

Abstract
Aims
Myocarditis is often associated with parvovirus B19 (B19V) persistence, which can induce vascular damage. Based on the antiviral and anti‐inflammatory properties of telbivudine, we aimed to evaluate its efficacy to protect B19V‐infected endothelial cells in vitro and to treat chronic lymphocytic myocarditis patients with B19V transcriptional activity.
Methods and results
We evaluated the endothelial‐protective potential of telbivudine in human microvascular endothelial cells‐1, which were infected with B19V. Treatment with 10 ng/mL of telbivudine decreased the B19V‐induced endothelial cell apoptosis and endothelial‐to‐mesenchymal transition. Along with this finding, telbivudine reduced the expression of transforming growth factor‐β1 and of tenascin‐C. The endothelial‐protective properties of telbivudine were also found in tumour necrosis factor‐α‐stressed human microvascular endothelial cells‐1. In addition, oxidative stress in angiotensin II‐stressed and transforming growth factor‐β1‐stressed HL‐1 cardiomyocytes and fibroblasts, respectively, was reduced upon telbivudine treatment, illustrating that telbivudine exerts multimodal protective effects. Based on these in vitro findings, four patients severely suffering from an endomyocardial biopsy‐proven myocarditis associated with B19V transcriptional activity (VP1/VP2‐mRNA positive) were treated with telbivudine (600 mg/dL) for 6 months in a single‐patient‐use approach. Follow‐up biopsies 6 months after treatment showed that VP1/VP2‐mRNA levels and CD3 cells decreased in all patients and were associated with an improvement in ejection fraction and New York Heart Association class. These findings were paralleled by a drop in tenascin‐C expression as shown via matrix‐assisted laser desorption ionization–imaging mass spectrometry.
Conclusions
Telbivudine exerts endothelial‐protective effects in B19V‐infected endothelial cells and improves chronic myocarditis associated with B19V transcriptional activity. These findings will be further evaluated in the clinical exploratory trial: the PreTopic study.
Keywords: Myocarditis, Parvovirus B19, Telbivudine, Endothelial cells, Pleiotropic effects
Introduction
Parvovirus B19 (B19V) is a small non‐enveloped DNA virus of the Parvoviridae family that preferentially replicates in erythroid progenitor cells. Despite its strong erythroid tropism and related acute disease association, B19V persists in many other non‐erythroid tissues, including the liver, skin, brain, and heart.1 Thereby, it is a vasculotrope virus,2, 3 related to microangiopathy, endothelial,4 and diastolic dysfunction5 and elevated circulating endothelial microparticles,6 owing to its capacity to infect endothelial cells. Parvovirus B19 has often been linked to the pathogenesis of myocarditis and its progression to dilated cardiomyopathy (DCM).7 However, the etiological role of B19V in the development of myocarditis still remains unclear because cardiac B19V is also often found in myocardial autopsy samples of subjects without myocarditis and DCM.8, 9 Therefore, findings of cardiac B19V DNA could also belong to an unspecific innocent bystander effect rather than necessarily reflecting the pathogen causing myocarditis.10 At least systemic B19V infection can aggravate several clinical scenarios, including hepatitis,11 rheumatoid arthritis,12 post‐transplants,13 and glomerulonephritis.14
Myocarditis usually differs from these scenarios because it is rarely associated with a systemic B19V infection, despite findings of cardiac B19V DNA persistence.15 Only high copy numbers of B19V DNA are currently thought to be myocarditis related,3 but these are rarely found in endomyocardial biopsy (EMB). Recent findings further indicate that a subgroup of myocarditis patients characterized by transcriptionally active cardiotropic B19V have an altered cardiac gene expression compared with control patients and myocarditis patients with latent B19V, suggesting that probably active replicating B19V with detectable viral RNA is related to myocarditis.16 Besides the extent of the viral load17, 18 and the replicative status/transcriptional activity of B19V,16 other studies further suggest that the relevance of B19V in myocarditis and DCM depends on the presence of other cardiotropic viruses.17
The unclarity about the pathogenic role of B19V in myocarditis and DCM, partly due to the absence of experimental in vivo models, has hampered the search for potential B19V‐targeted therapies.19 Nevertheless, there is evidence from cell culture experiments showing that B19V modulates inflammatory signalling and apoptosis in endothelial cells20 and that its capsid protein VP1 damages circulating angiogenic cells,21 resulting in dysfunctional endogenous vascular repair.21 The same authors further demonstrated that B19V infection has a direct VP2‐mediated negative impact on trafficking of circulating angiogenic cells in the presence of impaired cardiac regeneration.22 In addition, although the virus does not infect the cardiomyocytes, it is discussed that myocarditis can arise from immunological cross‐reaction to epitopes shared between the virus and the myocardium,23 indicating the need for the development of not only antiviral but also immunomodulatory interventions in patients with B19V‐associated myocarditis.
On the search for antiviral components that possess immunomodulatory and/or antiviral properties meaningful in the context of lympohycytic myocarditis and B19V persistance, we found the mode of action of telbivudine interesting. Although this antiviral nucleoside analogue reverse transcriptase inhibitor is especially effective for retroviral and para‐retroviral (hepatitis B viruses) infections, telbivudine has some pleiotropic immunomodulatory/anti‐inflammatory24, 25, 26, 27 and interesting antiviral properties discussed to be theoretically able to interfere with the unique replication mode of B19V, too. In more detail, the finding that the single‐stranded B19V DNA genome replicates through a specific rolling hairpin mechanism to generate a double‐stranded DNA molecule mimicking DNA synthesis during the reverse transcription process comparable with retroviruses and hepatitis B viruses,28 the ability of telbivudine to preferentially inhibit the DNA‐dependent second strand DNA synthesis and its anti‐inflammatory effects,24, 25, 26, 27 prompted us to investigate whether treatment with telbivudine is effective in myocarditis associated with B19V transcriptional activity. Therefore, we investigated telbivudine in different in vitro settings to understand the possible mode of action and proved whether the results can be successfully translated to the clinical scenario by initiating a single‐patient‐use programme for patients severely suffering from EMB‐proven chronic lymphocytic myocarditis associated with cardiac B19V transcriptional activity.
Materials and methods
Cell culture
Human microvascular endothelial cells (HMEC)‐1 were cultured in endothelial cell growth (ECG) culture medium (PromoCell, Heidelberg, Germany) plus supplements (Supplement Mix for ECG medium; PromoCell) and 1% penicillin/streptomycin (P/S; Gibco, Paisley, UK) on T75 cm2 culture flasks (Falcon, NC, USA) pre‐coated with 0.02% gelatin and 125 mg/100 mL fibronectin solution (both from Sigma‐Aldrich, St. Louis, MO, USA) until experiments with B19V (see infra) or tumour necrosis factor (TNF)‐α. For TNF‐α supplementation experiments, HMEC‐1 were plated at a density of 225 000 cells/6‐well or 10 000 cells/96‐well for flow cytometry and Sirius Red/crystal violet staining, respectively. After 24 h, the medium was replaced by serum starvation medium [basal endothelial growth medium supplemented with 0.01% fetal bovine serum (FBS) and 1% P/S] with/out 10 ng/mL TNF‐α (BD Biosciences, Heidelberg, Germany) in the presence or absence of 10 ng/mL telbivudine (Santa Cruz, Heidelberg, Germany). Twenty‐four hours after culture, the cells were collected for Annexin V/7AAD flow cytometry or stained with Sirius Red and crystal violet.
UT7/EPO S1 cells (kindly provided by Dr Fechner, TU, Berlin) were cultured in Iscove's modified Dulbecco's medium (GE Healthcare Life Sciences HyClone Laboratories, Logan, UT, USA) supplemented with 10% FBS, 1% P/S, and 75 μL EPO (4000 i.E) in T75 flasks until B19V infection experiments (see infra). HL‐1 cardiomyoyctes were cultured in Claycomb media (Sigma‐Aldrich) supplemented with 10% FBS (Gibco Life Technologies), 1% P/S (Gibco), 0.1 mM norepinephrine (Sigma‐Aldrich), and 2 mM l‐glutamine (Biochrom, Berlin, Germany) on T75 culture flasks pre‐coated with 0.02% gelatin and 125 mg/100 mL fibronectin solution (both from Sigma‐Aldrich). For angiotensin (Ang) II stimulation experiments, HL‐1 cells were plated at a density of 200 000 cells/6‐well and 24 h later stimulated with/out 1 μM Ang II in the presence or absence of 10 ng/mL telbivudine (Santa Cruz). Twenty‐four hours after stimulation, cells were collected for reactive oxygen species (ROS) and sarcomeric myosin heavy chain flow cytometry. Murine fibroblasts were cultured in Iscove's modified Dulbecco's medium (Sigma‐Aldrich) supplemented with 10% FBS (Gibco Life Technologies) and 1% P/S (Gibco). For stimulation experiments with transforming growth factor‐β1 (TGF‐β1), murine fibroblasts were plated at a density of 100 000 cells/6‐well. After 24 h of culture, the medium was aspirated and replaced with serum starvation medium (0.05% FBS + 1% P/S) with/out 5 ng/mL TGF‐β1 in the presence or absence of 10 ng/mL telbivudine (Santa Cruz) for 24 h. Thereafter, cells were collected for ROS flow cytometry.
Parvovirus B19 infection
In order to infect HMEC‐1 with B19V, HMEC‐1 were plated at a cell density of 225 000 cells/6‐well on gelatin/fibronectin pre‐coated 6‐well plates. Twenty‐four hours later, the cells were infected with 10 000 B19V particles per cell in 500 μL supplement‐free medium containing 0.1% FBS, 1% P/S, and 0.5 mM glucose for 2 h at 4°C and 1 h at 37°C, as described previously.20 In parallel, cells without B19V infection were cultured under the same serum starvation conditions for 2 h at 4°C and 1 h at 37°C. Next, HMEC‐1 were washed twice with phosphate‐buffered saline (PBS) (Gibco) and subsequently cultured in full ECG medium with/out 10 ng/mL telbivudine (Santa Cruz) for 30 min for RNA isolation, for 1 h and 24 h for DNA and RNA isolation, and Annexin V/7AAD flow cytometry, and for 72 h for α‐smooth muscle actin/VE‐cadherin flow cytometry purposes. In another setting, HMEC‐1 were pre‐treated with telbivudine for 2 h followed by B19V infection, as described previously. Twenty‐four hours after B19V infection, uninfected HMEC‐1, untreated B19V‐infected HMEC‐1, and telbivudine pre‐treated B19V‐infected HMEC‐1 were collected for co‐culture with HL‐1 (see infra).
For the infection of UT7 cells, UT7 cells were plated at a density of 500 000 cells/12‐well. After 24 h, cells were infected with 10 000 viral particles/cell in 1 mL full UT7 medium with/out 10 ng/mL telbivudine (Santa Cruz) and cultured for 2 h at 4°C and 1 h at 37°C. One and 24 h later, cells were collected for DNA and RNA isolation.
Co‐culture of HL‐1 with parvovirus B19‐infected human microvascular endothelial cells‐1
DiO‐labelled HL‐1 were plated at a density of 250 000 cells/6‐well. Twenty‐four hours later, uninfected HMEC‐1 and infected HMEC‐1 treated with/out telbivudine were collected (see supra) and added to the HL‐1 at a ratio of 1:10. Next, cells were co‐cultured for 24 h and collected for the evaluation of DiO+ Annexin V+/7AAD‐ HL‐1 cells via flow cytometry.
Gene expression analysis
Cells were collected in Trizol (Sigma‐Aldrich), and RNA was isolated via the Trizol method. Next, RNA was DNase treated with the NucleoSpin RNA II Kit (Macherey‐Nagel, Düren, Germany) and subsequently reverse transcribed via High Capacity cDNA Reverse Transcription Kit from Applied Biosystems by Thermo Fisher Scientific (Carlsbad, CA, USA). To assess the mRNA expression of the target genes tenascin‐C, TGF‐β, and IFN‐β, mRNA expression was analysed via real‐time PCR using gene expression assays for human tenascin‐C (Hs01115665_m1), TGF‐β (Hs00171257_m1), and interferon (IFN)‐β (Hs01077958_s1) from Applied Biosystems by Thermo Fisher Scientific. mRNA expression was normalized to the housekeeping gene CDKN1b (Hs00153277_m1) and relatively expressed with the control group set as 1. For the analysis of B19V copy number and cDNA in B19V‐infected UT7 cells, the VP1/NS1 forward primer (5′‐CCTAgAAAACCCATCCTCTCTgTT‐3′), reverse primer (5′‐CCAggCTTgTgTAAg TCTTCACTAg‐3′), probe (FAM‐5′‐CCTCTAggTTCTgCATgACTgCTAC‐3′‐TAMRA) (TIB Molbiol, Berlin, Germany), and the Parvovirus B19 Standard (pB19V‐CR2.1T) from GenExpress (Berlin, Germany) were used.
Flow cytometry analysis
For the measurement of (DiO+) Annexin V+/7AAD‐, α‐smooth muscle actin+/VE‐cadherin‐, sarcomeric myosin heavy chain, and 5‐(and‐6)‐chloromethyl‐2′,7′‐dichlorodihydrofluorescein diacetate, acety ester (CM‐H2DCFDA) + cells, cells were stained with the appropriate antibodies and/or dyes and measured as described in detail previously.29, 30, 31 Samples were analysed on a MACSQuant Analyser (Miltenyi Biotec, Bergisch Gladbach, Germany). Re‐analysis of flow cytometry data was performed using FlowJo software version 8.8.6. (Tree Star Inc., Ashland, OR, USA).
Collagen content
Human microvascular endothelial cells‐1 were 24 h after stimulation with/out TNF‐α in the presence or absence of telbivudine, fixed in methanol overnight at −20°C, washed once with PBS, and incubated in 0.1% Direct Red 80 (Sirius Red) staining solution at room temperature for 60 min. After a second washing with PBS, the Sirius Red staining of the HMEC‐1 was eluted in 0.1 N sodium hydroxide at room temperature for 60 min on a rocking platform. As described previously,30 the optical density representative for the accumulation of Collagen I and III was measured at 540 nm. For normalization to cell amount, HMEC‐1 were stained with crystal violet and the absorbance was measured at 495 nm. Data are represented as the ratio of the absorbance at 540 nm (Sirius Red) towards the absorbance at 495 nm (crystal violet).
Patients
Patients with clinical evidence for myocarditis underwent left‐ventricular EMB.32 Endomyocardial biopsies were analysed by a histological, immunohistological, and molecular workup as previously described.33 One female and three male consecutive patients [mean (±SD) age 45 ± 13 years] with myocarditis [mean (±SD) 17 ± 3 CD3 cells/mm2]34 and transcriptionally active B19V infection determined by the presence of B19V DNA and VP1/VP2‐mRNA complained for symptoms of heart failure, chest discomfort, and atypical angina attacks [mean (±SD) left‐ventricular ejection fraction (EF) 43 ± 13% and mean left‐ventricular end‐diastolic diameter 59 ± 7.9; New York Heart Association (NYHA) II‐III]. The symptoms persisted for more than 4 months despite an 8 week treatment with heart failure medication. Medication remained unchanged during the complete study period. Patients underwent EMB before (pre) and immediately after termination of the antiviral treatment (post). All patients gave written informed consent to participate in the pilot trial.
Analysis of myocardial inflammation and parvovirus B19 DNA and RNA
Quantitative evaluation of inflammatory cells was performed on snap frozen sections of EMB as described previously.35, 36 The EMB diagnosis of myocardial inflammation was based on histomorphological criteria37 in combination with immunohistochemical data.15 Nucleic acids were isolated from EMB of the patients, and B19V‐DNA and B19V‐RNA loads were determined as described previously using B19V‐VP1/2‐specific quantitative PCR and reverse transcription real‐time PCR.3, 16 Parvovirus B19 RNA loads were normalized by quantification of isolated mRNAs calculated as expression of the housekeeping gene hypoxanthine‐guanine phosphoribosyltransferase (Applied Biosystems, Darmstadt, Germany).
Left ventricle analysis by matrix‐assisted laser desorption ionization–imaging mass spectrometry
Sections of 7 μm were prepared from paraffin blocks and transferred onto indium‐tin‐oxide slides (Bruker Daltonics, Billerica, MA, USA). Next, sections were dewaxed twice in xylene for 3 min each and passed through decreasing concentrations of ethanol (2 × 100%, 95%, and 70%) for 1 min each. An endonuclease and trypsin solution was directly applied onto the section using an automated spraying device. Tissue incubation with the trypsin solution was performed for 3 h at 37°C in a moist chamber. Following trypsinization, the matrix solution (α‐Cyano‐4‐hydroxycinnamic acid) was applied in ImagePrep using the manufacturer's protocol.
Matrix‐assisted laser desorption ionization (MALDI)–imaging mass spectrometry (IMS) data acquisition was performed on an Autoflex III MALDI‐TOF/TOF with flexControl 3.0 and flexImaging 3.0 software (Bruker Daltonik) in a positive ion reflector mode and in a range of m/z 800–3500 Da and a raster width of 80 μm. After MALDI‐IMS experiments, the matrix was removed with 70% ethanol and the tissue sections were stained with haematoxylin and eosin.
The software SCiLS Lab (SCiLS Lab 2015a) was used for unsupervised characterization of the heart tissue biopsies data. This pipeline starts with data pre‐processing, including baseline removal, normalization, and peak selection. Data mining was performed first by using principal component analysis described by Klerk et al.38 for mass spectra. Followed to principal component analysis, data from each group were compared pairwise in order to obtain discriminative m/z values for the groups by using receiver operating characteristic (area under the curve < 0.4) analysis. P values (<0.05) were calculated by the Wilcoxon rank‐sum test.
Discriminative m/z values were identified by bottom up by a ‘bottom‐up’‐nano liquid chromatography–tandem mass spectrometry (MS/MS) protein identification approach from an adjacent tissue section. The raw spectra from the ‘bottom‐up’‐nano liquid chromatography–MS/MS analysis were all converted to mascot generic files (.mgf) using the ProteoWizard software. The resulting .mgf files were then searched against a Homo sapiens protein database downloaded from the UniProt database UniProt using the Mascot search engine (version 2.4, MatrixScience, Boston, MA, USA). Comparison of MALDI‐IMS and LC‐MS/MS m/z values requires the identification of more than one peptide (mass differences < 0.9 Da).
Statistical analysis
Statistical analysis was performed using Prism 7 for Mac OS X (GraphPad Software, Inc., La Jolla, CA, USA). Ordinary one‐way ANOVA was used for statistical analysis of the data with correction for multiple comparisons via the Tukey test. Pairwise comparison was performed via the paired Student's t‐test. Data are presented as mean ± SEM. Differences were considered to be significant when the two‐sided P value was lower than 0.05.
Results
Telbivudine reduces parvovirus B19‐induced damage in human microvascular endothelial cells
Parvovirus B19 induced endothelial damage in HMEC‐1 as obviated by a 2.3‐fold (P < 0.0001) induction of apoptosis and a 2.7‐fold (P < 0.0001) increase in endothelial‐to‐mesenchymal transition (EndMT), a stress‐induced phenomena by which endothelial cells differentiate into fibroblasts30 contributing to cardiac fibrosis.39 In parallel to the induction of EndMT, B19V up‐regulated tenascin‐C mRNA. Telbivudine abrogated the B19V‐induced apoptosis and EndMT by 1.3‐fold (P = 0.01) and 1.6‐fold (P < 0.005), respectively (Figure 1 A‐C). Concomitantly, treatment with telbivudine was also associated with a 2.9‐fold (P < 0.0005) and 1.8‐fold (P < 0.05) drop in tenascin‐C and TGF‐β mRNA expression, respectively (Figure 1 D‐E), and a 3.4‐fold (P < 0.05) increase in IFN‐β mRNA expression (Figure 1 F). Interestingly, HMEC‐1 infected with B19V and subsequently co‐cultured with HL‐1 cells induced apoptosis in HL‐1 cells by 3.4‐fold (P < 0.0001), whereas uninfected HMEC‐1 did not induce apoptosis in HL‐1 cells. This pro‐apoptotic effect of B19V‐infected HMEC‐1 on HL‐1 cells could be partly abrogated (1.5‐fold; P < 0.0005) when HMEC‐1 were treated with telbivudine before B19V infection (Supporting Information, Figure S1 ). In frame with the non‐permissive nature of HMEC‐1 for B19V,40 B19V did not replicate in HMEC‐1. In fact, B19V copy number and cDNA decreased over time post‐B19V infection (Figure 2 B‐C). Here, telbivudine did not affect B19V DNA and cDNA copy number 24 h post‐infection (Figure 2 B‐C). In contrast to HMEC‐1, B19V copy number and cDNA increased by 3.4‐fold (P < 0.00001) and 1633‐fold (P < 0.00001) between 1 h and 24 h post‐infection in the permissive cell line UT7/EPO respectively. Whereas telbivudine did not affect B19V copy number 24 h post‐infection (Figure 3 B), it moderately (1.3‐fold; P < 0.005) decreased B19V cDNA expression 24 h post‐infection vs. untreated B19V‐infected cells (Figure 3 C).
Figure 1.
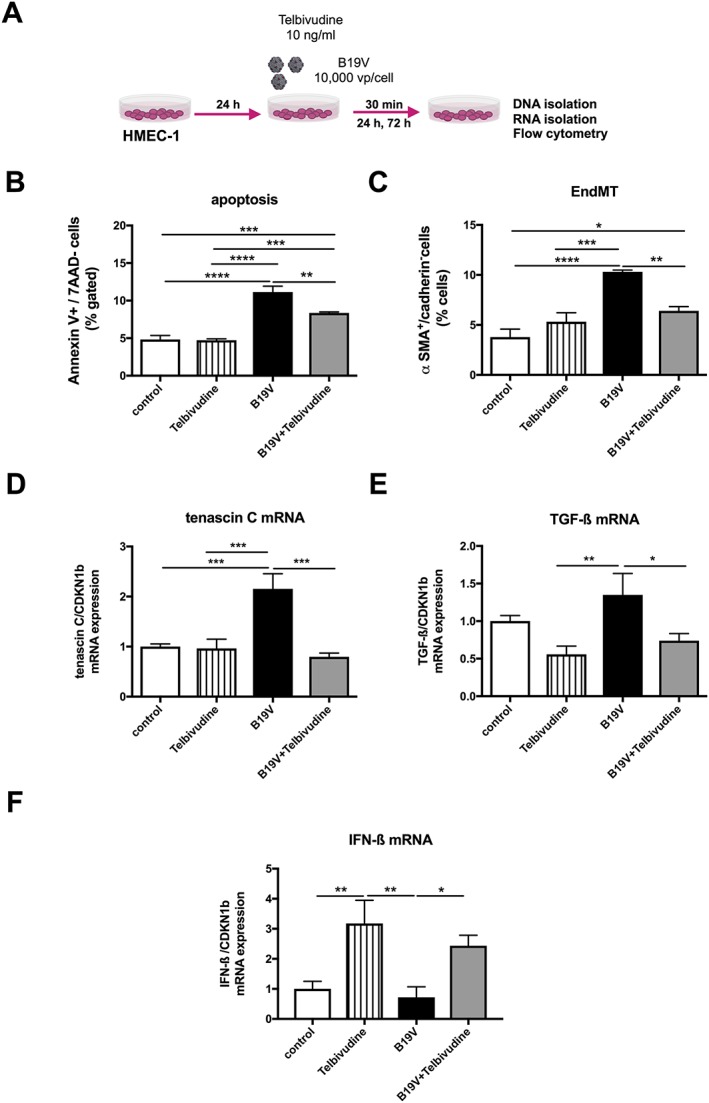
Impact of telbivudine on parvovirus B19‐infected human microvascular endothelial cells. (A) Experimental design: human microvascular endothelial cells (HMEC)‐1 were 24 h after plating, infected with 10 000 viral particles/cell with/out 10 ng/mL of telbivudine, and collected after 30 min for the analysis of tenascin‐C and TGF‐β1 mRNA expression, after 24 h for the analysis of apoptosis, and after 72 h for the anaylsis of endothelial‐to‐mesenchymal transition (EndMT). Bar graphs represent the mean ± SEM of (B) Annexin V/7AAD‐ cells (% gated), (C) α‐smooth muscle actin (SMA)+/VE‐cadherin‐ cells (% gated), (D) tenascin‐C, (E) transforming growth factor (TGF)‐β, and (F) interferon (IFN)‐β mRNA expression, as indicated, with n = 4–6/group and *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.
Figure 2.
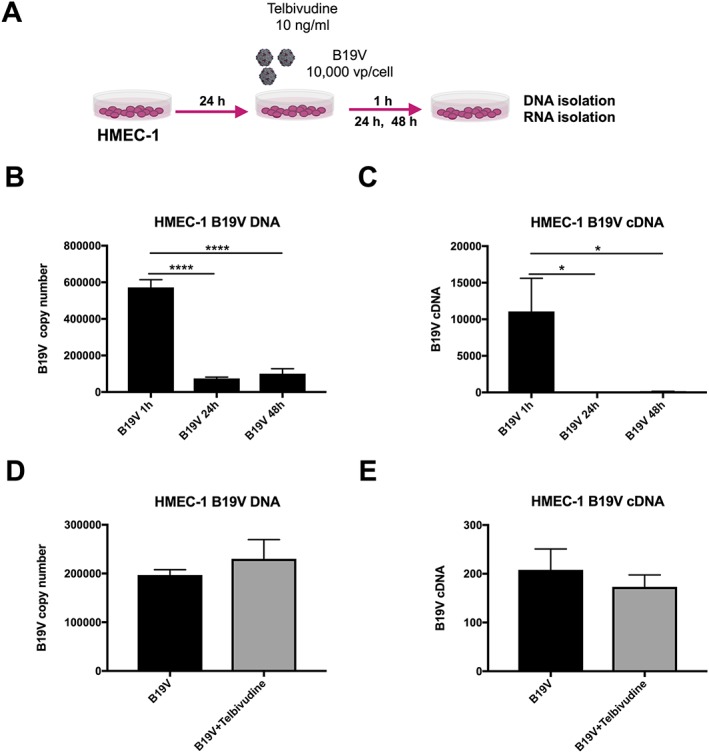
Impact of telbivudine on parvovirus B19 (B19V) copy number and expression in B19V‐infected human microvascular endothelial cells. (A) Experimental design: human microvascular endothelial cells (HMEC‐1) were 24 h after plating, infected with 10 000 viral particles/cell with/out 10 ng/mL of telbivudine, and collected after 1, 24, or 48 h for the analysis of B19V DNA copy number and mRNA expression. Bar graphs represent the mean ± SEM of (B) B19V copy number and (C) B19V cDNA, 1, 24, and 48 h post‐B19V infection in HMEC‐1 with n = 5–6/group and *P < 0.05 and ****P < 0.0001, (D) B19V copy number (n = 4/group), and (E) B19V cDNA expression (n = 6/group) 24 h post‐infection with/out telbivudine treatment.
Figure 3.
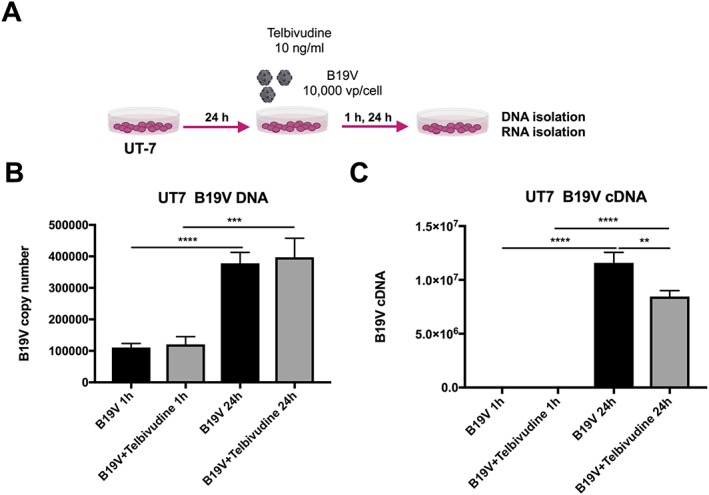
Impact of telbivudine on parvovirus B19 (B19V) copy number and expression in B19V‐infected UT7/EPO S1 cells. (A) Experimental design: UT7/EPO S1 cells were 24 h after plating, infected with 10 000 viral particles/cell with/out 10 ng/mL of telbivudine, and collected after 1 and 24 h for the analysis of B19V DNA copy number and mRNA expression. Bar graphs represent the mean ± SEM of (B) B19V copy number and (C) B19V cDNA, 1 and 24 h post‐B19V infection with/out telbivudine treatment in UT7/EPO S1 cells with n = 4–6/group and **P < 0.005, ***P < 0.0005, and ****P < 0.0001.
Telbivudine exerts pleiotropic effects
To investigate whether the endothelial‐protective effects of telbivudine were also beyond B19V‐induced stress conditions, HMEC‐1 were stressed with TNF‐α and treated with telbivudine (Figure 4 A). Similar to B19V conditions, telbivudine decreased the TNF‐α‐induced endothelial apoptosis by 1.5‐fold (P < 0.0001) (Figure 4 B). Furthermore, telbivudine reduced the TNF‐α‐up‐regulated collagen deposition in HMEC‐141 by 1.2‐fold (P < 0.05) (Figure 4 C). To next evaluate whether the protective effects of telbivudine were also present in other cells than endothelial cells, HL‐1 cells were stressed with Ang II and murine fibroblasts with TGF‐β1 and the impact of telbivudine on oxidative stress and/or cardiomyocyte hypertrophy was analysed (Figure 4 D and G). In HL‐1 cells, Ang II induced ROS by 2.0‐fold (P < 0.0001), whereas telbivudine decreased ROS by 1.3‐fold (P < 0.001) (Figure 4 E). The reduction in oxidative stress was paralleled by a decline in cardiomyocyte hypertrophy as indicated by 1.6‐fold (P < 0.0001) lower sarcomeric myosin heavy chain levels (Figure 4 F). In murine fibroblasts, TGF‐β1 induced ROS by 5.5‐fold (P < 0.0001), which could be 2.1‐fold (P < 0.0001) abrogated by telbivudine treatment (Figure 4 H).
Figure 4.
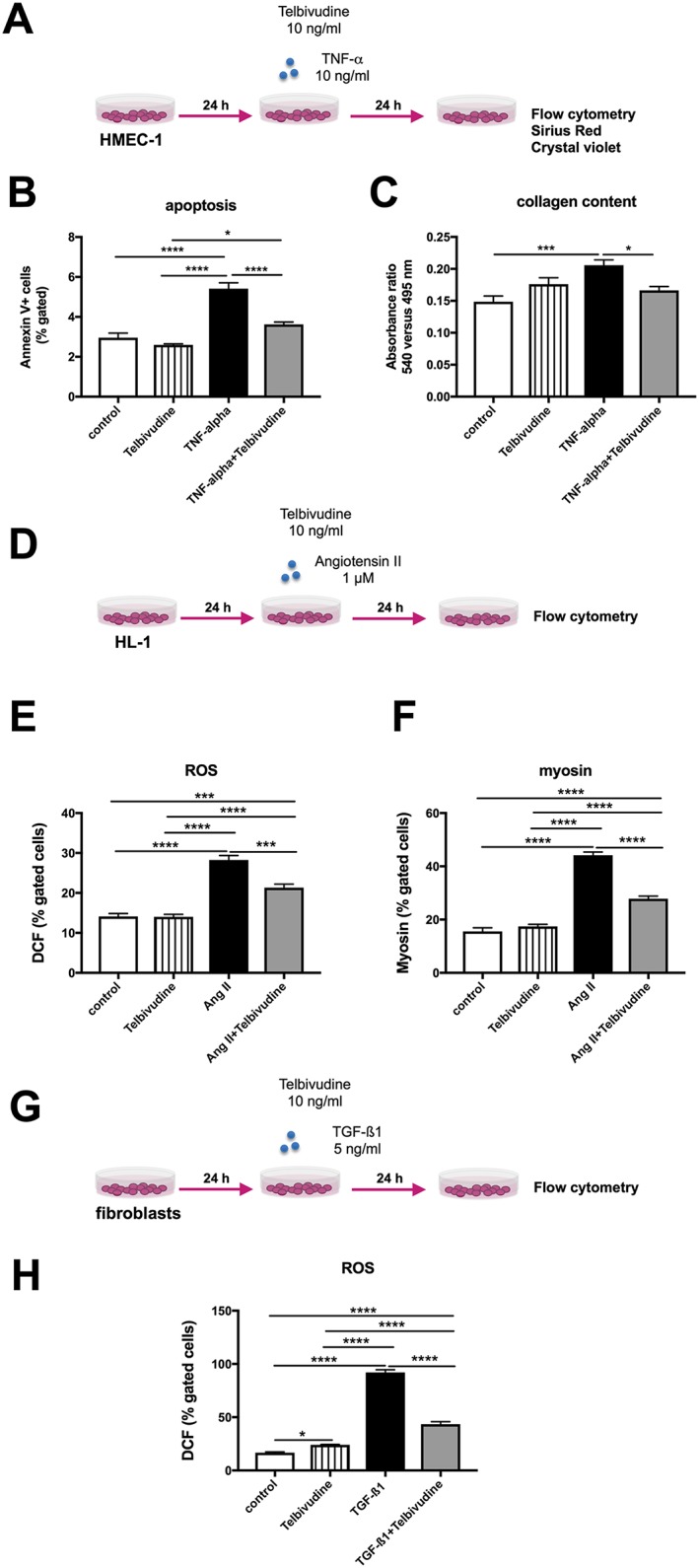
Pleiotropic effects of telbivudine. The pleiotropic effects of telbivudine were evaluated in tumour necrosis factor (TNF)‐α‐stimulated human microvascular endothelial cells (HMEC)‐1 (A‐C), angiotensin (Ang) II‐stimulated HL‐1 cells (D‐F), and transforming growth factor (TGF)‐β1‐stressed fibroblasts (G‐H). (A) Experimental design: HMEC‐1 were 24 h after plating, stimulated with/out 10 ng/mL of TNF‐α in the presence or absence of 10 ng/mL of telbivudine, and collected after 24 h for the analysis of apoptosis, or stained with Sirius Red and crystal violet for the quantification of collagen deposition. Bar graphs represent the mean ± SEM of (B) Annexin V/7AAD‐ cells (% gated) (n = 5–6/group), (C) the ratio of the absorbances at 495 to 405 nm (n = 10–12/group). (D) Experimental design: HL‐1 were 24 h after plating, stimulated with/out 1 μM Ang II in the presence or absence of 10 ng/mL of telbivudine, and collected after 24 h for the analysis of reactive oxygen species (ROS) and myosin. Bar graphs represent the mean ± SEM of (E) DCF (% gated cells) (n = 5–6/group), (F) myosin (% gated cells) (n = 6/group). (G) Experimental design: fibroblasts were 24 h after plating, stimulated with/out 5 ng/mL of TGF‐β1 in the presence or absence of 10 ng/mL of telbivudine, and collected after 24 h for the analysis of ROS. Bar graphs represent the mean ± SEM of (H) DCF (% gated cells) (n = 6/group). For the entire panel: *P < 0.05, ***P < 0.0005, and ****P < 0.0001.
Impact of telbivudine on parvovirus B19‐positive patients
Following the promising cell culture data with telbivudine, a single‐patient use was started in which the efficacy of telbivudine in clinically suffering myocarditis patients with persistent B19V transcriptional activity was evaluated. Upon telbivudine treatment, B19V transcriptional activity decreased to undetectable levels along with a reduction in CD3/macrophage numbers (cells/mm2) (Table 1). In parallel, telbivudine resulted in an amelioration of EF by 10% (P < 0.005; paired t‐test) and a reduction in chest pain, reflected in an improvement of NYHA class (Table 1). No side effects (changes in troponin T, creatinine kinase, and transaminases levels, electrocardiogram changes, arrhythmias, or muscle pain) were observed. Imaging mass spectrometry and subsequent principal component analysis could distinguish the protein signatures of B19V mRNA+ and B19V mRNA− EMB (Figure 5 A). Peptides from the extracellular matrix glycoprotein tenascin‐C (1718 and 1740 Da) show significantly lower intensities (receiver operating characteristic > 0.65; P < 0.01) in B19V mRNA− compared with B19V mRNA+ EMB (Figure 5 B).
Table 1.
Patient characteristics before and after telbivudine treatment
| CD3/macrophages (cells/mm2) |
DNA (copy number) |
mRNA (copy number) |
Ejection fraction (%) | New York Heart Association | |
|---|---|---|---|---|---|
| Before | |||||
| Patient 1 | 9/31 | 2642 | 1583 | 57 | II |
| Patient 2 | 9/34 | 16 | 90 | 35 | II |
| Patient 3 | 21/9 | 15 | 53 | 30 | III |
| Patient 4 | 28/178 | 145 | 371 | 50 | III |
| After | |||||
| Patient 1 | 0/16 | 196 | ND | 64 | I‐II |
| Patient 2 | 0/8 | 72 | ND | 45 | I‐II |
| Patient 3 | 7/11 | 189 | ND | 42 | II |
| Patient 4 | 6/31 | 418 | ND | 61 | II |
ND, not detectable.
Figure 5.
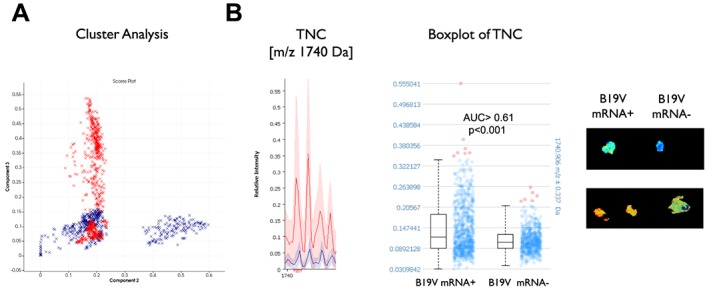
In situ tissue typing of endomyocardial biopsies in parvovirus B19‐positive myocarditis patients before (mRNA+) and after telbivudine treatment (mRNA−) via imaging mass spectrometry. (A) Probabilistic latent semantic analysis could distinguish the protein signatures of the B19V mRNA+ (red) and B19V mRNA− (blue) patients (Patients 3 and 4 of Table 1). (B) Peptides from tenascin‐C (TNC; 1740 Da) show significantly lower intensities (receiver operating characteristic > 0.72; P < 0.001) in B19 mRNA− compared with B19V mRNA+ endomyocardial biopsy.
Study design of PreTopic study
Based on the results of the pilot trial, the proof of concept ‘PreTopic’ study (EudraCT‐Number: 2016‐004825‐17) was designed: a Proof of concept study to investigate the Efficacy of telbivudine Over Placebo in patients with parvovirus‐associated Inflammatory Cardiomyopathy, a mono‐centre, interventional, placebo‐controlled, randomized, double‐blind, and exploratory trial (Figure 6 ). Patients suspected for inflammatory cardiomyopathy will be screened. Twenty‐six patients who are mRNA B19V positive, but adenovirus/enterovirus negative, and who have an EF ≤ 50% for >6 weeks will be included. Thirteen patients will receive placebo, and 13 will receive verum on top of normal heart failure treatment for 3 months (Figure 6 ). The primary end point is the change of the patient's quality of life between the baseline visit (Week 0) and the last visit (Week 12). The quality of life will be assessed via the Minnesota Living with Heart Failure Questionnaire (Annex I). Secondary end points include changes in cardiac function, biomarker levels, and safety.
Figure 6.
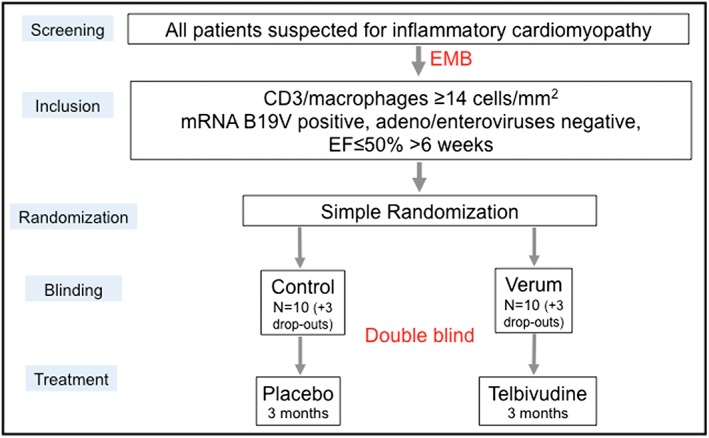
Study design of the PreTopic study. EF, ejection fraction; EMB, endomyocardial biopsy.
Discussion
The present study demonstrates that telbivudine reduces B19V‐induced endothelial apoptosis and EndMT in B19V‐infected endothelial cells without affecting B19V copy number. In permissive UT7/EPO cells, telbivudine slightly reduced B19V cDNA. Further experiments in TNF‐α‐stressed endothelial cells indicate that the telbivudine‐mediated endothelial‐protective effects also take place independent of B19V. This finding could further be supported in HL‐1 cells supplemented with Ang II and TGF‐β‐stressed fibroblasts. In patients with transcriptional active B19V‐associated inflammatory cardiomyopathy, telbivudine improved cardiac function and clinical symptoms and reduced the presence of inflammatory infiltrates, which was accompanied with an elimination of B19V cDNA levels.
Parvovirus B19 is implicated as a pathogen in the majority of chronic myocarditis cases.3 However, direct evidence showing a direct causal relationship between B19V cardiac presence and myocarditis is still missing, as well as therapies for B19V‐associated inflammatory cardiomyopathy.19
The strongest risk factor for impaired outcome in chronic myocarditis and inflammatory cardiomyopathy is the persistence of cardiac inflammation.42 Based on single‐centre experiences,43, 44, 45 the current European Society of Cardiology scientific statement recommends in EMB‐proven, virus‐negative patients with lymphocytic myocarditis an immunosuppressive therapy with prednisone and azathioprine to improve cardiac function.34 Based on the hypothesis that the detection of viral genomes indicates active infection, immunosuppression is avoided in cardiotropic viral genome‐positive patients. The implication is that immunosuppression is withheld in most of these patients, because in ~70% of those patients, a B19V persistence can be detected.15, 46, 47 Thus, for this clinical scenario, there is an unmet need to develop new immunomodulatory or antiviral therapy strategies that do not carry the risk of viral stimulation due to global immune suppression as it is known for cortisone. The antiviral and anti‐inflammatory properties of telbivudine24, 25, 26, 27 inspired us to evaluate its potential to treat B19V‐associated inflammatory cardiomyopathy. Given the vasculotrope nature of B19V2, 3 and the evidence that B19V modulates inflammatory signalling and apoptosis in endothelial cells,20 we evaluated at first potential endothelial‐protective effects of telbivudine in B19V‐infected endothelial cells. In agreement with the pro‐apoptotic and pro‐inflammatory potential of B19V, endothelial cell apoptosis and the inflammation/TGF‐β‐triggered phenomena,30 EndMT, were observed following B19V infection. Accordingly, mRNA expression of TGF‐β1 and of tenascin‐C, which is well known to induce epithelial‐to‐mesenchymal transition,48 was increased upon B19V infection. Interestingly, B19V‐infected endothelial cells induced apoptosis in HL‐1 cardiomyocytes upon co‐culture, a finding that could not be observed with non‐infected endothelial cells. This suggests that the inflammatory status of the B19V‐infected endothelial cell was transferred on the HL‐1 cardiomyocytes, confirming the relevance of endothelial cell–cardiomyocyte interactions.49 In agreement with the non‐permissive nature of HMEC‐1 for B19V,40 B19V did not replicate in HMEC‐1. Here, telbivudine did not affect B19V copy numbers, nor cDNA expression, but decreased B19V‐induced endothelial apoptosis, EndMT, TGF‐β1, and tenascin‐C mRNA expression. In addition, it abrogated the pro‐apoptotic effect of B19V‐infected HMEC‐1 on HL‐1 cardiomyocytes and induced the mRNA expression of antiviral IFN‐β. Interferon‐β has previously been shown to diminish B19V‐induced endothelial damage in patients with cardiac B19V persistence.4 The endothelial‐protective effects of telbivudine were further confirmed in TNF‐α‐stimulated endothelial cells where telbivudine decreased the TNF‐α‐induced apoptosis and collagen expression. Furthermore, telbivudine could block the Ang II‐induced and TGF‐β1‐induced oxidative stress in HL‐1 cardiomyocytes and fibroblasts, respectively, indicating multimodal protective effects of telbivudine on different cells.
Following our in vitro results and because of the absence of B19V animal models, a single‐patient use was initiated in the attempt to improve the status of clinically severely suffering patients with EMB‐proven chronic lymphocytic myocarditis associated with B19V‐transcriptional activity. Four patients, complaining for atypical chest pain for more than 6 months, were treated with telbivudine (600 mg/day for 6 months) on top of symptomatic standard therapy. Telbivudine treatment decreased the cardiac presence of transcriptional active B19V in all treated patients and improved their clinical symptoms as shown by an improvement in EF and a reduction in NYHA class. This finding cannot be completely explained by our in vitro experiments because B19V copy numbers were not affected in endothelial cells and only slightly reduced in UT7/EPO cells by telbivudine. Therefore, we suggest that a direct antiviral B19V effect of telbivudine seems to be unlikely, as proposed by others.28 Though, in vitro, telbivudine increased antiviral IFN‐β in B19V‐infected cells, which implicates at least indirect antiviral and anti‐inflammatory endothelial‐beneficial effects, which might belong to the effect we found in patients. However, in vivo immune responses are complex and cannot be completely mimicked in isolated cell culture settings. Nevertheless, it is an important finding per se that B19V replication did not boost following telbivudine. The pleiotropic immunomodulatory effects of telbivudine are therefore unique because they are at least not associated with a stimulation of B19V replication.
Matrix‐assisted laser desorption ionization–imaging mass spectrometry further illustrated a change in cardiac protein pattern between B19V‐positive patients before and after telbivudine treatment. In agreement with our in vitro findings, IMS revealed a decrease in tenascin‐C expression before and after telbivudine treatment. Besides its potential to induce the inflammation‐triggered processes epithelial‐mesenchymal transition48 and EndMT, the matricellular protein tenascin‐C also promotes a cardiac pro‐inflammatory micro‐environment and macrophage migration.50 Along with the decreased expression of tenascin‐C, patients with B19V‐associated inflammatory cardiomyopathy (≥14 CD3 cells and macrophages/mm2) exhibited a lower number of CD3 infiltrates after telbivudine treatment.
Findings from a single‐patient‐use approach have a case report character and do not include a placebo control. Therefore, we cannot completely exclude that the decrease in B19V mRNA and the improvement in clinical phenotype were not also influenced due to a natural course of the disease, although we started with our intervention >6 months after disease onset. Importantly, patients improved during telbivudine treatment safely without aggravation of viral load. Our promising results need further validation in a clinical exploratory trial: the PreTopic study (EudraCT‐Number: 2016‐004825‐17).
We conclude that telbivudine exerts endothelial‐protective effects in B19V‐infected endothelial cells and can improve B19V‐associated inflammatory cardiomyopathy following early findings of patients investigated in a single‐use approach. Hitherto, we do not recommend the general use of telbivudine in B19V‐associated inflammatory cardiomyopathy until its effectivity has been proven in clinical placebo‐controlled studies.
Conflict of interest
None.
Funding
This study was supported by the Berlin‐Brandenburg Center for Regenerative Therapies—BCRT (Bundesministerium für Bildung und Forschung—0313911) to C.T.
Supporting information
Figure S1. Telbivudine pretreatment of parvovirus B19‐infected endothelial cells reduces the pro‐apoptotic potential of parvovirus B19‐infected endothelial cells upon co‐culture in HL‐1 cardiomyocytes. A. Experimental design: HMEC‐1 were 24 h after plating, treated with/out Telbivudine for 2 h, next infected with 10 000 viral particles/cell and collected after 24 h for co‐culture on DiO‐labelled HL‐1 cells, which were plated 24 h before. After 24 h of co‐culture, cells were collected for the anaylsis of DiO+Annexin V/7AAD‐ cells by flow cytometry. Bar graphs represent the mean ± SEM of B. DiO+Annexin V/7AAD‐ HL‐1 cells (% gated), with n=6/group and ***p<0.0005 and ****p<0.0001.
Acknowledgements
We acknowledge Annika Koschel and Kerstin Puhl for excellent technical assistance.
Van Linthout, S. , Elsanhoury, A. , Klein, O. , Sosnowski, M. , Miteva, K. , Lassner, D. , Abou‐El‐Enein, M. , Pieske, B. , Kühl, U. , and Tschöpe, C. (2018) Telbivudine in chronic lymphocytic myocarditis and human parvovirus B19 transcriptional activity. ESC Heart Failure, 5: 818–829. 10.1002/ehf2.12341.
References
- 1. Adamson‐Small LA, Ignatovich IV, Laemmerhirt MG, Hobbs JA. Persistent parvovirus B19 infection in non‐erythroid tissues: possible role in the inflammatory and disease process. Virus Res 2014; 190: 8–16. [DOI] [PubMed] [Google Scholar]
- 2. Bultmann BD, Sotlar K, Klingel K. Parvovirus B19. N Engl J Med 2004; 350: 2006–2007 author reply 2006‐7. [DOI] [PubMed] [Google Scholar]
- 3. Bock CT, Klingel K, Kandolf R. Human parvovirus B19‐associated myocarditis. N Engl J Med 2010; 362: 1248–1249. [DOI] [PubMed] [Google Scholar]
- 4. Schmidt‐Lucke C, Spillmann F, Bock T, Kuhl U, Van Linthout S, Schultheiss HP, Tschope C. Interferon beta modulates endothelial damage in patients with cardiac persistence of human parvovirus B19 infection. J Infect Dis 2010; 201: 936–945. [DOI] [PubMed] [Google Scholar]
- 5. Tschope C, Bock CT, Kasner M, Noutsias M, Westermann D, Schwimmbeck PL, Pauschinger M, Poller WC, Kuhl U, Kandolf R, Schultheiss HP. High prevalence of cardiac parvovirus B19 infection in patients with isolated left ventricular diastolic dysfunction. Circulation 2005; 111: 879–886. [DOI] [PubMed] [Google Scholar]
- 6. Bachelier K, Biehl S, Schwarz V, Kindermann I, Kandolf R, Sauter M, Ukena C, Yilmaz A, Sliwa K, Bock CT, Klingel K, Bohm M. Parvovirus B19‐induced vascular damage in the heart is associated with elevated circulating endothelial microparticles. PLoS One 2017; 12: e0176311. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kuhl U, Lassner D, Pauschinger M, Gross UM, Seeberg B, Noutsias M, Poller W, Schultheiss HP. Prevalence of erythrovirus genotypes in the myocardium of patients with dilated cardiomyopathy. J Med Virol 2008; 80: 1243–1251. [DOI] [PubMed] [Google Scholar]
- 8. Schenk T, Enders M, Pollak S, Hahn R, Huzly D. High prevalence of human parvovirus B19 DNA in myocardial autopsy samples from subjects without myocarditis or dilative cardiomyopathy. J Clin Microbiol 2009; 47: 106–110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Lotze U, Egerer R, Gluck B, Zell R, Sigusch H, Erhardt C, Heim A, Kandolf R, Bock T, Wutzler P, Figulla HR. Low level myocardial parvovirus B19 persistence is a frequent finding in patients with heart disease but unrelated to ongoing myocardial injury. J Med Virol 2010; 82: 1449–1457. [DOI] [PubMed] [Google Scholar]
- 10. Koepsell SA, Anderson DR, Radio SJ. Parvovirus B19 is a bystander in adult myocarditis. Cardiovasc Pathol 2012; 21: 476–481. [DOI] [PubMed] [Google Scholar]
- 11. Bihari C, Rastogi A, Saxena P, Rangegowda D, Chowdhury A, Gupta N, Sarin SK. Parvovirus B19 associated hepatitis. Hepat Res Treat 2013; 2013: 472027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Naciute M, Mieliauskaite D, Rugiene R, Maciunaite G, Mauricas M, Murovska M, Girkontaite I. Parvovirus B19 infection modulates the levels of cytokines in the plasma of rheumatoid arthritis patients. Cytokine 2017; 96: 41–48. [DOI] [PubMed] [Google Scholar]
- 13. Gama BE, Emmel VE, Oliveira‐Silva M, Gutiyama LM, Arcuri L, Colares M, de Cassia Tavares R, Bouzas LF, Abdelhay E, Hassan R. Parvovirus B19 in the context of hematopoietic stem cell transplantation: evaluating cell donors and recipients. Transplant Direct 2017; 3: e217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Kupin WL. Viral‐associated GN: hepatitis B and other viral infections. Clin J Am Soc Nephrol 2017; 12: 1529–1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Kuhl U, Pauschinger M, Noutsias M, Seeberg B, Bock T, Lassner D, Poller W, Kandolf R, Schultheiss HP. High prevalence of viral genomes and multiple viral infections in the myocardium of adults with “idiopathic” left ventricular dysfunction. Circulation 2005; 111: 887–893. [DOI] [PubMed] [Google Scholar]
- 16. Kuhl U, Lassner D, Dorner A, Rohde M, Escher F, Seeberg B, Hertel E, Tschope C, Skurk C, Gross UM, Schultheiss HP, Poller W. A distinct subgroup of cardiomyopathy patients characterized by transcriptionally active cardiotropic erythrovirus and altered cardiac gene expression. Basic Res Cardiol 2013; 108: 372. [DOI] [PubMed] [Google Scholar]
- 17. Bock CT, Duchting A, Utta F, Brunner E, Sy BT, Klingel K, Lang F, Gawaz M, Felix SB, Kandolf R. Molecular phenotypes of human parvovirus B19 in patients with myocarditis. World J Cardiol 2014; 6: 183–195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Dennert R, van Paassen P, Wolffs P, Bruggeman C, Velthuis S, Felix S, van Suylen RJ, Crijns HJ, Cohen Tervaert JW, Heymans S. Differences in virus prevalence and load in the hearts of patients with idiopathic dilated cardiomyopathy with and without immune‐mediated inflammatory diseases. Clin Vaccine Immunol 2012; 19: 1182–1187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Van Linthout S, Tschope C, Schultheiss HP. Lack in treatment options for virus‐induced inflammatory cardiomyopathy: can iPS‐derived cardiomyocytes close the gap? Circ Res 2014; 115: 540–541. [DOI] [PubMed] [Google Scholar]
- 20. Duechting A, Tschope C, Kaiser H, Lamkemeyer T, Tanaka N, Aberle S, Lang F, Torresi J, Kandolf R, Bock CT. Human parvovirus B19 NS1 protein modulates inflammatory signaling by activation of STAT3/PIAS3 in human endothelial cells. J Virol 2008; 82: 7942–7952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Schmidt‐Lucke C, Zobel T, Schrepfer S, Kuhl U, Wang D, Klingel K, Becher PM, Fechner H, Pozzuto T, Van Linthout S, Lassner D, Spillmann F, Escher F, Holinski S, Volk HD, Schultheiss HP, Tschope C. Impaired endothelial regeneration through human parvovirus B19‐infected circulating angiogenic cells in patients with cardiomyopathy. J Infect Dis 2015; 212: 1070–1081. [DOI] [PubMed] [Google Scholar]
- 22. Schmidt‐Lucke C, Zobel T, Escher F, Tschope C, Lassner D, Kuhl U, Gubbe K, Volk HD, Schultheiss HP. Human parvovirus B19 (B19V) up‐regulates CXCR4 surface expression of circulating angiogenic cells: implications for cardiac ischemia in B19V cardiomyopathy. J Infect Dis 2018; 217: 456–465. [DOI] [PubMed] [Google Scholar]
- 23. Dettmeyer R, Kandolf R, Baasner A, Banaschak S, Eis‐Hubinger AM, Madea B. Fatal parvovirus B19 myocarditis in an 8‐year‐old boy. J Forensic Sci 2003; 48: 183–186. [PubMed] [Google Scholar]
- 24. Meng N, Gao X, Yan W, Wang M, Liu P, Lu XD, Zhang SJ, Lu YQ, Tang WX. Efficacy of telbivudine in the treatment of chronic hepatitis B and liver cirrhosis and its effect on immunological responses. J Huazhong Univ Sci Technolog Med Sci 2015; 35: 230–234. [DOI] [PubMed] [Google Scholar]
- 25. Pan X, Yao W, Fu J, Liu M, Li L, Gao X. Telbivudine improves the function of myeloid dendritic cells in patients with chronic hepatitis B. Acta Virol 2012; 56: 31–38. [DOI] [PubMed] [Google Scholar]
- 26. Li J, Jia M, Liu Y, She W, Li L, Wang J, Jiang W. Telbivudine therapy may shape CD4(+) T‐cell response to prevent liver fibrosis in patients with chronic hepatitis B. Liver Int 2015; 35: 834–845. [DOI] [PubMed] [Google Scholar]
- 27. Chen Y, Li X, Ye B, Yang X, Wu W, Chen B, Pan X, Cao H, Li L. Effect of telbivudine therapy on the cellular immune response in chronic hepatitis B. Antiviral Res 2011; 91: 23–31. [DOI] [PubMed] [Google Scholar]
- 28. Ozawa K, Kurtzman G, Young N. Productive infection by B19 parvovirus of human erythroid bone marrow cells in vitro . Blood 1987; 70: 384–391. [PubMed] [Google Scholar]
- 29. Van Linthout S, Savvatis K, Miteva K, Peng J, Ringe J, Warstat K, Schmidt‐Lucke C, Sittinger M, Schultheiss HP, Tschope C. Mesenchymal stem cells improve murine acute coxsackievirus B3‐induced myocarditis. Eur Heart J 2011; 32: 2168–2178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Spillmann F, Miteva K, Pieske B, Tschope C, Van Linthout S. High‐density lipoproteins reduce endothelial‐to‐mesenchymal transition. Arterioscler Thromb Vasc Biol 2015; 35: 1774–1777. [DOI] [PubMed] [Google Scholar]
- 31. Miteva K, Van Linthout S, Pappritz K, Muller I, Spillmann F, Haag M, Stachelscheid H, Ringe J, Sittinger M, Tschope C. Human endomyocardial biopsy specimen‐derived stromal cells modulate angiotensin II‐induced cardiac remodeling. Stem Cells Transl Med 2016; 5: 1707–1718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Tschope C, Kherad B, Schultheiss HP. How to perform an endomyocardial biopsy? Turk Kardiyol Dern Ars 2015; 43: 572–575. [DOI] [PubMed] [Google Scholar]
- 33. Dominguez F, Kuhl U, Pieske B, Garcia‐Pavia P, Tschöpe C. Update on myocarditis and inflammatory cardiomyopathy: reemergence of endomyocardial biopsy. Rev Esp Cardiol (Engl Ed) 2016; 69: 178–187. [DOI] [PubMed] [Google Scholar]
- 34. Caforio AL, Pankuweit S, Arbustini E, Basso C, Gimeno‐Blanes J, Felix SB, Fu M, Helio T, Heymans S, Jahns R, Klingel K, Linhart A, Maisch B, McKenna W, Mogensen J, Pinto YM, Ristic A, Schultheiss HP, Seggewiss H, Tavazzi L, Thiene G, Yilmaz A, Charron P, Elliott PM, M. European Society of Cardiology Working Group on, D. Pericardial . Current state of knowledge on aetiology, diagnosis, management, and therapy of myocarditis: a position statement of the European Society of Cardiology Working Group on Myocardial and Pericardial Diseases. Eur Heart J 2013; 34: 2636–2648, 2648a‐2648d. [DOI] [PubMed] [Google Scholar]
- 35. Noutsias M, Seeberg B, Schultheiss HP, Kuhl U. Expression of cell adhesion molecules in dilated cardiomyopathy: evidence for endothelial activation in inflammatory cardiomyopathy. Circulation 1999; 99: 2124–2131. [DOI] [PubMed] [Google Scholar]
- 36. Noutsias M, Pauschinger M, Ostermann K, Escher F, Blohm JH, Schultheiss H, Kuhl U. Digital image analysis system for the quantification of infiltrates and cell adhesion molecules in inflammatory cardiomyopathy. Med Sci Monit 2002; 8: MT59–MT71. [PubMed] [Google Scholar]
- 37. Aretz HT. Myocarditis: the Dallas criteria. Hum Pathol 1987; 18: 619–624. [DOI] [PubMed] [Google Scholar]
- 38. Klerk LA, Dankers PY, Popa ER, Bosman AW, Sanders ME, Reedquist KA, Heeren RM. TOF‐secondary ion mass spectrometry imaging of polymeric scaffolds with surrounding tissue after in vivo implantation. Anal Chem 2010; 82: 4337–4343. [DOI] [PubMed] [Google Scholar]
- 39. Zeisberg EM, Tarnavski O, Zeisberg M, Dorfman AL, McMullen JR, Gustafsson E, Chandraker A, Yuan X, Pu WT, Roberts AB, Neilson EG, Sayegh MH, Izumo S, Kalluri R. Endothelial‐to‐mesenchymal transition contributes to cardiac fibrosis. Nat Med 2007; 13: 952–961. [DOI] [PubMed] [Google Scholar]
- 40. Guan W, Wong S, Zhi N, Qiu J. The genome of human parvovirus B19 can replicate in nonpermissive cells with the help of adenovirus genes and produces infectious virus. J Virol 2009; 83: 9541–9553. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41. Rieder F, Kessler SP, West GA, Bhilocha S, de la Motte C, Sadler TM, Gopalan B, Stylianou E, Fiocchi C. Inflammation‐induced endothelial‐to‐mesenchymal transition: a novel mechanism of intestinal fibrosis. Am J Pathol 2011; 179: 2660–2673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Kindermann I, Kindermann M, Kandolf R, Klingel K, Bultmann B, Muller T, Lindinger A, Bohm M. Predictors of outcome in patients with suspected myocarditis. Circulation 2008; 118: 639–648. [DOI] [PubMed] [Google Scholar]
- 43. Frustaci A, Chimenti C, Calabrese F, Pieroni M, Thiene G, Maseri A. Immunosuppressive therapy for active lymphocytic myocarditis: virological and immunologic profile of responders versus nonresponders. Circulation 2003; 107: 857–863. [DOI] [PubMed] [Google Scholar]
- 44. Frustaci A, Russo MA, Chimenti C. Randomized study on the efficacy of immunosuppressive therapy in patients with virus‐negative inflammatory cardiomyopathy: the TIMIC study. Eur Heart J 2009; 30: 1995–2002. [DOI] [PubMed] [Google Scholar]
- 45. Escher F, Kuhl U, Lassner D, Poller W, Westermann D, Pieske B, Tschope C, Schultheiss HP. Long‐term outcome of patients with virus‐negative chronic myocarditis or inflammatory cardiomyopathy after immunosuppressive therapy. Clin Res Cardiol 2016; 105: 1011–1020. [DOI] [PubMed] [Google Scholar]
- 46. Kuhl U, Pauschinger M, Seeberg B, Lassner D, Noutsias M, Poller W, Schultheiss HP. Viral persistence in the myocardium is associated with progressive cardiac dysfunction. Circulation 2005; 112: 1965–1970. [DOI] [PubMed] [Google Scholar]
- 47. Verdonschot J, Hazebroek M, Merken J, Debing Y, Dennert R, Brunner‐La Rocca HP, Heymans S. Relevance of cardiac parvovirus B19 in myocarditis and dilated cardiomyopathy: review of the literature. Eur J Heart Fail 2016; 18: 1430–1441. [DOI] [PubMed] [Google Scholar]
- 48. Takahashi Y, Sawada G, Kurashige J, Matsumura T, Uchi R, Ueo H, Ishibashi M, Takano Y, Akiyoshi S, Iwaya T, Eguchi H, Sudo T, Sugimachi K, Yamamoto H, Doki Y, Mori M, Mimori K. Tumor‐derived tenascin‐C promotes the epithelial‐mesenchymal transition in colorectal cancer cells. Anticancer Res 2013; 33: 1927–1934. [PubMed] [Google Scholar]
- 49. Brutsaert DL. Cardiac endothelial‐myocardial signaling: its role in cardiac growth, contractile performance, and rhythmicity. Physiol Rev 2003; 83: 59–115. [DOI] [PubMed] [Google Scholar]
- 50. Abbadi D, Laroumanie F, Bizou M, Pozzo J, Daviaud D, Delage C, Calise D, Gaits‐Iacovoni F, Dutaur M, Tortosa F, Renaud‐Gabardos E, Douin‐Echinard V, Prats AC, Roncalli J, Parini A, Pizzinat N. Local production of tenascin‐C acts as a trigger for monocyte/macrophage recruitment that provokes cardiac dysfunction. Cardiovasc Res 2018; 114: 123–137 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Telbivudine pretreatment of parvovirus B19‐infected endothelial cells reduces the pro‐apoptotic potential of parvovirus B19‐infected endothelial cells upon co‐culture in HL‐1 cardiomyocytes. A. Experimental design: HMEC‐1 were 24 h after plating, treated with/out Telbivudine for 2 h, next infected with 10 000 viral particles/cell and collected after 24 h for co‐culture on DiO‐labelled HL‐1 cells, which were plated 24 h before. After 24 h of co‐culture, cells were collected for the anaylsis of DiO+Annexin V/7AAD‐ cells by flow cytometry. Bar graphs represent the mean ± SEM of B. DiO+Annexin V/7AAD‐ HL‐1 cells (% gated), with n=6/group and ***p<0.0005 and ****p<0.0001.