very-low-frequency oscillations of cerebral hemodynamics and blood pressure are affected by aging and cognitive load,” Neuroimage 85(Pt1), 608–615 (2014). 10.1016/j.neuroimage.2013.04.107 [DOI] [PubMed] [Google Scholar]
- 136.Peng T., et al. , “The effects of age on the spontaneous low-frequency oscillations in cerebral and systemic cardiovascular dynamics,” Physiol. Meas. 29, 1055–1069 (2008). 10.1088/0967-3334/29/9/005 [DOI] [PubMed] [Google Scholar]
- 137.Oudegeest-Sander M. H., et al. , “Assessment of dynamic cerebral autoregulation and cerebrovascular reactivity in ageing by measurements of cerebral blood flow and cortical oxygenation,” Exp. Physiol. 99, 586–598 (2014). 10.1113/expphysiol.2013.076455 [DOI] [PubMed] [Google Scholar]
- 138.Li Z., et al. , “Wavelet coherence analysis of prefrontal oxygenation signals in elderly subjects with hypertension,” Physiol. Meas. 35, 777–791 (2014). 10.1088/0967-3334/35/5/777 [DOI] [PubMed] [Google Scholar]
- 139.Gao Y., et al. , “Cerebral autoregulation in response to posture change in elderly subjects-assessment by wavelet phase coherence analysis of cerebral tissue oxyhemoglobin concentrations and arterial blood pressure signals,” Behav. Brain Res. 278, 330–336 (2015). 10.1016/j.bbr.2014.10.019 [DOI] [PubMed] [Google Scholar]
- 140.Cui R., et al. , “Wavelet coherence analysis of spontaneous oscillations in cerebral tissue oxyhemoglobin concentrations and arterial blood pressure in elderly subjects,” Microvasc. Res. 93, 14–20 (2014). 10.1016/j.mvr.2014.02.008 [DOI] [PubMed] [Google Scholar]
- 141.Wang B., et al. , “Posture-related changes in brain functional connectivity as assessed by wavelet phase coherence of NIRS signals in elderly subjects,” Behav. Brain Res. 312, 238–245 (2016). 10.1016/j.bbr.2016.06.037 [DOI] [PubMed] [Google Scholar]
- 142.Kainerstorfer J. M., Sassaroli A., Fantini S., “Optical oximetry of volume-oscillating vascular compartments: contributions from oscillatory blood flow,” J. Biomed. Opt. 21, 101408 (2016). 10.1117/1.JBO.21.10.101408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Scholkmann F., et al. , “A review on continuous wave functional near-infrared spectroscopy and imaging instrumentation and methodology,” Neuroimage 85(Pt1), 6–27 (2014). 10.1016/j.neuroimage.2013.05.004 [DOI] [PubMed] [Google Scholar]
- 144.Tzeng Y. C., Panerai R. B., “CrossTalk proposal: dynamic cerebral autoregulation should be quantified using spontaneous blood pressure fluctuations,” J. Physiol. 596, 3–5 (2018). 10.1113/JP273899 [DOI] [PMC free article] [PubMed] [Google Scholar]