

Abstract
Chinese materia medica (CMM) are essential components of traditional Chinese medicine, and Chinese medicinal formulas consisting of 2 or more types of CMM are widely used. These formulations have played a pivotal role in health protection and disease control for thousands of years. Guzhi Zengsheng Zhitongwan (GZZSZTW), which represents one of the Chinese medicinal formulations, has been used for several decades to treat joint diseases. However, the exact molecular mechanism underlying its efficacy in treating osteoarthritis remains to be elucidated. In the present study, we investigated the effects of GZZSZTW on primary chondrocytes. We demonstrated that GZZSZTW significantly promoted chondrocyte viability, maintained chondrocytes in a continuous proliferative state, and prevented their further differentiation. These effects were achieved by the synergistic interactions of various herbs and their active components in GZZSZTW through an increase in the expression levels of functional genes participating in chondrocyte commitment and proliferation and a decrease in the expression levels of genes involved in chondrocyte differentiation. GZZSZTW treatment also decreased the expression levels of genes that inhibited chondrocyte proliferation. Thus, this study has greatly deepened the current knowledge about the molecular effects of GZZSZTW on chondrocytes. It has also shed new light on possible strategies to further prevent and treat cartilage-related diseases by using traditional Chinese medicinal formulations.
1. Introduction
It is well-known that Youyou Tu won the 2015 Nobel Prize in Physiology or Medicine for her discoveries concerning a novel therapy for malaria using artemisinin, which is derived from a traditional herbal medicine called qinghaosu. Since then, Chinese materia medica (CMM) has attracted worldwide attention and has extended to the forefront of the global research community [1].
CMM is an essential component of traditional Chinese medicine; it relies on Chinese medicinal formulas consisting of 2 or more types of CMM and has played a pivotal role in health protection and disease control for thousands of years [2]. In other words, CMM have already provided outstanding clinical results, since they have been tested in human experiments with long-term observation periods and in clinical trials [3]. For instance, Guzhi Zengsheng Zhitongwan (GZZSZTW), a Chinese medicinal formulation created by the national medical master professor Bailing Liu, has been used in the Affiliated Hospital of Changchun University of Chinese Medicine for several decades to treat joint diseases such as osteoarthritis. Clinically, this formula is routinely taken in the form of orally administered pills, which are only produced in the hospital's internal preparation center to treat patients, and it is not yet available commercially. Although it is effective and widely used in the treatment of joint diseases in our hospital, the precise molecular mechanism underlying its efficacy in treating osteoarthritis remains to be elucidated.
Osteoarthritis (OA) is the most common degenerative joint and cartilage disease and is associated with pain and disability. However, no suitable treatment for OA exists, because of the poor self-healing capacity of damaged cartilage [4]. Chondrocytes are the only cells residing in cartilage that control cartilage structure and function by regulating extracellular matrix turnover and maintaining cartilage homeostasis [5]. Thus, investigating the effect of the Chinese medicinal formulation GZZSZTW on chondrocytes is therefore very important for further dissecting the molecular mechanism behind its successful clinical treatment of osteoarthritis.
In the present study, we prepared aqueous extracts of GZZSZTW. We isolated primary chondrocytes from neonatal mouse rib cartilage and investigated the effects of GZZSZTW on the proliferation of chondrocytes. We further performed RNA-seq to analyze the gene expression pattern of chondrocytes in response to treatment with GZZSZTW. We demonstrated that GZZSZTW significantly promoted chondrocyte viability, maintained chondrocytes in a continuous proliferative state, and suppressed their further differentiation by increasing the expression levels of genes that promote chondrocyte commitment and proliferation and a decrease in the expression levels of genes that inhibit chondrocyte proliferation and promote chondrocyte differentiation.
2. Materials and Methods
2.1. Preparation of the GZZSZTW Aqueous Extract
GZZSZTW was obtained from the Affiliated Hospital of Changchun University of Chinese Medicine (Changchun, China). The formulation of GZZSZTW consisted of 7 types of CMM, namely, Rehmannia glutinosa (Gaertn.) DC., Spatholobus suberectus Dunn, Epimedium brevicornu Maxim (K.S.Hao), Raphanus sativus L. (Hook. f. & T. Anderson) (baked), Drynaria fortunei (Kunze ex Mett.) J.Sm. (baked), Cynomorium coccineum subsp. songaricum (Rupr.) (J.Léonard), and Cibotium barometz (L.) (J.Sm). The CMM mixture of GZZSZTW was extracted with distilled water by a reflux method and was then filtered through a 0.45-μm Hollow Fiber Cartridge (GE Healthcare, USA). The filtrate was freeze-dried by a Heto PowerDry LL3000 Freeze Dryer (Thermo, USA) and stored at -80°C.
2.2. Isolation of Primary Chondrocytes
Animal experiments were approved by the Ethical Committee for Animal Research of Changchun University of Chinese Medicine. Primary chondrocytes from the rib cartilage of neonatal mice were isolated as previously described [6]. Briefly, cartilage from the rib cages of neonatal C57BL/6J mice was digested for 45 minutes with 3 mg/ml collagenase D and then overnight with 0.5 mg/ml collagenase D (Sigma, USA). The cells were centrifuged and resuspended in DMEM/F12 medium (Thermo, USA) containing 5% FCS (Thermo, USA) and 1% penicillin/streptomycin (Sigma, USA).
2.3. Cell Proliferation Assay
The Cell Counting Kit-8 (CCK-8) (Sigma, USA) was used to evaluate chondrocyte proliferation according to the manufacturer's protocol. Briefly, primary chondrocytes were seeded into 96-well cell culture plates at a density of 5 × 104 cells/ml (100 μl per well) and cultured at 37°C in a 5% CO2 incubator (Thermo, USA) for 4 h. The cell culture medium was then discarded, and each well was rinsed thoroughly with fresh culture medium. The chondrocytes were treated with GZZSZTW at different concentrations (0, 0.2, 0. 4, 0.6, 0.8, and 1.0 mg/ml dissolved in culture medium; 100 μl per well) and subsequently cultured for 24 h. Deer antler extract (DAE) was used as a reference drug and added to the cells in the same way as GZZSZTW. Next, 10 μl of CCK-8 reagent was added, and the cells were incubated for 1 h. The absorbance was measured at 450 nm using an Infinite 200 PRO plate reader (Life Sciences, USA). The cell proliferation rate was calculated as the percentage of cell viability after treatment with GZZSZTW and DAE at different concentrations, respectively.
2.4. RNA Purification and Illumina Sequencing
Primary chondrocytes were isolated as described above, seeded into 6-well cell culture plates at a density of 1 × 105 cells/well, and incubated for 4 h. Chondrocytes were treated with GZZSZTW for 24 h or left untreated. Then, the cell culture medium was discarded, and each well was rinsed thoroughly with cold PBS buffer. Total RNA was isolated using TRIzol (Invitrogen, USA) in accordance with the manufacturer's protocol. RNA integrity was evaluated using the Agilent 2100 Bioanalyzer (Agilent Technologies, USA). Paired-end mRNA libraries were generated using the TruSeq Stranded mRNA kit (Illumina, USA) according to the manufacturer's protocol. High-throughput sequencing of the mRNA libraries was performed on an Illumina HiSeq 2500 platform (Illumina, USA).
2.5. RNA-Seq Data Analysis
After RNA-seq, clean reads were obtained by trimming the raw reads to remove the low-quality reads and adapter sequences. The data sets were deposited in the NCBI Sequence Read Archive (SRA) database under accession number SRP125978. The clean reads were aligned with the mouse (Mus musculus) reference genome using HISAT [7]. The FPKM algorithm was used to measure the gene expression levels [8]. BLAST was used to perform annotations against the nonredundant (NR) and Swiss-Prot protein databases. DEGseq was used to analyze the differentially expressed genes [9]. Genes with a log2 fold change ≥ 1 or ≤ -1 and with a p value ≤ 0.001 were considered to be differentially expressed.
2.6. Quantitative Real-Time PCR (qRT-PCR) Verification
The expression levels of differentially expressed genes were validated by qRT-PCR. Briefly, total RNA was isolated using TRIzol (Invitrogen, USA) in accordance with the manufacturer's protocol. cDNA was synthesized using the iScript cDNA Synthesis Kit (Bio-Rad, USA) and amplified using SsoAdvanced Universal SYBR® Green Supermix (Bio-Rad, USA) on a CFX Connect Real-Time PCR Detection System (Bio-Rad, USA) under standard amplification conditions. The gene expression levels were normalized to the mouse beta-actin gene (Actb) and calculated using the 2−ΔΔCT method [10].
3. Results
3.1. GZZSZTW Promotes Proliferation in Primary Chondrocytes in a Dose-Dependent Manner
The effect of GZZSZTW on chondrocyte proliferation was measured by the CCK-8 assay. Deer antler extract (DAE), which has been shown to promote chondrocyte viability and keep chondrocytes proliferating continuously, while blocking maturation and further differentiation in a dose-dependent manner, was used as a reference drug [11]. As shown in Figure 1, chondrocyte viability was significantly increased in a dose-dependent manner with GZZSZTW treatment compared with the untreated control group (0 mg/ml). Since treatment with GZZSZTW at the concentrations of 0.8 mg/ml and 1.0 mg/ml had a similar effect on chondrocyte viability, a concentration of 0.8 mg/ml GZZSZTW was selected for use in the following experiments.
Figure 1.
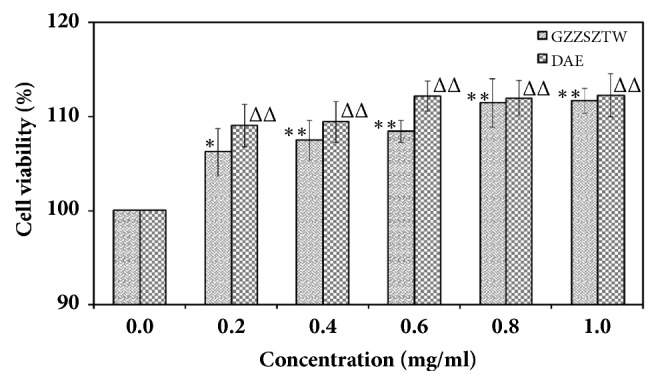
Cell viability assay in primary chondrocytes treated with GZZSZTW and DAE. The CCK-8 kit was used to detect chondrocyte proliferation following treatment with GZZSZTW and DAE at increasing concentrations (0, 0.2, 0.4, 0.6, 0.8, and 1.0 mg/ml) for 24 h, respectively. Cell viabilities were normalized and calculated relative to the untreated group (0 mg/ml). Data are presented as the mean and standard deviation for technical triplicates in an experiment representative of several other independent experiments. ∗ (Δ) represents p <0.01 and ∗∗ (ΔΔ) represents p <0.001, computed by the t-test for the difference in cell viability in response to GZZSZTW and DAE treatment, respectively.
3.2. Sequencing, Genome Mapping, and Functional Annotation
After Illumina sequencing and data processing, 40,958,510 and 40,961,200 clean reads were obtained from primary chondrocytes not treated with GZZSZTW (Blank) and those treated with GZZSZTW, respectively, as shown in Table 1. The quality assessment showed that the Q30 percentages were greater than 94%, and the GC content percentages were approximately 52%. For the Blank and GZZSZTW-treated samples, 36,258,888 and 36,392,470 reads were aligned to the mouse genome, respectively. In total, 15,788 out of 15,908 (Blank) and 15,566 out of 15,676 (GZZSZTW) transcripts were annotated against the nonredundant (NR) NCBI protein database and Swiss-Prot database, respectively.
Table 1.
Statistics for the sequencing and assembly results.
| Statistics | Blank | GZZSZTW |
|---|---|---|
| Clean reads | 40,958,510 | 40,961,200 |
| Q30 percentage | 94.38 | 94.42 |
| GC percentage | 52.31 | 52.49 |
| Total mapped reads | 36,258,888 | 36,392,470 |
| Total transcripts | 15,908 | 15,676 |
| Known transcripts | 15,788 | 15,566 |
3.3. Comparative Analysis of Differentially Expressed Genes
The differential expression analysis identified 229 genes that were significantly differentially expressed between the GZZSZTW-treated and Blank groups (log2 fold change ≥ 1 or ≤ -1 and p ≤0.001), including 139 upregulated genes and 90 downregulated genes (GZZSZTW versus Blank), as shown in Table 2.
Table 2.
Statistical analysis of differentially expressed genes (GZZSZTW versus Blank).
| Statistics | Number |
|---|---|
| Differentially expressed mRNAs | 229 |
| Upregulated mRNAs | 139 |
| Downregulated mRNAs | 90 |
3.4. GZZSZTW Increases the Expression Levels of Multiple Genes That Positively Regulate Chondrocyte Proliferation
Because GZZSZTW was able to promote proliferation in primary chondrocytes, we first analyzed the differentially expressed genes that positively regulated chondrocyte proliferation. Based on the results from the RNA-seq analysis, we identified 10 differentially expressed genes that directly promoted cell proliferation, including Tnfaip2, Chi3l1, Tnf, Pfkfb3, Sox8, Jag1, Mafb, Pla2g7, Hnrnpa1, and E2f3. The expression levels of these genes were significantly increased in response to GZZSZTW treatment, as shown in Table 3.
Table 3.
Differentially expressed genes that positively regulate chondrocyte proliferation.
| Gene name | Blank (FPKM) |
GZZSZTW (FPKM) |
log2 fold change (GZZSZTW /Blank) | p value |
|---|---|---|---|---|
| Tumor necrosis factor alpha-induced protein 2 (Tnfaip2) | 37.69 | 129.86 | 1.78 | 0 |
| Chitinase-3-like protein 1 (Chi3l1) | 40.76 | 122.14 | 1.58 | 1.55E-121 |
| Tumor necrosis factor (Tnf) | 9.18 | 24.24 | 1.40 | 7.31E-21 |
| 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (Pfkfb3) | 9.68 | 22.87 | 1.24 | 1.76E-51 |
| Transcription factor SOX-8 (Sox8) | 2.99 | 15.93 | 2.41 | 1.25E-50 |
| Protein jagged-1 (Jag1) | 6.29 | 13.38 | 1.09 | 3.80E-27 |
| Transcription factor MafB (Mafb) | 5.08 | 10.76 | 1.08 | 6.05E-14 |
| Platelet-activating factor acetylhydrolase (Pla2g7) | 4.33 | 9.88 | 1.19 | 1.02E-08 |
| Heterogeneous nuclear ribonucleoprotein A1 (Hnrnpa1) | 0.01 | 6.60 | 9.37 | 1.87E-21 |
| Transcription factor E2F3 (E2f3) | 2.47 | 5.44 | 1.14 | 2.88E-11 |
3.5. GZZSZTW Treatment Maintains Chondrocyte Proliferation by Decreasing the Expression Levels of Inhibitors of Cell Proliferation
We then analyzed the differentially expressed genes that participated in the inhibition of cell proliferation. In total, the expression levels of 7 genes were significantly decreased in response to GZZSZTW treatment. These genes are typically downregulated when cells are in a proliferative state and play a crucial role in inhibiting cell proliferation, as shown in Table 4.
Table 4.
Differentially expressed genes that negatively regulate chondrocyte proliferation.
| Gene name | Blank (FPKM) |
GZZSZTW (FPKM) |
log2 fold change (GZZSZTW /Blank) | p value |
|---|---|---|---|---|
| Rho-related GTP-binding protein RhoB (Rhob) | 118.64 | 53.33 | -1.15 | 1.51E-97 |
| Dual specificity protein phosphatase 6 (Dusp6) | 18.05 | 7.41 | -1.28 | 1.37E-23 |
| Serine/threonine-protein kinase PLK3 (Plk3) | 16.41 | 6.27 | -1.39 | 3.09E-19 |
| Fibroblast growth factor 21 (Fgf21) | 14.80 | 7.37 | -1.01 | 7.27E-05 |
| Cell cycle checkpoint control protein RAD9A (Rad9a) | 13.54 | 6.37 | -1.09 | 1.81E-10 |
| Filamin A-interacting protein 1-like (Filip1l) | 9.78 | 4.84 | -1.01 | 4.77E-11 |
| Ras-like protein family member 11B (Rasl11b) | 8.48 | 3.96 | -1.10 | 1.45E-06 |
3.6. GZZSZTW Treatment Maintains Chondrocyte Proliferation by Promoting Chondrocyte Proliferation and Suppressing Chondrocyte Differentiation
According to the RNA-seq analysis, the expression levels of pancartilaginous early chondrocyte markers including Sox9, Sox5, Sox6, Acan, Col2a1, Col9a1, Col11a1, Hapln1, and Wwp2 were slightly increased in response to GZZSZTW treatment, as shown in Table 5.
Table 5.
Gene expression levels of pancartilaginous early chondrocyte markers.
| Gene name | Blank (FPKM) |
GZZSZTW (FPKM) |
log2 fold change (GZZSZTW /Blank) | p value |
|---|---|---|---|---|
| Transcription factor SOX-9 (Sox9) | 22.35 | 28.25 | 0.34 | 1.39E-06 |
| Transcription factor SOX-5 (Sox5) | 2.53 | 5.17 | 1.03 | 7.95E-14 |
| Transcription factor SOX-6 (Sox6) | 11.16 | 13.09 | 0.23 | 1.24E-10 |
| Aggrecan core protein (Acan) | 439.94 | 554.62 | 0.33 | 7.55E-177 |
| Collagen alpha-1(II) chain (Col2a1) | 16294.32 | 16870.22 | 0.05 | 3.43E-96 |
| Collagen alpha-1(IX) chain(Col9a1) | 1024.49 | 1188.74 | 0.21 | 4.10E-92 |
| Collagen alpha-1(XI) chain (Col11a1) | 838.24 | 1020.09 | 0.28 | 1.47E-232 |
| Hyaluronan and proteoglycan link protein 1 (Hapln1) | 117.28 | 132.39 | 0.17 | 5.60E-10 |
| NEDD4-like E3 ubiquitin-protein ligase WWP2 (Wwp2) | 140.53 | 166.42 | 0.24 | 2.99E-14 |
Furthermore, the expression levels of proliferating and prehypertrophic chondrocyte markers including Fgfr3, Matn1, Comp, Ptch1, Runx2, and Runx3 were also slightly increased in response to GZZSZTW treatment. However, the expression levels of the major prehypertrophic and hypertrophic chondrocyte markers including Pth1r, Sp7, Ihh, Bmp6, and Ibsp either decreased slightly or exhibited almost no change in response to GZZSZTW treatment, as shown in Table 6.
Table 6.
Gene expression levels of growth plate chondrocyte markers.
| Gene name | Blank (FPKM) |
GZZSZTW (FPKM) |
log2 fold change (GZZSZTW /Blank) | p value |
|---|---|---|---|---|
| Fibroblast growth factor receptor 3 (Fgfr3) | 57.13 | 97.35 | 0.77 | 2.34E-80 |
| Cartilage matrix protein (Matn1) | 102.96 | 124.45 | 0.27 | 1.06E-08 |
| Cartilage oligomeric matrix protein (Comp) | 712.93 | 775.11 | 0.12 | 1.77E-13 |
| Protein patched homolog 1 (Ptch1) | 3.92 | 5.44 | 0.47 | 2.92E-03 |
| Runt-related transcription factor 2 (Runx2) | 7.43 | 10.01 | 0.43 | 3.02E-05 |
| Runt-related transcription factor 3 (Runx3) | 4.68 | 6.26 | 0.42 | 7.23E-03 |
| Parathyroid hormone/parathyroid hormone-related peptide receptor (Pth1r) | 55.23 | 54.53 | -0.02 | 8.96E-01 |
| Transcription factor Sp7 (Sp7) | 3.76 | 4.04 | 0.10 | 4.23E-01 |
| Indian hedgehog protein (Ihh) | 0.57 | 0.51 | -0.16 | 4.99E-01 |
| Collagen alpha-1(X) (Col10a1) | 92.05 | 122.37 | 0.41 | 1.71E-25 |
| Bone morphogenetic protein 6 (Bmp6) | 4.38 | 5.37 | 0.29 | 1.19E-01 |
| Bone sialoprotein 2 (Ibsp) | 78.3 | 72.73 | -0.11 | 1.04E-01 |
3.7. Validation of RNA-Seq Data by qRT-PCR
To validate the accuracy of the RNA-seq results, we selected 8 of the differentially expressed genes (Tnfaip2, Chi3l1, Sox8, Jag1, Rhob, Dusp6, Rad9a, and Filip1l) and verified their expression profiles using qRT-PCR. The specific primers used in this experiment are listed in Table 7.
Table 7.
Primer sequences used in qRT-PCR validation.
| Gene | Primer | Sequence |
|---|---|---|
| Tnfaip2 | Forward primer | AGGAGGAGTCTGCGAAGAAGA |
| Reverse primer | GGCAGTGGACCATCTAACTCG | |
| Chi3l1 | Forward primer | GTACAAGCTGGTCTGCTACTTC |
| Reverse primer | ATGTGCTAAGCATGTTGTCGC | |
| Sox8 | Forward primer | CGAGGGGATACTGCTGAGG |
| Reverse primer | AGCTCTGCGTTATGGAGATGC | |
| Jag1 | Forward primer | CCTCGGGTCAGTTTGAGCTG |
| Reverse primer | CCTTGAGGCACACTTTGAAGTA | |
| Rhob | Forward primer | GTGCCTGCTGATCGTGTTCA |
| Reverse primer | CCGAGAAGCACATAAGGATGAC | |
| Dusp6 | Forward primer | ATAGATACGCTCAGACCCGTG |
| Reverse primer | ATCAGCAGAAGCCGTTCGTT | |
| Rad9a | Forward primer | GGCTGTCCATTCGCTATCCC |
| Reverse primer | GTGGGGCAAAAAGGAAGCAG | |
| Filip1l | Forward primer | AGCACTCAGTCGGCAAATTGA |
| Reverse primer | AGCCTCTTATTGAGGTCTCTGC | |
| Actb | Forward primer | ACCTTCTACAATGAGCTGCG |
| Reverse primer | CTGGATGGCTACGTACATGG |
The relative fold change of each gene was normalized to the internal reference gene Actb. The expression levels of the selected differentially expressed genes measured by qRT-PCR were consistent with the results of the RNA-seq analysis, as shown in Figure 2.
Figure 2.

Gene expression levels of differentially expressed genes validated by qRT-PCR. Gene expression levels for individual genes are presented as the fold change between the GZZSZTW-treated group and the untreated control group.
4. Discussion
GZZSZTW, a Chinese medicinal formulation widely used for treating joint diseases, has been used in the Affiliated Hospital of Changchun University of Chinese Medicine for several decades. However, the precise molecular mechanism underlying the ability of GZZSZTW to treat these diseases remains to be elucidated. In the present study, we investigated the effects of GZZSZTW on primary mouse chondrocytes using state-of-the-art RNA-seq technology. According to the results from the CCK-8 assay, GZZSZTW significantly promoted proliferation in primary chondrocytes in a dose-dependent manner. Furthermore, GZZSZTW almost showed the same effect as DAE at the concentrations of 0.8 mg/ml and 1.0 mg/ml. We then performed RNA-seq to further explore the mechanisms responsible for regulating cell proliferation in response to GZZSZTW treatment.
We first analyzed the differentially expressed genes involved in enhancing cell proliferation. Our results indicated that GZZSZTW significantly increased the expression levels of multiple genes involved in promoting cell proliferation, most of which are highly expressed in tumor cells, such as Tnfaip2, Chi3l1, Tnf, Pfkfb3, Sox8, Jag1, Mafb, Pla2g7, Hnrnpa1, and E2f3. Expression of the Tnfaip2 gene is induced in response to Tnf, which plays a pivotal role in tumor formation and growth. Overexpression of Tnfaip2 significantly promotes tumor cell proliferation, and silencing of Tnfaip2 suppresses proliferation [12]. Chi3l1 is highly expressed in various tumors and possesses oncogenic properties. Chi311 promotes cell proliferation in a similar manner to insulin-like growth factor 1 [13]. Pfkfb3 is overexpressed in many cancers, and it promotes cell proliferation through accelerating cell cycle progression and suppressing apoptosis [14]. Sox8 is highly expressed in many tumor cells, and downregulation of Sox8 suppresses tumor cell proliferation [15]. Jag1, a ligand for the Notch family of receptors, is highly expressed in a variety of tumors. Overexpression of Jag1 enhances tumor cell proliferation [16]. Mafb is a member of the Maf family of transcription factors. Overexpression of Mafb enhances tumor cell proliferation, whereas its knockdown inhibits tumor cell proliferation [17]. Pla2g7 is a cancer-selective biomarker; silencing of Pla2g7 is an antiproliferative and proapoptotic therapeutic approach in cancer treatment [18]. Hnrnpa1 is a member of the heterogeneous nuclear ribonucleoprotein family and is highly expressed in growing mammalian cells. Knockdown of Hnrnpa1 inhibits tumor cell proliferation [19]. E2f3, a member of the E2F family of transcription factors, is critical for the transcriptional activation of genes that control proliferation in both normal and tumor cells [20]. Our results indicated that GZZSZTW treatment significantly promoted chondrocyte proliferation by upregulating the expression levels of functional genes involved in the promotion of cell proliferation.
Consistent with the results discussed above, the expression levels of several functional genes involved in the inhibition of cell proliferation were significantly decreased in response to GZZSZTW treatment, most of which act as tumor suppressors, including Rhob, Dusp6, Plk3, Fgf21, Rad9a, Filip1l, and Rasl11b. Rhob is a member of the Ras homolog family. Rhob serves as a tumor suppressor, and loss of Rhob expression has been reported in a variety of tumor cells [21]. Dusp6 is a negative regulator of tumor cell proliferation, and its expression is significantly decreased in many types of invasive tumor cells. Overexpression of Dusp6 suppresses tumor cell proliferation [22]. Plk3 is a member of the Plk family. Plk3 is downregulated in various types of tumor cells and inhibits tumor cell proliferation and tumorigenesis [23]. Fgf21 is an endocrine factor that inhibits chondrocyte proliferation and thus reduces skeletal growth [24]. Rad9a is a cell cycle checkpoint control protein and is required for cell cycle arrest and DNA damage repair in response to DNA damage [25]. Filip1l is downregulated in many types of tumor cells, and its overexpression results in inhibition of cell proliferation and increased apoptosis [26]. Rasl11b is a member of the Ras-like protein family, and its overexpression leads to decreased cell proliferation [27]. Our results indicated that GZZSZTW treatment significantly promoted chondrocyte proliferation by downregulating the expression levels of functional genes involved in the inhibition of cell proliferation.
According to our RNA-seq analysis, the expression levels of the functional genes involved in chondrocyte proliferation and differentiation were also changed slightly in response to GZZSZTW treatment. For instance, the expression levels of pancartilaginous early chondrocyte markers, including Sox9, Sox5, Sox6, Acan, Col2a1, Col9a1, Col11a1, Hapln1, and Wwp2, were slightly increased in response to GZZSZTW treatment. Sox9 is a master transcription factor that plays a major role during cartilage development through regulation of its target genes, including Sox5, Sox6, Acan, Col2a1, Col9a1, Col11a1, Hapln1, and Wwp2 [28–30]. We further investigated the expression levels of growth plate cartilage markers at different stages of differentiation. The expression levels of markers of proliferating and prehypertrophic chondrocytes, including Fgfr3, Matn1, Comp, Ptch1, Runx2, and Runx3, were slightly increased in response to GZZSZTW treatment. However, the expression levels of the major markers of prehypertrophic and hypertrophic chondrocytes, including Pth1r, Sp7, Ihh, Bmp6, and Ibsp, either decreased slightly or exhibited almost no change in response to GZZSZTW treatment. These results indicated that GZZSZTW treatment facilitated chondrogenesis, maintained chondrocytes in a proliferative state, and prevented their further differentiation into hypertrophic chondrocytes. However, we also observed that the expression level of Col10a1, a specific marker of prehypertrophic and hypertrophic chondrocytes in growth plate cartilage, was slightly increased in response to GZZSZTW treatment. This observation might be explained by the regulatory function of Sox9, which also serves as an upstream regulator of Col10a1 during the progression toward the hypertrophic stage. Sox9 sustains proliferation in columnar chondrocytes and is also required for chondrocyte hypertrophy, a stage that also includes Col10a1 expression [31].
GZZSZTW is designed according to professor Bailing Liu's clinical experiences under the guidance of fundamental theories of traditional Chinese medicine. Among the seven types of CMM in GZZSZTW, Rehmannia glutinosa and Cynomorium coccineum subsp. songaricum (Rupr.) are important CMM with the ability to nourish the “kidney”. In tradition Chinese medicine, kidney is never a definition in anatomic sense but a functional system which plays crucial roles in storing essence and regulating growth, development, and reproduction [32, 33]. Epimedium brevicornu Maxim, a traditional Chinese herbal medicine, has been widely used in China for the treatment of various diseases, such as arthritis and osteoporosis [34]. Icariin, which is considered the major bioactive component of this herbal medicine, has been shown to suppress articular cartilage and bone loss and prevents joint destruction [35]. The traditional Chinese herbal medicine Drynaria fortunei is commonly used to treat musculoskeletal traumatic disorders, such as bone fracture and osteoarthritis [36, 37]. Propinqualin, which is similar in chemical structure to the natural phytoestrogens naringenin and genistein, has been considered as the putative active ingredient of Drynaria fortunei [38]. However, the role of propinqualin in the regulation of chondrocyte is still largely unknown. Cibotium barometz is widely used in Chinese medicinal formulations for the clinical treatment of osteoarthritis. Polysaccharides, one of the most important bioactive components extracted from Cibotium barometz, have been shown to stimulate chondrocyte proliferation in vitro by promoting G1/S cell cycle transition [39].
Spatholobus suberectus Dunn and Raphanus sativus L. are two types of medicinal and edible plants in traditional Chinese medicine. Spatholobus suberectus Dunn has been widely used as dietary supplements in addition to its traditional prescription for treating anemia, arthralgia, inflammation, and arthritis. It has been shown that gallic acid is the major compound with the anti-inflammatory effects in Spatholobus suberectus Dunn [40]. Im and colleagues have shown that Spatholobus suberectus Dunn has a potential therapeutic effect for treating osteoarthritis through preventing extracellular matrix destruction in articular cartilage [41]. Raphanus sativus L. is an edible root vegetable that is commonly used around the world. The seeds of Raphanus sativus L have long been used as anti-inflammatory traditional herbal medicine, and it has been shown that sinapic acid is the major active constituent [42]. Taken together, our results indicated that various herbs and their active components in GZZSZTW synergistically promoted chondrocyte viability and proliferation, but inhibited chondrocyte differentiation, extracellular matrix destruction, and inflammation.
5. Conclusions
The present study demonstrated that the Chinese medicinal formulation GZZSZTW, which has been used for several decades to treat joint diseases (e.g., osteoarthritis), significantly promoted chondrocyte viability and proliferation, maintained chondrocytes in a continuous proliferative state, and inhibited further differentiation. These effects were achieved by the synergistic interactions of various herbs and their active components in GZZSZTW, which increased the expression levels of functional genes participating in chondrocyte commitment and proliferation and decreasing the expression levels of genes involved in chondrocyte differentiation. GZZSZTW treatment also decreased the expression levels of genes that inhibit chondrocyte proliferation. Thus, this study has greatly deepened the current knowledge about the molecular effects of GZZSZTW on chondrocytes. This study has also shed new light on possible strategies to further prevent and treat cartilage-related diseases by using traditional Chinese medicinal formulations.
Acknowledgments
This work was supported by the Science and Technology Development Project of Jilin Province (Grant No. 20170520044JH), the Science and Technology Project of the Jilin Provincial Education Department (Grant No. JJKH20170721KJ), the TCM Clinical Research Center for Bone diseases of Jilin Province (Grant No. 20180623048TC), and the National Natural Science Foundation of China (Grant No. 81702136).
Contributor Information
Xiangyang Leng, Email: leng_xiangyang@163.com.
Daqing Zhao, Email: zhaodaqing1963@163.com.
Data Availability
The data sets were deposited in the NCBI Sequence Read Archive (SRA) database under accession number SRP125978.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this paper.
References
- 1.Mullard A. Chinese biopharma starts feeding the global pipeline. Nature Reviews Drug Discovery. 2017;16(7):443–446. doi: 10.1038/nrd.2017.94. [DOI] [PubMed] [Google Scholar]
- 2.Zhou X., Seto S. W., Chang D., et al. Synergistic effects of Chinese herbal medicine: a comprehensive review of methodology and current research. Frontiers in Pharmacology. 2016;7, article 201 doi: 10.3389/fphar.2016.00201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bay D. Traditional Chinese materia medica: a retrospect and prospect. Memórias do Instituto Oswaldo Cruz. 1991;86(2):1–4. doi: 10.1590/S0074-02761991000600003. [DOI] [PubMed] [Google Scholar]
- 4.Zhang W., Ouyang H., Dass C. R., Xu J. Current research on pharmacologic and regenerative therapies for osteoarthritis. Bone Research. 2016;4, article 15040 doi: 10.1038/boneres.2015.40. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Akkiraju H., Nohe A. Role of chondrocytes in cartilage formation, progression of osteoarthritis and cartilage regeneration. Journal of Developmental Biology. 2015;3(4):177–192. doi: 10.3390/jdb3040177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Gosset M., Berenbaum F., Thirion S., Jacques C. Primary culture and phenotyping of murine chondrocytes. Nature Protocols. 2008;3(8):1253–1260. doi: 10.1038/nprot.2008.95. [DOI] [PubMed] [Google Scholar]
- 7.Kim D., Langmead B., Salzberg S. L. HISAT: a fast spliced aligner with low memory requirements. Nature Methods. 2015;12(4):357–360. doi: 10.1038/nmeth.3317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Trapnell C., Williams B. A., Pertea G., et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature Biotechnology. 2010;28(5):511–515. doi: 10.1038/nbt.1621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wang L., Feng Z., Wang X., Zhang X. DEGseq: an R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics. 2010;26(1):136–138. doi: 10.1093/bioinformatics/btp612. [DOI] [PubMed] [Google Scholar]
- 10.Schmittgen T. D., Livak K. J. Analyzing real-time PCR data by the comparative CT method. Nature Protocols. 2008;3(6):1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 11.Yao B., Zhang M., Leng X., et al. Antler extracts stimulate chondrocyte proliferation and possess potent anti-oxidative, anti-inflammatory, and immune-modulatory properties. In Vitro Cellular & Developmental Biology - Animal. 2018;54(6):439–448. doi: 10.1007/s11626-018-0266-2. [DOI] [PubMed] [Google Scholar]
- 12.Xie Y., Wang B. Downregulation of TNFAIP2 suppresses proliferation and metastasis in esophageal squamous cell carcinoma through activation of the Wnt/β-catenin signaling pathway. Oncology Reports. 2017;37(5):2920–2928. doi: 10.3892/or.2017.5557. [DOI] [PubMed] [Google Scholar]
- 13.Areshkov P. O., Avdieiev S. S., Balynska O. V., LeRoith D., Kavsan V. M. Two closely related human members of chitinase-like family, CHI3L1 and CHI3L2, activate ERK1/2 in 293 and U373 cells but have the different in-fluence on cell proliferation. International Journal of Biological Sciences. 2011;8(1):39–48. doi: 10.7150/ijbs.8.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Yalcin A., Clem B. F., Imbert-Fernandez Y., et al. 6-Phosphofructo-2-kinase (PFKFB3) promotes cell cycle progression and suppresses apoptosis via Cdk1-mediated phosphorylation of p27. Cell Death & Disease. 2014;5(7) doi: 10.1038/cddis.2014.292.e1337 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Xie C., Han Y., Liu Y., Han L., Liu J. miRNA-124 down-regulates SOX8 expression and suppresses cell proliferation in non-small cell lung cancer. International Journal of Clinical and Experimental Pathology. 2014;7:7518–7526. [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 16.Simon D. P., Giordano T. J., Hammer G. D. Upregulated JAG1 enhances cell proliferation in adrenocortical carcinoma. Clinical Cancer Research. 2012;18(9):2452–2464. doi: 10.1158/1078-0432.CCR-11-2371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yu H., Jiang H.-L., Xu D., et al. Transcription factor MafB promotes hepatocellular carcinoma cell proliferation through up-regulation of cyclin D1. Cellular Physiology and Biochemistry. 2016;39(2):700–708. doi: 10.1159/000445661. [DOI] [PubMed] [Google Scholar]
- 18.Vainio P., Lehtinen L., Mirtti T., et al. Phospholipase PLA2G7, associated with aggressive prostate cancer, promotes prostate cancer cell migration and invasion and is inhibited by statins. Oncotarget . 2011;2(12):1176–1190. doi: 10.18632/oncotarget.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Liu X., Zhou Y., Lou Y., Zhong H. Knockdown of HNRNPA1 inhibits lung adenocarcinoma cell proliferation through cell cycle arrest at G0/G1 phase. Gene. 2016;576(2):791–797. doi: 10.1016/j.gene.2015.11.009. [DOI] [PubMed] [Google Scholar]
- 20.Humbert P. O., Verona R., Trimarchi J. M., Rogers C., Dandapani S., Lees J. A. E2f3 is critical for normal cellular proliferation. Genes & Development. 2000;14(6):690–703. [PMC free article] [PubMed] [Google Scholar]
- 21.Chen W., Niu S., Ma X., et al. RhoB acts as a tumor suppressor that inhibits malignancy of clear cell renal cell carcinoma. PLoS ONE. 2016;11(7) doi: 10.1371/journal.pone.0157599.e0157599 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Piya S., Kim J. Y., Bae J., Seol D.-W., Moon A. R., Kim T.-H. DUSP6 is a novel transcriptional target of p53 and regulates p53-mediated apoptosis by modulating expression levels of Bcl-2 family proteins. FEBS Letters. 2012;586(23):4233–4240. doi: 10.1016/j.febslet.2012.10.031. [DOI] [PubMed] [Google Scholar]
- 23.Lv H., Gao G., Zhang L., Sun Y. Polo-like kinase 3 inhibits osteosarcoma cell proliferation and tumorigenesis via cooperative interaction with p21. Molecular Medicine Reports. 2015;12(5):6789–6796. doi: 10.3892/mmr.2015.4304. [DOI] [PubMed] [Google Scholar]
- 24.Wu S., Levenson A., Kharitonenkov A., De Luca F. Fibroblast growth factor 21 (FGF21) inhibits chondrocyte function and growth hormone action directly at the growth plate. The Journal of Biological Chemistry. 2012;287(31):26060–26067. doi: 10.1074/jbc.M112.343707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Weinert T. A., Hartwell L. H. Characterization of RAD9 of Saccharomyces cerevisiae and evidence that its function acts posttranslationally in cell cycle arrest after DNA damage. Molecular and Cellular Biology. 1990;10(12):6554–6564. doi: 10.1128/MCB.10.12.6554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Burton E. R., Gaffar A., Lee S. J., et al. Downregulation of filamin A interacting protein 1-like is associated with promoter methylation and induces an invasive phenotype in ovarian cancer. Molecular Cancer Research. 2011;9(8):1126–1138. doi: 10.1158/1541-7786.MCR-11-0162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Emerson S. E., St. Clair R. M., Waldron A. L., et al. Identification of target genes downstream of semaphorin6A/PlexinA2 signaling in zebrafish. Developmental Dynamics. 2017;246(7):539–549. doi: 10.1002/dvdy.24512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Nakamura Y., Yamamoto K., He X., et al. Wwp2 is essential for palatogenesis mediated by the interaction between Sox9 and mediator subunit 25. Nature Communications. 2011;2(1, article 251) doi: 10.1038/ncomms1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Leung V. Y. L., Gao B., Leung K. K. H., et al. SOX9 governs differentiation stage-specific gene expression in growth plate chondrocytes via direct concomitant transactivation and repression. PLoS Genetics. 2011;7(11) doi: 10.1371/journal.pgen.1002356.e1002356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Urano T., Narusawa K., Shiraki M., et al. Single-nucleotide polymorphism in the hyaluronan and proteoglycan link protein 1 (HAPLN1) gene is associated with spinal osteophyte formation and disc degeneration in Japanese women. European Spine Journal. 2011;20(4):572–577. doi: 10.1007/s00586-010-1598-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dy P., Wang W., Bhattaram P., et al. Sox9 directs hypertrophic maturation and blocks osteoblast differentiation of growth plate chondrocytes. Developmental Cell. 2012;22(3):597–609. doi: 10.1016/j.devcel.2011.12.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zhang R. X., Li M. X., Jia Z. P. Rehmannia glutinosa: review of botany, chemistry and pharmacology. Journal of Ethnopharmacology. 2008;117(2):199–214. doi: 10.1016/j.jep.2008.02.018. [DOI] [PubMed] [Google Scholar]
- 33.Liu H.-P., Chang R.-F., Wu Yi.-S., Lin W.-Y., Tsai F.-J. The Yang-Tonifying Herbal Medicine Cynomorium songaricum extends lifespan and delays aging in Drosophila. Evidence-Based Complementary and Alternative Medicine. 2012;2012:11. doi: 10.1155/2012/735481.735481 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ma H., He X., Yang Y., Li M., Hao D., Jia Z. The genus Epimedium: an ethnopharmacological and phytochemical review. Journal of Ethnopharmacology. 2011;134(3):519–541. doi: 10.1016/j.jep.2011.01.001. [DOI] [PubMed] [Google Scholar]
- 35.Wei C. C., Ping D. Q., You F. T., Qiang C. Y., Tao C. Icariin prevents cartilage and bone degradation in experimental models of arthritis. Mediators of Inflammation. 2016;2016 doi: 10.1155/2016/9529630.9529630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Wong R. W. K., Rabie A. B. M. Effect of gusuibu graft on bone formation. Journal of Oral and Maxillofacial Surgery. 2006;64(5):770–777. doi: 10.1016/j.joms.2006.01.008. [DOI] [PubMed] [Google Scholar]
- 37.Chen F.-P., Chang C.-M., Hwang S.-J., Chen Y.-C., Chen F.-J. Chinese herbal prescriptions for osteoarthritis in Taiwan: analysis of national health insurance dataset. BMC Complementary and Alternative Medicine. 2014;14, article 91 doi: 10.1186/1472-6882-14-91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sun J.-S., Thériault B. L., Anderson G. I. The effect of Gu-Sui-Bu (Drynaria fortunei) on bone cell activity. American Journal of Chinese Medicine. 2004;32(5):737–753. doi: 10.1142/S0192415X0400234X. [DOI] [PubMed] [Google Scholar]
- 39.Fu C., Zheng C., Lin J., et al. Cibotium barometz polysaccharides stimulate chondrocyte proliferation in vitro by promoting G1/S cell cycle transition. Molecular Medicine Reports. 2017;15(5):3027–3034. doi: 10.3892/mmr.2017.6412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Mohibbullah M., Ye J. L., Park H. J., et al. The medicinal herb Spatholobus suberectus with promising in vitro antioxidant and anti-inflammatory potentials and its phytochemical characterization by RP-HPLC analysis. J Food Biochem. 2017;2017e12480 [Google Scholar]
- 41.Im N.-K., Lee S.-G., Lee D.-S., Park P.-H., Lee I.-S., Jeong G.-S. Spatholobus suberectus inhibits osteoclastogenesis and stimulates chondrogenesis. American Journal of Chinese Medicine. 2014;42(5):1123–1138. doi: 10.1142/S0192415X14500700. [DOI] [PubMed] [Google Scholar]
- 42.Kook S.-H., Choi K.-C., Lee Y.-H., Cho H.-K., Lee J.-C. Raphanus sativus L. seeds prevent LPS-stimulated inflammatory response through negative regulation of the p38 MAPK-NF-κB pathway. International Immunopharmacology. 2014;23(2):726–734. doi: 10.1016/j.intimp.2014.11.001. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data sets were deposited in the NCBI Sequence Read Archive (SRA) database under accession number SRP125978.