

Abstract
Myocardial infarction triggers massive biochemical changes, even cardiac cell death. Endoplasmic reticulum stress is involved in the pathology of myocardial infarction-mediated apoptosis. In the present study, myocardial cell line H9c2 cells were treated with cobalt chloride (CoCl2) to induce hypoxia. Isoproterenol was used for two successive days to induce myocardial infarction in SD rats. The cardioprotective effect of olive leaf extract (OLE) and its main constituent hydroxytyrosol and the underlying mechanisms were evaluated. The results showed that hydroxytyrosol markedly protected H9c2 cells against CoCl2-induced apoptosis. Hydroxytyrosol could reduce the mRNA and protein expression of GRP78 and CHOP induced by CoCl2 in vitro. In vivo, the decreased ejection fraction and fractional shortening, increased heart weight/body ratio, the formation of infarction, disordered cardiac muscle fibers and infiltration of inflammatory cells induced by isoproterenol could be significantly ameliorated by pretreatment with OLE for a month. Similarly, OLE could also reverse the increase of GRP78 and CHOP expression induced by isoproterenol. Therefore, OLE and hydroxytyrosol exert a cardioprotective effect through endoplasmic reticulum stress, which could be a new target for the prevention and treatment of cardiovascular diseases.
Keywords: myocardial infarction, olive leaf extract, hydroxytyrosol, endoplasmic reticulum stress
Introduction
Myocardial infarction (MI) leads to heart failure and subsequent death[1]. It is still a main cause of death in the world despite advances in treatments[2–3]. Studies have shown that endoplasmic reticulum (ER) stress in cardiomyocytes can generate and promote cell apoptosis[4]. The C/EBP homologous protein (CHOP) pathway is the major signal pathway through which ER stress induces apoptosis. GRP78, one kind of heat shock protein 70, is the main protein in the CHOP pathway[5]. The expression of heat shock proteins is an essential way to antagonize this stress[6]. Studies have shown that GRP78 responds to hypoxia and ischemia effectively[7]. CHOP and GRP78 have been widely used as specific markers for ER stress[8].
Olive leaf extract (OLE) contains abundant polyphenols including oleuropein, hydroxytyrosol and other flavonoids that have been investigated to fight against cardiovascular diseases[9–10]. Besides, Mediterranean diet with plentiful olive substances has been proven associated with a low incidence of cardiovascular diseases, cancer and stroke[11–13]. Hydroxytyrosol, one major component in OLE polyphenols, has been demonstrated to protect cardiomyocytes H9c2 against 4-hydroxynonenal induced cytotoxicity[14]. However, the mechanisms of hydroxytyrosol against hypoxia and MI are obscure.
MI is an imbalance between coronary blood supply and myocardial blood demand. In this study, cobalt chloride (CoCl2) was used as a hypoxia-inducing agent to mimic MI. Isoproterenol is a synthetic catecholamine and β-adrenergic agonist to induce MI. At a high concentration, isoprenaline produces severe stress, leading to infarct or infarct-like lesion of the myocardium[15–16]. In this study, isoprenaline was used to induce MI by subcutaneous injection for two successive days in vivo. We investigated the protective effect of OLE on MI induced by isoprenaline in vivo, and the protective effect of hydroxytyrosol on cardiomyocyte H9c2 induced by CoCl2 in vitro through the ER stress pathway.
Materials and methods
Reagents
CoCl2·6H2O was purchased from Sigma Chemical Co (USA). Isoproterenol Yi Feixue Bio, Nanjing, China. Hydroxytyrosol (98%) was purchased from Aladdin Biological Reagent Company (Shanghai, China). OLE (25%) was provided by Sinolife United Biotech Company. H9c2 cells were obtained from the Cell Bank of Shanghai Institute of Biochemistry and Cell Biology. Dulbecco's Modified Eagle's Medium (DMEM) and FBS were purchased from ScienCell Research Laboratories (CA, USA). Primary antibodies against GRP78, CHOP and β-actin were purchased from Cell Signaling Technology (Danvers, MA, USA). Goat anti-rabbit IgG and goat anti-mouse IgG antibodies were purchased from ZSGB-BIO (Beijing, China). Hochest 33258 was obtained from the Beyotime Institute of Biotechnology (Shanghai, China). Primers used for quantitative RT-PCR were provided by Invitrogen (Carlsbad, Calif, USA). GRP78:(Forward-5'-TCAGCCCACCGTAACAATCAAGG-3'; Reverse-5'-CTTCCTCAGCAAACTTCTCGGCG-3'). CHOP: (Forward-5'-GCACCTCCCAAAGCCCTCGC-3'; Reverse-5'-CCGTTTCCTAGTTCTTCCTT-3')
Cell culture and treatment
H9c2 cells were cultured in high-glucose DMEM supplemented with 10% FBS containing 5% CO2 at 37 °C. In order to induce cell hypoxia, 1×104 cells/well were seeded in 96-well plates and cultured overnight. The cells were treated with CoCl2 at different concentrations (0, 50, 100, 200, 400, 600, 800 and 1,000 μmol/L) for 24 hours. Then, cell viability was measured by MTT assay. For the protective effects of hydroxytyrosol, hydroxytyrosol (0, 1, 5, 10, 20, or 40 μmol/L) was used for pretreating H9c2 cells for 24 hours before CoCl2 (400 μmol/L) inducement.
Then, 5 × 105 H9c2 cells/well were seeded in a six-well plate with a glass coverslip in each well. The cells were treated with CoCl2 (400 μmol/L), hydroxytyrosol (40 μmol/L) and CoCl2 (400 μmol/L) plus hydroxytyrosol (40 μmol/L) for 24 hours. Cells with fragmented and condensed nuclei were determined using Hoechst 33258 staining (Beyotime) and annexin-V FITC/PI kit (KeyGEN, Nanjing, China) by flow cytometry according to the introductions.
Animals and experimental protocols
All animal experiments were conducted in accordance with the Institutional Animal Care and Use Committee of the Jiangsu Province Institute of Traditional Chinese Medicine and written up following the ARRIVE guidelines. Male SD rats of (200±20) g were supplied by Shanghai Silaike Laboratory Animal Ltd and allowed to adapt to the laboratory conditions for one week before experiment. Rats were divided randomly into 4 groups (n=6). The normal control group was pretreated with physiological saline by oral gavage one month before subcutaneous injection of physiological saline for two days. The isoproterenol group was pretreated with ddH2O by oral gavage one month before subcutaneous injection of isoproterenol [85 mg/(kg·day)] for two days. The isoproterenol plus OLE group was pretreated with OLE [200 mg/(kg·day)] by oral gavage one month before subcutaneous injection of isoproterenol for two days, and the OLE [200 mg/(kg·day)] group was treated with OLE by oral gavage for one month before subcutaneous injection of physiological saline for two days.
Western blotting
Cells were seeded in a six-well plate (5 × 105 H9c2 cells/well) cultured in high-glucose DMEM supplemented with 10% FBS and then treated with CoCl2 or HT for indicated doses and time. Proteins were isolated by lysis buffer (Beyotime).
After the animals were sacrificed, the heart tissues were disrupted by homogenization on ice with lysis buffer. After centrifugation, protein extracts were collected.
Protein lysates were separated on 10% SDS-PAGE, transferred onto the PVDF membranes, and blocked with 5% non-fat milk for one hour, and then incubated with primary antibodies overnight at 4 °C and secondary antibodies for one hour at room temperature. Membranes were again washed with TBST and immunoreactive proteins were visualized using ECL Western blotting detection reagents (Cell Signaling Technology) were used to detect immunoreactive proteins[17].
Quantitative RT-PCR
Total RNA was extracted from H9c2 cells or heart tissues using RNAiso Plus (TaKaRaBio Technology, Dalian, China). Then, RNA was converted to cDNA using Prime Script TM RT Master Mix (TaKaRa). Real-time qPCR analysis for mRNA expression was performed using SYBR Green qPCR Master Mix (Yi FeiXue Bio, Nanjing, China) and ABI 7900. mRNA expression was normalized against GAPDH expression[18].
Echocardiography
After anesthetizing with 10% chloral hydrate, the rats were placed in decubitus supine position on the heat pad. The transthoracic echocardiography was carried out using GE ViVid-q ultrasound systems with 3-MHz linear transducer and 2-cm depth two-dimensional imaging (GE Systems, Hayozma, Tirat Carmel, Israel) when prewarmed echo transmission gel was applied to the hairless chest after injection of ISO for two days.
Statistical analysis
Statistical analysis was performed using SPSS 18.0 and GraphPad Prism v5.0 (Graphpad Software Inc) software. Data were presented as mean±SD. Unpaired Student's t tests was used to compare the means of two groups, and one-way analysis of variance (ANOVA) was used to compare the means of at least three groups. P<0.05 was considered to be statistically significant.
Results
Protective role of hydroxytyrosol against CoCl2 treatment
H9c2 cells were treated with CoCl2 at different concentrations (0–1,000 μmol/L) for 24 hours to induce hypoxia. As shown in Fig. 1A, the cell viability revealed a dose-dependent decrease. When the concentration reached 200 μmol/L, CoCl2 significantly decreased cell viability. Because of the cell viability of 400 μmol/L CoCl2 treatment was about 50%, this concentration of CoCl2 was used to induce cell hypoxia in subsequent experiments. When the cells were pretreated with hydroxytyrosol (Fig. 1B) for 24 hours and then treated with CoCl2 for 24 hours, cell viability recovered in a concentration-dependent way. At 40 μmol/L hydroxytyrosol, significantly increased cell viability was observed, compared with CoCl2 treatment.
Fig.1.
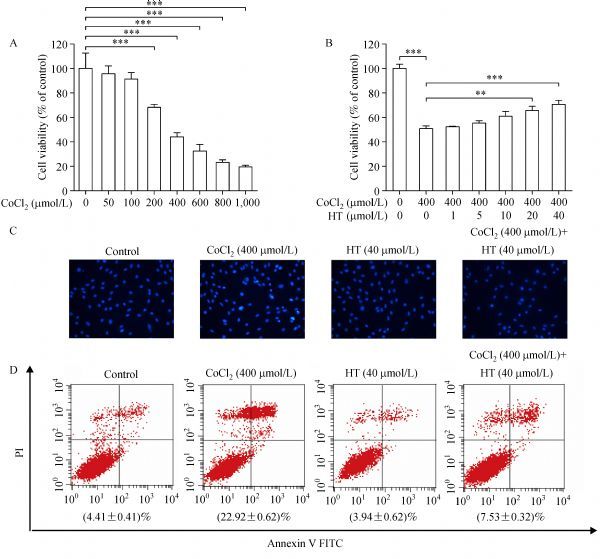
Hydroxytyrosol (HT) protects against CoCl2 induced apoptosis.
To confirm the results, Hochest 33258 staining was used to detect apoptotic cell death. H9c2 cells in the control group showed weak fluorescent spots, whereas the cells exposed to 400 μmol/L CoCl2 for 24 hours showed strong fluorescent spots; furthermore, pretreatment of hydroxytyrosol reversed the apoptosis of H9c2 cells induced by CoCl2. These indicate the protective role of HT against the damage effect of CoCl2 (Fig. 1C). In addition, to further confirm this effect, Annexin-V FITC/propidium iodide staining with flow cytometry was performed, which also showed that hydroxytyrosol could alleviate H9c2 apoptosis induced by CoCl2 (Fig. 1D).
Expression of GRP78 and CHOP after CoCl2 treatment
The induction of hypoxic stress was in parallel with induction of GRP78 and CHOP[19]. Western blotting analysis showed that 400 μmol/L CoCl2 markedly increased the expression of GRP78 time-dependently at 0-12 hours. At 24 hours, the expression of GRP78 slightly decreased (Fig. 2A). The expression of CHOP showed a time-dependent increase at 0–24 hours. When the cells were treated with CoCl2 at various concentrations for 12 hours, the expression of GRP78 peaked at 200 µmol/L and then decreased at 400 µmol/L. Besides, the expression of CHOP was increased significantly in a dose-dependent manner (Fig. 2B). As expected, the expression of GRP78 and CHOP in mRNA were consistent with change of protein at different time points and concentrations (Fig. 2C, D, E and F).
Fig.2.
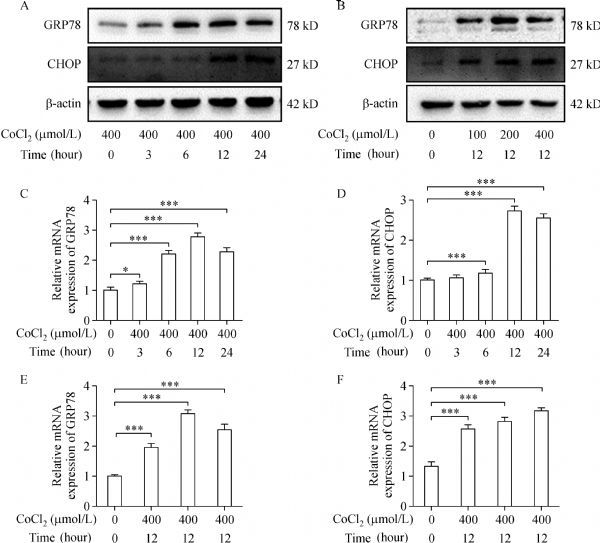
The expression of GRP78 and CHOP after CoCl2 treatment.
Hydroxytyrosol inhibited the expression of GRP78 and CHOP after CoCl2 treatment
According to the above results, HT could protect H9c2 cells against the damage of CoCl2. CoCl2 treatment could increase the expression of GRP78 and CHOP, which might be a result of hypoxic stress. As shown in Fig. 3A, H9c2 cells were pretreated with hydroxytyrosol at the indicated concentrations for 24 hours, and then treated with CoCl2 (400 µmol/L) for 12 hours. The expression of GRP78 and CHOP was inhibited dramatically by hydroxytyrosol, as compared to the treatment of CoCl2. In order to confirm the results, the mRNA levels of GRP78 and CHOP were detected. Fig. 3B–C shows that the mRNA levels of GRP78 and CHOP were upregulated. However, hydroxytyrosol at 10–40 μmol/L could reverse GRP78 levels significantly, and CHOP levels at 1–40 μmol/L. Therefore, the protective effects of hydroxytyrosol against the damage of CoCl2 might result from the inhibition of GRP78 and CHOP expression.
Fig.3.
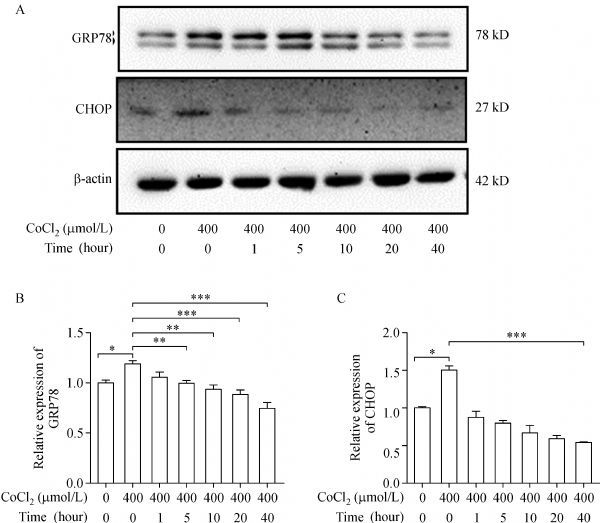
Hydroxytyrosol inhibits the expression of GRP78 and CHOP after CoCl2 treatment.
Protective effects of OLE against isoproterenol-induced MI in rats
According to the in vitro results, we evaluated the protective effects of OLE against isoproterenol-induced MI in vivo. OLE [200 mg/(kg·day)] was administered by gavage for one month before subcutaneous injection of isoproterenol [85 mg/(kg·day)] for two days. H&E staining displayed that rats receiving OLE treatment had normal cardiac fibers and no inflammatory cell infiltration. Rats receiving isoproterenol showed infarction areas, characterized by disordered cardiac muscle fibers and infiltration of inflammatory cells. However, after the pretreatment with OLE, these pathological changes were dramatically ameliorated (Fig. 4A).
Fig.4.
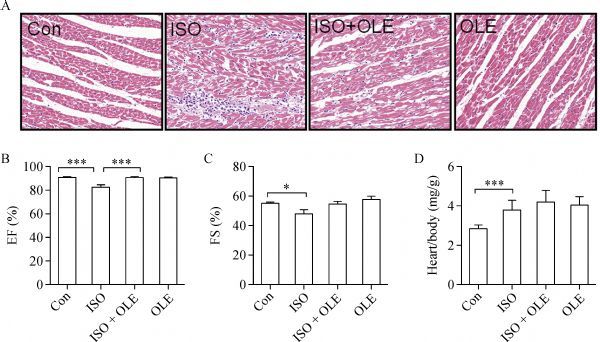
OLE exerts cardioprotection on rats subjected to isoproterenol .
As shown in Fig. 4B–C, echocardiography showed that heart function in isoproterenol-treated rats was significantly decreased in ejection fraction and fraction shortening. However, OLE treatment considerably improved heart function with increased ejection fraction and fraction shortening. The heart weight over body weight ratio was higher in the isoproterenol group; however, this increase could be reversed by OLE (Fig. 4D).
Protective effect of OLE on isoproterenol-induced MI through upregulating GRP78 and CHOP in rats
Expression levels of GRP78 and CHOP in vivo were also detected. As expected, proteins extracted from hearts tissues treated with OLE [200 mg/(kg·day)] showed a low expression of GRP78 and CHOP, which was similar to that of the control group. However, isoproterenol [85 mg/(kg·day)] could enhance the expression of GRP78 and CHOP significantly. When treated with isoproterenol and OLE, the expression of these two proteins was reversed dramatically (Fig. 5A).
Fig.5.
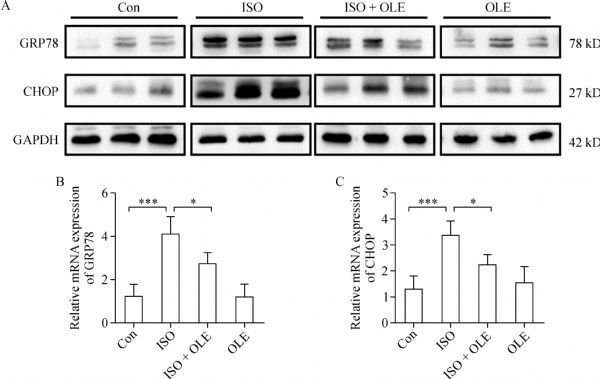
Effects of OLE on protein and mRNA levels of GRP78 and CHOP in rats.
The mRNA levels of GRP78 and CHOP in heart tissues were also detected by quantitative RT-PCR. The expression levels of CHOP and GRP78 were higher in the group treated with isoproterenol [85 mg/(kg·day)] than that in control and OLE [200 mg/(kg·day)] treatment groups. However, combination of isoproterenol with OLE [200 mg/(kg·day)] could reverse the high expression of CHOP and GRP78 induced by isoproterenol [85 mg/(kg·day)] (Fig. 5A and B). Similar results were observed in the protein expression. These results demonstrated that the protection of OLE against isoproterenol-induced MI in vivo involved the inhibition of GRP78 and CHOP expression. As shown in Fig. 6A, OLE and the main component, hydroxytyrosol can protect MI induced by isoproterenol in vivo and H9c2 cells hypoxia induced by CoCl2 in vitro.
Fig.6.
GRP78 and CHOP are involved in cardioprotective effect of hydroxytyrosol and OLE A schematic summary for the protection and mechanisms of hydroxytyrosol and olive leaf extract on myocardial infarction in vitro and in vivo.
Discussion
This study elaborated that OLE can reverse isoproterenol-induced MI and its major component hydroxytyrosol can protect H9c2 cells against apoptosis induced by CoCl2 through the ER stress pathway in vivo and in vitro. To our knowledge, it is the first report that hydroxytyrosol attenuates CoCl2-induced hypoxia in H9c2 cells and OLE reverses MI through regulating molecules GRP78 and CHOP in the ER stress pathway.
CoCl2 is a well-known inducer of hypoxia in cultured cardiomyocytes. Also, isoproterenol is extensively used to induce acute MI in animal models that mimic myocardial ischemic-anoxic injury[20–24]. In our study, CoCl2 induces the apoptosis of H9c2 cells and reduces their viability in a dose-dependent manner, which is consistent with the other reports that CoCl2 induced hypoxia and apoptosis of H9c2 cells[25–26]. However, this is the first report of the antagonistic effect of hydroxytyrosol on CoCl2 showing hydroxytyrosol protected H9c2 cells against the cytotoxicity of CoCl2. Besides, xanthine/xanthine oxidase has been suggested as a risk factor for heart disease. Xanthine/xanthine oxidase induces cardiomyocyte apoptosis and death through generating superoxide and hydrogen peroxide[27]. In contrast, CoCl2 can replace ferrous ions of proline hydroxylase, which leads cells into a hypoxic state. Moreover, hydroxytyrosol maintains cardiomyocyte viability through inhibiting intracellular ROS generation and mediating the phosphorylation of survival or death related proteins in H9c2 cells subjected to xanthine/xanthine oxidase-induced oxidative stress[27]. In vivo, when the rats were treated with isoproterenol, MI was obviously induced as ejection fraction, fractional shortening, disarranged cardiac muscle fibers and inflammatory cell infiltration decreased, which is consistent with the finding of previous studies[28–29].
The ER is a crucial component of the cellular reticular network that allows cells to adjust to a variety of conditions[30], like protein folding, calcium homeostasis, and lipid biosynthesis. Stimuli such as oxidative stress and ischemic insults enhance the expression of normal and/or folding-defective proteins, leading to the accumulation of unfolded proteins. This phenomenon is called ER stress that activates unfolded protein reaction[31]. Three major branches of ER stress sensors are protein kinase ER-like kinase (PERK), inositol-requiring protein 1 (IRE1) and activating transcription factor 6 (ATF6), which are involved in unfolded protein reaction[32]. Under the normal condition, the ER stress sensors are bound to GRP78 and inactivated. But when stimulated, GRP78 is dissociated from ER sensors, ER sensors are activated, and unfolded protein reaction is initiated to reduce cellular dysfunction and promote cell survival. However, excessive unfolded protein reaction will initiate CHOP-mediating cell apoptosis[33]. Studies show that ER stress plays an important role in the pathogenesis of MI, ischemic heart diseases and heart failure[30, 34–35]. Suppressing ER stress and associated apoptosis provides protective effects on cardiovascular system[36]. In addition, ER stress induces the expression of CHOP and then inhibits the expression of anti-apoptotic proteins Bcl-2 and Bnip3[37–38]. In our study, we observed CoCl2 could induce the overexpression of CHOP and GRP78 in vitro and isoproterenol induced the similar results in vivo. Pretreated with hydroxytyrosol or OLE, the abnormal expression of CHOP and GRP78 was inhibited significantly.
The therapeutic effect of OLE and its main constituent hydroxytyrosol is notable. The formation of infarction, disordered cardiac muscle fibres and infiltration of inflammatory cells are the typical pathological characteristics of MI[39]. All of these pathologic changes could be ameliorated by OLE significantly.
In conclusion, this study demonstrates that OLE and its main constituent hydroxytyrosol can prevent H9c2 cell damage and MI induced by CoCl2 and isoproterenol. Defense ER stress is involved in this cardioprotective mechanism. The expression of GRP78 and CHOP, specific markers of ER stress, can be decreased significantly by OLE and hydroxytyrosol. Therefore, the OLE and hydroxytyrosol are effective and promising agents for treatment of MI. However, detailed molecular mechanisms of OLE and hydroxytyrosol still need to be studied in order to confirm these protective functions.
Acknowledgment
This work was supported by the National Natural Science Foundation of China (81472977), foundation from the Priority Academic Program Development of Jiangsu Higher Education Institutions, Graduate Student Practice and Innovation Project of Jiangsu Province Ordinary University (SJZZ15_0117).
References
- 1. Janahmadi Z, Nekooeian AA, Moaref AR, et al. Oleuropein offers cardioprotection in rats with acute myocardial infarction[J]. Cardiovasc Toxicol, 2015, 15(1): 61–68. . [DOI] [PubMed] [Google Scholar]
- 2. Jing L, Wang Y, Zhao XM, et al. Cardioprotective effect of hydrogen-rich saline on isoproterenol-induced myocardial infarction in rats[J]. Heart Lung Circ, 2015, 24(6): 602–610. . [DOI] [PubMed] [Google Scholar]
- 3. Aronow WS. Epidemiology, pathophysiology, prognosis, and treatment of systolic and diastolic heart failure[J]. Cardiol Rev, 2006, 14(3): 108–124. . [DOI] [PubMed] [Google Scholar]
- 4. Szegezdi E, Duffy A, O'Mahoney ME, et al. ER stress contributes to ischemia-induced cardiomyocyte apoptosis[J]. Biochem Biophys Res Commun, 2006, 349(4): 1406–1411. . [DOI] [PubMed] [Google Scholar]
- 5. Yu H, Zhang H, Zhao W, et al. Gypenoside protects against myocardial ischemia-reperfusion injury by inhibiting cardiomyocytes apoptosis via inhibition of CHOP pathway and activation of PI3K/Akt pathway in vivo and in vitro[J]. Cell Physiol Biochem, 2016, 39(1): 123–136. . [DOI] [PubMed] [Google Scholar]
- 6. Hardy B, Raiter A. Peptide-binding heat shock protein GRP78 protects cardiomyocytes from hypoxia-induced apoptosis[J]. J Mol Med (Berl), 2010, 88(11): 1157–1167. . [DOI] [PubMed] [Google Scholar]
- 7. Shintani-Ishida K, Nakajima M, Uemura K, et al. Ischemic preconditioning protects cardiomyocytes against ischemic injury by inducing GRP78[J]. Biochem Biophys Res Commun, 2006, 345(4): 1600–1605. . [DOI] [PubMed] [Google Scholar]
- 8. Schröder M, Kaufman RJ. The mammalian unfolded protein response[J]. Annu Rev Biochem, 2005, 74(1): 739–789. . [DOI] [PubMed] [Google Scholar]
- 9. Poudyal H, Campbell F, Brown L. Olive leaf extract attenuates cardiac, hepatic, and metabolic changes in high carbohydrate-, high fat-fed rats[J]. J Nutr, 2010, 140(5): 946–953. . [DOI] [PubMed] [Google Scholar]
- 10. El SN, Karakaya S. Olive tree (Olea europaea) leaves: potential beneficial effects on human health[J]. Nutr Rev, 2009, 67(11): 632–638. . [DOI] [PubMed] [Google Scholar]
- 11. Dekanski D, Selaković V, Piperski V, et al. Protective effect of olive leaf extract on hippocampal injury induced by transient global cerebral ischemia and reperfusion in Mongolian gerbils[J]. Phytomedicine, 2011, 18(13): 1137–1143. . [DOI] [PubMed] [Google Scholar]
- 12. Fung TT, Rexrode KM, Mantzoros CS, et al. Mediterranean diet and incidence of and mortality from coronary heart disease and stroke in women[J]. Circulation, 2009, 119(8): 1093– 1100. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Boss A, Bishop KS, Marlow G, et al. Evidence to support the anti-cancer effect of olive leaf extract and future directions[J]. Nutrients, 2016, 8(8): 8. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Bali EB, Ergin V, Rackova L, et al. Olive leaf extracts protect cardiomyocytes against 4-hydroxynonenal-induced toxicity in vitro: comparison with oleuropein, hydroxytyrosol, and quercetin[J]. Planta Med, 2014, 80(12): 984–992. . [DOI] [PubMed] [Google Scholar]
- 15. Othman AI, Elkomy MM, El-Missiry MA, et al. Epigallocatechin-3-gallate prevents cardiac apoptosis by modulating the intrinsic apoptotic pathway in isoproterenol-induced myocardial infarction[J]. Eur J Pharmacol, 2017, 794: 27–36. . [DOI] [PubMed] [Google Scholar]
- 16. Akila P, Vennila L. Chlorogenic acid a dietary polyphenol attenuates isoproterenol induced myocardial oxidative stress in rat myocardium: An in vivo study[J]. Biomed Pharmacother, 2016, 84: 208–214. . [DOI] [PubMed] [Google Scholar]
- 17. Zhou Q, Chen B, Wang X, et al. Sulforaphane protects against rotenone-induced neurotoxicity in vivo: Involvement of the mTOR, Nrf2, and autophagy pathways[J]. Sci Rep, 2016, 6: 32206. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Chen G, Yang Y, Hu C, et al. Protective effects of Huangqin Decoction against ulcerative colitis and associated cancer in mice[J]. Oncotarget, 2016, 7(38): 61643–61655. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Sawada N, Yao J, Hiramatsu N, et al. Involvement of hypoxia-triggered endoplasmic reticulum stress in outlet obstruction-induced apoptosis in the urinary bladder[J]. Lab Invest, 2008, 88(5): 553–563. . [DOI] [PubMed] [Google Scholar]
- 20. Baines CP, Molkentin JD. Adenine nucleotide translocase-1 induces cardiomyocyte death through upregulation of the pro-apoptotic protein Bax[J]. J Mol Cell Cardiol, 2009, 46(6): 969–977. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Xu ZW, Chen X, Jin XH, et al. SILAC-based proteomic analysis reveals that salidroside antagonizes cobalt chloride-induced hypoxic effects by restoring the tricarboxylic acid cycle in cardiomyocytes[J]. J Proteomics, 2016, 130: 211–220. . [DOI] [PubMed] [Google Scholar]
- 22. Gallo S, Gatti S, Sala V, et al. Agonist antibodies activating the Met receptor protect cardiomyoblasts from cobalt chloride-induced apoptosis and autophagy[J]. Cell Death Dis, 2014, 5: e1185. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Dhivya V, Priya LB, Chirayil HT, et al. Piperine modulates isoproterenol induced myocardial ischemia through antioxidant and anti-dyslipidemic effect in male Wistar rats[J]. Biomed Pharmacother, 2017, 87: 705–713. . [DOI] [PubMed] [Google Scholar]
- 24. Akila P, Asaikumar L, Vennila L. Chlorogenic acid ameliorates isoproterenol-induced myocardial injury in rats by stabilizing mitochondrial and lysosomal enzymes[J]. Biomed Pharmacother, 2017, 85: 582–591. . [DOI] [PubMed] [Google Scholar]
- 25. Ashok A, Kanwar JR, Krishnan UM, et al. SurR9C84A protects and recovers human cardiomyocytes from hypoxia induced apoptosis[J]. Exp Cell Res, 2017, 350(1): 19–31. . [DOI] [PubMed] [Google Scholar]
- 26. Sajjad A, Novoyatleva T, Vergarajauregui S, et al. Lysine methyltransferase Smyd2 suppresses p53-dependent cardiomyocyte apoptosis[J]. Biochim Biophys Acta, 2014, 1843(11): 2556–2562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Ozbek N, Bali EB, Karasu C. Quercetin and hydroxytyrosol attenuates xanthine/xanthine oxidase-induced toxicity in H9c2 cardiomyocytes by regulation of oxidative stress and stress-sensitive signaling pathways[J]. Gen Physiol Biophys , 2015, 34(4): 407–414. . [DOI] [PubMed] [Google Scholar]
- 28. Kumar M, Kasala ER, Bodduluru LN, et al. Baicalein protects isoproterenol induced myocardial ischemic injury in male Wistar rats by mitigating oxidative stress and inflammation[J]. Inflamm Res, 2016, 65(8): 613–622. . [DOI] [PubMed] [Google Scholar]
- 29. Kocak C, Kocak FE, Akcilar R, et al. Molecular and biochemical evidence on the protective effects of embelin and carnosic acid in isoproterenol-induced acute myocardial injury in rats[J]. Life Sci, 2016, 147: 15–23. . [DOI] [PubMed] [Google Scholar]
- 30. Groenendyk J, Agellon LB, Michalak M. Coping with endoplasmic reticulum stress in the cardiovascular system[J]. Annu Rev Physiol, 2013, 75: 49–67. . [DOI] [PubMed] [Google Scholar]
- 31. Minamino T, Komuro I, Kitakaze M. Endoplasmic reticulum stress as a therapeutic target in cardiovascular disease[J]. Circ Res, 2010, 107(9): 1071–1082. . [DOI] [PubMed] [Google Scholar]
- 32. Cubillos-Ruiz JR, Bettigole SE, Glimcher LH. Tumorigenic and immunosuppressive effects of endoplasmic reticulum stress in cancer[J]. Cell, 2017, 168(4): 692–706. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Walter P, Ron D. The unfolded protein response: from stress pathway to homeostatic regulation[J]. Science, 2011, 334(6059): 1081–1086. . [DOI] [PubMed] [Google Scholar]
- 34. Luo T, Chen B, Wang X. 4-PBA prevents pressure overload-induced myocardial hypertrophy and interstitial fibrosis by attenuating endoplasmic reticulum stress[J]. Chem Biol Interact, 2015, 242: 99–106. . [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Wu H, Ye M, Yang J, et al. Nicorandil protects the heart from ischemia/reperfusion injury by attenuating endoplasmic reticulum response-induced apoptosis through PI3K/Akt signaling pathway[J]. Cell Physiol Biochem , 2015, 35(6): 2320–2332. . [DOI] [PubMed] [Google Scholar]
- 36. Miyazaki Y, Kaikita K, Endo M, et al. C/EBP homologous protein deficiency attenuates myocardial reperfusion injury by inhibiting myocardial apoptosis and inflammation[J]. Arterioscler Thromb Vasc Biol, 2011, 31(5): 1124–1132. . [DOI] [PubMed] [Google Scholar]
- 37. Oyadomari S, Mori M. Roles of CHOP/GADD153 in endoplasmic reticulum stress[J]. Cell Death Differ, 2004, 11(4): 381–389. . [DOI] [PubMed] [Google Scholar]
- 38. Fang SJ, Li PY, Wang CM, et al. Inhibition of endoplasmic reticulum stress by neuregulin-1 protects against myocardial ischemia/reperfusion injury[J]. Peptides, 2017, 88: 196–207. . [DOI] [PubMed] [Google Scholar]
- 39. Griswold DE, Hillegass LM, Hill DE, et al. Method for quantification of myocardial infarction and inflammatory cell infiltration in rat cardiac tissue[J]. J Pharmacol Methods, 1988, 20(3): 225–235. . [DOI] [PubMed] [Google Scholar]