

Abstract
Background
Plant pathogenic fungi of the genus Fusarium infect a wide array of crops and produce numerous health-threatening mycotoxins. Recently, we found that larvae of the common pest of stored products Tenebrio molitor preferably fed on grains colonized with Fusarium proliferatum. We draw the hypothesis that the increased attractiveness of infected grains for mealworms facilitates dispersal of the fungus. In this work we examined the dissemination of F. proliferatum and further Fusarium spp. by adults of T. molitor.
Results
Mealworm beetle Tenebrio molitor transmitted Fusarium species F. avenaceum, F. culmorum, F. poae, and F. proliferatum to wheat grains with varying efficiency. F. proliferatum was disseminated most efficiently: 20 days after feeding on Fusarium cultures, the beetles still transmitted F. proliferatum to most grains exposed to feeding. The transmission of F. culmorum gradually declined over time and the transmission of the other Fusarium spp. ceased completely 20 d after beetles feeding of fungal cultures. Propagules of F. proliferatum and F. culmorum were traceable in beetles' feces for 20 days while no colonies of F. poae and F. avenaceum were detectable after 5 days. Because F. proliferatum was transmitted by mealworms most efficiently, this species was further investigated. Mealworm beetles T. molitor preferred feeding on grains colonized with F. proliferatum as compared to uninfected grains. Male beetles infected with F. proliferatum transmitted the fungus by copulation.
Conclusions
Efficient dissemination of F. proliferatum by mealworm beetle together with the feeding preference of the beetle for grains colonized with F. proliferatum show that the chemical phenotype of the fungus responsible for the enhanced attractiveness of infected grains is subjected to positive selection. This indicates that adaptation of F. proliferatum to transmission by insects involved an alteration of insects' feeding preferences.
Introduction
Colonization of grains by Fusarium species impairs food security by diminishing the food quality by mycotoxins contamination. F. proliferatum has a wide host range. The pathogen has been most frequently isolated from maize, rice, sorghum and asparagus, but it was also found in banana [1], citrus fruits [2], date palm [3] and pine seedlings [4]. Since 1990, F. proliferatum is known to infect wheat [5–9]. F. verticillioides and F. proliferatum are the main source of mycotoxins fumonisins in food and feed products [8, 10]. Contamination with fumonisins raises food safety concerns because fumonisins exert acute toxicity as well as carcinogenic and teratogenic effects [11]. Many surveys reported association between high levels of fumonisins in maize with outbreaks of equine leukoencephalomalacia [12] and swine pulmonary edema [13]. Besides fumonisins, F. proliferatum is capable of synthesizing mycotoxins beauvericin [14], fusaproliferin [15], fusarins [16], and moniliformin [17].
Insects represent an important route in the transmission of numerous pathogens among plants [18]. Fermaud and Menn [19] reported that the grape berry moth Lobesia botrana transmitted fungus Botrytis cinerea from infected to healthy berries. Paine et al. [20] discussed that fungi of genus Ophiostoma may benefit from the association with bark beetles by being transported to new host trees. Beetles (Dendroctonus sp.) may benefit from the association by feeding on the fungus, or because pathogenic fungi kill the host tree. Adult corn earworm feeding on the honeydew secreted by Claviceps africana transmits ergot fungus from diseased to healthy panicles [21]. The rust fungus Puccinia monoica inhibits flowering of its host plants (Arabis species) and transforms infected leaves in a way mimicking true flowers, attracting pollinating insects [22]. Weevil benefit from feeding on rust-infected tissue and the fungus benefits from the transmission of its spores by weevils [23].
Mealworm beetle (syn. darkling beetle) Tenebrio molitor and its larvae (mealworms or yellow mealworms) are pests of stored grains. Recently we found that colonization of wheat grains with Fusarium spp. affected the food choice of mealworms [24]. Grains colonized with F. proliferatum were more attractive to larvae of T. molitor than uninfected grains. Because wheat plants can be systemically infected with F. proliferatum via colonized grains [5], we hypothesized in the current study that the attraction of T. molitor to grains infected with F. proliferatum facilitates dissemination of the fungus. Here we tested this hypothesis by investigating the dissemination of F. proliferatum and other Fusarium species in wheat grains by the beetles T. molitor.
Materials and methods
Organisms and media
T. molitor was reared on whole wheat flour with 5% yeast extract in a climate chamber in darkness at 27 ± 2°C and a relative humidity of 65 ± 5%. Before the experiments, the beetles were starved for 24 h for cleaning up the remaining food and also to ensure feeding on the mycelia [25]. For surface sterilization, beetles were carefully washed with autoclaved distilled water, soaked in 1.2–1.3% sodium hypochlorite for 3 min followed by 3-times rinsing with autoclaved distilled water for 2 min each. The beetles were dried on autoclaved paper and covered by sterile plastic lid under laminar flow cabinet.
Fungal strains (Table 1) have been described in previous work [24]. Fungal cultures were grown on potato dextrose agar (PDA, Merck, Darmstadt, Germany) plates in darkness at 23°C.
Table 1. Fungal strains.
| Species | Host | Year | Origin |
|---|---|---|---|
| Fusarium avenaceum 1.27 | wheat grains | 2008 | Poppelsdorf, Bonn, Germany |
| Fusarium culmorum 3.37 | wheat | 2004 | Klein-Altendorf, Bonn, Germany |
| Fusarium poae DSM 62376 | oat | 1990 | DSMZ, Braunschweig, Germany |
| Fusarium proliferatum 21.1 | maize | 2007 | Hainichen, Germany |
| Beauveria bassiana Bea2 | black vine weevil | 1989 | Stuttgart, Germany |
CLA (Carnation Leaf-piece Agar) was prepared by placing two sterile pieces of carnation leaves onto 2% water agar plate (20 g agar in 1 L of water) [26]. Carnation leaves were harvested from plants grown without fungicide or insecticide application and dried in an oven at 70°C for 3–4 h until brittle. Dried leaves were sterilized with 1.5% sodium hypochlorite and dried at room temperature under a clean bench. PDB (potato dextrose broth) was purchased from Merck (Darmstadt, Germany). CZID (Czapek-Dox-Iprodione-Dichloran) agar was prepared as described [27, 28] and amended with streptomycin, ampicillin and chlortetracycline (Applichem, Darmstadt, Germany), each at 0.1 g/L.
Fungal DNA content and survival of F. proliferatum in T. molitor
For the estimation of fungal DNA and mycotoxin content in the insects, ten T. molitor beetles for each of four time points were allowed to graze for 24 h on F. avenaceum, F. culmorum, F. poae, and F. proliferatum cultures on PDA plates and subsequently transferred into sterile Petri dishes with autoclaved wheat grains. Entomopathogenic fungus B. bassiana was used as a control. Beetles were harvested on 1, 5, 10 and 15 d and DNA content of Fusarium DNA and mycotoxins was determined as described below. To determine whether F. proliferatum survived in the bodies of beetles, 10 T. molitor beetles were fed F. proliferatum mycelia on PDA plates until death (longest time period 15 d). Dead beetles were frozen at -20°C, surface-sterilized with 1% sodium hypochlorite for 3 min, rinsed 3 times with sterile distilled water and placed on CZID plates. The outgrowing mycelia were monitored and examined for taxonomic affiliation with F. proliferatum.
Dissemination of Fusarium spp. by T. molitor
Transmission of fungal inoculum by T. molitor to healthy grains was investigated by placing individual beetles that had been previously fed on fungal cultures into arenas with uncontaminated wheat grains and monitoring grain infection as well as the presence of fungal propagules in beetles' excreta over a time course. The experiment was started by feeding 10 mealworm beetles on cultures of F. avenaceum, F. culmorum, F. poae, F. proliferatum, and B. bassiana on PDA plates for 24 h. After feeding, the beetles were placed into sterile Petri dishes with 50–60 autoclaved wheat grains each, a single beetle per plate. The grains were changed every 24 h. After 1, 5, 10, 15, and 20 d, fifteen grains were randomly selected from each Petri dish, placed on CZID plates and kept in darkness at 25°C to determine fungal colonization. Grains with outgrowing mycelia were counted, generating a colonization score from 0 to 15 for each animal and time point. At the same time, excreta (feces) of the beetles were collected, mashed in sterile water and spread on CZID plates. Fungal colonies on the plates with excreta were counted, resulting in a cfu value for each animal and time point. The experiment was started with 10 beetles for each fungal species but not all beetles survived till the last sampling. B. bassiana proved lethal: all beetles died within 5 days after feeding. These data were excluded from the analysis. Some beetles fed on cultures of Fusarium spp. died, too, but at least 5 beetles from each Fusarium culture survived till the end of the experiment. Therefore we used data from 5 beetles for each Fusarium spp. in the analysis, keeping the number of replicates at the same value of 5. The entire experiment was repeated twice; because the results were essentially the same, only data from one experiment are shown.
Food choice experiment with F. proliferatum-infected grains
Wheat grains were soaked in distilled water for 18 h at room temperature. 20 g of soaked grains with 5 ml water were autoclaved in 100 ml Erlenmeyer flask for 20 min at 121°C, inoculated with 2 ml F. proliferatum conidia (1.3×106 spores/ml) and incubated for 7 days to ensure complete colonization. Control wheat grains were treated in a same way without inoculation. Food preference of mealworm beetle T. molitor was investigated in Petri dishes of a diameter 142 mm marked to generate four equal sectors in the form of pie slices. 6 g of non-colonized wheat grains each were placed into opposing sectors of the plates and 6 g grains colonized with F. proliferatum to the other sectors. Ten T. molitor beetles were placed into the center of the dishes. After 20 min in dark, beetles in each sector were counted. Controls were prepared in the same way with all four sectors filled with autoclaved not infected grains and two opposite sectors randomly selected for counting. Ten replicates with 10 animals each were performed and the experiment was repeated twice.
Microscopic analyses
Beetles fed on 2-weeks-old F. proliferatum culture for 24 h were sputter-coated with gold and examined by scanning electronic microscopy on Phenom G2 Pro (Phenom World, Eindhoven, Netherland) for fungal mycelia and conidia attached to their body. Beetles tested for fungal colonization were surface sterilized with 1.2–1.3% sodium hypochlorite for 3 min, rinsed 3 times with sterile distilled water for 2 min and placed on CZID plates. Images of the specimens observed under light microscope were recorded with a camera incorporated to the Leitz DMRB Leica light microscope using software Diskus 4.2 (Hilgers, Königswinter, Germany).
Re-isolation of fungi from grains and beetles
Contaminated wheat grains and excreta, gut and eggs of beetles were surface sterilized with 1.2–1.3% sodium hypochlorite and cultured on CZID plates. Single colony or hyphal tips were transferred to both PDA and CLA plates. The taxonomic affiliation of fungal isolates was confirmed by morphological features, such as pigmentation of the mycelia and the form of micro- and macro-conidia. For DNA extraction, single colonies were cultivated in 50 ml PDB at 25°C for 4 d. The mycelia were freeze-dried and ground into fine powder for DNA extraction.
Mycotoxin analysis
Beetles were dried in vacuum at 40°C overnight and ground. Mycotoxins were extracted as described before [29]. Beauvericin, fumonisin B1 and enniatins were separated on an RP column at 40°C followed by electrospray ionization in a positive mode and analysis on ion trap 500 MS (Varian, Darmstadt, Germany) [29]. The samples for trichothecenes B and zearalenone quantification were defatted with cyclohexane. Deoxynivalenol and zearalenone were separated on the same HPLC system and detected by tandem mass spectrometry using triple quadrupole 1200 L (Varian, Darmstadt, Germany) according to published methods [30, 31]. Two mass transitions were used for each toxin.
Calibration curves were constructed using analytical standards dissolved in methanol/water (1:1). The estimated limits of quantification (LOQ) and detection (LOD) for beauvericin and enniatins A, B, A1, and B1 were 150 ng/g and 60 ng/g, respectively. LOQ and LOD for fumonisin B1 were 390 ng/g and 190 ng/g, respectively. LOD for deoxynivalenol and zearalenone were 200 ng/g and 20 ng/g, respectively.
DNA extraction, species-specific detection and quantification of fungal DNA
Total DNA of T. molitor was extracted using a CTAB method [32] and dissolved in TE buffer (10 mM Tris, 1 mM EDTA, pH 8.0). The quality and quantity of DNA were estimated by gel electrophoresis in 0.8% agarose gels (Cambrex, Rockland, USA) prepared in TAE buffer [33]. The electrophoresis was carried out at 4 V/cm for 60 min. The gel was stained with ethidium bromide (2 mg/l) and documented with a digital imaging system (Vilber Lourmat, Marne la Vallee, France). Prior PCR analysis, an inhibition assay was carried out to test the effect of matrix on DNA amplification.
DNA of fungal cultures re-isolated from wheat grains, insect excreta, and gut and eggs and grown in PDB was extracted using DNeasy Plant Mini kit (QIAGEN, Hilden, Germany) according to the manufacturer’s protocol. The DNA quality was monitored by electrophoresis in 1.0% agarose gels (Agarose NEEO Ultra-Quality, Carl Roth, Karlsruhe, Germany) prepared in TAE buffer [33]. The electrophoresis was carried out at 4 V/cm for 30 min. The agarose gel was stained with 10,000 × dilution Gel Red (Biotium, Darmstadt, Germany) and documented with a digital imaging system (Gel Doc, Bio-Rad, Munich, Germany).
Thermocycler (CFX384, BioRad, Munich, Germany) was used for real-time PCR analysis. Primers MGBF/R [34] and Fp 82F/R [35] were used for species-specific quantification of F. avenaceum and F. poae, respectively. The reactions were carried out in polymerase buffer (16 mM (NH4)2SO4, 67 mM Tris-HCl, 0.01% (v/v) Tween-20, pH 8.8 at 25°C) with 0.15 mM of each dNTP (Bioline, Luckenwalde, Germany), 2.5 mM MgCl2, 0.1 U of Taq DNA polymerase (BIOTaq, Bioline, Luckenwalde, Germany), 0.3 μM of each primer, and 0.1 × SYBR Green I (Invitrogen, Karlsruhe, Germany). F. culmorum and F. proliferatum DNA was quantified according to established protocols [29, 32]. The lowest standards set as limits of quantification were 0.169 pg/μl for F. avenaceum, F. culmorum, F. poae and 2.09 fg/μl for F. proliferatum.
Taxonomic identity of re-isolated F. proliferatum was confirmed by PCR with species-specific primers PRO1/PRO2 [36]. The reaction mixture contained Dream Taq buffer (Thermo Fisher Scientific, Darmstadt, Germany) with 4 mM MgCl2, 0.2 mM of each of the four deoxynucleoside triphosphates, 0.1 U of Dream Taq DNA polymerase (Thermo Fisher Scientific, Darmstadt, Germany), 0.5 μM of each primer and 2 μl sample DNA. The following cycling conditions were used: 1 cycle of 10 min at 94°C, 35 cycles of 60 s at 94°C, 30 s at 60°C, and 60 s at 72°C, followed by a final extension cycle at 72°C for 10 min [37]. 1% agarose gel was prepared in TAE buffer [33] and the electrophoresis was carried out at 4 V/cm for 30 min. The gel was stained with Gel Red and documented with a digital imaging system (Bio-Rad, Munich, Germany).
Statistical analyses
Statistical analysis was performed with R 3.5.0 [38]. Data for infected grains were analyzed using logistic regression. Colonies counts were analyzed using a log-linear model. The results of the food choice experiments were evaluated using Chi-Square Goodness of Fit test. Box plots drawn using Excel 2016 show interquartile range with the median and mean; whiskers indicate the largest and smallest observation or 1.5-fold of the interquartile range, whichever was smaller/larger. The number of replicates and the number of grains or animals per replicate are specified in the description of the experiments in Material and Methods section.
Results
Dissemination of Fusarium spp. to new grains by mealworm beetle T. molitor
Transmission of fungal inoculum to healthy grains by the beetle T. molitor was investigated by placing beetles fed on fungal cultures into Petri dishes with wheat grains. The grains were replaced daily and their contamination with Fusarium spp. was monitored for 20 d (Fig 1). F. proliferatum was disseminated most efficiently: no decline in the contamination of new grains with F. proliferatum was observed with contamination rates remaining above 90% till the end of the experiment. The contamination with F. culmorum gradually decreased over time while F. avenaceum and F. poae became undetectable after 10 d and 15 d, respectively. B. bassiana killed all beetles within 5 d, preventing transmission to new grains. The data fitted a generalized linear model with quasibinominal error structure and revealed that the contamination rates of F. proliferatum were significantly high (p < 0.01) when compared to that of F. avenaceum, F. poae and control. The original data were shown in S1 Table.
Fig 1. Transmission of Fusarium spp. to wheat grains by Tenebrio molitor.

The graph shows percentage of wheat grains that became infected with Fusarium spp. after exposure to beetles of T. molitor that were previously fed on fungal cultures. Means ± S.D. are shown (n = 10).
Survival of propagules of Fusarium spp. in the digestive track of mealworm beetle Tenebrio molitor
Dissemination of ingested fungal propagules by feces is only possible if the propagules survive the gut passage. To investigate this possibility, feces of beetles fed on fungal cultures were collected for 20 d, suspended in water and plated on Fusarium-specific agar media. Propagules of all fungal Fusarium species tested survived the gut passage (Fig 2). F. proliferatum was disseminated most efficiently, followed by F. culmorum. The density of F. avenaceum and F. poae propagules in the excreta declined rapidly, becoming undetectable after 5 d. The data fitted a generalized linear model and revealed that colony forming units of F. proliferatum were significantly different (p < 0.01) compared to that of F. avenaceum, F. culmorum and F. poae. Dissemination of B. bassiana could not be detected because all beetles fed on B. bassiana culture died within 5 d. No colonies of Fusarium spp. grew out of feces of beetles that were not fed on Fusarium cultures. The original data were shown in S2 Table.
Fig 2. Dissemination of propagules of Fusarium species by feces of mealworm beetle Tenebrio molitor that has been fed on Fusarium cultures.

The density of fungal propagules determined as the number of Cfu (colony forming units) in the excreta of T. molitor adults fed on cultures of Fusarium spp. is shown as mean ± S.D.
Fungal proliferation and mycotoxin accumulation in T. molitor beetles
Dissemination of fungal propagules by mealworm beetles is expected to last longer if the fungus multiplies in beetles' digestion track. T. molitor beetles were allowed to graze on F. avenaceum, F. culmorum, F. poae, F. proliferatum, and B. bassiana cultures, transferred into sterile Petri dishes with autoclaved wheat grains, harvested at different time points and the content of Fusarium DNA in the beetles was determined. DNA of all Fusarium species except F. culmorum was detected in most beetles until the last time point at 15 d (Table 2). Beetles fed on F. culmorum died within 10 d. Because only a single animal fed on F. culmorum survived till 15 d, the result of qPCR analysis was disregarded. Large variation in the content of fungal DNA among individuals at all time points (Table 2) prevented us from testing whether fungal biomass in the beetles during the study period was growing.
Table 2. Fungal DNA in beetles grazing on Fusarium spp. mycelia and conidia.
|
Fungal species |
Species-specific fungal DNA in beetles (ng/g) | |||
|---|---|---|---|---|
| 1 d | 5 d | 10 d | 15 d | |
| Fusarium avenaceum | 87 ± 27 (7/10) |
47 ± 18 (5/10) |
106 ± 22 (9/10) |
153 ± 67 (4/4) |
| Fusarium culmorum | 110 ± 16 (10/10) |
34 ± 8 (7/10) |
186 ± 35 (9/10) |
<LOQ (1/1) |
| Fusarium poae | 3 (1/10) |
5 ± 4 (2/10) |
41 ± 40 (3/10) |
3 ± 1 (7/8) |
| Fusarium proliferatum | 258 ± 45 (10/10) |
35 ± 4 (8/10) |
100 ± 30 (10/10) |
29 ± 14 (8/8) |
Means and standard error of the mean of positive samples are shown. The number of beetles with detectable fungal DNA and the total number of beetles investigated are shown in brackets.
Mycotoxin content in beetles fed on Fusarium cultures was analyzed. Beauvericin was produced by three Fusarium species including the most efficiently disseminated F. proliferatum. Examination of the presence of beauvericin in the beetles at different time points showed that beetles fed on F. poae were consistently contaminated with beauvericin, while beetles grazing on F. proliferatum only rarely contained beauvericin (Table 3). Beetles consuming B. bassiana died within 5 days; all beetles contained beauvericin at this time point (Table 3). Deoxynivalenol, nivalenol and zearalenone were not detectable in beetles fed on F. culmorum (Table 4). Enniatins were detected in beetles fed on all fungal species; it turned out that wheat grains used for the experiment were naturally contaminated with enniatins at 9.0 ± 1.0 ng/g.
Table 3. Beauvericin contamination of T. molitor beetles.
| Fungal species | Time | Beetles contaminated with beauvericin |
|---|---|---|
|
F. poae |
1 d | 4/10a |
| 5 d | 1/10 | |
| 10 d | 10/10 | |
| 15 d | 4/8 | |
| F. proliferatum | 1 d | 2/10 |
| 5 d | 2/10 | |
| 10 d | 1/10 | |
| 15 d | 0/9 | |
| B.bassiana | 1 d | 1/10 |
| 5 d | 10/10 | |
| 10 d | / | |
| 15 d | / |
The limits of quantification (LOQ) for beauvericin was 150 ng/g. Slash indicates that all beetles died before sampling.
a Number of beetles contaminated with beauvericin/total number of beetles.
Table 4. Mycotoxin content in T. molitor beetles.
| Fungal species | Beauvericin | Fumonisin B1 | Enniatin A | Enniatin B | Enniatin A1 | Enniatin B1 | Deoxynivalenol | Zearalenone | |
|---|---|---|---|---|---|---|---|---|---|
| (ng/g) | |||||||||
| Control | 1 d | <LOQ | <LOQ | <LOQ | 6 ± 1 | <LOQ | 2 ± 0 | <LOQ | <LOQ |
| 5 d | <LOQ | <LOQ | <LOQ | 5 ± 0 | <LOQ | 7 ± 4 | <LOQ | <LOQ | |
| 10 d | <LOQ | <LOQ | <LOQ | 15 ± 1 | 2 ± 1 | 7 ± 1 | <LOQ | <LOQ | |
| 15 d | <LOQ | <LOQ | <LOQ | 5± 0 | <LOQ | 3 ± 0 | <LOQ | <LOQ | |
| F. avenaceum | 1 d | <LOQ | <LOQ | <LOQ | 246 ± 32 b | 275 ± 8 b | 228 ± 42 | / | / |
| 5 d | <LOQ | <LOQ | <LOQ | 406 ± 47 | 261 ± 18 b | 296 ± 30 | / | / | |
| 10 d | <LOQ | <LOQ | <LOQ | 552 ± 49 b | 291 ± 22 b | 355 ± 37 b | / | / | |
| 15 d | <LOQ | <LOQ | <LOQ | 215 ± 20 b | <LOQ | 177 ± 5 b | / | / | |
| F. poae | 1 d | 251 ± 28 b | <LOQ | <LOQ | <LOQ | 202 ± 10 b | 156 ± 9 | / | / |
| 5 d | 176 b | <LOQ | <LOQ | 365 b | 259 ± 25 | 220 ± 32 | / | / | |
| 10 d | 843 ± 187 b | <LOQ | <LOQ | 234 ± 12 b | 336 ± 26 | 305 ± 33 | / | / | |
| 15 d | 215 ± 28 b | <LOQ | 212 b | <LOQ | 221 ± 7 | 175 ± 6 | / | / | |
| F. proliferatum | 1 d | 1383 ± 1171 b | <LOQ | <LOQ | 218 ± 70 b | 265 ± 74 b | 230 ± 55 | / | / |
| 5 d | 499 ± 330 b | <LOQ | <LOQ | 257 ± 17 b | 207 ± 8 b | 203 ± 15 | / | / | |
| 10 d | <LOQ | <LOQ | <LOQ | 466 ± 46 b | 209 ± 14 b | 309 ± 31 b | / | / | |
| 15 d | <LOQ | <LOQ | <LOQ | 185 ± 11 b | 225 ± 6 b | 166 ± 4 | / | / | |
| F. culmorum | 1 d | / | / | / | / | / | / | <LOQ | <LOQ |
| 5 d | / | / | / | / | / | / | <LOQ | <LOQ | |
| 10 d | / | / | / | / | / | / | <LOQ | <LOQ | |
| 15 d | / | / | / | / | / | / | <LOQ | <LOQ | |
| B. bassiana | 1 d | 161b | <LOQ | <LOQ | 240 ± 15 b | 330 ± 33 | 283 ± 37 b | / | / |
| 5 d | 1759 ± 511 | <LOQ | 413 ± 81 b | 232 ± 26 | 279 ± 15 | 224 ± 14 | / | / | |
Notes
The values indicated mean values and respective standard error of mean. Slash indicated that the mycotoxin was not analyzed. The limits of quantification (LOQ) of beauvericin, enniatin A, enniatin B, enniatin A1, enniatin B1 were 150 ng/g while LOQ of fumonisin B1 was 390 ng/g.
b not all 10 samples contained detectable amount of the toxin.
Food preference of T. molitor for grains colonized with F. proliferatum
Because of long-lasting dissemination of F. proliferatum by mealworm beetles that were previously fed on F. proliferatum (Fig 1), we investigated how colonization with F. proliferatum affected the food preference of the beetles. T. molitor strongly preferred grains infected with F. proliferatum over uninfected grains (Fig 3). The experiment was repeated twice with essentially the same result.
Fig 3. Food preference of Tenebrio molitor for wheat grains colonized with Fusarium proliferatum.
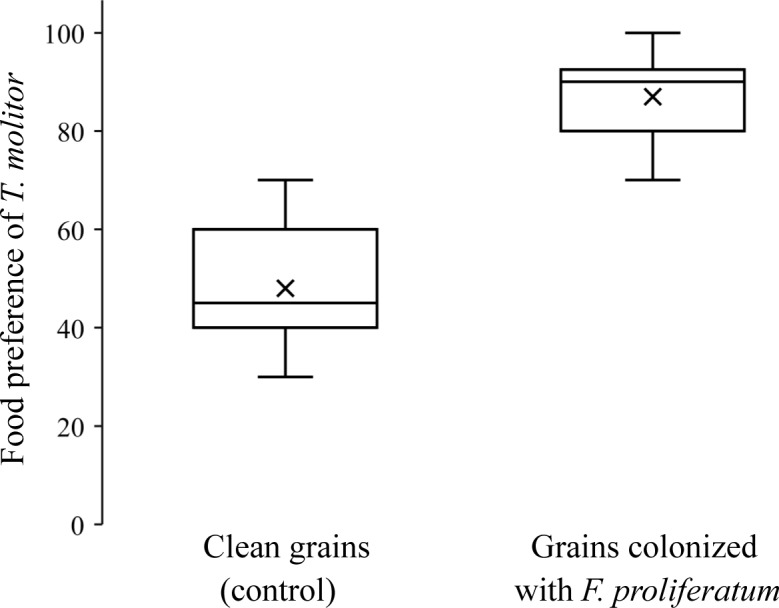
Mealworm beetles T. molitor were offered wheat grains colonized with F. proliferatum and uninfected grains. After 20 min, beetles feeding on grains with F. proliferatum were counted and the count was expressed as a percentage of all beetles in the arena. In the control, both food choices consisted of uninfected grains. The preference of the beetles for infected grains was statistically significant (chi-square test, Χ2 = 59.3, p < 0.001).
F. proliferatum propagules adhere to the body of T. molitor
The presence of propagules of the F. proliferatum on the surface of beetles' bodies was examined by electron microscopy. Fungal mycelia (Fig 4A) and conidia (Fig 4B) were observed on the surface of beetles fed on F. proliferatum cultures. The fungus isolated from these samples was confirmed to be F. proliferatum. We also found fungal conidia attached on antennae (Fig 5B with the control 5A), mouthparts (Fig 5D with the control 5C), wings (Fig 5F with the control 5F) and legs (Fig 5H with the control 5G) of the beetles. No mycelia or conidia were found on beetles that did not have contact with a fungal culture.
Fig 4. Hyphae and conidia on the body of Tenebrio molitor fed on Fusarium proliferatum.

Scanning electron microscopy photographs show F. proliferatum hyphae (a) and conidia (b) adhering to the cuticle of T. molitor beetles that were fed on F. proliferatum culture.
Fig 5. Hyphae and conidia on the extremities of Tenebrio molitor fed on Fusarium proliferatum.

Scanning microscopy photographs show F. proliferatum conidia attached to the antennae (b; a is a control), mouthparts (d; c is a control), wings (f, e is a control), and legs (h; g is a control).
Fungal mycelia grew out of dead bodies of beetles fed on F. proliferatum cultures (Fig 6A and 6B). The mycelia were confirmed to belong to F. proliferatum by PCR with species-specific primers.
Fig 6. Growth of Fusarium proliferatum from dead bodies, feces and eggs of Tenebrio molitor and from contaminated grains.

The upper part of the figure shows fungal hyphae growing out of the bodies of dead beetles fed on F. proliferatum (a and b) and wheat grains contaminated with F. proliferatum by the beetles (c and d). The lower part of the figure shows fungal colonies growing from feces of beetles suspended in water and plated on agar medium (e), and an egg of T. molitor contaminated with F. proliferatum via copulation (f).
Transmission of F. proliferatum to the next generation of T. molitor by copulation
The long-lasting dissemination of F. proliferatum by mealworm beetles (Fig 4) raised the question whether the fungus can be transmitted from male beetles to offspring by copulation. Five male beetles fed on F. proliferatum for 48 h were mated with five females. After 7 d, the females were dissected and 9–11 eggs from each female (a total of 50 eggs) were picked up with sterilized forceps and placed on CZID plates for 3–4 d at 25°C. The emergence of Fusarium mycelia (Fig 6F) was recorded. 50 eggs from 5 female beetles after copulation with males fed on clean grains were used as negative controls. Out of the eggs from female beetles inseminated by males fed on F. proliferatum, outgrowth of fungal mycelia was detected in 30 eggs, with 44–80% contaminated eggs per beetle. The taxonomic assignment of fungal colonies to F. proliferatum was confirmed by morphology and species-specific PCR. Except for a single case of contamination with an Aspergillus species, no fungal growth was observed on control eggs.
Discussion
Seed-borne infection of wheat plants with F. proliferatum causes systemic colonization of the plants and mycotoxins accumulation in wheat grains [5]. Transmission of F. proliferatum in storage from infected to uninfected grains might increase the incidence of wheat plants grown from stored seeds. This work provides evidence that T. molitor is capable of disseminating F. proliferatum efficiently among stored grains.
It is well known that pest insects may increase mold incidence in stored grains [39]. To our knowledge, this phenomenon has not been exploited in an ecological context, especially regarding the food preference of pests. Our previous work showed that colonization of grains with F. proliferatum increased their attractiveness to mealworms (larvae of Tenebrio molitor) [24]. In the current work, adult beetles of T. molitor exhibited a strong preference for grains colonized with F. proliferatum and T. molitor beetles fed on F. proliferatum disseminated the fungus to fresh grains at a high rate for an extended period of time. It is tempting to hypothesize that the increased attractiveness of grains infected with F. proliferatum enhances the fitness of the fungus by accelerating its dissemination. Positive selection may maintain the chemical phenotype of F. proliferatum that is responsible for its attractiveness to the beetles. Furthermore, the dissemination of F. proliferatum by T. molitor may select F. proliferatum strains that allow host beetles to survive the infection for an extended time period. A rigorous test of this hypothesis would require quantitative estimation of fungal fitness, accounting for dissemination by T. molitor as well as for the loss of fungal propagules due to feeding. The strong food preference of the beetles for infected grains and the high efficiency and long persistence of the dissemination of fungal propagules by T. molitor however suggest that the dissemination of F. proliferatum by insects is an integral part of its life history.
Why was only F. proliferatum disseminated efficiently by T. molitor, while the other Fusarium species disappeared completely 20 days after the contact with the beetles? All four Fusarium species infect flowers of grain crops, followed by the colonization of developing grains. In grain crops, F. proliferatum occurs mainly in maize; infection of wheat with F. proliferatum has recently been established but the fungus appears to be only a minor pathogen of wheat. It has not been reported from other small-grain cereals so far. The other three Fusarium species occur frequently on maize as well as on small-grain cereals and all of them are components of the Fusarium Head Blight disease complex. None of the species form races. Thus specialization for host plants is unlikely to explain the differences in the dissemination rates of Fusarium species by T. molitor.
F. proliferatum however differs from the other three Fusarium species regarding its association with insects. A recent survey of entomopathogenic Fusarium species associated with Tribolium beetles recovered only two Fusarium species: F. keratoplasticum and F. proliferatum [40]. Furthermore, F. proliferatum was used to control wheat flour beetle Tribolium castaneum [41]. The ability of F. proliferatum to infect insects may be related to its long-term dissemination by T. molitor beetles that had been exposed to F. proliferatum culture. Association of the other three Fusarium species used in this study with insects has been occasionally reported, too. For instance, F. avenaceum, F. culmorum, and F. poae were isolated from the housefly Musca domestica, the clover leaf weevil Hypera punctata, and the grass hopper Melanoplus bivittatus, respectively [42]. These reports however reflect rare and spurious observations rather than established fungus-insect associations. A recent review concluded that in spite of an increase of Fusarium diseases of grain crops with herbivore infestation reported in many studies, vector activity of the insects has rarely been demonstrated [43]. In a few publications reporting direct transmission of Fusarium spp. by insects, either the acquisition of inoculum from an ecologically relevant source was not shown or the success rate of the transmission was not determined [43].
Propagules of F. proliferatum and to a lesser extent other Fusarium species survived passage through the digestion tract of the insects. Survival of a gut passage is a key feature of entomopathogenic fungi [44]. In contrast to entomopathogenic fungus B. bassiana, F. proliferatum has not killed beetles that ingested fungal cultures. Low concentrations of beauvericin found in beetles fed on F. proliferatum indicate that the fungus ceased producing the mycotoxin in insect bodies, which may be part of its adaptation to dissemination by the beetles. Long survival of a plant pathogenic fungus in the digestion track of pests points out at a potentially important route of disease spread, establishing a new link between storage pest management and the control of fungal diseases.
Our results showed that F. proliferatum can be transmitted by mating of T. molitor to the eggs, which may lead to dissemination of the fungus by the new generation of the pest. Transmission of fungi to next generations by mating has been described for classical entomopathogens Metarhizium anisopliae and Beauveria bassiana [45]. Thus survival in the digestion track of insects and transmission to a next generation via copulation are features shared by F. proliferatum and entomopathogenic fungi. Apart from insect hosts to which it is pathogenic [41], F. proliferatum appears to be adapted to insects to which it does not cause visible disease. The adaptation of F. proliferatum to T. molitor may not allow appreciable proliferation of the fungus within insect host (Table 2) but it certainly facilitates its dissemination.
While preferentially feeding on grains infected with Fusarium spp., T. molitor is likely to ingest mycotoxins. Furthermore, fungus surviving in the digestive track of the insect might continue producing mycotoxins. Contamination of T. molitor with mycotoxins is relevant for food safety because the insect belongs to alternative proteins sources for food [46] and for life support systems for astronauts [47]. Surprisingly, mycotoxin fumonisin B1 was not detectable in beetles fed on F. proliferatum. The synthesis of fumonisin in F. verticillioides, which is closely related to F. proliferatum, underlies a complex control with epigenetic regulation [48]. We assume that the amount of fumonisin ingested with fungal mycelia was too small to be detected and that the synthesis of fumonisins was down-regulated in the insects. Alternatively, insects may have detoxified fumonisin, as indicated by the results of feeding and injecting mealworms with pure fumonisin B1 [49].
In conclusion, our study has shown that beetles of T. molitor are attracted to wheat grains infected with F. proliferatum and serve as a vehicle for the transmission of the fungus to uninfected grains. F. proliferatum survived passage via the digestive tract of insects and was transmitted to eggs by copulation, which suggests that F. proliferatum is adapted to dissemination by insect hosts.
Supporting information
(ZIP)
(ZIP)
Acknowledgments
We wish to express our gratitude to Dr. Ulrike Steiner for her guidance and comments on the scanning electronic microscopy. We thank Kerstin Lange, Heike Rollwage and Ruth Pilot for assistance and help in the lab. We also acknowledge anonymous reviewers for very helpful comments on the first version of this manuscript.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This research was partly supported by German Research Foundation (DFG, AL902/4-1 to B.A. www.dfg.de), the PLANTFOODSEC project of the European Union (FP7/2007-2013, No. 261752, https://cordis.europa.eu/project/rcn/100052_en.html) and China Postdoctoral Science Foundation (No. 2017M622304 support to ZG, www.chinapostdoctor.org.cn). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Jimenez M, Logrieco A, Bottalico A. Occurrence and pathogenicity of Fusarium species in banana fruits. Journal of Phytopathology. 1993; 137(3):214–20. 10.1111/j.1439-0434.1993.tb01341.x [DOI] [Google Scholar]
- 2.Hyun JW, Lee SC, Kim DH, Ko SW, Kim KS. Fusarium fruit rot of citrus in Jeju Island. Mycobiology. 2000; 28(3):158–62. [Google Scholar]
- 3.Abdalla MY, Al-Rokibah A, Moretti A, Mule G. Pathogenicity of toxigenic Fusarium proliferatum from date palm in Saudi Arabia. Plant Disease. 2000; 84(3):321–4. 10.1094/PDIS.2000.84.3.321 [DOI] [PubMed] [Google Scholar]
- 4.Ocamb CM, Juzwik J, Martin FB. Fusarium spp. and Pinus strobus seedlings: root disease pathogens and taxa associated with seed. New forests. 2002; 24(1):67–79. 10.1023/A:1020589121134 [DOI] [Google Scholar]
- 5.Guo Z, Pfohl K, Karlovsky P, Dehne H-W, Altincicek B. Fumonisin B-1 and beauvericin accumulation in wheat kernels after seed-borne infection with Fusarium proliferatum. Agricultural and Food Science. 2016; 25(2):138–45. http://orcid.org/0000-0001-8913-8900 [Google Scholar]
- 6.Clear RM, Patrick SK. Fusarium species isolated from wheat samples containing tombstone (scab) kernels from Ontario, Manitoba, and Saskatchewan. Canadian Journal of Plant Science. 1990; 70(4):1057–69. 10.4141/cjps90-128. [DOI] [Google Scholar]
- 7.Conner RL, Hwang SF, Stevens RR. Fusarium proliferatum: A new causal agent of black point in wheat. Can J Plant Pathol-Rev Can Phytopathol. 1996; 18(4):419–23. 10.1080/07060669609500598 [DOI] [Google Scholar]
- 8.Desjardins AE, Busman M, Proctor RH, Stessman R. Wheat kernel black point and fumonisin contamination by Fusarium proliferatum. Food additives and contaminants. 2007; 24(10):1131–7. 10.1080/02652030701513834 [DOI] [PubMed] [Google Scholar]
- 9.Amato B, Pfohl K, Tonti S, Nipoti P, Dastjerdi R, Pisi A, et al. Fusarium proliferatum and fumonisin B1 co-occur with Fusarium species causing Fusarium Head Blight in durum wheat in Italy. Journal of Applied Botany and Food Quality. 2015; 88:228–33. 10.5073/JABFQ.2015.088.033 [DOI] [Google Scholar]
- 10.Jurado M, Marín P, Callejas C, Moretti A, Vázquez C, González-Jaén MT. Genetic variability and fumonisin production by Fusarium proliferatum. Food microbiology. 2010; 27(1):50–7. 10.1016/j.fm.2009.08.001 [DOI] [PubMed] [Google Scholar]
- 11.Joint FAO/WHO Expert Committee on Food Additives. Safety evaluation of certain mycotoxins in food. 2001; Rome.
- 12.Wilson TM, Ross PF, Rice LG, Osweiler GD, Nelson HA, Owens DL, et al. Fumonisin B1 levels associated with an epizootic of equine leukoencephalomalacia. Journal of Veterinary Diagnostic Investigation. 1990; 2(3):213–6. 10.1177/104063879000200311 [DOI] [PubMed] [Google Scholar]
- 13.Osweiler GD, Ross PF, Wilson TM, Nelson PE, Witte ST, Carson TL, et al. Characterization of an epizootic of pulmonary edema in swine associated with fumonisin in corn screenings. Journal of Veterinary Diagnostic Investigation. 1992; 4(1):53–9. 10.1177/104063879200400112 [DOI] [PubMed] [Google Scholar]
- 14.Moretti A, Logrieco A, Bottalico A, Ritieni A, Randazzo G. Production of beauvericin by Fusarium proliferatum from maize in Italy. Mycotoxin Research. 1994; 10(2):73–8. 10.1007/BF03192255 [DOI] [PubMed] [Google Scholar]
- 15.Ritieni A, Fogliano V, Randazzo G, Scarallo A, Logrieco A, Moretti A, et al. Isolation and characterization of fusaproliferin, a new toxic metabolite from Fusarium proliferatum. Natural toxins. 1995; 3(1):17–20. 10.1002/nt.2620030105 [DOI] [PubMed] [Google Scholar]
- 16.Miller JD, Savard ME, Schaafsma AW, Seifert KA, Reid LM. Mycotoxin production by Fusarium moniliforme and Fusarium proliferatum from Ontario and occurrence of fumonisin in the 1993 corn crop. Canadian Journal of Plant Pathology. 1995; 17:233–9. [Google Scholar]
- 17.Marasas WFO, Thiel PG, Rabie CJ, Nelson PE, Toussoun TA. Moniliformin production in Fusarium section Liseola. Mycologia. 1986: 242–7. 10.2307/3793169 [DOI] [Google Scholar]
- 18.Eigenbrode SD, Bosque-Perez NA, Davis TS. Insect-borne plant pathogens and their vectors: ecology, evolution, and complex interactions In: Berenbaum MR, editor. Annual Review of Entomology, Vol 63 Annual Review of Entomology. 63. Palo Alto: Annual Reviews; 2018. p. 169–91. 10.1146/annurev-ento-020117-043119 [DOI] [PubMed] [Google Scholar]
- 19.Fermaud M, Menn R Le. Transmission of Botrytis cinerea to grapes by grape berry moth larvae. Phytopathology. 1992; 82(12):1393–8. [Google Scholar]
- 20.Paine TD, Raffa KF, Harrington TC. Interactions among scolytid bark beetles, their associated fungi, and live host conifers. Annual review of entomology. 1997; 42(1):179–206. 10.1146/annurev.ento.42.1.179 [DOI] [PubMed] [Google Scholar]
- 21.Prom LK, Lopez JD Jr, Latheef MA. Transmission of Claviceps africana spores from diseased to non-infected sorghum by corn earworm moths, Helicoverpa zea. Journal of Sustainable Agriculture. 2003; 21(4):49–58. 10.1300/J064v21n04_05 PMID: 15279250 [DOI] [Google Scholar]
- 22.Roy BA. Floral mimicry by a plant pathogen. Nature 1993; 362:56–8. [Google Scholar]
- 23.Friedli J, Bacher S. Mutualistic interaction between a weevil and a rust fungus, two parasites of the weed Cirsium arvense. Oecologia. 2001; 129(4):571–6. 10.1007/s004420100763 [DOI] [PubMed] [Google Scholar]
- 24.Guo Z, Döll K, Dastjerdi R, Karlovsky P, Dehne H-W, Altincicek B. Effect of fungal colonization of wheat grains with Fusarium spp. on food choice, weight gain and mortality of meal beetle larvae (Tenebrio molitor). PLOS ONE. 2014; 9(6):e100112 10.1371/journal.pone.0100112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Davis GR, Smith JD, Schiefer B, Loew FM. Screening for mycotoxins with larvae of Tenebrio molitor. Journal of invertebrate pathology. 1975; 26(3):299–303. 10.1016/0022-2011(75)90225-6 [DOI] [PubMed] [Google Scholar]
- 26.Leslie JF, Summerell BA. The Fusarium laboratory manual: Blackwell publishing; 2006. 10.1002/9780470278376 [DOI] [Google Scholar]
- 27.Abildgren MP, Lund F, Thrane U, Elmholt S. Czapek-Dox agar containing iprodione and dicloran as a selective medium for the isolation of Fusarium species. Letters in Applied Microbiology. 1987; 5(4):83–6. 10.1111/j.1472-765X.1987.tb01620.x [DOI] [Google Scholar]
- 28.Thrane U. Comparison of three selective media for detecting Fusarium species in foods: a collaborative study. International journal of food microbiology. 1996; 29(2):149–56. 10.1016/0168-1605(95)00040-2 [DOI] [PubMed] [Google Scholar]
- 29.Nutz S, Döll K, Karlovsky P. Determination of the LOQ in real-time PCR by receiver operating characteristic curve analysis: application to qPCR assays for Fusarium verticillioides and F. proliferatum. Analytical and bioanalytical chemistry. 2011; 401(2):717–26. 10.1007/s00216-011-5089-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Adejumo TO, Hettwer U, Karlovsky P. Survey of maize from south-western Nigeria for zearalenone, α- and β-zearalenols, fumonisin B1 and enniatins produced by Fusarium species. Food additives and contaminants. 2007; 24(9):993–1000. 10.1080/02652030701317285 [DOI] [PubMed] [Google Scholar]
- 31.Adejumo TO, Hettwer U, Karlovsky P. Occurrence of Fusarium species and trichothecenes in Nigerian maize. International journal of food microbiology. 2007; 116(3):350–7. 10.1016/j.ijfoodmicro.2007.02.009 [DOI] [PubMed] [Google Scholar]
- 32.Brandfass C, Karlovsky P. Upscaled CTAB-based DNA extraction and real-time PCR assays for Fusarium culmorum and F. graminearum DNA in plant material with reduced sampling error. International journal of molecular sciences. 2008; 9(11):2306–21. 10.3390/ijms9112306 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual Cold Spring Harbor, NY,: Cold Spring Harbor Laboratory Press; 1989. No.Ed. 2 pp. xxxviii + 1546 pp. [Google Scholar]
- 34.Waalwijk C, van der Heide R, de Vries I, van der Lee T, Schoen C, Costrel-de Corainville G, et al. Quantitative detection of Fusarium species in wheat using TaqMan Molecular Diversity and PCR-detection of Toxigenic Fusarium Species and Ochratoxigenic Fungi: Springer; 2004. p. 481–94. 10.1007/978-1-4020-2285-2_3 [DOI] [Google Scholar]
- 35.Parry DW, Nicholson P. Development of a PCR assay to detect Fusarium poae in wheat. Plant pathology. 1996; 45(2):383–91. 10.1046/j.1365-3059.1996.d01-133.x [DOI] [Google Scholar]
- 36.Mulè G, Susca A, Stea G, Moretti A. A species-specific PCR assay based on the calmodulin partial gene for identification of Fusarium verticillioides, F. proliferatum and F. subglutinans. European Journal of Plant Pathology. 2004; 110(5–6):495–502. 10.1023/B:EJPP.0000032389.84048.71 [DOI] [Google Scholar]
- 37.Waskiewicz A, Golinski P, Karolewski Z, Irzykowska L, Bocianowski J, Kostecki M, et al. Formation of fumonisins and other secondary metabolites by Fusarium oxysporum and F. proliferatum: a comparative study. Food additives and contaminants. 2010; 27(5):608–15. 10.1080/19440040903551947 [DOI] [PubMed] [Google Scholar]
- 38.R Development Core Team (2018). R: A language and environment for statistical computing. ISBN 3–900051–07–0. Available: http://www.R-project.org. R Foundation for Statistical Computing, Vienna, Austria.
- 39.Dunkel FV. The relationship of insects to the deterioration of stored grain by fungi. International journal of food microbiology. 1988; 7(3):227–44. 10.1016/0168-1605(88)90042-6 [DOI] [PubMed] [Google Scholar]
- 40.Chehri K. Molecular identification of entomopathogenic Fusarium species associated with Tribolium species in stored grains. J Invertebr Pathol. 2017;144: 1–6. 10.1016/j.jip.2017.01.003 [DOI] [PubMed] [Google Scholar]
- 41.Laith AA. First record of use Fusarium proliferatum fungi in direct treatment to control the adult of wheat flour beetle Tribolium confusum, as well as, use the entomopathogenic fungi Beauveria bassiana. Ecology, Environment and Conservation, in press. [Google Scholar]
- 42.Gordon WL. The occurrence of Fusarium species in Canada: VI. taxonomy and geographic distribution of Fusarium species on plants, insects, and fungi. Canadian Journal of Botany. 1959; 37(2):257–90. 10.1139/b59-021 PMID: 27055573 [DOI] [Google Scholar]
- 43.Drakulic J, Bruce TJA, Ray RV. Direct and host-mediated interactions between Fusarium pathogens and herbivorous arthropods in cereals. Plant Pathology 2016; 66:3–13. 10.1111/ppa.12546 [DOI] [Google Scholar]
- 44.Teetor-Barsch GH, Roberts DW. Entomogenous Fusarium species. Mycopathologia. 1983; 84(1):3–16. 10.1007/BF00436991 [DOI] [PubMed] [Google Scholar]
- 45.Maniania NK, Ouna E, Ahuya P, Frérot B, Félix AE, Le Ru B, et al. Dissemination of entomopathogenic fungi using Busseola fusca male as vector. Biological Control. 2011; 58(3):374–8. [Google Scholar]
- 46.Grau T, Vilcinskas A, Joop G. Sustainable farming of the mealworm Tenebrio molitor for the production of food and feed. Z Naturforsch, C, J Biosci. 2017;72: 337–349. 10.1515/znc-2017-0033 [DOI] [PubMed] [Google Scholar]
- 47.Li L, Zhao Z, Liu H. Feasibility of feeding yellow mealworm (Tenebrio molitor L.) in bioregenerative life support systems as a source of animal protein for humans. Acta Astronautica. 2013;92: 103–109. 10.1016/j.actaastro.2012.03.012 [DOI] [Google Scholar]
- 48.Visentin I, Montis V, Döll K, Alabouvette C, Tamietti G, Karlovsky P, et al. Transcription of genes in the biosynthetic pathway for fumonisin mycotoxins is epigenetically and differentially regulated in the fungal maize pathogen Fusarium verticillioides. Eukaryotic Cell. 2012;11: 252–259. 10.1128/EC.05159-11 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Abado‐Becognee K, Fleurat‐Lessard F, Creppy EE, Melcion D. Effects of fumonisin B1 on growth and metabolism of larvae of the yellow mealworm, Tenebrio molitor. Entomologia experimentalis et applicata. 1998; 86(2):135–43. 10.1046/j.1570-7458.1998.00274.x [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(ZIP)
(ZIP)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.