


Keywords: nerve regeneration, YL-IPA08, hippocampus, dentate gyrus, lipopolysaccharide, neuroinflammation, depression, translocator protein, neural regeneration
Abstract
Translocator protein has received attention for its involvement in the pathogenesis of depression. This study assessed the effects of the new translocator protein ligand, YL-IPA08, on alleviating inflammation-induced depression-like behavior in mice and investigated its mechanism of action. Mice were intracerebroventricularly injected with 1, 10, 100 or 1000 ng lipopolysaccharide. The tail-suspension test and the forced swimming test confirmed that 100 ng lipopolysaccharide induced depression-like behavior. A mouse model was then established by intraventricular injection of 100 ng lipopolysaccharide. On days 16–24 after model establishment, mice were intragastrically administered 3 mg/kg YL-IPA08 daily. Immunohistochemistry was used to determine BrdU and NeuN expression in the hippocampus. YL-IPA08 effectively reversed the depression-like behavior of lipopolysaccharide-treated mice, restored body mass, increased the number of BrdU-positive cells, and the number and proportion of BrdU and NeuN double-positive cells. These findings indicate that YL-IPA08 can attenuate lipopolysaccharide-induced depression-like behavior in mice by promoting the formation of hippocampal neurons.
Introduction
Depression is a chronic, severe, widespread and burdensome psychiatric illness. Unfortunately, the pathophysiological mechanism underlying depression and anxiety remains obscure. Over the past two decades, evidence has accumulated to show that inflammation can contribute to the etiology of clinical depression in co-morbid accompanying diseases (Smith, 1991; Maes, 2009; Raison and Miller, 2011; Beumer et al., 2012; Berk et al., 2013). Components that mediate inflammation in the immune system can participate or be involved in the pathology of depression (Slavich and Irwin, 2014). Inflammation can in turn induce behavioral performance changes, including low mood, anhedonia, social-behavioral withdrawal and psychomotor retardation, which are the initial symptoms of depression (Lotrich, 2015). Knock-out or down-regulation of cytokines can exert anti-depression-like effects (Lawson et al., 2013a). These results led to potential therapies for clinical depression targeting inflammation (Catena-Dell’Osso et al., 2011).
In the central nervous system, translocator protein (TSPO) is mainly located in the outer membrane of glial cell mitochondria and acts as a mediator in the translocation of cholesterol from the outer to the inner mitochondrial membrane, which is the rate-limiting step in the synthesis of neurosteriods, including allopregnanolone (Nothdurfter et al., 2012; Hatty and Banati, 2015). Moreover, TSPO participates in inflammation and regulates microglial activation (Karlstetter et al., 2014). Chronic stress and depression induce a decrease in TSPO density in the brain and peripheral organs in rodents (Milenkovic et al., 2015; Wang et al., 2015; Zhang et al., 2016). Moreover, TSPO ligand, PK 11195, demonstrated significant neuroprotective effects by inhibiting microglial activation and cytokine accumulation (Leaver et al., 2012). Allopregnanolone is one of the most important neurosteroids and alleviates neuroinflammation and contributes to neural regeneration in many rodent models (Wang et al., 2010; Nezhadi et al., 2016). Overexpression of TSPO and increased allopregnanolone levels reversed lipopolysaccharide (LPS)-induced inflammation and neural impairment (Wang et al., 2016a). Thus, TSPO is a promising target for anti-depressant and anxiolytic drugs, but its specific mechanism of action remains to be determined.
To search for new TSPO ligands for the therapeutic treatment of stress-related disorders, a series of novel compounds was designed and synthesized. Among these compounds, N-ethyl-N-(2-pyridinyl methyl)-2-(3,4-ichlorophenyl)-7-methylimidazo 1,2-a] pyridine-3-acetamide hydrochloride (YL-IPA08) was identified for its high TSPO affinity and strong anti-depressant and anxiolytic-like effects in the absence of tolerance and withdrawal liabilities. The current study first investigated whether intracerebroventricular LPS could induce depression-like behavior in mice, and then tested the hypothesis that YL-IPA08 could attenuate inflammation involved in depression-like behavior. We also examined hippocampal neurogenesis after LPS and YL-IPA08 administration.
Materials and Methods
Animals
One hundred and twenty specific-pathogen-free adult male C57BL/6 mice weighing 18–22 g and aged 6–8 weeks were obtained from the Beijing SPF Animal Technology Company, Beijing, China (Animal license No. SCXK 2016-0002). All mice were housed with controlled temperature (23 ± 1°C), humidity (45%), and lighting (12 hours per day). Food and water were available ad libitum unless specified. All procedures were conducted according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals (8th edition). The experimental procedures were approved by the Institutional Committee on Animal Care and Use of the Academy of Military Medical Sciences (approval No. IACUC.20094). All efforts were made to minimize animal suffering and reduce the number of animals used for the experiments.
Group assignment
Sixty mice were randomly assigned into five groups and administrated with 0, 1, 10, 100, or 1000 ng LPS (Sigma-Aldrich, St. Louis, MO, USA) by intracerebroventricular microinjection (n = 12 per group; Table 1). Tail suspension and forced swimming tests were performed 24 hours after LPS administration to determine the optimal dose of LPS to induce depression-like behavior.
Table 1.
Group assignment

An additional sixty mice were assigned into four groups: control, LPS (100 ng), YL-IPA08, and LPS + YL-IPA08 (n = 15 per group; Table 1). Two weeks after BrdU injection (day 1), mice were treated with LPS by intracerebroventricular injection (day 16).
Intervention
YL-IPA08 (purity ≥ 99%) (a selective TSPO ligand that has entered phase II clinical trials) was synthesized by the Department of Medicinal Chemistry in the Academy of Military Medical Sciences, Beijing, China (Figure 1). Bromodeoxyuridine (BrdU) and LPS were purchased from Sigma-Aldrich. BrdU was dissolved in 0.9% saline containing 2.5% dimethyl sulfoxide. For evaluation of neurogenesis, BrdU (100 mg/kg) was administered intraperitoneally three times with 3-hour intervals two weeks before LPS administration.
Figure 1.
Chemical structure of YL-IPA08.
For the intracerebroventricular injection of LPS, mice were anesthetized by intraperitoneal injection of Avertin (200 mg/kg; Sigma) and placed on a stereotactic apparatus (Kopf Instruments, Tujunga, CA, USA) for stereotactic injection of LPS. Injection was made through a hole drilled in the skull into the lateral ventricle using the following the coordinates of the mouse brain atlas (Franklin and Paxinos, 2008) (in mm): 0.5 posterior, +1.0 lateral and 2.0 ventral from bregma. The injection speed was set at 0.667 μL/min and the needle was left in place for 1 minute following injection.
LPS was dissolved in artificial cerebrospinal fluid and administered by intracerebroventricular injection in a volume of 2 μL. The artificial cerebrospinal fluid vehicle contained 140 mM NaCl, 3.0 mM KCl, 2.5 mM CaCl2, 1.0 mM MgCl2, and 1.2 mM Na2HPO4, adjusted to pH 7.4. YL-IPA08 (3 mg/kg) was administered by intragastric gavage at a volume of 2 mL/kg from 8:00 to 9:00 a.m., daily from the third week to the end of the experiment (days 16–24). The dose and route of administration are based on our previous study (Zhang et al., 2014b) and preliminary exploration. Behavioral tests were performed 60 minutes after YL-IPA08 administration (day 17).
Body weight of mice in all four groups was determined daily using electronic scales (Scientech, Boulder, CO, USA). One week after behavioral tests, the mice were euthanized by CO2 asphyxiation followed by transcardial perfusion with ice-cold phosphate buffered saline (PBS) (day 24). The brains were immediately removed, fixed in 4% paraformaldehyde for 48 hours, cryoprotected in 30% sucrose at 4°C for 48 hours and then processed for BrdU/NeuN histological analysis.
Behavioral experiments
Open field test
To evaluate whether the reversion of depression-like behavior by YL-IPA08 depends on affecting locomotor activity, the number of line crossings and rearings 24 hours after LPS administration was assessed as described in our previous studies (Qiu et al., 2013; Zhang et al., 2016). Mice were placed in the corner of a plastic box (36 cm × 29 cm × 23 cm; XR-XZ301, Xinruan Technology Co., Shanghai, China) in which the base was divided into equal sectors and recorded for 5 minutes. The number of crossings (all four paws placed into a new square) and rearings (both front paws raised from the floor) were counted by an observer who was blind to the group allocation.
Tail suspension test
The tail suspension test was performed as previously described (Steru et al., 1985), with minor modifications. The mice were suspended for 6 minutes from the top of the apparatus (Biowill Co., Ltd., Shanghai, China) using adhesive tape placed approximately 1 cm from the tip of the tail. The duration of immobility during the last 4 minutes of the 6 minutes was measured. The mice were judged to be immobile when they hung passively without moving. YL-IPA08 was administered 60 minutes before the test.
Forced swimming test
The forced swimming test was performed following the protocol of Porsolt et al. (1978), with minor modifications. All of the mice received a single YL-IPA08 administration. Sixty minutes after drug administration, the mice were individually placed in cylindrical containers (diameter 12 cm, height 20 cm, containing 10 cm of water maintained at 25°C; Biowill Co., Ltd.) for 6 minutes. The duration of immobility during the last 4 minutes of the 6 minutes was recorded. The mice were considered to be immobile when they floated motionless, only making movements necessary to keep their heads above the water.
Immunohistochemistry
Immunohistochemistry was performed as described in our previous report (Li et al., 2009). After behavioral testing, mice that received intraperitoneal injections of BrdU were deeply anesthetized and transcardially perfused with 0.9% NaCl. Brains were then removed and fixed for 24 hours in 4% buffered formalin before cryoprotection in 30% sucrose in PBS. For immunofluorescent staining of BrdU, free-floating brain sections were incubated in 2× standard saline citrate/50% formamide at 65°C for 2 hours, and then in 2 N HCl at 37°C for 30 minutes and 0.1 M boric acid (pH 8.5) for 10 minutes before blocking with PBS-plus for 60 minutes. Brain sections were incubated for 1 day in cold PBS-plus containing both rat anti-BrdU antibody (1:200; Abcam, Cambridge, MA, USA) and mouse anti-NeuN antibody (1:1000; Chemicon, Temecula, CA, USA). After rinsing with PBS, the sections were incubated with Red-X-conjugated goat anti-rat IgG and FITC-conjugated goat anti-mouse IgG (both at a dilution of 1:200; Jackson, MS, USA) in PBS for 2 hours before mounting with Vectashield (Vector Laboratories, Burlingame, CA, USA). The sections were analyzed using a confocal laser-scanning microscope (Zeiss LSM510, Thornwood, NY, USA). BrdU-positive cells were counted using a modified stereological protocol. In brief, every sixth section throughout the entire hippocampus was analyzed. All BrdU-labeled cells in the granular cell layer and hilus were counted through a 60× objective lens to distinguish individual cells and multiplied by six, to record as the total number of labeled cells in the dentate gyrus. At least fifty BrdU-positive cells in the DG were randomly identified from each animal to determine the percentage of newborn neurons among the BrdU-labeled cells.
Statistical analysis
Data are represented as the mean ± SEM. Data were analyzed using GraphPad Prism 6 software (Graphpad Prism Institute Inc., La Jolla, CA, USA) by an observer who was blind to the experimental protocol. The dose effect of LPS was analyzed using one-way analysis of variance with Dunnett's test for multiple comparisons. All other measures were analyzed using two-way analysis of variance with Bonferroni test for multiple comparisons. When the two-way interaction P value was < 0.05, post-hoc analysis using Fisher's protected least significant difference test was employed to test differences among means.
Results
Effect of intracerebroventricular injection of LPS on depression-like behavior in mice
To test the hypothesis that central LPS would induce depression-like behavior, tail suspension and forced swimming tests were employed following intracerebroventricular treatment of LPS. In the tail suspension test, a single intracerebroventricular injection of LPS increased immobility 24 hours after treatment (LPS main effect, F4,52 = 7.961, P < 0.001; Figure 2A). Doses of 100–1000 ng significantly increased immobility (P100 ng < 0.001, P1000 ng < 0.0001 vs. vehicle group; Dunnett's test; Figure 2A). Likewise, in the forced swimming test, a single intracerebroventricular injection of LPS increased immobility 24 hours after treatment (LPS main effect, F4,53 = 3.619, P < 0.05; Figure 2B). Doses of 10–100 ng significantly induced depression-like behavior (P10 ng < 0.001, P100 ng < 0.0001 vs. control group; Dunnett's test; Figure 2B). These results indicated that a single intracerebroventricular injection of LPS could induce depression-like behavior in C57 mice.
Figure 2.
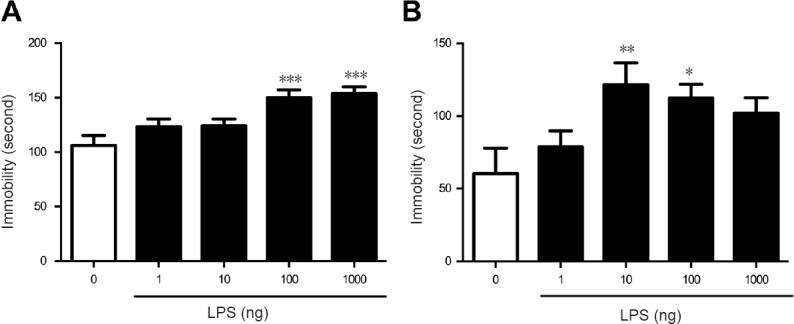
Intracerebroventricular LPS induced depression-like behavior in mice.
Twenty-four hours after a single intracerebroventricular injection of LPS, immobility was increased in the tail suspension test (A) and the forced swimming test (B). Effective doses were 100–1000 ng and 10–100 ng, respectively. Data are expressed as the mean ± SEM (n = 12; one-way analysis of variance followed by Dunnett's test). *P < 0.05, **P < 0.01, ***P < 0.001, vs. 0 ng LPS. LPS: Lipopolysaccharide.
Anti-depression-like effect of YL-IPA08 in mice presenting with LPS-induced depression-like behavior
Following the LPS dose-effect experiment, 100 ng LPS was chosen as the dose to induce depression-like behavior and the effects of LPS and YL-IPA08 on locomotor activity were tested. As shown in Figure 2A, neither LPS nor YL-IPA08 changed the number of line crossings or rearings in the open field test (FLPS = 1.248, FDFO = 0.3903, FLPS×DFO = 0.2423, for line crossings, P > 0.05; FLPS = 0.976, FDFO = 1.5102, FLPS×DFO = 1.836, for rearings, P > 0.05; Figure 3A, B), indicating that differences in behavioral performance in other tests was not because of differences in locomotor activity.
Figure 3.

YL-IPA08 attenuated 100 ng LPS-induced depression-like behavior and sickness response in mice.
LPS was dissolved in aCSF. (A, B) Line crossings (A) and rearings (B) in the open field test were measured to estimate the effect of LPS and/or YL-IPA08 (a selective translocator protein ligand) on locomotor activity. (C, D) Duration of immobility in the tail suspension test (C) and forced swimming test (D) was determined 24 hours after LPS microinjection. LPS-treated mice spent more time immobile compared to vehicle-treated mice, while the duration of immobility in LPS-exposed mice was reduced by YL-IPA08. (E) Body weight was measured 24 hours after LPS injection. Data are expressed as the mean ± SEM (n = 15; two-way analysis of variance with Bonferroni test for multiple comparisons). *P < 0.05, **P < 0.01; #P < 0.05. LPS: Lipopolysaccharide; aCSF: artificial cerebrospinal fluid.
YL-IPA08 protected mice from LPS-induced depression-like behavior in the tail suspension test and the forced swimming test. YL-IPA08 attenuated the LPS-induced increase in immobility in the tail suspension test (FLPS = 9.623, P = 0.0038; FDFO = 19.24, P = 0.0199; FLPS×DFO = 14.186, P = 0.0001; Figure 3C) and the forced swimming test (FLPS = 9.623, P = 0.0038; FDFO = 19.24, P = 0.0199; FLPS×DFO = 14.186, P = 0.0001; Figure 3D).
Changes in body weight were analyzed to assess a sickness response induced by LPS. As expected, mice lost body weight 24 hours following LPS treatment, which was reversed by YL-IPA08 (FLPS = 8.749, P = 0.036 FDFO = 7.55, P = 0.0192; FLPS×DFO = 17.55, P = 0.0002; Figure 3E).
Effects of LPS and YL-IPA08 on neurogenesis in the hippocampus
Many studies have shown that inflammation reduces neurogenesis and that this reduction might be involved with depression-like behavior. We used immunofluorescence histology to label BrdU- and NeuN-positive cells in the dentate gyrus to measure neurogenesis (Figure 4A, B). As shown in Figure 4, the number of BrdU-positive cells was reduced in the LPS group (FLPS = 14.56, P = 0.0051; Figure 4C) as was the number of cells co-labeled with BrdU and NeuN (FLPS = 18.85, P = 0.0022; Figure 4D). The percentage of BrdU- and NeuN-positive cells among total BrdU-positive cells was also reduced (FLPS = 13.78, P = 0.0051; Figure 4E). YL-IPA08 alleviated these reductions (FYL-IPA08 = 8.313, P = 0.0360, FYL-IPA08×LPS = 8.122, P = 0.037 for BrdU+ cells; FYL-IPA08 = 8.211, P = 0.005, FYL-IPA08×LPS = 6.323, P = 0.022 for BrdU+/NeuN+ co-labeled cells; FYL-IPA08 = 5.987, P = 0.0462, FYL-IPA08×LPS = 7.021, P = 0.0356, for the percentage of co-labeled BrdU- and NeuN-positive cells among total BrdU+ cells; Figure 4C-E). These data indicated that YL-IPA08 attenuated the impairment of hippocampal neurogenesis induced by LPS.
Figure 4.

YL-IPA08 (a selective translocator protein ligand) attenuates the reduction in neurogenesis caused by 100 ng LPS.
LPS was dissolved in aCSF. (A) BrdU cells are labeled red in the micrographs of the hippocampal dentate gyrus. Scale bar: 500 μm. (B) Confocal micrographs of NeuN+ (green) and BrdU+ (red) cells in the hippocampal dentate gyrus. Scale bar: 50 μm. (C–E) The number of BrdU-labeled (C), NeuN+/BrdU+ cells (D) and the percentage of BrdU+/NeuN+ cells to total BrdU+ cells (E) were reduced in the LPS group, but the reductions were reversed in the YL-IPA08 + LPS group. Data are expressed as the mean ± SEM (n = 5; two-way analysis of variance with Bonferroni test for multiple comparisons). **P < 0.01; #P < 0.05. LPS: Lipopolysaccharide; aCSF: artificial cerebrospinal fluid.
Discussion
Inflammation in the central nervous system can cause depression-like behavior in many rodent models (Kojima et al., 2014; Byrne et al., 2015; Kiecolt-Glaser et al., 2015). TSPO as a modulator that participates in the pathology of neuroinflammation also plays an important role in the development of depression, but the mechanism remains unclear. This study tested the hypothesis that TSPO ligand, YL-IPA08, could attenuate lipopolysaccharide-induced depression-like behavior in mice.
LPS acts as a bacterial Toll-like receptor 4 ligand and can activate the innate immune response. LPS administered by systemic injection, intracerebral microinjection or chronic infusion induces inflammation in rodents. LPS can result in accumulation of pro-inflammatory cytokines and neuronal cell death via apoptosis (Bossù et al., 2012; Huang et al., 2012; Zhang et al., 2015; Wang et al., 2016b; Yang et al., 2017). It is known that cytokines disrupt neuronal functions and cause many mental disorders. In the current study, intracerebroventricular injection of LPS was used to establish an inflammation-induced depression-like behavior mouse model. Compared with peripheral inflammation, central inflammation is more involved with depression and dominates the effect of peripheral inflammation, which will affect an animal's performance in behavioral tests. Furthermore, TSPO has been reported to play an important role in microglial activation and expression of pro-inflammatory cytokines (Liu et al., 2014). Studies showed that TSPO ligands such as PK11195 and XBD173 could target to TSPO to exert a broad influence on the inflammatory signaling in microglia (Nothdurfter et al., 2012a; Karlstetter et al., 2014a; Scholz et al., 2015). These knowledges support our hypothesis that the amelioration of TSPO ligand YL-IPA08 on the immune response elicited by LPS was attributed to the effect of TSPO, giving that the YL-IPA08 alone had no significant effect on depression-like behaviors or inflammatory markers compared with the control group.
The tail suspension and forced swimming tests are two widely used screening tests for the detection of anti-depressive effects in mice (Castagne et al., 2011). In this study, intracerebroventricular injection of 100 ng LPS induced depression-like behavior in both tests without changing locomotor activity. The difference in dose-dependence between the two models may result from differences in test sensitivity.
LPS-induced body weight loss indicated a sickness response, which is consistent with our previous study (Zhang et al., 2015). Body weight change is a clinical symptom in depression patients (Krimpuri et al., 2018), and body weight loss is considered part of the sickness response of many mental disorders (Lawson et al., 2013b). However, this impairment was remarkably attenuated by treatment with YL-IPA08. This effect might be related to neurosteroid biosynthesis in which TSPO participates.
Over the past twenty years, neurosteroids have been considered potential treatments for many mental disorders, including anxiety and depression, and impaired neurosteroid synthesis contributes to the pathology of these mental impairments (Pinna et al., 2006). Clinical research also discovered decreased levels of neurosteroids in plasma, serum, and cerebrospinal fluid of depression patients. Of these neurosteroids, the progesterone metabolite allopregnanolone is a potent positive modulator of the action of γ-aminobutyric acid (Nin et al., 2011; Zorumski et al., 2013; Comenencia-Ortiz et al., 2014). Importantly, neuroactive steroids elicit anxiolytic and anti-depressant properties in many different animal models (Eser et al., 2006, 2008; Brinton, 2013; Jahn, 2016). Recently, the role of TSPO has drawn increased attention in the pathophysiology of depression-related disorders (Schüle et al., 2014; Maciukiewicz et al., 2015; Setiawan et al., 2015). The TSPO ligand, Ro5-4864, reduced microglial activation and neuronal apoptosis induced by quinolinic acid (Ryu et al., 2005). R05-4684 and another ligand, PK11195, exerts an anti-inflammatory effect in many animal models of acute inflammation, including arthritis (Waterfield et al., 1999), pleural exudation (Torres et al., 2000) and inflammation in the central nervous system (Ryu et al., 2005; Veiga et al., 2005), indicating anti-inflammatory properties in addition to regulation of depression and stress. YL-IPA08, as a new TSPO ligand with high TSPO affinity, exerts potent antidepressant-like effects (Zhang et al., 2014a, b), which is consistent with the inflammation-induced depression-like model used in this study. The dose of YL-IPA08 was chosen as a therapeutic dose according to our previous study (Zhang et al., 2014a). YL-IPA08 attenuated LPS-induced depression-like behavior without changing locomotor activity, indicating that the decrease of immobility in the behavioral tests was not a false-positive phenomenon resulting from stimulation of the central nervous system, further verifying the anti-depression effect of YL-IPA08. In addition, compared with the control group, YL-IPA08 alone did not exert an anti-depression effect. Combined with previous studies detailing the anti-depressant effect of YL-IPA08 and the depression-like effect induced by LPS, TSPO may participate in inflammation-induced depression-like behaviors, and YL-IPA08 may exert an anti-depressant effect by alleviating the inflammation. However, the molecular mechanism for this needs to be determined.
One of the most significant brain regions involved in the pathology of depression is the limbic system (Anthes, 2014; Hoogenboom et al., 2014). The prefrontal cortex, the amygdala and the hippocampus are three brain regions within the limbic system, and are the most clearly altered brain regions in depression (Anthes, 2014; Hoogenboom et al., 2014). Adult hippocampal neurogenesis was demonstrated to be intimately involved with emotion, and impaired hippocampal neurogenesis contributes to the development of depression (Masi and Brovedani, 2011; Eisch and Petrik, 2012). Moreover, dysfunctional hippocampal neurogenesis could be caused by neuroinflammation (Chugh et al., 2015; Vay et al., 2016). Furthermore, anti-depressants protect neural cell regeneration in the hippocampus (Chugh et al., 2015; Vay et al., 2016). Allopregnanolone, a product downstream of TSPO, might modulate neurogenesis in rodents (Brinton and Wang, 2006; Charalampopoulos et al., 2008). Therefore, we hypothesized that TSPO ligand YL-IPA08 may be beneficial for the improvement of inflammation-induced reduction in neural regeneration by increasing subsequent synthesis of allopregnanolone and neurogenesis. To address our hypothesis, we measured the effect of YL-IPA08 on hippocampal neurogenesis. Intracerebroventricular LPS decreased hippocampal neurogenesis, while YL-IPA08 obviously ameliorated the impaired hippocampal neurogenesis, suggesting that TSPO could participate in neuroinflammation-induced depression-like behavior, which is consistent with previous studies (Brinton and Wang, 2006; Charalampopoulos et al., 2008; Wang et al., 2016a). Thus, these results lead to the idea that reversal of impaired neurogenesis plays an important role in the neuroprotection effect of YL-IPA08. Importantly, we observed depressive-like behaviors in mice exposed to LPS ten days before BrdU/NeuN analysis. The timing of depressive-like behaviors in LPS-mice seems to be coincident with the loss of mature newborn neurons. Interestingly, LPS mice showed decreases in both BrdU+ cells and BrdU+/NeuN+ cells as well as depressive-like behaviors. One explanation may be that LPS impaired neurogenesis in the hippocampal dentate gyrus. This idea is supported by the acute strong inflammation, including cytokine accumulation, which induces immune reactions against neuronal bodies and glial cells for the survival and conditioning of neurons to regenerate severed nerves (Dubový et al., 2013; Molinos et al., 2015). The damaged neural cells lead to anxiety and depressive-like behaviors (McHugh et al., 2004). These results indicate a possible cause-and-effect relationship between the LPS-impaired neurogenesis and the LPS-induced depression-like behavior. Anti-depressants may exert their behavioral effects by increasing neurogenesis in the hippocampus (Banasr et al., 2006). In particular, YL-IPA08 in LPS mice not only reduced the loss of BrdU+ cells, but also relieved the depressive-like behaviors. This neural protective effect of YL-IPA08 is consistent with our hypothesis.
In summary, the TSPO ligand, YL-IPA08, attenuated LPS-induced depression-like behavior In C57 mice and YL-IPA08 neuroprotection may involve hippocampal neurogenesis. This study might provide a new strategy for depression treatment. A limitation of this study is a lack of measurement of neurosteroids inside and outside of mitochondria.
Additional file: Open peer review reports 1 (46.1KB, pdf) , 2 (7.8KB, pdf) .
Footnotes
Conflicts of interest: The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Financial support: This study was supported by the National Natural Science Foundation of China, No. 8167050047. The funder had no role in the study design, data collection or analysis; the decision to publish; or the preparation of the manuscript.
Institutional review board statement: This study was approved by the Institutional Committee on Animal Care and Use of Academy of Military Medical Sciences (approval No. IACUC.20094).
Copyright license agreement: The Copyright License Agreement has been signed by all authors before publication.
Data sharing statement: Datasets analyzed during the current study are available from the corresponding author on reasonable request.
Plagiarism check: Checked twice by iThenticate.
Peer review: Externally peer reviewed.
Open peer reviewers: Jigar Pravinchandra Modi, Florida Atlantic University, USA; Simone Molz, Contestado University, Brazil.
Funding: This study was supported by the National Natural Science Foundation of China, No. 8167050047.
(Copyedited by Yu J, Li CH, Qiu Y, Song LP, Zhao M)
References
- 1.Anthes E. Depression: a change of mind. Nature. 2014;515:185–187. doi: 10.1038/515185a. [DOI] [PubMed] [Google Scholar]
- 2.Banasr M, Soumier A, Hery M, Mocaer E, Daszuta A. Agomelatine, a new antidepressant, induces regional changes in hippocampal neurogenesis. Biol Psychiatry. 2006;59:1087–1096. doi: 10.1016/j.biopsych.2005.11.025. [DOI] [PubMed] [Google Scholar]
- 3.Belarbi K, Jopson T, Tweedie D, Arellano C, Luo W, Greig NH, Rosi S. TNF-alpha protein synthesis inhibitor restores neuronal function and reverses cognitive deficits induced by chronic neuroinflammation. J Neuroinflammation. 2012;9:23. doi: 10.1186/1742-2094-9-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Berk M, Williams LJ, Jacka FN, O’Neil A, Pasco JA, Moylan S, Allen NB, Stuart AL, Hayley AC, Byrne ML, Maes M. So depression is an inflammatory disease, but where does the inflammation come from? BMC Med. 2013;11:200. doi: 10.1186/1741-7015-11-200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Beumer W, Gibney SM, Drexhage RC, Pont-Lezica L, Doorduin J, Klein HC, Steiner J, Connor TJ, Harkin A, Versnel MA, Drexhage HA. The immune theory of psychiatric diseases: a key role for activated microglia and circulating monocytes. J Leukoc Biol. 2012;92:959–975. doi: 10.1189/jlb.0212100. [DOI] [PubMed] [Google Scholar]
- 6.Bossù P, Cutuli D, Palladino I, Caporali P, Angelucci F, Laricchiuta D, Gelfo F, De Bartolo P, Caltagirone C, Petrosini L. A single intraperitoneal injection of endotoxin in rats induces long-lasting modifications in behavior and brain protein levels of TNF-alpha and IL-18. J Neuroinflammation. 2012;9:101. doi: 10.1186/1742-2094-9-101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Brinton RD. Neurosteroids as regenerative agents in the brain: therapeutic implications. Nat Rev Endocrinol. 2013;9:241–250. doi: 10.1038/nrendo.2013.31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Brinton RD, Wang JM. Preclinical analyses of the therapeutic potential of allopregnanolone to promote neurogenesis in vitro and in vivo in transgenic mouse model of Alzheimer's disease. Curr Alzheimer Res. 2006;3:11–17. doi: 10.2174/156720506775697160. [DOI] [PubMed] [Google Scholar]
- 9.Byrne ML, O’Brien-Simpson NM, Mitchell SA, Allen NB. Adolescent-onset depression: are obesity and inflammation developmental mechanisms or outcomes? Child Psychiatry Hum Dev. 2015;46:839–850. doi: 10.1007/s10578-014-0524-9. [DOI] [PubMed] [Google Scholar]
- 10.Castagne V, Moser P, Roux S, Porsolt RD. Rodent models of depression: forced swim and tail suspension behavioral despair tests in rats and mice. Curr Protoc Neurosci Chapter 8:Unit 8.10A. 2011 doi: 10.1002/0471142301.ns0810as55. [DOI] [PubMed] [Google Scholar]
- 11.Catena-Dell’Osso M, Bellantuono C, Consoli G, Baroni S, Rotella F, Marazziti D. Inflammatory and neurodegenerative pathways in depression: a new avenue for antidepressant development? Curr Med Chem. 2011;18:245–255. doi: 10.2174/092986711794088353. [DOI] [PubMed] [Google Scholar]
- 12.Charalampopoulos I, Remboutsika E, Margioris AN, Gravanis A. Neurosteroids as modulators of neurogenesis and neuronal survival. Trends Endocrinol Metab. 2008;19:300–307. doi: 10.1016/j.tem.2008.07.004. [DOI] [PubMed] [Google Scholar]
- 13.Chugh D, Ali I, Bakochi A, Bahonjic E, Etholm L, Ekdahl CT. Alterations in brain inflammation, synaptic proteins, and adult hippocampal neurogenesis during epileptogenesis in mice lacking synapsin2. PLoS One. 2015;10:e0132366. doi: 10.1371/journal.pone.0132366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Comenencia-Ortiz E, Moss SJ, Davies PA. Phosphorylation of GABAA receptors influences receptor trafficking and neurosteroid actions. Psychopharmacology (Berl) 2014;231:3453–3465. doi: 10.1007/s00213-014-3617-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dubový P, Jancalek R, Kubek T. Role of inflammation and cytokines in peripheral nerve regeneration. Int Rev Neurobiol. 2013;108:173–206. doi: 10.1016/B978-0-12-410499-0.00007-1. [DOI] [PubMed] [Google Scholar]
- 16.Eisch AJ, Petrik D. Depression and hippocampal neurogenesis: a road to remission? Science. 2012;338:72–75. doi: 10.1126/science.1222941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Eser D, Baghai TC, Schule C, Nothdurfter C, Rupprecht R. Neuroactive steroids as endogenous modulators of anxiety. Curr Pharm Des. 2008;14:3525–3533. doi: 10.2174/138161208786848838. [DOI] [PubMed] [Google Scholar]
- 18.Eser D, Schule C, Romeo E, Baghai TC, di Michele F, Pasini A, Zwanzger P, Padberg F, Rupprecht R. Neuropsychopharmacological properties of neuroactive steroids in depression and anxiety disorders. Psychopharmacology (Berl) 2006;186:373–387. doi: 10.1007/s00213-005-0188-z. [DOI] [PubMed] [Google Scholar]
- 19.Franklin KBJ, Paxinos G. San Diego: Academic Press; 2008. The Mouse Brain in Stereotaxic Coordinates. [Google Scholar]
- 20.Hatty CR, Banati RB. Protein-ligand and membrane-ligand interactions in pharmacology: the case of the translocator protein (TSPO) Pharmacol Res. 2015;100:58–63. doi: 10.1016/j.phrs.2015.07.029. [DOI] [PubMed] [Google Scholar]
- 21.Hoogenboom WS, Perlis RH, Smoller JW, Zeng-Treitler Q, Gainer VS, Murphy SN, Churchill SE, Kohane IS, Shenton ME, Iosifescu DV. Limbic system white matter microstructure and long-term treatment outcome in major depressive disorder: a diffusion tensor imaging study using legacy data. World J Biol Psychiatry. 2014;15:122–134. doi: 10.3109/15622975.2012.669499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Huang HJ, Chen YH, Liang KC, Jheng YS, Jhao JJ, Su MT, Lee-Chen GJ, Hsieh-Li HM. Exendin-4 protected against cognitive dysfunction in hyperglycemic mice receiving an intrahippocampal lipopolysaccharide injection. PLoS One. 2012;7:e39656. doi: 10.1371/journal.pone.0039656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Jahn H. Steroid-synthesis inhibition in depression: a good idea? Lancet Psychiatry. 2016;3:92–93. doi: 10.1016/S2215-0366(15)00494-0. [DOI] [PubMed] [Google Scholar]
- 24.Karlstetter M, Nothdurfter C, Aslanidis A, Moeller K, Horn F, Scholz R, Neumann H, Weber BHF, Rupprecht R, Langmann T. Translocator protein (18 kDa) (TSPO) is expressed in reactive retinal microglia and modulates microglial inflammation and phagocytosis. J Neuroinflammation. 2014a;11:3. doi: 10.1186/1742-2094-11-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Karlstetter M, Nothdurfter C, Aslanidis A, Moeller K, Horn F, Scholz R, Neumann H, Weber BH, Rupprecht R, Langmann T. Translocator protein (18 kDa) (TSPO) is expressed in reactive retinal microglia and modulates microglial inflammation and phagocytosis. J Neuroinflammation. 2014b;11:3. doi: 10.1186/1742-2094-11-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kiecolt-Glaser JK, Derry HM, Fagundes CP. Inflammation: depression fans the flames and feasts on the heat. Am J Psychiatry. 2015;172:1075–1091. doi: 10.1176/appi.ajp.2015.15020152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kojima M, Kojima T, Suzuki S, Takahashi N, Funahashi K, Kato D, Hanabayashi M, Hirabara S, Asai S, Ishiguro N. Alexithymia, depression, inflammation, and pain in patients with rheumatoid arthritis. Arthritis Care Res (Hoboken) 2014;66:679–686. doi: 10.1002/acr.22203. [DOI] [PubMed] [Google Scholar]
- 28.Krimpuri RD, Yokley JM, Seeholzer EL, Horwath EL, Thomas CL, Bardaro SJ. Qualifying for bariatric surgery: is preoperative weight loss a reliable predictor of postoperative weight loss? Surg Obes Relat Dis. 2018;14:60–64. doi: 10.1016/j.soard.2017.07.012. [DOI] [PubMed] [Google Scholar]
- 29.Lawson MA, McCusker RH, Kelley KW. Interleukin-1 beta converting enzyme is necessary for development of depression-like behavior following intracerebroventricular administration of lipopolysaccharide to mice. J Neuroinflammation. 2013a;10:54. doi: 10.1186/1742-2094-10-54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lawson MA, Parrott JM, McCusker RH, Dantzer R, Kelley KW, O’Connor JC. Intracerebroventricular administration of lipopolysaccharide induces indoleamine-2, 3-dioxygenase-dependent depression-like behaviors. J Neuroinflammation. 2013b;10:87. doi: 10.1186/1742-2094-10-87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Leaver KR, Reynolds A, Bodard S, Guilloteau D, Chalon S, Kassiou M. Effects of translocator protein (18 kDa) ligands on microglial activation and neuronal death in the quinolinic-acid-injected rat striatum. ACS Chem Neurosci. 2012;3:114–119. doi: 10.1021/cn200099e. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Li YF, Huang Y, Amsdell SL, Xiao L, O’Donnell JM, Zhang HT. Antidepressant- and anxiolytic-like effects of the phosphodiesterase-4 inhibitor rolipram on behavior depend on cyclic AMP response element binding protein-mediated neurogenesis in the hippocampus. Neuropsychopharmacology. 2009;34:2404–2419. doi: 10.1038/npp.2009.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu GJ, Middleton RJ, Hatty CR, Kam WW, Chan R, Pham T, Harrison-Brown M, Dodson E, Veale K, Banati RB. The 18 kDa translocator protein, microglia and neuroinflammation. Brain Pathol. 2014;24:631–653. doi: 10.1111/bpa.12196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lotrich FE. Inflammatory cytokine-associated depression. Brain Res. 2015;1617:113–125. doi: 10.1016/j.brainres.2014.06.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Maciukiewicz M, Marshe VS, Tiwari AK, Fonseka TM, Freeman N, Rotzinger S, Foster JA, Kennedy JL, Kennedy SH, Muller DJ. Genetic variation in IL-1beta, IL-2, IL-6, TSPO and BDNF and response to duloxetine or placebo treatment in major depressive disorder. Pharmacogenomics. 2015;16:1919–1929. doi: 10.2217/pgs.15.136. [DOI] [PubMed] [Google Scholar]
- 36.Maes M. “Functional” or “psychosomatic” symptoms, e.g. a flu-like malaise, aches and pain and fatigue, are major features of major and in particular of melancholic depression. Neuro Endocrinol Lett. 2009;30:564–573. [PubMed] [Google Scholar]
- 37.Masi G, Brovedani P. The hippocampus, neurotrophic factors and depression: possible implications for the pharmacotherapy of depression. CNS Drugs. 2011;25:913–931. doi: 10.2165/11595900-000000000-00000. [DOI] [PubMed] [Google Scholar]
- 38.McHugh SB, Deacon RM, Rawlins JN, Bannerman DM. Amygdala and ventral hippocampus contribute differentially to mechanisms of fear and anxiety. Behav Neurosci. 2004;118:63–78. doi: 10.1037/0735-7044.118.1.63. [DOI] [PubMed] [Google Scholar]
- 39.Milenkovic VM, Rupprecht R, Wetzel CH. The translocator protein 18 kDa (TSPO) and its role in mitochondrial biology and psychiatric disorders. Mini Rev Med Chem. 2015;15:366–372. doi: 10.2174/1389557515666150324122642. [DOI] [PubMed] [Google Scholar]
- 40.Molinos M, Almeida CR, Caldeira J, Cunha C, Goncalves RM, Barbosa MA. Inflammation in intervertebral disc degeneration and regeneration. J R Soc Interface. 2015;12:20141191. doi: 10.1098/rsif.2015.0429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nezhadi A, Sheibani V, Esmaeilpour K, Shabani M, Esmaeili-Mahani S. Neurosteroid allopregnanolone attenuates cognitive dysfunctions in 6-OHDA-induced rat model of Parkinson's disease. Behav Brain Res. 2016;305:258–264. doi: 10.1016/j.bbr.2016.03.019. [DOI] [PubMed] [Google Scholar]
- 42.Nin MS, Martinez LA, Pibiri F, Nelson M, Pinna G. Neurosteroids reduce social isolation-induced behavioral deficits: a proposed link with neurosteroid-mediated upregulation of BDNF expression. Front Endocrinol (Lausanne) 2011;2:73. doi: 10.3389/fendo.2011.00073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nothdurfter C, Baghai TC, Schule C, Rupprecht R. Translocator protein (18 kDa) (TSPO) as a therapeutic target for anxiety and neurologic disorders. Eur Arch Psychiatry Clin Neurosci. 2012a;262(Suppl 2):S107–112. doi: 10.1007/s00406-012-0352-5. [DOI] [PubMed] [Google Scholar]
- 44.Nothdurfter C, Baghai TC, Schüle C, Rupprecht R. Translocator protein (18 kDa) (TSPO) as a therapeutic target for anxiety and neurologic disorders. Eur Arch Psychiatry Clin Neurosci. 2012b;262:107–112. doi: 10.1007/s00406-012-0352-5. [DOI] [PubMed] [Google Scholar]
- 45.Pinna G, Costa E, Guidotti A. Fluoxetine and norfluoxetine stereospecifically and selectively increase brain neurosteroid content at doses that are inactive on 5-HT reuptake. Psychopharmacology. 2006;186:362–372. doi: 10.1007/s00213-005-0213-2. [DOI] [PubMed] [Google Scholar]
- 46.Porsolt RD, Anton G, Blavet N, Jalfre M. Behavioural despair in rats: a new model sensitive to antidepressant treatments. Eur J Pharmacol. 1978;47:379–391. doi: 10.1016/0014-2999(78)90118-8. [DOI] [PubMed] [Google Scholar]
- 47.Qiu ZK, Zhang LM, Zhao N, Chen HX, Zhang YZ, Liu YQ, Mi TY, Zhou WW, Li Y, Yang RF, Xu JP, Li YF. Repeated administration of AC-5216, a ligand for the 18 kDa translocator protein, improves behavioral deficits in a mouse model of post-traumatic stress disorder. Prog Neuropsychopharmacol Biol Psychiatry. 2013;45:40–46. doi: 10.1016/j.pnpbp.2013.04.010. [DOI] [PubMed] [Google Scholar]
- 48.Raison CL, Miller AH. Is depression an inflammatory disorder? Curr Psychiatry Rep. 2011;13:467–475. doi: 10.1007/s11920-011-0232-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ryu JK, Choi HB, McLarnon JG. Peripheral benzodiazepine receptor ligand PK11195 reduces microglial activation and neuronal death in quinolinic acid-injected rat striatum. Neurobiol Dis. 2005;20:550–561. doi: 10.1016/j.nbd.2005.04.010. [DOI] [PubMed] [Google Scholar]
- 50.Schüle C, Nothdurfter C, Rupprecht R. The role of allopregnanolone in depression and anxiety. Prog Neurobiol. 2014;113:79–87. doi: 10.1016/j.pneurobio.2013.09.003. [DOI] [PubMed] [Google Scholar]
- 51.Schnydrig S, Korner L, Landweer S, Ernst B, Walker G, Otten U, Kunz D. Peripheral lipopolysaccharide administration transiently affects expression of brain-derived neurotrophic factor, corticotropin and proopiomelanocortin in mouse brain. Neurosci Lett. 2007;429:69–73. doi: 10.1016/j.neulet.2007.09.067. [DOI] [PubMed] [Google Scholar]
- 52.Scholz R, Caramoy A, Bhuckory MB, Rashid K, Chen M, Xu H, Grimm C, Langmann T. Targeting translocator protein (18 kDa) (TSPO) dampens pro-inflammatory microglia reactivity in the retina and protects from degeneration. J Neuroinflammation. 2015;12:201. doi: 10.1186/s12974-015-0422-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Setiawan E, Wilson AA, Mizrahi R, Rusjan PM, Miler L, Rajkowska G, Suridjan I, Kennedy JL, Rekkas PV, Houle S, Meyer JH. Role of translocator protein density, a marker of neuroinflammation, in the brain during major depressive episodes. JAMA Psychiatry. 2015;72:268–275. doi: 10.1001/jamapsychiatry.2014.2427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Slavich GM, Irwin MR. From stress to inflammation and major depressive disorder: a social signal transduction theory of depression. Psychol Bull. 2014;140:774–815. doi: 10.1037/a0035302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Smith RS. The macrophage theory of depression. Med Hypotheses. 1991;35:298–306. doi: 10.1016/0306-9877(91)90272-z. [DOI] [PubMed] [Google Scholar]
- 56.Steru L, Chermat R, Thierry B, Simon P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology. 1985;85:367–370. doi: 10.1007/BF00428203. [DOI] [PubMed] [Google Scholar]
- 57.Torres SR, Frode TS, Nardi GM, Vita N, Reeb R, Ferrara P, Ribeiro-do-Valle RM, Farges RC. Anti-inflammatory effects of peripheral benzodiazepine receptor ligands in two mouse models of inflammation. Eur J Pharmacol. 2000;408:199–211. doi: 10.1016/s0014-2999(00)00760-3. [DOI] [PubMed] [Google Scholar]
- 58.Vay SU, Blaschke S, Klein R, Fink GR, Schroeter M, Rueger MA. Minocycline mitigates the gliogenic effects of proinflammatory cytokines on neural stem cells. J Neurosci Res. 2016;94:149–160. doi: 10.1002/jnr.23686. [DOI] [PubMed] [Google Scholar]
- 59.Veiga S, Azcoitia I, Garcia-Segura LM. Ro5-4864, a peripheral benzodiazepine receptor ligand, reduces reactive gliosis and protects hippocampal hilar neurons from kainic acid excitotoxicity. J Neurosci Res. 2005;80:129–137. doi: 10.1002/jnr.20430. [DOI] [PubMed] [Google Scholar]
- 60.Wang DS, Tian Z, Guo YY, Guo HL, Kang WB, Li S, Den YT, Li XB, Feng B, Feng D, Zhao JN, Liu G, Zhao MG. Anxiolytic-like effects of translocator protein (TSPO) ligand ZBD-2 in an animal model of chronic pain. Mol Pain. 2015;11:16. doi: 10.1186/s12990-015-0013-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wang JM, Singh C, Liu L, Irwin RW, Chen S, Chung EJ, Thompson RF, Brinton RD. Allopregnanolone reverses neurogenic and cognitive deficits in mouse model of Alzheimer's disease. Proc Natl Acad Sci U S A. 2010;107:6498–6503. doi: 10.1073/pnas.1001422107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Wang W, Zhang L, Zhang X, Xue R, Li L, Zhao W, Fu Q, Mi W, Li Y. Lentiviral-mediated overexpression of the 18 kDa translocator protein (TSPO) in the hippocampal dentate gyrus ameliorates LPS-induced cognitive impairment in mice. Front Pharmacol. 2016a;7:384. doi: 10.3389/fphar.2016.00384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wang ZH, Wang SH, Wu Y, Li Z, Liao XL, Qin TH. Dexamethasone inhibits the expression of microRNA-155 in macrophages induced by lipopolysaccharide. Zhongguo Zuzhi Gongcheng Yanjiu. 2016b;20:1591–1596. [Google Scholar]
- 64.Waterfield JD, McGeer EG, McGeer PL. The peripheral benzodiazepine receptor ligand PK 11195 inhibits arthritis in the MRL-lpr mouse model. Rheumatology (Oxford) 1999;38:1068–1073. doi: 10.1093/rheumatology/38.11.1068. [DOI] [PubMed] [Google Scholar]
- 65.Yang H, Cheng X, Yang YL, Wang YH, Du GH. Ramulus Cinnamomi extract attenuates neuroinflammatory responses via downregulating TLR4/MyD88 signaling pathway in BV2 cells. Neural Regen Res. 2017;12:1860–1864. doi: 10.4103/1673-5374.219048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Zhang LM, Qiu ZK, Zhao N, Chen HX, Liu YQ, Xu JP, Zhang YZ, Yang RF, Li YF. Anxiolytic-like effects of YL-IPA08, a potent ligand for the translocator protein (18 kDa) in animal models of post-traumatic stress disorder. Int J Neuropsychopharmacol. 2014a;17:1659–1669. doi: 10.1017/S1461145714000479. [DOI] [PubMed] [Google Scholar]
- 67.Zhang LM, Qiu ZK, Chen XF, Zhao N, Chen HX, Xue R, Zhang YZ, Yang RF, Li YF. Involvement of allopregnanolone in the anti-PTSD-like effects of AC-5216. J Psychopharmacol. 2016;30:474–481. doi: 10.1177/0269881115625115. [DOI] [PubMed] [Google Scholar]
- 68.Zhang LM, Zhao N, Guo WZ, Jin ZL, Qiu ZK, Chen HX, Xue R, Zhang YZ, Yang RF, Li YF. Antidepressant-like and anxiolytic-like effects of YL-IPA08, a potent ligand for the translocator protein (18 kDa. Neuropharmacology. 2014b;81:116–125. doi: 10.1016/j.neuropharm.2013.09.016. [DOI] [PubMed] [Google Scholar]
- 69.Zhang XY, Cao JB, Zhang LM, Li YF, Mi WD. Deferoxamine attenuates lipopolysaccharide-induced neuroinflammation and memory impairment in mice. J Neuroinflammation. 2015;12:20. doi: 10.1186/s12974-015-0238-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Zorumski CF, Paul SM, Izumi Y, Covey DF, Mennerick S. Neurosteroids, stress and depression: Potential therapeutic opportunities. Neurosci Biobehav Rev. 2013;37:109–122. doi: 10.1016/j.neubiorev.2012.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.