

Abstract
Analyses of steroid receptors are important for understanding molecular details of transcriptional control, as well as providing insight as to how an individual transacting factor contributes to cell identity and function. These studies have led to the identification of a superfamily of regulatory proteins that include receptors for thyroid hormone and the vertebrate morphogen retinoic acid. Although animals employ complex and often distinct ways to control their physiology and development, the discovery of receptor-related molecules in a wide range of species suggests that mechanisms underlying morphogenesis and homeostasis may be more ubiquitous than previously expected.
STEROID AND THYROID HORMONES ACT TO COORDINATE complex events involved in development, differentiation, and physiological response to diverse stimuli. These molecules, through binding to specific intracellular receptors, coordinate the components of behavioral and physiological repertoires by activat-ing the expression of gene networks. Thus, the hormone-receptor complex may function as a key constituent in determining commit-ment to specific cell lineages, as well as provokin differentiation, in already determined cells. The purposes of this review are (i) to establish the historical perspective that associated these molecules with the control of differential patterns of gene expression; (ii) to describe the striking evolution of our understanding of the structure/function relationships between receptors and the implications for regulation of gene activity; and (iii) to present emerging issues on the physiology and the molecular basis of hormone action.
Past
Diseases that we now understand to be associated with defects in steroid and thyroid hormone function were identified relatively early in medical history; it was only since the early part of this century that a foundation for physiological studies was supplied by the isolation and structural analyses of these hormones. It was known from the work of Huxley and others that extracts from thyroids could control the metamorphosis of amphibians, but it was not until 1915 that Kendall was able to crystallize the molecule involved and show that it was composed of two iodinated tyrosine residues (1, 2). Ten years later, both Kendall and Reichstein completed the structural analysis of cortisol purified from the adrenal cortex, which led to the realization that it was (as are all other steroid hormones) derived from cholesterol (3, 4). While many considered this to be an achievement of modern endocrinology, one is humbled by the fact that Chinese alchemists (5), for medicinal reasons, developed empirical methods between the 10th and 16th centuries to purify steroids to near homogeneity.
From the early 1900s to the present, there has been a tremendous increase in our knowledge of endocrine organs and the diverse physiology that they coordinate (3, 6). Three major classes of steroid hormones have been described on the basis of biological assays: the adrenal steroids (including cortisol and aldosterone), the sex steroids (progesterone, estrogen, and testosterone), and vitamin D3. These molecules were shown to be profoundly important for correct vertebrate development and physiology and, consequently, each has become a major focus of biological and clinical investigation (7). The adrenal steroids widely influence body homeostasis, controlling glycogen and mineral metabolism as well as mediating the stress response. They have widespread effects on the immune and nervous systems and influence the growth and differentiation of cultured cells. The sex steroids provoke the development and determination of the embryonic reproductive system, masculinize or feminize the brain at birth, control reproduction and reproductive behavior in the adult, and control development of secondary sexual characteristics. Vitamin D is necessary for normal bone development and plays a critical role in calcium metabolism and bone differentiation. Aberrant production of these hormones has been associated with a broad spectrum of clinical disease including cancer.
Both thyroid and steroid hormones can be important in metamorphosis. A thyroidectomized tadpole will not develop to a frog, but addition of thyroxin to the water induces all of the changes for development to a terrestrial adult (1). Similarly, ecdysteroids act as metamorphic hormones in insects (8). It was possible to associate the action of ecdysone directly to induced changes in chromosome structure (8) during ecdysone-induced chromosome puffing, suggesting a link between steroid hormones and activation of gene expression.
How can small, relatively simple molecules elicit such a diversity of complex responses? The first clue was provided by the identification of steroid and thyroid hormone receptors through the use of radioactively labeled ligands in the early 1970s (9). In each case the hormone induced a change in the receptor such that it associated with high-affinity binding sites in chromatin. This, in turn, led to the induction or repression of a limited number of genes (approximately 50 to 100 per cell) (10). Selectivity is achieved, in part, by restricted expression of the different receptors in specific cells and tissues. Because the chromatin structure of each cell type is uniquely organized, different sets of genes may be accessible to the hormone receptor complex.
Attempts were initiated to purify the steroid and thyroid hormone receptors despite the sobering realization that these molecules were present in only trace amounts (103 to 104 per cell) and would thus require enrichments of 105- to 106-fold to achieve homo geneity. The development of high-affinity synthetic analogs of the ligands overcame many of the difficulties of receptor isolatioln and has revolutionized both clinical and biochemical studies (6, 7). By the early 1980s all but the androgen, mineralocorticoid, and thyroid hormone receptors were purified (11, 12). Each receptor undergoes a structural alteration or “transformation” upon hormone binding, which in turn enables DNA binding. Analysis of the purified glucocorticoid receptor revealed that DNA binding and honmone binding properties, although present in a single molecule, coufld be separated by limited proteolysis, leading to the first suggestioln of a domain structure (13, 14).
Purification and biochemical characterization of the glucocorticoid receptor was accompanied by the identification of a variety of glucocorticoid-responsive genes (11, 12, 15). Many of these genes have been isolated and shown to be transcriptionally regulated by glucocorticoids. Gene transfer studies, particularly with the mouse mammary tumor virus (MMTV) promoter and the human metallothionein IIA promoter, identified short cis-acting sequences (about 20 bp in size) that are required for hormonal activation of transcription (16, 17). The attachment of these elements to an otherwise hormone-nonresponsive gene causes that gene to be hormone-responsive (18). These sequences, or hormone response elements (HREs), function in a position- and orientation-independent fashion and thus behave like transcriptional enhancers (19 ,20). Unlike other enhancers, their activity is dependent upon the presence or absence of ligand. These studies suggest that transcriptional regulation derives from the bindibg of hormone-receptor complexes to HRE sites on DNA. This interpretatio has been verified by in vitro footprint analyses which reveal that purified glucocorticoid, estrogen, progesterone, and thyroid hormone receptors bind to the upstream DNA of responsive genes at sites which correspond to the genetically identified HREs (16, 19–25). The apparent dyad symmetry of these elements (Fig. 1) suggests that they interact with receptor dimers.
Fig. 1.

Alignment of nucleic acid sequences in regions correspond to identified HREs (15–25). Numbering refers to nucleotide position relative to the start of transcription. Arrows indicate dyad axis of symme consensus sequence is derived from nucleotides conserved with a frequency of 50% or more. Specific references for HREs can be found in Fig. 2. GRE glucocorticoid response element; MMTV, mouse mammary tumor virus; hGH, human growth hormone; MSV, murine sarcoma virus; hMTIIA, human metallothionein; TO, tyrosine oxidase; TAT, tyrosine aminotransferase; ERE, estrogen response element; xVit, Xenopus vitellogenin; cVit, chicken vitellogenin; Oval, chicken ovalbumin; rPrl, rat prolactin;TRE thyroid hormone response element; and rGH, rat growth hormone.
Present
Comparative anatomy.
Analysis of the hormone receptors is essential for understanding both the origins of complex regulatory systems and how they contribute to the maintenance of the organism. The isolation of steroid receptor complementary DNAs (cDNAs) has identified a family of related genes that bind ligands of remarkable diversity. The interaction between steroid receptor genes, the genetic circuits that they control, and their contributions to spatial organization in the embryo and organ physiology in the adult can now be elucidated.
The expression cloning of the human glucocorticoid receptor (hGR) provided the first completed structure of a steroid receptor and revealed a segment with astonishing relatedness to the viral oncogene erbA (26–28). This relationship of the hormone receptors to erbA was independently confirmed by the cloning of the human estrogen, progesterone, aldosterone, and vitamin D receptors (29–36). Two groups initiated the characterization of the erbA protooncogene product that led to its startling identification as the thyroid hormone receptor (37, 38). This represented a critical advance, for it suggested a unifying hypothesis for receptor structure and hormone action.
Although steroid and thyroid hormones are neither structurally nor biosynthetically related, the existence of a common structure for their receptors supports the proposal that there is a large superfamily of genes whose products are ligand-responsive transcription factors. The presence of a highly conserved DNA sequence element initiated searches for such cryptic receptor genes. By means of low stringency hybridization techniques at least five new gene products have been identified. Two of these, referred to as estrogen receptor–related genes 1 and 2 (ERR1 and ERR2), are more related to the steroid than to the thyroid hormone receptors but do not bind any of the major classes of known steroid hormones (39). The remaining are closer to the thyroid hormone receptor. Indeed, one of them is a second thyroid hormone receptor (40, 41). Another is the apparent receptor for the vitamin A–related metabolite retinoic acid (42, 43). The third is closely related to the receptor for retinoic acid; although its ligand is not known (44, 45) the receptor has been implicated in the etiology of hepatocellular carcinoma, and has been named HAP.
The recent characterization of the E75 locus from Drosophila predicts the existence of a protein with overall structural properties similar to the steroid and thyroid hormone receptors (46). Structur al comparisons of the E75 gene product with the vertebrate homologs indicate remarkable relatedness to the thyroid hormone and vitamin D receptors. Perhaps this is a receptor for the insect steroid ecdysone or the isoprenoid juvenile hormone.
Schematic results of molecular cloning studies are presented in Fig. 2 in which the molecules have been aligned on the basis of regions of maximum protein homology (47). The numbers indicate the extent of sequence identity to the hGR. The central core sequence is rich in Cys, Lys, and Arg residues and is highly conserved (homologies ranging from 42 to 94%). The homology in the ligand-binding domain is more graded and generally parallels the structural relatedness of the hormones themselves. Although the NH2-terminus is not conserved, it may contribute to important functional differences between receptors.
Fig. 2.
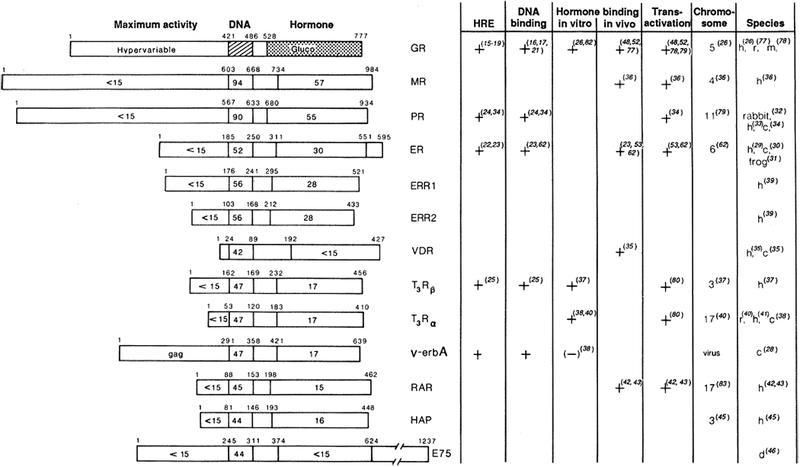
Schematic amino acid comparison of members of the steroid hormone receptor superfamily. Primary amino acid sequences have been aligned on the basis of regions of maximum amino acid similarity, with the percentage amino acid identity indicated for each region in relation to the hGR (55). Domains shown are a domain at the NH2-terminal end, required for “Maximum activity”; the 66- to 68-amino acid DNA-binding core (“DNA”); and the 25-amino acid hormone-binding domain (“Hormone”). The amino acid position of each domain boundary is shown. Amino acid numbers for all receptors represent the human forms with the exception of v-erbA and E75 (46). Functional assignments have been determined by characterization of the glucocorticoid and estrogen receptors. Designations are as follows: GR, glucocorticoid receptor; MR, mineralocorticoid receptor; PR, progesterone receptor; ER, estrogen receptor; ERR1 or ERR2, estrogenr eceptor–related1 or 2; VDR, vitaminD3 receptor;and T3Rβ and T3Rα, thyroid hormone receptors. The (+) or (−) indicates whether a particular property has been demonstrated for the products of cloned receptor cDNA or with purified receptor. HRE, hormone response element. This relates to whether the binding site has been identified structurally and whether its enhancement properties have been demonstrated by gene transfer studies. For PR, DNA-binding properties have been shown only with the native purified receptor. “Hormone binding in vitro” indicates whether this property has been demonstrated by translation in a rabbit reticulocyte lysate system (26). “Hormone binding in vivo” refers to expression of the cloned receptor in transfected cells. “Chromosome” indicates the human chromosome location. Species are as follows: h, human; r, rat; m, mouse; c, chicken; and d, Drosophila.
Functional domains.
The classic model for steroid/thyroid hor mone action proposes that binding of the ligand to the receptor induces an allosteric change that allows the receptor-hormone complex to bind to its DNA response element in the promoter region of a target gene. It is this binding that leads to modulation of gene expression. The cloning of receptor cDNAs provides the first opportunity to dissect the molecular basis of steroid action.
The identification of functional domains for hormone binding, DNA binding, and transactivation was facilitated by a screening assay that uses cultured cells transfected with two DNA expression vectors (Fig. 3). The trans-vector provides for the efficient production of the receptor in cells that do not normally express the receptor gene. The cis-vector contains a luciferase gene (or any other easily monitored function) coupled to a hormone-responsive promoter. Applications of hormone or an experimental agonist will activate the luciferase gene, causing light to be emitted from cell extracts. The level of light emitted is directly proportional to the effectiveness of the hormone receptor complex in activating gene expression.
Fig. 3.
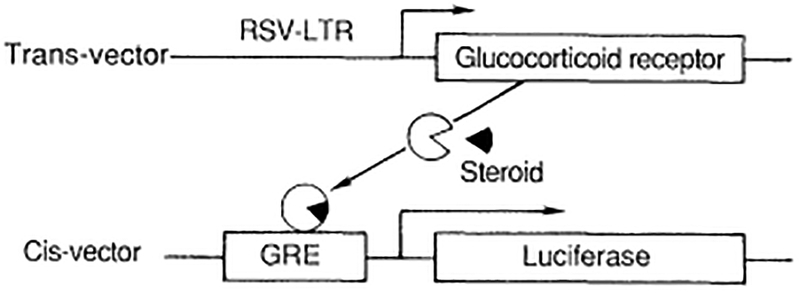
The cotransfection assay. Cultured cells are transfected with the receptor cDNA in an expression vector (the trans-vector). The function of this transcription factor can be monitored by the activity of a reporter gene (the luciferase gene) linked to an appropriate hormone-responsive promoter. In this case, the promoter is from the MMTV virus carrying a GRE enhancer. The trans-vector encodes the hGR, shown combining with the steroid hormone (triangle).
In the case of the glucocorticoid receptor, the cis-vector contains the mammary tumor virus (MTV) promoter, which has a well-characterized glucocorticoid response element (GRE). In the cotransfection assay, expression of the cis-vector is induced about 500-fold in a hormone-dependent fashion. By means of this assay it is possible to investigate the effects of in vitro mutations on receptor activity.
Sequence comparisons in combination with related functional studies have given rise to an emerging picture for a common structure for the receptor superfamily gene products (Fig. 2). An unexpected and revealing result from the mutational studies is that loss of a portion of the hormone-binding region of the glucocorticoid receptor engenders a constitutively active molecule (48, 49). The results provide the first mechanistic insight into the process of activation: neither the steroid-binding domain nor the steroid hormone itself is needed for DNA binding or transcriptional enhancement. Instead, it seems that the hormone-binding region normally prevents the domains for DNA binding and transcriptional activation from functioning. The addition of hormone apparently relieves this inhibition (50, 51).
Our initial suspicion was that the DNA-binding domain is included within the highly conserved central core of the protein. Three features supported this suggestion: (i) the clustering of basic residues likely to interact with DNA, (ii) the presence of a Cys-rich motif, and (iii) the high homology of this core among receptors (27). To test this assignment, different parts of this region were mutated (48, 49, 52, 53). Mutants continue to bind hormone, indicating that the structure of the protein is intact; however, they do not bind DNA. A direct proof of function was provided by substituting the putative DNA-binding domain of the human estrogen receptor (hER) with that of the hGR, resulting in a hybrid receptor with the predicted switch in template specificity (54). This suggested a general strategy, referred to as the finger swap, which has been successfully exploited to characterize novel hormone receptors (Fig. 4).
Fig. 4.
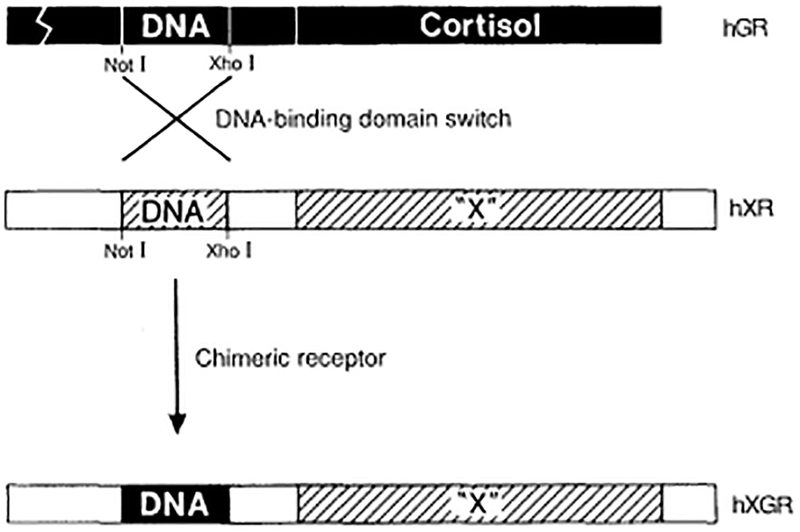
The finger swap.The modular structure of the steroid receptors allows the exchange of one domain for another to create a functional hybrid. Thus, if the DNA-binding domain of a candidate receptor is substituted with the corresponding region from the glucocorticoid receptor, the resulting chimeric receptor should stimulate the MTV promoter when exposed to the appropriate ligand. This approach was used to functionally identify the retinoic acid receptor (42, 43) and alter the binding specificity of the estrogen receptor (54).
These issues raise the question of whether there are structural aspects of the DNA-binding region that can explain its properties. The most striking feature is the conservation of Cys residues. A comparison of the amino acid sequences in the DNA-binding domain of the hormone receptors (Fig. 5) reveals significant identity and similarity over these evolutionary divergent molecules. Out of 65 residues, 20 are invariant, an additional 7 are conserved in 7/8 of the gene products, and more than half are conserved in 5/8 of the molecules. Nine of the invariant residues are Cys, with one invariant His (Fig. 5). The positioning of the residues is similar to a motif originally observed in the 5S gene transcription factor TFIIIA (55) in which multiple Cys- and His-rich repeating units apparently fold into a “fingered” structure coordinated by a zinc ion (Fig. 5). This finger of amino acids is proposed to interact with a half turn of DNA.
Fig. 5.
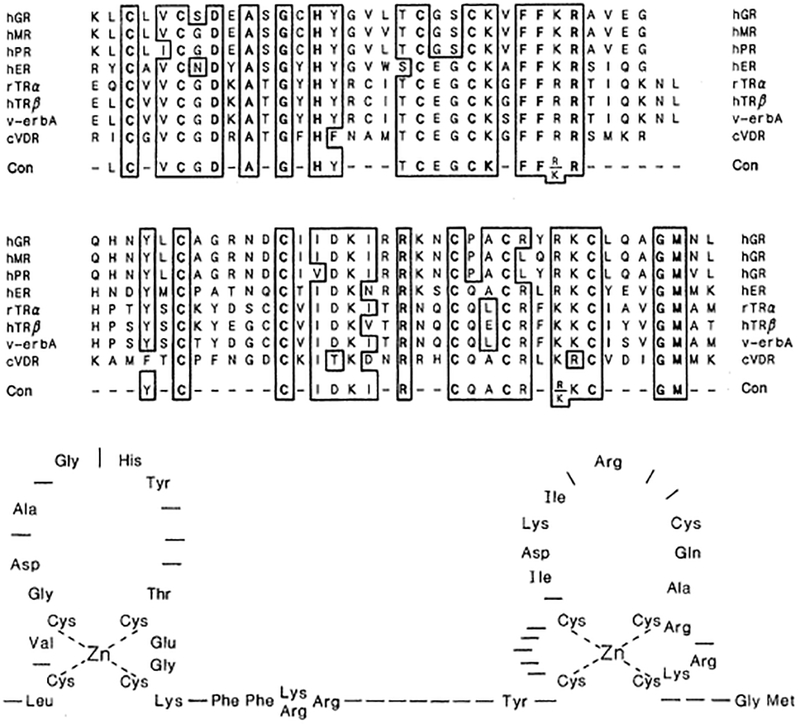
(Top) Amino acid sequence comparison of DNA-binding domains. A computer program for the concurrent comparison of three or more amino acid sequences was used (47). Amino acid residues matched in at least five of the eight polypeptides are boxed and designated in the consensus (Con) sequence. Hyphens indicate divergent sequences; gaps indicate no comparable amino acids. Absolutely conserved residues are in bold print. (Bottom) Hypothetical structure of the DNA-binding domain of the hormone receptors. This domain is configured into two putative zinc-binding fingers with each zinc ion forming a tetrahedral coordination complex with Cys residues. Alternative coordination positions might include the Cys in the second finger and its proximal Cys, shortening the finger and shifting the last conserved Cys into the “trailer” region.
Are such structures important for receptor function? Several results imply they are [see (56) for review]. Site-directed mutagenesis has shown that conserved Cys residues are required for DNA binding (54, 57). Furthermore, recent evidence suggests that the binding of zinc by the receptors is required for DNA binding in vitro (58). Genomic analysis indicates that fingers are encoded by separate exons (36, 59) and an examination of the proposed structure suggests that these fingers are structurally distinct (56). This is most readily seen by the spacing between the cysteines that would be involved in the putative zinc coordination complex. In addition, comparative studies show that the more NH2-terminal “first finger” is more highly conserved among receptors than the more COOH-terminal “second finger.” The first finger is relatively more hydrophilic and has few basic amino acids that might be expected to interact with DNA. The second finger is rich in Lys and Arg residues and is highly basic. Although attention has been focused on zinc fingers, the residues between the two fingers and the residues immediately after the second finger are also highly conserved. This raises the possibility that these stretches mediate part of the DNA-binding function.
In contrast to the highly conserved DNA-binding domain, the NH2-terminal extension of the receptors is hypervariable in size and amino acid composition (Fig. 2). Nevertheless, evidence suggests it contributes to function. Deletions in this region of the glucocorticoid receptor reduce activity by 10- to 20-fold (48, 60). Genetic evidence for a functional role for the NH2-terminus also comes from analysis of the NTi (nuclear transfer increased) glucocorticoid receptor mutants (61). NTi glucocorticoid receptors appear to contain an altered NH2-terminus and, although they can bind hormone, they are not biologically functional. Similarly, an estrogen receptor with an NH2-terminal deletion is able to regulate the vitellogenin promoter in a normal fashion, but is tenfold less active in regulating the expression of the estrogen-responsive promoter p52 (62). Finally, preliminary evidence with the progesterone receptor indicates that the A and B forms, which differ by 128 amino acids at the NH2-terminus, may have strikingly different capacities to regulate gene expression (63). Such results further support the hypothesis that this domain may modulate receptor function by influencing transactivation, DNA binding, or both.
Subfamilies and superfamilies.
The startling discovery of a common structure for the steroid and thyroid hormone receptors and our ability to isolate new receptors by homology suggest that other proteins that contain similar structural features are likely to be hormone- or ligand-responsive transcription factors (LTFs). Apparently it is the analogous action of the hormones that is reflected in the homologous structure of their receptors. An extension of this proposal predicts that other small, hydrophobic molecules may interact with structurally related intracellular receptors. For example, production of cholesterol is regulated by feedback mechanisms that maintain overall levels by monitoring dietary intake and controlling synthesis accordingly (64). Recent evidence demonstrates that at least some of this regulation is transcriptional (65). Since cholesterol is structurally related to steroid hormones, and indeed serves as their biosynthetic precursor, it seems logical to predict both the existence of a cholesterol receptor and its membership in this superfamily. The herbicide TCDD (dioxin) shows close structural relatedness to thyroid hormones and mediates a variety of metabolic effects as a consequence of its action on gene expression. A dioxin receptor has been identified (66), and it now seems likely that this receptor too, will ultimately be part of the LTF superfamily. One of the major issues to arise out of the characterization of this receptor is whether dioxin acts as an agonist or an antagonist for a natural endogenous ligand. Further investigation of this could reveal the existence of a new hormone that may have valuable physiologic and clinical implications.
Preliminary evidence suggests the existence of additional members of the LTF family. For example, the integration of the hepatitis virus in a human liver carcinoma identified a genetic locus (HAP) with striking homology to the DNA-binding fingers of the steroid and thyroid hormone receptors (45). Aberrant expression of HAP could possibly be involved in tumor formation. Indeed, if this locus encodes a new hormone receptor, what might its ligand be? Strong homology to the retinoic acid receptor hints that the product of the HAP locus may bind a related molecule, possibly retinoic acid itself. By extension, the identification of genes for new receptors, by means of low-stringency hybridization techniques, promises to be an exciting area. Already, two novel gene products related to the estrogen receptor have been identified (Fig. 2) (39). Is their expression tissue-specific? Do they bind estrogen? Might they bind other sex steroids and help to identify new hormone response systems? Such discoveries are likely to have an impact on health and human disease as well as expand our knowledge of basic human physiology.
Remarkably, single ligands may have multiple receptors. Currently, two thyroid hormone receptors have been identified and there may be more (37, 40, 41). What could the advantages be to having different receptors for the same hormone? One possibility is that they are expressed in a tissue-specific fashion. This notion has already been confirmed by the identification of an abundant neuronal form of the thyroid hormone receptor (40). Second, it is possible that they respond differently to thyroid hormone metabolites. Third, since their DNA-binding domains differ slightly, they might activate overlapping, yet partially distinct, genetic networks. Finally, multiple thyroid receptor genes provide multiple promoter enhancers that might be responsive to distinct metabolic or hormonal regulators.
The protein product of v-erbA is a derivative of the thyroid hormone receptor that has been proposed to promote leukemogenesis by acting as a thyroxin-independent transcription factor (37, 38). By unknown means, changes in the ligand-binding domain of the protein apparently activate the receptor, perhaps by forcing it into a configuration similar to that achieved by the binding of its physiological ligand. The activation of erbA may therefore be an example of how the loss of allosteric control can confer pathogenicity on the product of a proto-oncogene. In vitro studies already indicate that altered glucocorticoid receptors can be biologically active. It thus seems likely that truncations or mutations in other hormone receptors could lead to activated states perturbing homeostatic balance and abetting tumor progression. Although lacking decisive evidence, we can suggest that mutations in the estrogen and androgen receptors may contribute to the conversion of steroid-dependent breast tumors and prostate tumors to hormone-independent growth (67). As previously mentioned, the integration site of the hepatitis virus in a human tumor may lead to the identification of a new receptor in which a genetic lesion is associated with malignant transformation. A critical step will be the demonstration that mutant receptors contribute to tumorigenesis. It will then be necessary to determine how they exert their effects, whether it simply involves the constitutive activation of hormone-responsive genes or whether it includes an altered substrate specificity so that new genes come under the regulation of mutated receptors. Once a genetic lesion has been identified, this information can be used to contribute to diagnosis and treatment.
Although for many decades it has been understood that sex steroids can influence behavior, the role of other hormones in neurologic function is controversial. Since the 19th-century discovery by Addison of adrenal insufficiency (68), glucocorticoids have been associated with patients’ inability to concentrate, drowsiness, restlessness, insomnia, irritability, apprehension, disturbed sleep, and possibly psychotic episodes and manic-depressive disorders. The effects of thyroid hormones on neuronal development and the high level of expression of the thyroid hormone receptor in the adult nervous system lead to the prediction that aberrant hormonal production, variation in receptor expression, or receptor mutations influencing hormone-binding properties could mediate aberrant metabolic effects in the central nervous system (69). Thus, an important future area of investigation is the contribution of receptors to the etiology of psychiatric disorders.
Ontogeny and physiology.
Although it is widely believed that differential regulation of gene expression is the critical level at which early development is controlled, this does not provide a conceptual framework for the process by which spatial organization is achieved. Despite excellent evidence for graded positional information in Drosophila and nematodes, it is unclear how this relates to morphogenic signals in vertebrates. One long-standing theory is that pattern formation is achieved by the establishment of a gradient of a diffusable substance or morphogen. Work by numerous laboratories over the last several years has indicated that retinoic acid manifests morphogenic properties. Recently, Thaller and Eichele (70) demonstrated that retinoic acid is indeed the substance responsible for establishment of the anterior-posterior axis in the developing chick limb bud. The ability of retinoic acid to induce differentiation in teratocarcinoma cells (71) to parietal endoderm suggests a role for it in the earliest stages of embryonic development.
The discovery of the retinoic acid receptor (42, 43) was made possible from the demonstration that conserved regions in the receptors correspond to discrete functional domains. Thus, by exchanging the DNA-binding domain of the retinoic receptor for the comparable region from the glucocorticoid receptor, a hybrid molecule was generated that activates GRE-responsive promoters (such as the MMTV-LTR) in response to retinoic acid (Fig. 4) (42).
By analogy with steroid receptors, we can propose that the interaction of retinoic acid with its intracellular receptor triggers a cascade of regulatory events that results from the activation of specific sets of genes. Thus, for the first time in a vertebrate system, it should be possible to investigate the mechanism of morphogenesis by identifying a discrete complement of developmental control genes.
With regard to establishment of spatial information, one obvious question is whether there is a gradient of receptor itself. Furthermore, preliminary results reveal the presence of related genes. Might there be receptors for other morphogens and do they also contribute to development?
Mechanisms.
What are the molecular interactions between the ligand and the receptor that lead to activation? Once activated, how does this molecule find a particular binding site and what is the detailed nature of the DNA-protein interaction? What is the molecular interaction between the receptor and the transcriptional machinery? How do receptors and their potential interaction with other transactivators cause RNA polymerase II to initiate transcription? It must be emphasized that steroid and thyroid hormones can repress gene expression as well as activate it. It is important to determine whether repression and activation are mediated by the same types of DNA sequences and whether other protein factors are involved.
Once a receptor is bound to DNA, how does it activate transcription? Molecular interactions with cognate binding sequences have been analyzed for the transcriptional regulatory proteins lac, λ, and cro (72). Because the DNA-binding domain of the hormone receptors is fundamentally different from that of these molecules (which employ a helix-turn helix motif), it will be necessary to co-crystallize the receptors with cognate DNA-binding sites. These studies, along with site-directed mutagenesis of the receptor, should provide information on how the protein recognizes DNA, but may not reveal the dynamics of transactivation. It will be necessary to determine whether transactivation and DNA binding can be separated as they have been in other regulatory proteins such as GAL4 and GCN4 (73, 74). Assuming these functions are separable, it should be possible to identify receptor variants that bind normally to DNA but fail to transactivate (48, 52, 62). On the basis of this knowledge it will then be necessary to develop techniques to characterize the activation process itself. Does the receptor associate with other transcriptional regulatory proteins? Does this occur before the receptor binds to its HRE? Must the receptor remain bound to the DNA template for the associated gene to remain active or can a transiently bound receptor initiate permanent structural change?
The identification of a transactivation function (τl) in the NH2-terminus of the glucocorticoid receptor leads to an unexpected conclusion (52). Since the NH2-terminus is not conserved among different receptors, they each may achieve this function by distinct means. It has been suggested that the activation domain of yeast GAL4 includes a stretch of acidic amino acids configured in an amphipathic a helix (75). Apparently, overall structural features are critical, rather than the specific sequence. Likewise, the τl region is acidic and so is the activation domain of another yeast regulator, GCN4 (74). It remains to be seen if acidic domains embody the activation function of all the steroid receptors. If so, it might suggest that diverse groups of regulatory proteins from yeast to man employ a remarkably conserved approach to transcriptional control. If the receptors interact with other proteins through acid domains, it will be necessary to purify and characterize these molecules. Ultimately the role of individual components and the mechanism of transactivation must be confirmed by the development of receptor-dependent in vitro transcription systems.
Conclusion
In the 1920s, T. H. Morgan, who explored a genetic approach to development, asserted that to understand development one must understand the molecular basis of differential gene expression (76). Although animals develop in very diverse ways, the discovery of receptor-related molecules in a wide range of species suggests that molecular mechanisms underlying developmental and physiological homeostasis may be much more universal than was previously suspected. The cloning of the steroid and thyroid hormone receptors marks an important step forward in understanding fundamental mechanisms of gene regulation as well as hormone action. The paradoxical and reciprocal effects of gene regulation on the cell and that of the cell on the gene embody functional physiology in a profound sense. For this paradox reflects both the irreversible changes of embryonic development as well as the recurrent changes in organ physiology in the adult.
REFERENCES AND NOTES
- 1.Gudernatsch JG, Arch. Entwecklungsmech. Org 35, 457 (1912); Huxley JS, Nature (London) 123, 712, (1929); Elkin W, Mem. Soc. Endocrinol 18, 137 (1970). [Google Scholar]
- 2.Kendall EC,J.Am.Med.Assoc 64, 2042 (1915). [Google Scholar]
- 3.Gaunt R, in Handbook of Physiology: Endocrinology, Greep R and Astwood E, Eds. (American Physiological Society, Washington, DC, 1975), vol. 6, pp. 1–12. [Google Scholar]
- 4.Bentley PJ, Comparative Vertebrate Endocrinology, (Cambridge Univ. Press, Cambridge, 1986), pp. 65–76. [Google Scholar]
- 5.Needham J, Science and Civilisation in China (Cambridge Univ. Press, Cambridge, MA, 1983). [Google Scholar]
- 6.Krieger D, in Endocrinology and Metabolism, Felig P, Baxter JD, Broadus AE, Frohman LA, Eds. (McGraw-Hill, New York, 1981), pp. 125–149. [Google Scholar]
- 7.Baxter JD and Tyrrell JB, ibid, pp. 385–510; Troen P and Oshima H, ibid, pp. 627,–668; Broackus M, ibid, pp. 963–1080. [Google Scholar]
- 8.Ashbrener M, Nature (London) 225, 435 (1980). [Google Scholar]
- 9.Jensen EV and DeSombre ER, Annu. Rev. Biochem 41, 203 (1972); Tompkins GM, Harvey Lect 68 (1974); Tata J, Oppenheimer JH, Koerner D, Schwartz HL, Surks MI,J. Clin. Endocrinol. Metab 35, 330 (1972). [DOI] [PubMed] [Google Scholar]
- 10.Ivarie RD and O’Farrell PH, Cell 13, 41 (1978). [DOI] [PubMed] [Google Scholar]
- 11.Yamamoto K, Annu. Rev. Genet 19, 209 (1985). [DOI] [PubMed] [Google Scholar]
- 12.Ringold G, Annu. Rev. Pharmacol. Toxicol 25, 529 (1985). [DOI] [PubMed] [Google Scholar]
- 13.Wrange O and Gustaffson JA,J. Biol. Chem 253, 856 (1978); Wrange O, Carlstedt-Duke J, Gustaffson JA, ibid 254, 9284, (1979); Okret S, Carlstedt-Duke J, Wrange O, Carlstrom K, Gustafsson JA, Biochim. Biophys Acta 677, 205 (1982).621208 [Google Scholar]
- 14.Carlstedt-Duke J et al. , Proc. Natl. Acad. Sci. U.S.A 79, 4260 (1982); Dellwege HG et al. , EMBO J 1, 285, (1982); Wrange O et al. , J. Biol. Chem 259, 4534 (1984). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Parks WP, Scolnick EM, Kosikowski EH, Science 184, 158 (1974); Ringold G et al. , Cell 6, 299, (1975); Kurtz DT and Feigelson P, Proc. Natl. Acad. Sci. U.S A 74, 4791, (1977); Karin M et al. ,Nature (London) 286, 295, (1980); Hager LJ and Palmiter RD, ibid 291, 340, (1981); Spindler SR, Mellon SH, Baxter JD,J. Biol. Chem 257, 11627, (1982); Evans RM, Birnberg NC, Rosenfeld MG, Proc. Natl. Acad. Sci. U.S.A 79, 7659 (1982).4361099 [Google Scholar]
- 16.Ostrowski MC et al. , EMBOJ 3, 1891 (1984); Govindan MV, Spiess E, Majors J, Proc. Natl. Acad. Sci. U.S.A 79, 5157, (1982); Pfahl M, Cell 31, 475, (1982); Payvar F et al. , ibid 35, 381, (1983); Scheidereit C, Geisse S, Westphal HM, Beato M, Nature (London) 304, 749 (1983). [Google Scholar]
- 17.Karin M et al. , Nature (London) 308, 513 (1984). [DOI] [PubMed] [Google Scholar]
- 18.Robins DM, Pack I, Seeburg PH, Axel R, Cell 29, 623 (1982); Slater ER et al. , Mol. Cell. Biol 5, 2984 (1985). [DOI] [PubMed] [Google Scholar]
- 19.Chandler VL, Maler BA, Yamamoto KR, Cell 33, 489 (1983). [DOI] [PubMed] [Google Scholar]
- 20.Benoist C and Chambon P, Nature (London) 290, 304 (1981); Laimonis LA et al. , Proc. Natl. Acad. Sci. U.S.A 79, 6453, (1982); Banerji J, Olson L, Schaffner W, Cell 33, 729 (1983).6259538 [Google Scholar]
- 21.Jantzen H-M et al. , Cell 49, 29 (1987). [DOI] [PubMed] [Google Scholar]
- 22.Klein-Hitpass L et al. , ibid 46, 1053 (1986). [DOI] [PubMed] [Google Scholar]
- 23.Druege D et al. , Nucleic Acids Res. 14, 9329 (1986); Waterman M et al. , Mol. Endocrinol 2, 14 (1988); Maurer R, personal communication.3467302 [Google Scholar]
- 24.Compton J, Schrader W, O’Malley BW, Proc. Natl. Acad. Sci. U.S.A 80, 161 (1983); Chambon P et al. , Rec. Prog. Hormone Res 40, 1, (1984); von der Ahe D et al. , Nature (London) 313, 706 (1985). [Google Scholar]
- 25.Glass C et al. , Nature (London) 329, 738 (1987). [DOI] [PubMed] [Google Scholar]
- 26.Hollenberg SM et al. , ibid 318, 635 (1985). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Weinberger C et al. , ibid, p. 670. [Google Scholar]
- 28.Debuire B et al. , Science 224, 1456 (1984). [DOI] [PubMed] [Google Scholar]
- 29.Green S et al. , Nature (London) 320, 134 (1986); Greene GL et al. , Science 231, 1150 (1986).3754034 [Google Scholar]
- 30.Krust A et al. , EMBOJ 5, 891 (1986). [Google Scholar]
- 31.Weiler I, Lew D, Shapiro D, Mol. Endocrinol 1, 355 (1987). [DOI] [PubMed] [Google Scholar]
- 32.Loosfelt H et al. , Proc. Natl. Acad. Sci. U.S.A 83, 9045 (1986). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Misrahi M et al. , Biochem. Biophys. Res. Comm 143, 740 (1987). [DOI] [PubMed] [Google Scholar]
- 34.Conneelly O et al. ,Mol. Endocrinol 1,517 (1987); Gronemeyer H et al. , EMBOJ 6, 3985 (1987).3153474 [Google Scholar]
- 35.McDonnell DP et al. , Science 235, 1214 (1987); Baker A et al. , Proc. Natl. Acad. Sci. U.S.A, in press. [DOI] [PubMed] [Google Scholar]
- 36.Arriza J et al. , Science 237, 268 (1987). [DOI] [PubMed] [Google Scholar]
- 37.Weinberger C et al. , Nature (London) 324, 641 (1986). [DOI] [PubMed] [Google Scholar]
- 38.Sap J et al. , ibid, p. 635. [Google Scholar]
- 39.Giguere V, Yang N, Segui P, Evans RM, ibid 331, 91 (1988). [DOI] [PubMed] [Google Scholar]
- 40.Thompson CC, Weinberger C, Lebo R, Evans RM, Science 237, 1610 (1987). [DOI] [PubMed] [Google Scholar]
- 41.Cerelli G, Thompson C, Damm K, Evans RM, unpublished observation; Benbrook D and Pfahl M, Science 230, 788 (1987). [Google Scholar]
- 42.Giguere V et al. , Nature (London) 330, 624 (1987). [DOI] [PubMed] [Google Scholar]
- 43.Petkovich M et al. , ibid, p. 444; Robertson M, ibid, p. 420. [Google Scholar]
- 44.Dejean A et al. , ibid 322, 70 (1986). [Google Scholar]
- 45.deThe H et al. , ibid 330, 667 (1987). [Google Scholar]
- 46.Segraves W, thesis, Stanford University; (1988). [Google Scholar]
- 47.Johnson MS and Doolittle RF,J. Mol. Evol 23, 267 (1986). [DOI] [PubMed] [Google Scholar]
- 48.Hollenberg S et al. , Cell 49, 39 (1987). [DOI] [PubMed] [Google Scholar]
- 49.Godowski PJ et al. , Nature (London) 325, 365 (1987). [DOI] [PubMed] [Google Scholar]
- 50.Becker PB et al. , ibid 324, 606 (1986). [Google Scholar]
- 51.Wellman T and Beato M, ibid, p. 688. [Google Scholar]
- 52.Giguere V et al. , Cell 46, 645 (1986).3742595 [Google Scholar]
- 53.Kumar V, Green S, Staub A, Chambon P, EMBOJ 5, 2231 (1986). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Green S and Chambon P, Nature (London) 325, 75 (1987). [DOI] [PubMed] [Google Scholar]
- 55.Miller J, McLachlan A, Klug A, EMBOJ 4, 1609 (1985). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Evans RM and Hollenberg S, Cell 52, 1 (1988). [DOI] [PubMed] [Google Scholar]
- 57.Hollenberg S, Giguere V, Evans RM, unpublished observations.
- 58.Sabbah M et al. , J. Biol. Chem 262, 8631 (1987). [PubMed] [Google Scholar]
- 59.Huckaby C et al. , Proc. Natl. Acad. Sci. U.S.A 84, 8380 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Danielson M et al. , Mol. Endocrinol 1, 816 (1987). [DOI] [PubMed] [Google Scholar]
- 61.Yamamoto K et al. , Rec. Prog. Hormone Res 32, 3 (1976). [DOI] [PubMed] [Google Scholar]
- 62.Kumar V et al. , Cell 51, 941 (1987). [DOI] [PubMed] [Google Scholar]
- 63.O’Malley B, personal communication.
- 64.Brown MS and Goldstein JL, in The Pharmacological Basis of Therapeutics, Gilman AG, Goodman LS, Rall TW, Murad F, Eds. (Macmillan, New York, 1985), pp. 827–845. [Google Scholar]
- 65.Sudhof J, Aussel D, Brown M, Goldstein J, Cell 48, 1061 (1987). [DOI] [PubMed] [Google Scholar]
- 66.Poland A, Glover E, Kende A, J. Biol. Chem 251, 4936 (1976). [PubMed] [Google Scholar]
- 67.Sledge GW and McGuire WL, Adv. Cancer Res 38, 61 (1983). [DOI] [PubMed] [Google Scholar]
- 68.Addison T, On the Constitutional and Local Effects of Disease of the Suprarenal Capsules (S. Highley, London, 1855). [Google Scholar]
- 69.Sapolsky RM,Neurosci J. 6, 2240 (1986). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Thaller C and Eichele G, Nature (London) 327, 625 (1987). [DOI] [PubMed] [Google Scholar]
- 71.Strickland S and Mahdavi V, Cell 15, 393 (1978); Jetten A et al. , Exp. Cell. Res 124, 381, (1979); Wang S-Y et al. , Dev. Biol 107, 75 (1985). [DOI] [PubMed] [Google Scholar]
- 72.Ptashne M, A Genetic Switch (Cell Press, Cambridge, MA, 1986). [Google Scholar]
- 73.Keegan L, Gill G, Ptashne M, Science 231,699 (1986); Brent R and Ptashne M, Cell 43, 729 (1985). [DOI] [PubMed] [Google Scholar]
- 74.Hope I and Struhl K, Cell 46, 885 (1986). [DOI] [PubMed] [Google Scholar]
- 75.Giniger E and Ptashne M, Nature (London) 330, 670 (1987). [DOI] [PubMed] [Google Scholar]
- 76.Morgan TH, in Nobel Lectures in Molecular Biology, Baltimore D, Ed. (Academic Press, New York, 1977), pp. 3–18; Morgan TH , The Theory of the Gene (Yale Univ. Press, New Haven, 1926). [Google Scholar]
- 77.Miesfeld R et al. , Cell 46, 389 (1986). [DOI] [PubMed] [Google Scholar]
- 78.Danielson M et al. , EMBOJ 5, 2513 (1986). [Google Scholar]
- 79.Law ML et al. , Proc. Natl. Acad. Sci. U.S.A 84, 2877 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Thompson C and Evans RM, unpublished observations.
- 81.Cato A et al. , EMBOJ 5,2237 (1986). [Google Scholar]
- 82.Rusconi S et al. , ibid 6, 1309 (1987). [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mattei M et al. , Hum. Genet, in press. [Google Scholar]
- 84.I thank J. Arriza for helping to compile data, members of the Gene Expression Laboratory for discussion, and R. Doolittle for computer analysis of receptor homologies. I also thank G. Wahl, M. McKeown, B. Sefton, T. Hunter, and other members at the Salk Institute for advice and critical reading of the manuscript. I acknowledge many colleagues who shared data prior to publication and thank E. Stevens for expert administrative and secretarial assistance. Supported by the Howard Hughes Medical Institute and a grant from the National Institutes of Health.