

Abstract
Advancement in cell culture protocols, multidisciplinary research approach, and the need of clinical implication to reconstruct damaged or diseased tissues has led to the establishment of three-dimensional (3D) test systems for regeneration and repair. Regenerative therapies, including dental tissue engineering, have been pursued as a new prospect to repair and rebuild the diseased/lost oral tissues. Interactions between the different cell types, growth factors, and extracellular matrix components involved in angiogenesis are vital in the mechanisms of new vessel formation for tissue regeneration. In vitro pre-vascularization is one of the leading scopes in the tissue-engineering field. Vascularization strategies that are associated with co-culture systems have proved that there is communication between different cell types with mutual beneficial effects in vascularization and tissue regeneration in two-dimensional or 3D cultures. Endothelial cells with different cell populations, including osteoblasts, smooth muscle cells, and fibroblasts in a co-culture have shown their ability to advocate pre-vascularization. In this review, a co-culture perspective of human gingival fibroblasts and vascular endothelial cells is discussed with the main focus on vascularization and future perspective of this model in regeneration and repair.
Keywords: Co-culture, Endothelial cells, Gingival fibroblast, Tissue engineering, Vascularization
Introduction
In the nascent field of tissue engineering, it is vital to develop and improve the original structural, functional, and physiological condition of a tissue. To achieve this, the basic principle lies in the isolation, augmentation, and seeding of autologous cells into a suitable matrix/scaffold before in vivo implantation [1]. For a neovessel growth of a tissue (beyond 200 μm/day) [2, 3] in a body, the formation of a long-lasting vascular network is required for ample oxygen and nutrient supply [4] which is also true for tissue engineering applications dealing in angiogenesis related research. The in vitro development of vascularized and clinically relevant tissue engineered constructs that support and maintain the survival of viable implanted tissues is a key challenge in this field [5, 6]. Numerous approaches have been undertaken to engineer vascularized tissues based on either endothelial cells (ECs) and their ability to form new vessels (neoangiogenesis) and with the biologically-derived vessel systems using three-dimensional scaffold or both [7]. To explore the cellular based strategy on how cells react to certain stimuli, many types of single cells have been cultivated in vitro and much has been studied on how these stimuli influence the single cell population as a whole. However, to understand the direct paracrine interaction between ECs with different cell types affecting cell function and gene expression in two- and three-dimensional settings [8], the concept of heterotypic culture system (also known as co-culture system) has been established to address this complex process of vascularization.
Co-culture systems establish excellent atmosphere to study the interaction of heterotypic cell cultures. A range of factors are known to which heterogeneous cell population can be characterised. These include certain cell differentiation factors, various levels of activation and the presence of an enormous variety of signalling molecules in the surroundings. Due to this complexity, it is unfeasible to examine the biomarkers, gene expression or both for individual cells to be expressed in in vivo conditions, which are important for particular phenotype [8].
ECs are the most widely distributed cell type in the human body and forms the inner cellular lining of the entire vascular system [9]. Fibroblasts play an essential role in the angiogenic process through their production of extracellular matrix (ECM) molecules [10] and by the release of essential growth factors such as vascular endothelial growth factor (VEGF) [11], transforming growth factor-β (TGF-β) [12], and platelet-derived growth factor (PDGF) [13]. Following an injury to the tissue, the microcirculation becomes the rostrum for both the inflammatory response and the successive healing reaction to restore functional details of the damaged tissue or organ. To initiate vessel formation locally from the existing vasculature, ECs are activated at the damage sites in the microvasculature. This process of repair occurs in the milieu of ECM component, which was deposited initially by blood vessels and platelets in the surroundings [14]. In addition to this, rapid mobilization of fibroblasts occurs at the site of injury to produce an ECM, which helps in the repair of injured/damaged vasculature [14]. Thus, fibroblasts play an essential role in the angiogenic process by establishing a matrix that is very necessary and favorable for vascular repair [15]. Moreover, pro-angiogenic and anti-angiogenic factors also particularize by the fibroblasts that communicate with ECs to control the pattern and density of blood vessels [16, 17].
The paradigm of regenerative medicine that aims to develop biomaterials and cell-based therapies emerged as an alternative approach to tissue and organ reconstruction and has opened new prospects to the acceleration of the angiogenic response. Multiple cellular interactions are key elements in the microenvironment of co-culture. Despite numerous attempts in co-culturing ECs with different cell population, it is difficult to test all the novel biomaterials in vivo because of the certain challenges including a high diversity of cells differentiation and activation, a wide variety of signaling molecules, biocompatibility issues, and porosity [6]. However, the use of in vitro co-culture systems using the cells from human origin is being investigated to pre-assess and define the promising potential for success of these after implantation [18]. This review describes the cellular interactions between co-cultured human gingival fibroblasts (HGFs) and ECs, and the process of initiating vascularization that is pertinent to study regeneration in a tissue-engineered construct.
Vascular endothelial cells
Endothelial cells are known to be the major cellular resident of the entire vascular system (arteries, veins, and capillaries). They form a continuous lining at the interface between blood and tissue and are present in all blood vessels. Due to its unique strategic position at the interface between the blood and the tissue, it plays a vital role in providing the proper hemostatic balance. Several different EC types have been used for promoting angiogenesis and vasculogenesis in vitro [19–23] and in vivo [18, 24–27]. Among the mature EC types, human umbilical vein ECs (HUVECs) and human dermal microvascular ECs (HDMEC) are the most widely used cells in the tissue culture experiments [28, 29]. Besides its crucial role in providing the lining for the vessel walls, ECs also exhibit certain essential functions. They are known to be involved in the blood coagulation cascade (thrombosis and thrombolysis), platelet–blood vessel interaction, and act as a potential source of growth promoters (PDGF, endothelin-1 (ET-1), thrombin, fibroblast growth factor (FGF-2), and interleukin-1) and inhibitors (heparin sulfates, nitric oxide, TGF-β) [30, 31]. The migratory and proliferative capacity of ECs is regulated by these factors that play a vital role in the regulation of vascular growth. Thus, the endothelial layer can regulate and help in vascular tone and growth [30, 32].
ECs are also involved in the multiphasic angiogenic process involving a series of cellular events that lead to neovascularization [33]. In normal physiological settings, angiogenesis can be a benchmark of wound healing, the menstrual cycle and organ regeneration [34]. Besides normal conditions, certain pathologies also induce angiogenesis, such as metastatic growth, chronic kidney failure, atherosclerosis, diabetic retinopathy and some intraocular neo-vascular disorders [35–37]. In order to develop a functional vascular network, a complex organizational coordination between various cell populations undergoing remarkable changes and exquisitely dependent upon exchanging signals that promote vascular stability and maturation are necessary [38]. VEGF along with its receptors VEGFR1 and VEGFR2 considered being the most important growth factor and regulator, specific for the vascular endothelium and endothelial cell development respectively [30]. Sprouting of ECs from existing vasculature is a dynamic process that involves binding of VEGF to specific ligand moieties on the ECM and interaction with laminin and collagen IV. Maturation and stabilization of the newly formed vasculature require FGF-2, angiopoietins (Ang) 1, ephrinB2, pericytes and collagen IV [34, 39]. In another study [40], the in vitro morphological differentiation of ECs on a reconstituted gel and Matrigel [41] has been studied using basement membrane matrix proteins such as collagen I, IV and laminin. It has been found that ECs deprived of growth factors undergo morphological differentiation with tube formation after 6–12 weeks and are significantly enhanced by culturing on a reconstituted gel composed of basement membrane proteins. Tubule formation does not occur on tissue culture plastic coated with laminin or collagen IV, either alone or in combination, or on an agarose or a collagen I gel. However, ECs cultured on a collagen I gel supplemented with laminin form tubules, while supplementation with collagen IV induces less tubule formation. This was further studied by Stamati et al. [42] who controlled the in vitro clustering of ECs in three-dimensional collagen hydrogels, which are essential in the formation of different sized tubular structures. It was found that by varying the matrix composition and/or having a source of cell signalling angiogenic proteins, ECs supplemented with laminin showed significantly higher expression of α6 integrin and an increase (P < 0.05) in VEGFR2 positive ECs and increased uptake of VEGF which showed essential proangiogenic linkage between these two.
ECs are involved in the production of numerous matricellular proteins belonging to CCN [connective tissue growth factor (CTGF), cysteine-rich 61 (CYR61) and nephroblastoma overexpressed (NOV)] family that affect many diverse biological processes [43, 44]. Among these, CTGF and CYR61 also play prominent roles in the regulation of angiogenic process [45] which are dependent on the induction of TGFβ or FGF-2 [46]. It has been shown in in vitro studies that MDA231 cells produce higher levels of CTGF protein and more CTGF mRNA under hypoxic conditions [47, 48], suggesting a positive role of CTGF at relatively initial stages of an angiogenic cascade. Moreover, CTGF secreted by tumour cells in hypoxic conditions has been shown to up-regulate endothelial matrix metalloproteinases (MMPs) and to down-regulate endothelial tissue inhibitors of MMPs (TIMPs), thereby, stimulating ECM degradation and ECs migration [47]. Similar data have also been found for CYR61 which can also up-regulate the levels of MMPs [49].
Endothelial gene expression has been regulated during tumour angiogenesis by specific microRNAs (miRNAs), such as those of the miR-17-92 cluster. The effect of exosomal miRNAs derived from leukemic cells (K562) on HUVECs has been studied by co-culturing transfected (Cy3-labeled pre-miR-92a) K562 cells with HUVECs [50]. It has been shown that the exosome, derived from K562 cells with enforced miR-92a expression, did not affect the growth of HUVECs but did enhance ECs migration and tube formation indicating a vital role in neoplasia-to-endothelial cell communication [50].
ECs have been used to study the cellular elements that contribute to the formation of mature vasculature. Recently, endothelial/pericyte interactions has been studied [51] by adding vascular pericytes directly to the bead-bound endothelial monolayer. It has been observed that these pericytes formed close communication with the endothelial sprouts, enhancing the sprout number and vessel calibre. Moreover, the Jagged1 expression and Notch signalling are vital for the growth of both ECs and pericytes and may function in their interaction. These results substantially improve bead-capillary sprouting assays and offer an enhanced method of modelling for interactions between the endothelium and the vascular microenvironment [51].
Human gingival fibroblasts
Fibroblasts, the most abundant cells of the stroma are usually characterized by their morphology and their primary role is secretion of the components of the ECM for tissue maintenance and repair [52, 53]. Apart from their role as synthesizers and modifiers of the ECM, fibroblasts have a strong potential to induce an angiogenic response in the culture. They promote tubulogenesis by depositing ECM [54] and regulate the activation and propagation of EC by secreting various soluble angiogenic factors such as VEGF, PDGF, TGF-β1, FGF-2, and nitric oxide [54–56]. Among those, FGF-2 and VEGF are known to be best characterized which together delivers a synergistic angiogenic response. Besides their support in promoting the migration, survival, and proliferation of ECs, they also contribute towards the in vitro capillary formation [36–38] however, there is no specific reliable marker for fibroblasts [39, 40], and gene expression analysis study revealed that fibroblasts are quite different cells, depending on their tissue of origin [41].
Typically, there are three potential sources from where fibroblasts can be harvested in the oral cavity for the regeneration of gingival connective tissue. These include the gingiva [57], the periodontal ligament [58], and the dental pulp [59]. HGFs are the main resident of the gingival lamina propria and are considered as a major cell type in periodontal connective tissues. They are known to contribute towards the pathogenesis of periodontal disease in the inflammatory periodontium by an exuberant secretion of inflammatory mediators, matrix metalloproteinases, and cytokines [60, 61]. HGFs are considered as the most common cell type used for assessing the biocompatibility of implant prosthesis in the orofacial region [62, 63], for populating in vitro models of gingival connective tissues [64], soft tissue constructs [65], to develop physiologically relevant gingiva equivalent that resembles healthy or a neoplastic disease model [66] and can be a source of induced pluripotent stem cells (iPS) [67–71] for periodontal tissue engineering.
HGFs culture in different matrices has been studied that has shown promising results in soft tissue regeneration [72–74] and exhibit greater functional and biochemical activity in vitro such as increased cell adhesion, cell number and total protein count [75]. Mariotti and Cochran [76] compared the growth characteristics and macromolecular synthesis of HGF and human periodontal ligament fibroblast (HPDLF). They reported that in in vitro cell culture, HGF showed higher proliferative rate, total protein content and grew more rapidly than HPDLF. Moreover, the distribution of glycosaminoglycan, hyaluronic acid, and heparin was more dominant in the cellular segment of PDL tissue, which is indicative of fibroblasts heterogeneity [76]. In another study by Yoshino et al. [77], the relationship between mechanical stress and biochemical phenomena on angiogenic stimulator and inhibitor has been studied with HGFs and HPDLFs. It has been shown that when cultured on a flexible substrate, there is an increased production of VEGF by both cells (P < 0.01). Moreover, in vitro angiogenesis assay further revealed the increased tube like structures formed with HPLF (P < 0.01) which was not enhanced with HGFs because of the expression of pigment epithelium-derived factor (PEDF) [77]. Enamel matrix derivative (EMD) are known to enhance wound healing by inducing angiogenesis [78, 79]. Recently, effect of EMD and its mechanism of action on VEGF production has been studied using HGFs. It has been shown that EMD significantly enhanced the production of VEGF in a dose- and time-dependent manner (P < 0.05) which is mediated by ERK, p38 MAPK, and PI3K/Akt signalling pathways. In contrast, treatment of HGFs with antibodies to TGF-β1 or FGF-2 showed significantly lowered EMD-induced VEGF production, whereas exogenous addition of antibodies enhanced VEGF production. These results are suggestive of the proangiogenic activity of EMD in wound healing models which can help in tissue engineered constructs for early vascularization [80].
Co-culture systems in vascularization
Co-cultures are used in numerous fields of biological research and have progressed to the vanguard within recent history. The initial concept of co-culture systems was proposed in the early 1980s as a system for studying cell–cell communication [81] and subsequently, the use of co-culture systems in the field of tissue engineering and in regenerative medicine proceeded rapidly to their use with heterogeneous cell populations in direct and indirect settings (Fig. 1) [82, 83]. Co-culture models of heterogenic cell types more closely represent in vivo conditions in an ex vivo environment than do homotypic cell culture systems.
Fig. 1.
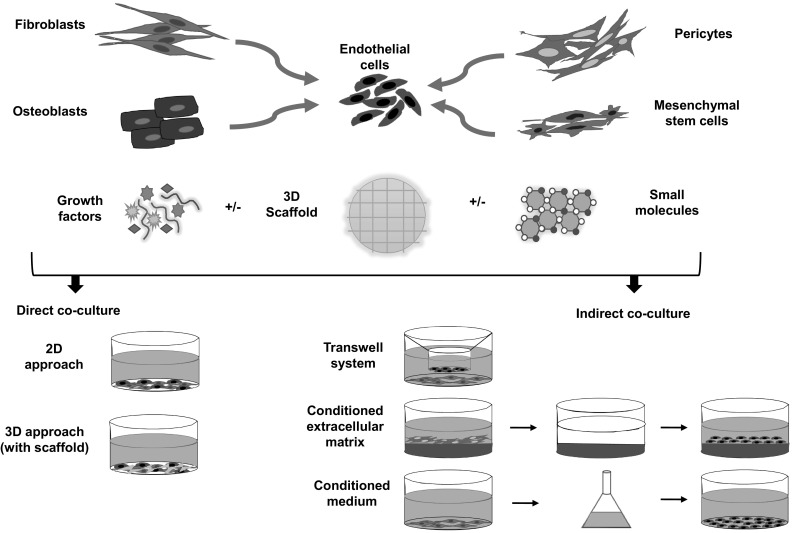
Overview of the cellular interaction between different cell types with endothelial cells in vascularization using a co-culture approach. Endothelial cells have been co-cultured with numerous supporting cells such as fibroblasts, osteoblasts, pericytes or mesenchymal stem cells etc. with or without scaffold through cell signaling and release of cytokines and growth factors or both to induce angiogenesis. These cell–cell communications occur mainly in two forms i.e. direct co-culture approach in two-dimensional or three-dimensional (with scaffold) in which cells interact through secreted signaling factors, cell–cell adhesion and cell–ECM adhesion whereas indirect co-culture approach either in transwell system using semi-permeable membrane, with conditioned medium containing soluble factors released by another cell or with conditioned extracellular matrix (ECM) produced by another cell. In this type, cells are not in direct contact, therefore, cell interaction occurs only through soluble factor signalling
Dynamic remodeling of vascular tissues is essential for the development of the complex angiogenic process. Using currently available model systems, much has already been studied that imitate the formation of vessel-like structures under defined in vitro conditions. These models have been established to identify and differentiate the assay molecules affecting angiogenesis [84]. The most widely functional in vitro and in vivo angiogenesis assays include ECs grown in different gels or mixed ECM gels with ECs [85, 86], microcarrier bead assays [87, 88], aortic rings embedded in gels systems [89–91], the matrigel implant [92], chick chorioallantoic membrane (CAM) [93], and the retinal angiogenesis assay [94]. In all these, ECs grow and differentiate to form loose network-like structures of cords in the presence of angiogenic growth factor e.g. VEGF [84]. Despite this, the possibility of the formation of vessels by ECs in the presence of an appropriate ECM, gel-type EC culture systems has been developed that undervalue the close association and/or involvement of living fibroblasts in an exceptional way [84]. Since the sprouting and maturation of vessels are tortuously tailored by distinct positive and negative regulators that are also shown to express by adjacent cell types, the pertinent nature of co-culture systems is clearly defined. Besides the complexity of this system, they nevertheless deliver a better approximation of a three-dimensional capillary-like association that would be found in vivo [95]. This assumption is advocated by the results of several in vitro studies, primarily dealing in cancer research. For example, Janvier et al. [96], developed a novel three-dimensional co-culture system to study the cellular interactions between fibroblasts and/or PC-3 human prostate adenocarcinoma cells. By co-culturing these cells in a collagen gel, capillary-like structures were evident which also concluded the pivotal role of fibroblasts in cancer neovascularization. In another study, Walter-Yohrling et al. [97] explained the invasion of ECs in a Matrigel-based three-dimensional culture system using a co-culture technique containing an ovarian carcinoma cell line (SKOV-3) with myofibroblasts/10T1/2 cells, suggesting an important cellular interaction between these cells. It has been shown that ECs rapidly form a vascular network within 5 h of culture on Matrigel. Moreover, the same ECs when co-cultured with ovarian carcinoma cell line, formed long, linear projections around the tumour cell cluster when seeded on a monolayer Matrigel within 5 h, which became more evident after 24 h. In contrast, when cultured in the presence of myofibroblasts, ECs adhered to the outside of the tumour cell cluster while myofibroblasts localized within the tumour cell mass. However, 10T1/2 cells (a pluripotent mouse mesenchymal cell line with pericyte like properties) did not establish the same invasive phenotype.
Perivascular cells such as pericytes (mural cells), smooth muscles cells, and adventitial fibroblasts, are coupled with the endothelium of capillaries and are known to be involved in the vessel maturation and stabilization during the later stages of angiogenesis [98]. The co-culture of ECs with these cells in vitro have confirmed that the release of soluble factors such as VEGF/FGF-2/PDGF or the deposition of ECM proteins such as fibronectin stimulate angiogenic processes in ECs such as increased plasminogen activity, proliferation, motility, and cord-like structure formation [99, 100].
Co-culture techniques find diverse applications in regenerative medicine and vascularization for studying natural or synthetic interactions between distinct cell populations. Many studies in the literature have proven success in establishing vascularized structures in co-cultures using biomaterial scaffold [101–104]. Fibroblasts stabilize the level of apoptosis when co-cultured with ECs in spheroid form [54]. In a study by Wenger et al. [54], a three-dimensional spheroidal co-culture system has been developed in vitro consisting of HUVECs and human primary fibroblasts (hFBs) and investigated the effects of human fibroblasts on ECs function with collagen-embedded spheroids. It has been shown that within 48 h of co-cultivation of HUVECs with hFBs, the level of apoptosis was strongly decreased (13%) and collagen-embedded HUVEC spheroids developed numerous lumenized capillary-like sprouts in comparison to solo culture of these cells in spheroids (i.e. HUVEC: 100%, hFBs: 6%). Moreover, it has been observed that due to the heterotypic cell interactions between HUVECs and hFBs within the co-spheroid, the quantification of cumulative sprout length depicts an approximately 35% reduction in ECs sprouting. This co-spheroid model of HUVEC/hFBs is highly suggestive for delivering a preformed capillary network ex vivo and may, therefore, be advantageous for enhancing angiogenesis in in vivo tissue engineering applications.
In another study by Choong et al. [55], polycaprolactone (gelatin-coated and hydroxyapatite-coated substrates) biomaterial surface has been studied in a co-culture setting that supports EC attachment and proliferation for inducing angiogenesis. It has been shown that within 12 days of co-culture between human bone marrow-derived fibroblasts (HBMFs) and human bone marrow endothelial cell line (HBMEC-60), HBMF cell proliferation and differentiation specifically enhanced (P < 0.05) and that this effect was not observed with co-culture with skin fibroblasts. Moreover, after 13 days of in vitro co-culture, tubular structures were evident which proved this co-culture model of ECs with HBMFs could be a promising model for bone vascularization [55].
A scope of co-culture consisting of heterotypic cells may also be utilized for the development of artificial mucosa equivalent. Recently, a pre-vascularized buccal mucosa equivalent has been generated [56] by using primary buccal epithelial cells, fibroblasts and microvascular ECs, in the presence of a native collagen membrane as a scaffold. Formation of dense capillary-like structures was evident within 3 weeks of cultivation with the lumen size of pre-formed blood vessels corresponded to the capillary size in vivo (10–30 μm). Moreover, when compared with a native highly cross-linked collagen membrane, enhanced formation of capillary-like structures was observed due to higher secretion of angiogenic factors such as PDGF, IL-8 and angiopoietin by the cells. These capillary-like structures became functional blood vessels through anastomosis with the host vasculature after implantation in nude mice which signifies the process and success of vascularization in tissue engineered construct [56]. In another study by Zhao et al. [105] using a three-dimensional co-culture collagen gel model for notch signalling pathway in angiogenesis, it has been demonstrated that compared to control group, Jagged1 significantly enhanced the formation of vessel like structures (i.e. more expression of angiogenic growth factors; P < 0.5) within 7 days and became denser after 14 days of co-culture while DAPT treatment attenuated this process. Moreover, it has been shown that VEGFA/B-Notch1/2-Hes1/Hey1-VEGFR1/3 signal axis played an important role which proves that notch signalling is critically involved in vascular development and disease [105]. Initial studies, particularly in angiogenesis, involved the co-culture of ECs with dermal fibroblasts for promoting vascularization in the skin [104]. It has been shown in a study by Guerreiro et al. [106] in which neonatal human dermal fibroblasts embedded in Matrigel plugs when implanted in mice produce collagen and induce angiogenesis from the host vasculature. It has been observed that fibroblasts could recruit ECs to vascularize the implanted matrix, which was further colonized after 1 week by murine functional blood vessels. Furthermore, levels of haemoglobin were significantly higher (P < 0.05 vs. control) when compared with the control matrix (implanted without fibroblasts) in which vessel formation was not observed. The fibroblasts implanted within Matrigel has been shown to increase ECs population (stained positive for CD 31 and for CD 34) along with the enhanced production of collagen which suggests that these cells take part in the recruitment of ECs in vivo. Hence, the interaction between fibroblasts and ECs has an essential role in the process of neovascularization.
Co-culture of human gingival fibroblasts and vascular endothelial cells
Many studies have been conducted to elucidate the choice of cell type(s) for inducing angiogenesis [107–109]. ECs have been co-cultured with numerous supporting cells such as smooth muscle cells [104], retinal cells [110], monocytes [111], mesenchymal stem cells [112], osteoblasts [113], epiCs [114], fibroblasts [104], and pericytes [115] etc. to induce angiogenesis (Fig. 1). These cell types have been shown to support angiogenesis via paracrine signaling or cell–cell interaction [104]. In physiological settings, ECs interact with different cell types in order to survive and proliferate [104]. However, in monoculture, ECs lose its ability to self-assemble into tube-like structures [116]. For example, Hofmann et al. [117] observed that in a monoculture, HUVECs died rapidly after 1 week. However, a long-term (6 weeks) co-culture of ECs with human osteoblasts in the presence of polyurethane scaffolds and platelet-rich plasma stimulated HUVEC maturation, expression of collagen type IV, and impressive formation of multiple tube-like structures (positive for CD31 and vWF; P < 0.05) which strongly signifies the need of co-cultures for vascular tissue engineering. Different experimental protocols have been instituted to explore fibroblast–EC interactions in culture systems [16, 17, 118–121]. In these culture systems, tube-like structures develop from single cell populations of vascular endothelial cells.
Since angiogenesis in many normal physiological processes depends on the activation and migration of fibroblasts, the in vitro cocultivation of fibroblasts and ECs is a coherent approach to augment the quality of vessel-like structures. Indeed, one of the most reliable in vitro angiogenesis test systems showed significant in vitro tubule formation in a monolayer co-culture of human diploid fibroblast and HUVECs [24, 122]. Particularly, no artificial or additional supplements were added, and during the 14-day period, vessel-like structures with true luminal development happened in multi-layered areas formed by the heterogeneous cell types. Moreover, the fibroblast’s crucial role was established, and the cell–cell interaction under these conditions allowed experimental analysis with comparative ease.
By optimizing the culture conditions, co-culture of HUVECs and HGFs can potentially be used in the regeneration of a vascularized gingival connective tissue by producing a tissue engineered construct. In a recent study [123], a co-culture of HUVECs with HGFs was used in a perfused degradable/polar/hydrophobic/ionic polyurethane (D-PHI) scaffold to ascertain the possibility of vessel formation in an in vitro bioreactor system. It has been demonstrated that over the 28 days of co-culture in the presence of scaffold with a cell seeding density of at least 80,000 cells/scaffold (P < 0.05) in a 50/50 mix of HUVEC and HGF media (by volume) showed increased cell growth, enhanced production of angiogenic factors (VEGF and FGF-2), clustering of HUVEC as well as decreased differentiation of myofibroblasts (P < 0.05 for days 14 and 28). It is also proved that these culture conditions coupled with pro-angiogenic effects are likely to be significant in the formation of a highly-vascularized tissue-engineered construct not only for regeneration of gingival lamina propria but also possibly for other soft tissues [123].
Fibroblast supports and modulates EC migration, viability, and network formation in a three-dimensional tissue-like stromal milieu. Kunz-Schughart et al. [84] studied this unique property by using human skin fibroblasts and ECs from HUVECs in distinct spheroid co-culture systems. A significant rise in EC clustering and vessel-like formation was evident when fibroblast to-EC ratios of 40 to 1 and 4 to 1 in a three-dimensional spheroid system were used. However, if this ratio is opposite in the spheroid (i.e. more ECs than fibroblasts), a substantial loss of the EC population as early as 3 days of culture was observed. Cheung et al. [123] also observed the same trends while co-culturing HGF and HUVEC in a ratio of 1:2 (HUVEC:HGF). It has been shown that greater proportion of HGF in the presence of D-PHI exhibit more cell growth while HUVEC favors capillary formation and more chances of survival at this lower concentration.
The ECM also attains a significant role in angiogenesis as it facilitates mechanical transduction to ECs with proteins from integrin family such as β1-integrins and αvβ3 integrin [124]. Due to the presence of TGF-β1, VEGF, and FGF-2, it is considered as a pool for angiogenic factors [125]. Collagen type I and III have been shown to stimulate EC tube formation and stabilize blood vessels [126], and it is revealed that ECs, when cultured on collagen-rich ECM, can develop into capillaries in 48 h [124]. Since fibroblasts mostly produce collagen (type 1 collagen specifically for HGFs) in addition to the angiogenic factors, this may well clarify the augmented angiogenic effects in EC–fibroblast co-cultures.
The use of HPDLF and HGF in the presence of ECs also proved to have a beneficial effect in regeneration. This was observed in a recent study [127] in which the effect of HUVEC on the migration of HPDLF and HGF has been investigated using a transwell co-culture model imitating the healing and regeneration of periodontal structures. It has been shown that the integrated option density of both the cells were significantly higher than each of the single culture (P < 0.01). Moreover, within 24 h of co-culture, the HGF promote increased horizontal migration (P < 0.01). However, in 7 days of vertical migration and wound healing model using scratch wound and round glass sheet assay, this effect was more pronounced on HPDLFs (P < 0.01). This co-culture model signifies the importance of ECs for vascularization and tissue engineering applications.
Future scenario and its role in tissue engineering and regenerative medicine
The advent of the multidisciplinary field of regenerative medicine, of which tissue engineering is a major component, has opened novel approaches to the acceleration of the healing response. Numerous types of single-cell in vitro culture systems have been developed to understand and study how cells react to certain stimuli. Using these systems, much is recognized how this influences the single-cell population as a whole. However, to study a direct interaction between two different cell types/soluble compounds (released by one cell and taken up by the neighbouring/surrounding cell) affecting the cell behaviour, genetic phenotype and the ability of ECs to perform angiogenesis is one of the present goals of the emerging studies encompassing two-dimensional and three-dimensional cell co-cultures.
One of the most definite tools to study the cellular interaction between heterogeneous populations is the co-culture system. However, it seems to be troubled with many technical issues that must be resolved before any significant data can be collected. Many advancements have been made in the culture and in in vitro evaluation of the mechanisms of formation of vascularization in the presence of both ECs and other relevant cells from the target tissue. These studies are beginning to show the complexity of the interaction and cell–cell communication taking place between the two heterogeneous cell types.
Different studies in the literature have been done using the co-culture of ECs with osteoblasts, smooth muscle cells, and keratinocytes in two-dimensional and three-dimensional settings. However, there is a dearth of literature on the co-culture of HGF and EC in the presence of biomaterial scaffold. So, future studies will be planned to use the scaffold/biomaterial approach for better understanding the cell–cell interaction between these two cells, and information from these studies should help to design and prepare new biomaterials for increasingly better survival and to function after implantation in vivo. It will help in the availability of a substrate for the establishment of the three-dimensional vascularization in tissue engineered construct.
Vascularization strategies in cell-based periodontal tissue engineering depend on optimal culture conditions. However, data on the optimal culture conditions for such co-cultures are deficient, and more precisely data that take both regenerative and angiogenic outcome parameters into account. Given the available literature on co-culture systems of HGF and ECs, it is notable that no consensus exists on the optimal conditions for such a co-culture system. Therefore, it is important to define the suitable medium for co-culture of these cells in the two-dimensional and three-dimensional environment. Establishing the optimal culture conditions for this co-culture system may be useful for improving angiogenesis in in vivo tissue engineering applications.
Even though all cell types in the living organism experience some form of mechano-stimulation, it is a fact that most culture systems are still established under static environment. However, in the human body, practically every cell type is subjected to some form of mechanical stimulation, whether compression, tension and/or shear forces. These conditions can be best controlled and monitored in bioreactors. So, future studies should be designed with the incorporation of bioreactors to better understand the co-culture mechanism between these two cells in relation to their natural counterpart and its successful application in clinical settings.
Conclusion
Tissue engineering in the context of regenerative dentistry has been acknowledged as one of the most important topics in dentistry in the twenty-first century. These progressing interdisciplinary fields utilize principles of engineering and life sciences to reproduce replicates, typically composed of biological and synthetic components that are meant to restore, maintain, or improve tissue function. Cellular strategies establish a favorable environment to achieve vascularization of tissue engineering constructs and precluding the troublesome side effects of delivery of growth factors, cytokines, hormones or other bioactive molecules. Methods for EC transplantation depend on the use of support cells to ensure the stabilization and maturation of newly formed vasculature. However, the clinical use of heterogeneous cell types would entail different isolation procedures, resulting in an increased morbidity of donor sites. Vascular tissue engineering aims at innovative technological approaches to be developed, as different cell populations necessitate the use of complex interactions to organize vascularization processes in very distinct organs in the human body. Co-culture techniques find multiple applications not only in regenerative medicine but also in vascularization strategies for studying natural or synthetic interactions between different cell populations. It is usually presumed that co-culture systems are more advantageous in vascularization strategies compared with monoculture in both in vitro and in vivo settings. Co-culture of HGFs with ECs has been proposed as a valuable pre-vascularization strategy in gingival tissue engineering. The flexibility of this system for enhanced graft vascularization will be explored under in vivo settings in future trials.
Acknowledgements
This review was supported by Universiti Sains Malaysia through the Research University Grant (RU-1001/PPSG/812168).
Conflict of interest
The authors have no financial conflicts of interest.
Ethical statement
There are no animal experiments carried out for this review article.
References
- 1.Bartold PM, McCulloch CA, Narayanan AS, Pitaru S. Tissue engineering: a new paradigm for periodontal regeneration based on molecular and cell biology. Periodontology. 2000;2000:253–269. doi: 10.1034/j.1600-0757.2000.2240113.x. [DOI] [PubMed] [Google Scholar]
- 2.Ausprunk DH, Folkman J. Migration and proliferation of endothelial cells in preformed and newly formed blood vessels during tumor angiogenesis. Microvasc Res. 1977;14:53–65. doi: 10.1016/0026-2862(77)90141-8. [DOI] [PubMed] [Google Scholar]
- 3.Rouwkema J, Rivron NC, van Blitterswijk CA. Vascularization in tissue engineering. Trends Biotechnol. 2008;26:434–441. doi: 10.1016/j.tibtech.2008.04.009. [DOI] [PubMed] [Google Scholar]
- 4.Fan H, Zeng X, Wang X, Zhu R, Pei G. Efficacy of prevascularization for segmental bone defect repair using β-tricalcium phosphate scaffold in rhesus monkey. Biomaterials. 2014;35:7407–7415. doi: 10.1016/j.biomaterials.2014.05.035. [DOI] [PubMed] [Google Scholar]
- 5.Rouwkema J, Koopman B, Blitterswijk C, Dhert W, Malda J. Supply of nutrients to cells in engineered tissues. Biotechnol Genet Eng Rev. 2010;26:163–178. doi: 10.5661/bger-26-163. [DOI] [PubMed] [Google Scholar]
- 6.Ikada Y. Challenges in tissue engineering. J R Soc Interface. 2006;3:589–601. doi: 10.1098/rsif.2006.0124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Novosel EC, Kleinhans C, Kluger PJ. Vascularization is the key challenge in tissue engineering. Adv Drug Deliv Rev. 2011;63:300–311. doi: 10.1016/j.addr.2011.03.004. [DOI] [PubMed] [Google Scholar]
- 8.Kirkpatrick CJ, Fuchs S, Unger RE. Co-culture systems for vascularization—learning from nature. Adv Drug Deliv Rev. 2011;63:291–299. doi: 10.1016/j.addr.2011.01.009. [DOI] [PubMed] [Google Scholar]
- 9.Cines DB, Pollak ES, Buck CA, Loscalzo J, Zimmerman GA, McEver RP, et al. Endothelial cells in physiology and in the pathophysiology of vascular disorders. Blood. 1998;91:3527–3561. [PubMed] [Google Scholar]
- 10.Newman AC, Nakatsu MN, Chou W, Gershon PD, Hughes CCW. The requirement for fibroblasts in angiogenesis: fibroblast-derived matrix proteins are essential for endothelial cell lumen formation. Mol Biol Cell. 2011;22:3791–3800. doi: 10.1091/mbc.E11-05-0393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kellouche S, Mourah S, Bonnefoy A, Schoëvaert D, Podgorniak M-P, Calvo F, et al. Platelets, thrombospondin-1 and human dermal fibroblasts cooperate for stimulation of endothelial cell tubulogenesis through vegf and pai-1 regulation. Exp Cell Res. 2007;313:486–499. doi: 10.1016/j.yexcr.2006.10.023. [DOI] [PubMed] [Google Scholar]
- 12.Paunescu V, Bojin FM, Tatu CA, Gavriliuc OI, Rosca A, Gruia AT, et al. Tumour-associated fibroblasts and mesenchymal stem cells: more similarities than differences. J Cell Mol Med. 2011;15:635–646. doi: 10.1111/j.1582-4934.2010.01044.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Antoniades HN, Galanopoulos T, Neville-Golden J, Kiritsy CP, Lynch SE. Injury induces in vivo expression of platelet-derived growth factor (pdgf) and pdgf receptor mrnas in skin epithelial cells and pdgf mrna in connective tissue fibroblasts. Proc Natl Acad Sci U S A. 1991;88:565–569. doi: 10.1073/pnas.88.2.565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Singer AJ, Clark RAF. Cutaneous wound healing. N Engl J Med. 1999;341:738–746. doi: 10.1056/NEJM199909023411006. [DOI] [PubMed] [Google Scholar]
- 15.Enzerink A, Vaheri A. Fibroblast activation in vascular inflammation. J Thromb Haemost. 2011;9:619–626. doi: 10.1111/j.1538-7836.2011.04209.x. [DOI] [PubMed] [Google Scholar]
- 16.Hudon V, Berthod F, Black AF, Damour O, Germain L, Auger FA. A tissue-engineered endothelialized dermis to study the modulation of angiogenic and angiostatic molecules on capillary-like tube formation in vitro. Br J Dermatol. 2003;148:1094–1104. doi: 10.1046/j.1365-2133.2003.05298.x. [DOI] [PubMed] [Google Scholar]
- 17.Black AF, Hudon V, Damour O, Germain L, Auger FA. A novel approach for studying angiogenesis: a human skin equivalent with a capillary-like network. Cell Biol Toxicol. 1999;15:81–90. doi: 10.1023/A:1007541713398. [DOI] [PubMed] [Google Scholar]
- 18.Staton CA, Reed MWR, Brown NJ. A critical analysis of current in vitro and in vivo angiogenesis assays. Int J Exp Pathol. 2009;90:195–221. doi: 10.1111/j.1365-2613.2008.00633.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bouis D, Hospers GA, Meijer C, Molema G, Mulder NH. Endothelium in vitro: a review of human vascular endothelial cell lines for blood vessel-related research. Angiogenesis. 2001;4:91–102. doi: 10.1023/A:1012259529167. [DOI] [PubMed] [Google Scholar]
- 20.Vailhe B, Vittet D, Feige JJ. In vitro models of vasculogenesis and angiogenesis. Lab Invest. 2001;81:439–452. doi: 10.1038/labinvest.3780252. [DOI] [PubMed] [Google Scholar]
- 21.Morin KT, Tranquillo RT. In vitro models of angiogenesis and vasculogenesis in fibrin gel. Exp Cell Res. 2013;319:2409–2417. doi: 10.1016/j.yexcr.2013.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Heiss M, Hellström M, Kalén M, May T, Weber H, Hecker M, et al. Endothelial cell spheroids as a versatile tool to study angiogenesis in vitro. FASEB J. 2015;29:3076–3084. doi: 10.1096/fj.14-267633. [DOI] [PubMed] [Google Scholar]
- 23.Zheng Y, Chen J, Craven M, Choi NW, Totorica S, Diaz-Santana A, et al. In vitro microvessels for the study of angiogenesis and thrombosis. Proc Natl Acad Sci U S A. 2012;109:9342–9347. doi: 10.1073/pnas.1201240109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Donovan D, Brown NJ, Bishop ET, Lewis CE. Comparison of three in vitro human ‘angiogenesis’ assays with capillaries formed in vivo. Angiogenesis. 2001;4:113–121. doi: 10.1023/A:1012218401036. [DOI] [PubMed] [Google Scholar]
- 25.Ribatti D, Vacca A. Models for studying angiogenesis in vivo. Int J Biol Markers. 1999;14:207–213. doi: 10.1177/172460089901400403. [DOI] [PubMed] [Google Scholar]
- 26.Fràter-Schröder M, Risau W, Hallmann R, Gautschi P, Böhlen P. Tumor necrosis factor type alpha, a potent inhibitor of endothelial cell growth in vitro, is angiogenic in vivo. Proc Natl Acad Sci U S A. 1987;84:5277–5281. doi: 10.1073/pnas.84.15.5277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cao Y, Linden P, Farnebo J, Cao R, Eriksson A, Kumar V, et al. Vascular endothelial growth factor c induces angiogenesis in vivo. Proc Natl Acad Sci U S A. 1998;95:14389–14394. doi: 10.1073/pnas.95.24.14389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Unger RE, Sartoris A, Peters K, Motta A, Migliaresi C, Kunkel M, et al. Tissue-like self-assembly in cocultures of endothelial cells and osteoblasts and the formation of microcapillary-like structures on three-dimensional porous biomaterials. Biomaterials. 2007;28:3965–3976. doi: 10.1016/j.biomaterials.2007.05.032. [DOI] [PubMed] [Google Scholar]
- 29.Bidarra SJ, Barrias CC, Barbosa MA, Soares R, Amedee J, Granja PL. Phenotypic and proliferative modulation of human mesenchymal stem cells via crosstalk with endothelial cells. Stem Cell Res. 2011;7:186–197. doi: 10.1016/j.scr.2011.05.006. [DOI] [PubMed] [Google Scholar]
- 30.Rajendran P, Rengarajan T, Thangavel J, Nishigaki Y, Sakthisekaran D, Sethi G, et al. The vascular endothelium and human diseases. Int J Biol Sci. 2013;9:1057–1069. doi: 10.7150/ijbs.7502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Rudijanto A. The role of vascular smooth muscle cells on the pathogenesis of atherosclerosis. Acta Med Indones. 2007;39:86–93. [PubMed] [Google Scholar]
- 32.Verhamme P, Hoylaerts MF. The pivotal role of the endothelium in haemostasis and thrombosis. Acta Clin Belg. 2006;61:213–219. doi: 10.1179/acb.2006.036. [DOI] [PubMed] [Google Scholar]
- 33.Ucuzian AA, Gassman AA, East AT, Greisler HP. Molecular mediators of angiogenesis. J Burn Care Res. 2010;31:158. doi: 10.1097/BCR.0b013e3181c7ed82. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Adams RH, Alitalo K. Molecular regulation of angiogenesis and lymphangiogenesis. Nat Rev Mol Cell Biol. 2007;8:464–478. doi: 10.1038/nrm2183. [DOI] [PubMed] [Google Scholar]
- 35.Folkman J. Angiogenesis: an organizing principle for drug discovery? Nat Rev Drug Discov. 2007;6:273–286. doi: 10.1038/nrd2115. [DOI] [PubMed] [Google Scholar]
- 36.Chung AS, Ferrara N. Developmental and pathological angiogenesis. Annu Rev Cell Dev Biol. 2011;27:563–584. doi: 10.1146/annurev-cellbio-092910-154002. [DOI] [PubMed] [Google Scholar]
- 37.Zoccali C. The endothelium as a target in renal diseases. J Nephrol. 2007;20:S39–S44. [PubMed] [Google Scholar]
- 38.Gale NW, Yancopoulos GD. Growth factors acting via endothelial cell-specific receptor tyrosine kinases: vegfs, angiopoietins, and ephrins in vascular development. Genes Dev. 1999;13:1055–1066. doi: 10.1101/gad.13.9.1055. [DOI] [PubMed] [Google Scholar]
- 39.Michiels C. Endothelial cell functions. J Cell Physiol. 2003;196:430–443. doi: 10.1002/jcp.10333. [DOI] [PubMed] [Google Scholar]
- 40.Kubota Y, Kleinman HK, Martin GR, Lawley TJ. Role of laminin and basement membrane in the morphological differentiation of human endothelial cells into capillary-like structures. J Cell Biol. 1988;107:1589–1598. doi: 10.1083/jcb.107.4.1589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Grant DS, Tashiro K, Segui-Real B, Yamada Y, Martin GR, Kleinman HK. Two different laminin domains mediate the differentiation of human endothelial cells into capillary-like structures in vitro. Cell. 1989;58:933–943. doi: 10.1016/0092-8674(89)90945-8. [DOI] [PubMed] [Google Scholar]
- 42.Stamati K, Priestley JV, Mudera V, Cheema U. Laminin promotes vascular network formation in 3d in vitro collagen scaffolds by regulating vegf uptake. Exp Cell Res. 2014;327:68–77. doi: 10.1016/j.yexcr.2014.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Bornstein P. Matricellular proteins: an overview. J Cell Commun Signal. 2009;3:163–165. doi: 10.1007/s12079-009-0069-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Bornstein P, Sage EH. Matricellular proteins: extracellular modulators of cell function. Curr Opin Cell Biol. 2002;14:608–616. doi: 10.1016/S0955-0674(02)00361-7. [DOI] [PubMed] [Google Scholar]
- 45.Brigstock DR. Regulation of angiogenesis and endothelial cell function by connective tissue growth factor (ctgf) and cysteine-rich 61 (cyr61) Angiogenesis. 2002;5:153–165. doi: 10.1023/A:1023823803510. [DOI] [PubMed] [Google Scholar]
- 46.Holbourn KP, Acharya KR, Perbal B. The ccn family of proteins: structure–function relationships. Trends Biochem Sci. 2008;33:461–473. doi: 10.1016/j.tibs.2008.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kondo S, Kubota S, Shimo T, Nishida T, Yosimichi G, Eguchi T, et al. Connective tissue growth factor increased by hypoxia may initiate angiogenesis in collaboration with matrix metalloproteinases. Carcinogenesis. 2002;23:769–776. doi: 10.1093/carcin/23.5.769. [DOI] [PubMed] [Google Scholar]
- 48.Shimo T, Kubota S, Kondo S, Nakanishi T, Sasaki A, Mese H, et al. Connective tissue growth factor as a major angiogenic agent that is induced by hypoxia in a human breast cancer cell line. Cancer Lett. 2001;174:57–64. doi: 10.1016/S0304-3835(01)00683-8. [DOI] [PubMed] [Google Scholar]
- 49.Chen CC, Mo FE, Lau LF. The angiogenic factor cyr61 activates a genetic program for wound healing in human skin fibroblasts. J Biol Chem. 2001;276:47329–47337. doi: 10.1074/jbc.M107666200. [DOI] [PubMed] [Google Scholar]
- 50.Umezu T, Ohyashiki K, Kuroda M, Ohyashiki JH. Leukemia cell to endothelial cell communication via exosomal mirnas. Oncogene. 2013;32:2747–2755. doi: 10.1038/onc.2012.295. [DOI] [PubMed] [Google Scholar]
- 51.Tattersall IW, Du J, Cong Z, Cho BS, Klein AM, Dieck CL, et al. In vitro modeling of endothelial interaction with macrophages and pericytes demonstrates notch signaling function in the vascular microenvironment. Angiogenesis. 2016;19:201–215. doi: 10.1007/s10456-016-9501-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Hinz B. Formation and function of the myofibroblast during tissue repair. J Investig Dermatol. 2007;127:526–537. doi: 10.1038/sj.jid.5700613. [DOI] [PubMed] [Google Scholar]
- 53.Wipff PJ, Hinz B. Myofibroblasts work best under stress. J Bodyw Mov Ther. 2009;13:121–127. doi: 10.1016/j.jbmt.2008.04.031. [DOI] [PubMed] [Google Scholar]
- 54.Wenger A, Kowalewski N, Stahl A, Mehlhorn AT, Schmal H, Stark GB, et al. Development and characterization of a spheroidal coculture model of endothelial cells and fibroblasts for improving angiogenesis in tissue engineering. Cells Tissues Organs. 2005;181:80–88. doi: 10.1159/000091097. [DOI] [PubMed] [Google Scholar]
- 55.Choong CS, Hutmacher DW, Triffitt JT. Co-culture of bone marrow fibroblasts and endothelial cells on modified polycaprolactone substrates for enhanced potentials in bone tissue engineering. Tissue Eng. 2006;12:2521–2531. doi: 10.1089/ten.2006.12.2521. [DOI] [PubMed] [Google Scholar]
- 56.Heller M, Frerick-Ochs EV, Bauer HK, Schiegnitz E, Flesch D, Brieger J, et al. Tissue engineered pre-vascularized buccal mucosa equivalents utilizing a primary triculture of epithelial cells, endothelial cells and fibroblasts. Biomaterials. 2016;77:207–215. doi: 10.1016/j.biomaterials.2015.10.073. [DOI] [PubMed] [Google Scholar]
- 57.Jin C, Ren LF, Ding HZ, Shi GS, Lin HS, Zhang F. Enhanced attachment, proliferation, and differentiation of human gingival fibroblasts on titanium surface modified with biomolecules. J Biomed Mater Res B Appl Biomater. 2012;100:2167–2177. doi: 10.1002/jbm.b.32784. [DOI] [PubMed] [Google Scholar]
- 58.Giannopoulou C, Cimasoni G. Functional characteristics of gingival and periodontal ligament fibroblasts. J Dent Res. 1996;75:895–902. doi: 10.1177/00220345960750030601. [DOI] [PubMed] [Google Scholar]
- 59.Buurma B, Gu K, Rutherford RB. Transplantation of human pulpal and gingival fibroblasts attached to synthetic scaffolds. Eur J Oral Sci. 1999;107:282–289. doi: 10.1046/j.0909-8836.1999.eos107408.x. [DOI] [PubMed] [Google Scholar]
- 60.Moharamzadeh K, Brook I, Van Noort R, Scutt A, Thornhill M. Tissue-engineered oral mucosa: a review of the scientific literature. J Dent Res. 2007;86:115–124. doi: 10.1177/154405910708600203. [DOI] [PubMed] [Google Scholar]
- 61.Daghigh F, Borghaei RC, Thornton RD, Bee JH. Human gingival fibroblasts produce nitric oxide in response to proinflammatory cytokines. J Periodontol. 2002;73:392–400. doi: 10.1902/jop.2002.73.4.392. [DOI] [PubMed] [Google Scholar]
- 62.Jin G, Prabhakaran MP, Ramakrishna S. Stem cell differentiation to epidermal lineages on electrospun nanofibrous substrates for skin tissue engineering. Acta Biomater. 2011;7:3113–3122. doi: 10.1016/j.actbio.2011.04.017. [DOI] [PubMed] [Google Scholar]
- 63.Ma Q, Mei S, Ji K, Zhang Y, Chu PK. Immobilization of ag nanoparticles/fgf-2 on a modified titanium implant surface and improved human gingival fibroblasts behavior. J Biomed Mater Res A. 2011;98:274–286. doi: 10.1002/jbm.a.33111. [DOI] [PubMed] [Google Scholar]
- 64.Blackwood KA, McKean R, Canton I, Freeman CO, Franklin KL, Cole D, et al. Development of biodegradable electrospun scaffolds for dermal replacement. Biomaterials. 2008;29:3091–3104. doi: 10.1016/j.biomaterials.2008.03.037. [DOI] [PubMed] [Google Scholar]
- 65.Chung TW, Wang SS, Wang YZ, Hsieh CH, Fu E. Enhancing growth and proliferation of human gingival fibroblasts on chitosan grafted poly(epsilon-caprolactone) films is influenced by nano-roughness chitosan surfaces. J Mater Sci Mater Med. 2009;20:397–404. doi: 10.1007/s10856-008-3586-z. [DOI] [PubMed] [Google Scholar]
- 66.Buskermolen JK, Reijnders CM, Spiekstra SW, Steinberg T, Kleverlaan CJ, Feilzer AJ, et al. Development of a full-thickness human gingiva equivalent constructed from immortalized keratinocytes and fibroblasts. Tissue Eng Part C Methods. 2016;22:781–791. doi: 10.1089/ten.tec.2016.0066. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Egusa H, Okita K, Kayashima H, Yu G, Fukuyasu S, Saeki M, et al. Gingival fibroblasts as a promising source of induced pluripotent stem cells. PLoS One. 2010;5:e12743. doi: 10.1371/journal.pone.0012743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Fournier BP, Ferre FC, Couty L, Lataillade JJ, Gourven M, Naveau A, et al. Multipotent progenitor cells in gingival connective tissue. Tissue Eng Part A. 2010;16:2891–2899. doi: 10.1089/ten.tea.2009.0796. [DOI] [PubMed] [Google Scholar]
- 69.Fournier BP, Larjava H, Häkkinen L. Gingiva as a source of stem cells with therapeutic potential. Stem Cells Dev. 2013;22:3157–3177. doi: 10.1089/scd.2013.0015. [DOI] [PubMed] [Google Scholar]
- 70.Wang F, Yu M, Yan X, Wen Y, Zeng Q, Yue W, et al. Gingiva-derived mesenchymal stem cell-mediated therapeutic approach for bone tissue regeneration. Stem Cells Dev. 2011;20:2093–2102. doi: 10.1089/scd.2010.0523. [DOI] [PubMed] [Google Scholar]
- 71.Ferré FC, Larjava H, Loison-Robert LS, Berbar T, Owen GR, Berdal A, et al. Formation of cartilage and synovial tissue by human gingival stem cells. Stem Cells Dev. 2014;23:2895–2907. doi: 10.1089/scd.2013.0547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Maia LP, Novaes AB, Jr, Souza SL, Grisi MF, Taba M, Palioto DB. In vitro evaluation of acellular dermal matrix as a three-dimensional scaffold for gingival fibroblasts seeding. J Periodontol. 2011;82:293–301. doi: 10.1902/jop.2010.100121. [DOI] [PubMed] [Google Scholar]
- 73.Rodrigues AZ, Oliveira PT, Novaes AB, Jr, Maia LP, Souza SL, Palioto DB. Evaluation of in vitro human gingival fibroblast seeding on acellular dermal matrix. Braz Dent J. 2010;21:179–189. doi: 10.1590/S0103-64402010000300001. [DOI] [PubMed] [Google Scholar]
- 74.Jhaveri HM, Chavan MS, Tomar GB, Deshmukh VL, Wani MR, Miller PD., Jr Acellular dermal matrix seeded with autologous gingival fibroblasts for the treatment of gingival recession: a proof-of-concept study. J Periodontol. 2010;81:616–625. doi: 10.1902/jop.2009.090530. [DOI] [PubMed] [Google Scholar]
- 75.Pelegrini CB, Maia LP, de Souza SL, Taba M, Jr, Palioto DB. Morphological, functional and biochemical characterization of canine gingival fibroblasts. Braz Dent J. 2013;24:128–135. doi: 10.1590/0103-6440201302144. [DOI] [PubMed] [Google Scholar]
- 76.Mariotti A, Cochran DL. Characterization of fibroblasts derived from human periodontal ligament and gingiva. J Periodontol. 1990;61:103–111. doi: 10.1902/jop.1990.61.2.103. [DOI] [PubMed] [Google Scholar]
- 77.Yoshino H, Morita I, Murota SI, Ishikawa I. Mechanical stress induces production of angiogenic regulators in cultured human gingival and periodontal ligament fibroblasts. J Periodontal Res. 2003;38:405–410. doi: 10.1034/j.1600-0765.2003.00660.x. [DOI] [PubMed] [Google Scholar]
- 78.Wennström JL, Lindhe J. Some effects of enamel matrix proteins on wound healing in the dento-gingival region. J Clin Periodontol. 2002;29:9–14. doi: 10.1034/j.1600-051x.2002.290102.x. [DOI] [PubMed] [Google Scholar]
- 79.Thoma DS, Villar CC, Carnes DL, Dard M, Chun YH, Cochran DL. Angiogenic activity of an enamel matrix derivative (EMD) and EMD-derived proteins: an experimental study in mice. J Clin Periodontol. 2011;38:253–260. doi: 10.1111/j.1600-051X.2010.01656.x. [DOI] [PubMed] [Google Scholar]
- 80.Sakoda K, Nakajima Y, Noguchi K. Enamel matrix derivative induces production of vascular endothelial cell growth factor in human gingival fibroblasts. Eur J Oral Sci. 2012;120:513–519. doi: 10.1111/j.1600-0722.2012.00999.x. [DOI] [PubMed] [Google Scholar]
- 81.Lawrence TS, Beers WH, Gilula NB. Transmission of hormonal stimulation by cell-to-cell communication. Nature. 1978;272:501–506. doi: 10.1038/272501a0. [DOI] [PubMed] [Google Scholar]
- 82.Paschos NK, Brown WE, Eswaramoorthy R, Hu JC, Athanasiou KA. Advances in tissue engineering through stem cell-based co-culture. J Tissue Eng Regen Med. 2015;9:488–503. doi: 10.1002/term.1870. [DOI] [PubMed] [Google Scholar]
- 83.Hendriks J, Riesle J, van Blitterswijk CA. Co-culture in cartilage tissue engineering. J Tissue Eng Regen Med. 2007;1:170–178. doi: 10.1002/term.19. [DOI] [PubMed] [Google Scholar]
- 84.Kunz-Schughart LA, Schroeder JA, Wondrak M, van Rey F, Lehle K, Hofstaedter F, et al. Potential of fibroblasts to regulate the formation of three-dimensional vessel-like structures from endothelial cells in vitro. Am J Physiol Cell Physiol. 2006;290:C1385–C1398. doi: 10.1152/ajpcell.00248.2005. [DOI] [PubMed] [Google Scholar]
- 85.Passaniti A, Taylor RM, Pili R, Guo Y, Long PV, Haney JA, et al. A simple, quantitative method for assessing angiogenesis and antiangiogenic agents using reconstituted basement membrane, heparin, and fibroblast growth factor. Lab Investig. 1992;67:519–528. [PubMed] [Google Scholar]
- 86.Bayless KJ, Davis GE. Sphingosine-1-phosphate markedly induces matrix metalloproteinase and integrin-dependent human endothelial cell invasion and lumen formation in three-dimensional collagen and fibrin matrices. Biochem Biophys Res Commun. 2003;312:903–913. doi: 10.1016/j.bbrc.2003.11.017. [DOI] [PubMed] [Google Scholar]
- 87.Nakatsu MN, Hughes CC. An optimized three-dimensional in vitro model for the analysis of angiogenesis. Methods Enzymol. 2008;443:65–82. doi: 10.1016/S0076-6879(08)02004-1. [DOI] [PubMed] [Google Scholar]
- 88.Nehls V, Drenckhahn D. A novel, microcarrier-based in vitro assay for rapid and reliable quantification of three-dimensional cell migration and angiogenesis. Microvasc Res. 1995;50:311–322. doi: 10.1006/mvre.1995.1061. [DOI] [PubMed] [Google Scholar]
- 89.Nicosia RF, Ottinetti A. Growth of microvessels in serum-free matrix culture of rat aorta. A quantitative assay of angiogenesis in vitro. Lab Investig. 1990;63:115–122. [PubMed] [Google Scholar]
- 90.Blacher S, Devy L, Burbridge MF, Roland G, Tucker G, Noël A, et al. Improved quantification of angiogenesis in the rat aortic ring assay. Angiogenesis. 2001;4:133–142. doi: 10.1023/A:1012251229631. [DOI] [PubMed] [Google Scholar]
- 91.Zhu WH, Iurlaro M, MacIntyre A, Fogel E, Nicosia RF. The mouse aorta model: influence of genetic background and aging on bfgf- and vegf-induced angiogenic sprouting. Angiogenesis. 2003;6:193–199. doi: 10.1023/B:AGEN.0000021397.18713.9c. [DOI] [PubMed] [Google Scholar]
- 92.Akhtar N, Dickerson EB, Auerbach R. The sponge/matrigel angiogenesis assay. Angiogenesis. 2002;5:75–80. doi: 10.1023/A:1021507031486. [DOI] [PubMed] [Google Scholar]
- 93.Forough R, Wang X, Martinez-Lemus LA, Thomas D, Sun Z, Motamed K, et al. Cell-based and direct gene transfer-induced angiogenesis via a secreted chimeric fibroblast growth factor-1 (sp-FGF-1) in the chick chorioallantoic membrane (CAM) Angiogenesis. 2003;6:47–54. doi: 10.1023/A:1025857229064. [DOI] [PubMed] [Google Scholar]
- 94.Uemura A, Kusuhara S, Katsuta H, Nishikawa S. Angiogenesis in the mouse retina: a model system for experimental manipulation. Exp Cell Res. 2006;312:676–683. doi: 10.1016/j.yexcr.2005.10.030. [DOI] [PubMed] [Google Scholar]
- 95.Nicosia RF, Villaschi S. Autoregulation of angiogenesis by cells of the vessel wall. Int Rev Cytol. 1999;185:1–43. doi: 10.1016/s0074-7696(08)60148-5. [DOI] [PubMed] [Google Scholar]
- 96.Janvier R, Sourla A, Koutsilieris M, Doillon CJ. Stromal fibroblasts are required for pc-3 human prostate cancer cells to produce capillary-like formation of endothelial cells in a three-dimensional co-culture system. Anticancer Res. 1997;17:1551–1557. [PubMed] [Google Scholar]
- 97.Walter-Yohrling J, Pratt BM, Ledbetter S, Teicher BA. Myofibroblasts enable invasion of endothelial cells into three-dimensional tumor cell clusters: a novel in vitro tumor model. Cancer Chemother Pharmacol. 2003;52:263–269. doi: 10.1007/s00280-003-0664-2. [DOI] [PubMed] [Google Scholar]
- 98.Senger DR, Davis GE. Angiogenesis. Cold Spring Harb Perspect Biol. 2011;3:a005090. doi: 10.1101/cshperspect.a005090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Edgar LT, Hoying JB, Utzinger U, Underwood CJ, Krishnan L, Baggett BK, et al. Mechanical interaction of angiogenic microvessels with the extracellular matrix. J Biomech Eng. 2014;136:021001. doi: 10.1115/1.4026471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Ucuzian AA, Bufalino DV, Pang Y, Greisler HP. Angiogenic endothelial cell invasion into fibrin is stimulated by proliferating smooth muscle cells. Microvasc Res. 2013;90:40–47. doi: 10.1016/j.mvr.2013.06.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Kaully T, Kaufman-Francis K, Lesman A, Levenberg S. Vascularization—the conduit to viable engineered tissues. Tissue Eng Part B Rev. 2009;15:159–169. doi: 10.1089/ten.teb.2008.0193. [DOI] [PubMed] [Google Scholar]
- 102.Li H, Chang J. Bioactive silicate materials stimulate angiogenesis in fibroblast and endothelial cell co-culture system through paracrine effect. Acta Biomater. 2013;9:6981–6991. doi: 10.1016/j.actbio.2013.02.014. [DOI] [PubMed] [Google Scholar]
- 103.Duttenhoefer F, Lara de Freitas R, Meury T, Loibl M, Benneker LM, Richards RG, et al. 3D scaffolds co-seeded with human endothelial progenitor and mesenchymal stem cells: evidence of prevascularisation within 7 days. Eur Cell Mater. 2013;26:49–65. doi: 10.22203/ecm.v026a04. [DOI] [PubMed] [Google Scholar]
- 104.Battiston KG, Cheung JW, Jain D, Santerre JP. Biomaterials in co-culture systems: towards optimizing tissue integration and cell signaling within scaffolds. Biomaterials. 2014;35:4465–4476. doi: 10.1016/j.biomaterials.2014.02.023. [DOI] [PubMed] [Google Scholar]
- 105.Zhao D, Xue C, Lin S, Shi S, Li Q, Liu M, et al. Notch signaling pathway regulates angiogenesis via endothelial cell in 3d co-culture model. J Cell Physiol. 2017;232:1548–1558. doi: 10.1002/jcp.25681. [DOI] [PubMed] [Google Scholar]
- 106.Guerreiro SG, Brochhausen C, Negrão R, Barbosa MA, Unger RE, Kirkpatrick CJ, et al. Implanted neonatal human dermal fibroblasts influence the recruitment of endothelial cells in mice. Biomatter. 2012;2:43–52. doi: 10.4161/biom.20063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Freiman A, Shandalov Y, Rozenfeld D, Shor E, Segal S, Ben-David D, et al. Adipose-derived endothelial and mesenchymal stem cells enhance vascular network formation on three-dimensional constructs in vitro. Stem Cell Res Ther. 2016;7:5. doi: 10.1186/s13287-015-0251-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Ribatti D, Crivellato E. Mast cells, angiogenesis, and tumour growth. Biochim Biophys Acta. 2012;1822:2–8. doi: 10.1016/j.bbadis.2010.11.010. [DOI] [PubMed] [Google Scholar]
- 109.Kang T, Jones TM, Naddell C, Bacanamwo M, Calvert JW, Thompson WE, et al. Adipose-derived stem cells induce angiogenesis via microvesicle transport of mirna-31. Stem Cells Transl Med. 2016;5:440–450. doi: 10.5966/sctm.2015-0177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Kumar R, Harris-Hooker S, Kumar R, Sanford G. Co-culture of retinal and endothelial cells results in the modulation of genes critical to retinal neovascularization. Vasc Cell. 2011;3:27. doi: 10.1186/2045-824X-3-27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.McDonald SM, Matheson LA, McBane JE, Kuraitis D, Suuronen E, Santerre JP, et al. Use of monocyte/endothelial cell co-cultures (in vitro) and a subcutaneous implant mouse model (in vivo) to evaluate a degradable polar hydrophobic ionic polyurethane. J Cell Biochem. 2011;112:3762–3772. doi: 10.1002/jcb.23307. [DOI] [PubMed] [Google Scholar]
- 112.Li Q, Wang Z. Influence of mesenchymal stem cells with endothelial progenitor cells in co-culture on osteogenesis and angiogenesis: an in vitro study. Arch Med Res. 2013;44:504–513. doi: 10.1016/j.arcmed.2013.09.009. [DOI] [PubMed] [Google Scholar]
- 113.Shah AR, Shah SR, Oh S, Ong JL, Wenke JC, Agrawal CM. Migration of co-cultured endothelial cells and osteoblasts in composite hydroxyapatite/polylactic acid scaffolds. Ann Biomed Eng. 2011;39:2501–2509. doi: 10.1007/s10439-011-0344-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Baiguera S, Ribatti D. Endothelialization approaches for viable engineered tissues. Angiogenesis. 2013;16:1–14. doi: 10.1007/s10456-012-9307-8. [DOI] [PubMed] [Google Scholar]
- 115.Tarallo S, Beltramo E, Berrone E, Porta M. Human pericyte–endothelial cell interactions in co-culture models mimicking the diabetic retinal microvascular environment. Acta Diabetol. 2012;49:S141–S151. doi: 10.1007/s00592-012-0390-5. [DOI] [PubMed] [Google Scholar]
- 116.Hashimoto A, Kuroyanagi Y. Standardization for mass production of allogeneic cultured dermal substitute by measuring the amount of VEGF, bFGF, HGF, TGF-beta, and il-8. J Artif Organs. 2008;11:225–231. doi: 10.1007/s10047-008-0436-1. [DOI] [PubMed] [Google Scholar]
- 117.Hofmann A, Ritz U, Verrier S, Eglin D, Alini M, Fuchs S, et al. The effect of human osteoblasts on proliferation and neo-vessel formation of human umbilical vein endothelial cells in a long-term 3D co-culture on polyurethane scaffolds. Biomaterials. 2008;29:4217–4226. doi: 10.1016/j.biomaterials.2008.07.024. [DOI] [PubMed] [Google Scholar]
- 118.Montesano R, Pepper MS, Orci L. Paracrine induction of angiogenesis in vitro by swiss 3t3 fibroblasts. J Cell Sci. 1993;105:1013–1024. doi: 10.1242/jcs.105.4.1013. [DOI] [PubMed] [Google Scholar]
- 119.Martin TA, Harding KG, Jiang WG. Regulation of angiogenesis and endothelial cell motility by matrix-bound fibroblasts. Angiogenesis. 1999;3:69–76. doi: 10.1023/A:1009004212357. [DOI] [PubMed] [Google Scholar]
- 120.Martin TA, Harding K, Jiang WG. Matrix-bound fibroblasts regulate angiogenesis by modulation of ve-cadherin. Eur J Clin Investig. 2001;31:931–938. doi: 10.1046/j.1365-2362.2001.00914.x. [DOI] [PubMed] [Google Scholar]
- 121.Velazquez OC, Snyder R, Liu ZJ, Fairman RM, Herlyn M. Fibroblast-dependent differentiation of human microvascular endothelial cells into capillary-like, three-dimensional networks. FASEB J. 2002;16:1316–1318. doi: 10.1096/fj.01-1011fje. [DOI] [PubMed] [Google Scholar]
- 122.Bishop ET, Bell GT, Bloor S, Broom IJ, Hendry NF, Wheatley DN. An in vitro model of angiogenesis: basic features. Angiogenesis. 1999;3:335–344. doi: 10.1023/A:1026546219962. [DOI] [PubMed] [Google Scholar]
- 123.Cheung JW, Jain D, McCulloch CA, Santerre JP. Pro-angiogenic character of endothelial cells and gingival fibroblasts cocultures in perfused degradable polyurethane scaffolds. Tissue Eng Part A. 2015;21:1587–1599. doi: 10.1089/ten.tea.2014.0548. [DOI] [PubMed] [Google Scholar]
- 124.Davis GE, Bayless KJ, Mavila A. Molecular basis of endothelial cell morphogenesis in three-dimensional extracellular matrices. Anat Rec. 2002;268:252–275. doi: 10.1002/ar.10159. [DOI] [PubMed] [Google Scholar]
- 125.Blumbach K, Zweers MC, Brunner G, Peters AS, Schmitz M, Schulz JN, et al. Defective granulation tissue formation in mice with specific ablation of integrin-linked kinase in fibroblasts–role of TGFβ1 levels and RhoA activity. J Cell Sci. 2010;123:3872–3883. doi: 10.1242/jcs.063024. [DOI] [PubMed] [Google Scholar]
- 126.Sottile J. Regulation of angiogenesis by extracellular matrix. Biochim Biophys Acta. 2004;1654:13–22. doi: 10.1016/j.bbcan.2003.07.002. [DOI] [PubMed] [Google Scholar]
- 127.Din J, Yin YZ, Huang JN, Hu CJ. The effect of vascular endothelial cells on the migration of periodontal ligament cells and gingival fibroblasts. Shanghai Kou Qiang Yi Xue. 2014;23:172–178. [PubMed] [Google Scholar]