

Abstract
Gynura procumbens (GP) is a medicinal herb that has long been known as anti-inflammatory and antihyperglycaemic. Recently, this herbal extract has been associated with a profertility effect, suggesting its applicability in treating both diabetes and male infertility. In this study, the effects of GP aqueous extract (GPAE) on diabetic rats were investigated through evaluating testes histology and androgen hormone levels as well as the implantation sites of female rats on copulation with the treated male rats. Three dosages of GPAE were used (150, 300, and 450 mg/kg), and there were three control groups [normal, diabetic, and metformin-treated diabetic]. Testes histology, androgen hormone levels, and number of implantation sites of the GPAE-treated groups matched those of the normal group in contrast to the diabetic and metformin-treated diabetic controls. Sperm proteomics analysis identified 666 proteins, but only 88 were consistently found in all the control and 450-mg/kg GPAE-treated groups. Four proteins, including cysteine-rich secretory protein 1, carboxylesterase 5A, zona pellucida binding protein, and phosphatidylethanolamine-binding protein 1, were significantly upregulated with GPAE treatment compared with the diabetic control, matching the protein levels of the normal group. These proteins were mainly involved in sperm maturation, sperm capacitation, and sperm-egg interaction, suggesting that GP treatment was able to restore the fertility of male diabetic rats at molecular protein level. In conclusion, GP treatment effectively treats infertility of male diabetic rats, possibly through the upregulation of proteins related to sperm maturation and sperm-egg interaction.
1. Introduction
Diabetes mellitus is one of the most prominent public health problems in modern societies and is rapidly increasing, specifically among those of reproductive age. According to Malaysia's National Health and Morbidity Survey [1], approximately 17.5% of individuals aged 18–36 years have diabetes mellitus, which is equivalent to 3.5 million individuals in Malaysia. Diabetes has negative effects on the male reproductive system, including testicular function, sperm maturation, and sexual hormone alteration [2]. Approximately 80% of males with diabetes mellitus experience frequent loss of libido as well as erectile and sexual dysfunctions [3]. Diabetes has also been shown to alter androgenic hormone levels, such as those of luteinising hormone (LH), follicle stimulating hormone (FSH), and testosterone [4]. High blood glucose induces a change in Leydig cells and pituitary-testicular axis, leading to decreased LH level and consequently testicular impairment [5]. Prolonged hyperglycaemia may also lead to the overproduction of ROS, which results in the disruption of spermatogenesis [6]. Hence, an effective treatment is urgently needed to overcome both hyperglycemia and fertility problem in diabetic patients.
A drug called metformin has been frequently used in treating hyperglycaemia; however, it has been found to be ineffective against infertility in diabetic patients [7]. Hence, studies are now shifting focus towards alternative herbal treatments to treat these problems. One alternative is the herb Gynura procumbens (GP). It is a medicinal plant of the Asteraceae family, known for its beneficial therapeutics effects, such as treating fever, rashes, hypertension, and diabetes mellitus [8]. This evergreen shrub grows extensively in Southeast Asia and is locally known in Malaysia as Sambung Nyawa. Several studies have reported that this herb possesses anti-inflammatory [9], antihypertensive [10], anti-cancer [11], and antihyperglycaemic [12, 13] activities and recently has been recognised as a profertility factor [14]. The administration of GP aqueous extract (GPAE) was reported to reduce blood glucose levels significantly and increased sperm quality [14]. Despite these findings, the effects of GPAE on diabetic male fertility health as well as sperm proteins have not been thoroughly elucidated.
The impact of herbal treatments on sperm proteins has been investigated in a few previous proteomics studies. For example, a two-dimensional gel electrophoresis (2DGE) study has reported that Centella asiatica (pegaga) extract caused modifications on sperm protein levels associated with energy generation process and sperm motility [15]. Additionally, administration of Lunasia amara Blanco (Sanrego) at 60 mg/kg also showed that the expression of sperm proteins involved in energy metabolism, sperm motility, amino acid metabolism, and cell signalling were highly increased, suggesting the herb profertility potential [16]. While most sperm protein studies have used 2DGE-based proteomics, another approach called shotgun proteomics, also can be performed whereby proteins are quantitatively compared on the basis of peptide signal intensities from mass spectrometry [17]. This approach provides higher identification of sperm proteins [18, 19] compared to 2DGE-based. However, application of shotgun proteomics in understanding the effect of herbal treatment especially GPAE on sperm proteins in diabetic patients is still limited and further experimentation is required.
In this study, the effects of three different doses of GPAE (150, 300, and 450 mg/kg per body weight) on spermatogenesis, hormonal levels, and implantation sites were investigated to further comprehend the ability of this herb to improve fertility in diabetic rats. Furthermore, shotgun proteomics were also conducted to analyse sperm protein changes due to the treatment which could highlight molecular significance related to the use of GPAE in diabetic rats.
2. Material and Methods
2.1. Preparation of GPAE
GP leaves were grown and collected from the Universiti Kebangsaan Malaysia glasshouse in Selangor, Malaysia (2.9300°N, 101.7774°E), and deposited in local herbarium with the voucher number, 40343 (KAK 01 (UKMB)). GPAE was prepared as described by Kamaruzaman and Mat Noor [20]. Briefly, 2.0 kg of GP leaves was dried in the oven at 48°C for 72 h. After drying, the leaves were ground to fine powder and mixed with water with the ratio of 1:20 before incubation in a water bath, at 60°C for 3 h. The extract was later filtered and freeze dried.
2.2. Diabetes Type I Induction and Experimental Design
This study was approved by the Animal Ethics Committee of Faculty of Medicine, Universiti Kebangsaan Malaysia (FST/2013/MAHANEM/31-JAN./492-FEB.-2013-FEB.-2015). A total of 42 proven fertile male 8-week-old Sprague-Dawley rats (Rattus norvegicus) were used in this study, and they were randomly divided into six groups. Three of the groups were considered as control groups: normal control (nontreated nondiabetic), diabetic control (nontreated diabetic), and metformin-treated diabetic control groups. Another three groups were considered as the treated diabetic groups with varying dosages of GPAE: 150, 300, and 450 mg/kg per body weight, respectively. These doses were chosen based on previous studies [13, 14] and also to determine the best dose among these three in treating both hyperglycaemia and fertility health.
Type I diabetes model was chosen for this study, using the chemotherapeutic drugs, streptozotocin (STZ). STZ is toxic to pancreatic β-cell, eventually induced inflammation in the pancreas causing diabetes condition in rats. STZ-induced hyperglycaemia was described as a good experimental model for type I diabetes [21]. All five groups except normal control were induced once, by a single intravenous injection of 50 mg/kg per body weight STZ at the root of the tail after overnight (12 hours) fasting. The STZ solution was prepared immediately prior to injection by dissolving the drug in a fresh, cold citrate buffer, pH 4.5. After 72 hours of induction, the fasting blood glucose level were measured using glucometer (Accucheck Performa). Rats possessing blood glucose levels of ≥ 13 mmol/L were considered diabetic. All rats were fed standard pellet diet and given access to water ad libitum. Subsequently, all treatments were given via oral gavage every day for 14 consecutive days. Nontreated normal and diabetic rats were maintained in similar conditions as the treated rats. Rats were sacrificed on day 15 for testes histology, androgen hormone levels, fertility study (implantation sites), and proteomics analysis.
2.3. Testes Histology
Testes were removed and fixed in a Bouin's Solution overnight. The testes were then dehydrated using alcohol and embedded in paraffin wax. Testes samples were sectioned at 5 μm in thickness and stained using Mallory staining. Testicular spermatogenesis was observed under light microscope.
2.4. Androgen Hormone Analyses (LH, FSH and Testosterone)
Blood samples were collected through cardiac puncture for the estimation of serum LH, FSH, and testosterone levels using a commercial kit as per manufacturer's instructions (Testosterone EIA, LH, and FSH kit by Cayman Chemical, Michigan, USA) [7].
2.5. Number of Implantation Sites
Each male rat from the six experimental groups was kept in a different cage together with two oestrous female rats. Healthy female rats were used in this study and made receptive for sexual activity by subcutaneous injections of 20 μg/rat of estradiol benzoate 52 h prior to mating and 1 mg/rat of progesterone 4 h earlier before mating. The males were allowed to be in the libido box for 10 min before the test for marking purposes [22]. Both males and females were allowed to mate for 7 days. Vaginal smears were performed to determine the gestation day. Pregnant female rats were later separated and kept for 16 days before being sacrificed, and the number of implantation sites in the uteri was recorded [23].
2.6. Sperm Proteomics Analysis
The sperm protein samples used in this study were from the three control groups and the best GPAE treatment dose group (450 mg/kg). Three biological replicates from each group were chosen for this study. The protein extraction was prepared as described in Yunianto et al. [15] with modifications. Sperm samples were harvested from the caudal epididymis. Sperms were allowed to “swim up” in Biggers–Whitten–Whittingham medium [24] for 30 min at 37°C in 5% CO2 incubator. These sperm samples were centrifuged and lysed with lysis buffer. Sodium dodecyl sulphate gel 12.5% was prepared for the electrophoresis process, which was done at the voltage of 75 V. Subsequently, protein digestion was carried out using in-gel digestion method as described by [25]. A total of 100 μg of protein lysate was incubated with dithiotreitol and iodoacetamide for reduction and alkylation steps, followed by overnight trypsinisation by adding 6 ng/μL trypsin (PROMEGA Gold, USA).
The peptide samples were then submitted into an LC system (Dionex 3000 Ultimate RSLCnano) coupled to an LTQ Orbitrap Fusion mass spectrometer (Thermo Fisher, Bremen, Germany). Digested samples (1 μL) were injected into a reverse phase column and eluted with a flow rate of 300 nL/min. Data were acquired in data-dependent mode. Precursors with an assigned monoisotopic m/z and charged 2–7 were further analysed. All precursors were filtered using a 20-s dynamic exclusion window with intensity threshold of 5000. The MS/MS spectra analysis was performed by using rapid scan rate and maximum injection time of 250 ms.
The LC-MS/MS data were analysed using MaxQuant software (version 1.5.3.30) as described by Iovinella et al. [26] and searched against the Rattus norvegicus protein sequences obtained from Uniprot database (proteome ID: UP000002494, accessed on February 2016). Peptide spectrum match and protein identification were filtered using a target-decoy approach with a false discover rate of 1%. The second peptide feature was enabled. The label-free quantification (LFQ) of protein was done using the MaxLFQ algorithm integrated in the MaxQuant software. Other MaxQuant settings were at default. All the information was reported in the “proteinGroups” output file, containing the full list of identified and quantified proteins.
The data were further analysed using Perseus software (version 1.5.4.1). The “proteinGroups” output file produced by MaxQuant was analysed. The hits to the reverse database, contaminants, and proteins identified with modified peptides were eliminated. Then, the LFQ intensity ratios were transformed by log2. Missing values were computed by drawing random numbers from a normal distribution to stimulate signals from low abundant proteins, using the default parameters [17]. The value of log2 (LFQ ratios) for each sample was then statistically analysed. Proteins identified were then annotated by using BLAST2GO (https://www.blast2go.com/) and WEGO analyses (https://biodb.swu.edu.cn/cgi-bin/wego/index.pl).
2.7. Statistical Analysis
Data were displayed as mean ± standard error of mean. The statistical analysis was performed in SPSS 22.0, using one-way analysis of variance (ANOVA) with a p value <0.05 considered statistically significant. For the proteomics analysis, protein LFQ intensities were compared using Perseus software (version 1.5.4.1) (one-way ANOVA, p < 0.05).
3. Results
3.1. Androgen Hormone Were Elevated after 14 Days of GPAE Treatment
Hormone levels for testosterone, LH, and FSH are shown in Table 1. On the basis of the results, all three hormones decreased in the diabetic rat group compared with the normal group. For example, the normal group displayed a higher level of testosterone (0.757 ± 0.014 ng/mL) compared with the negative control (0.151 ± 0.010 ng/mL). Similarly, both LH and FSH were lower in the diabetic rats (0.017 ± 0.002 ng/mL and 0.020 ± 0.008 ng/mL, respectively) compared with the normal group (0.042 ± 0.002 ng/mL and 0.037 ± 0.008 ng/mL, respectively). Furthermore, the use of metformin did not improve the levels of hormones in comparison with the diabetic group. Conversely, all GPAE treatment groups (14 days after treatment) showed increasing trends of testosterone, LH, and FSH hormone levels, specifically at the dosage of 450 mg/kg (0.802 ± 0.014 ng/mL, 0.041 ± 0.006 ng/mL, and 0.045 ± 0.008 ng/mL, respectively) (Table 1).
Table 1.
The effect of G. procumbens treatment on androgen hormone levels (luteinizing, follicle stimulating, and testosterone hormones) of diabetic male rats. Different letters beside the numbers suggest significantly different according to one-way ANOVA (p < 0.05).
| Group | Testosterone hormone (ng/mL) | Luteinizing hormone (ng/mL) | Follicle-stimulating hormone (ng/mL) |
|---|---|---|---|
| Normal | 0.757 ± 0.014b,c | 0.042 ± 0.002b,c | 0.037 ± 0.008b,c |
| Diabetes | 0.151 ± 0.010 | 0.017 ± 0.002 | 0.020 ± 0.008 |
| Metformin | 0.179 ± 0.009 | 0.021 ± 0.002 | 0.021 ± 0.004b |
| 150 mg/kg | 0.754 ± 0.008b,c | 0.031 ± 0.003b,c | 0.027 ± 0.007b,c |
| 300 mg/kg | 0.746 ± 0.009b,c | 0.032 ± 0.006b,c | 0.033 ± 0.008b,c |
| 450 mg/kg | 0.802 ± 0.014a,b,c | 0.041 ± 0.006b,c | 0.045 ± 0.008a,b,c |
aSignificantly increased compared to normal control (p<0.05); bsignificantly increased compared to diabetic control (p<0.05); csignificantly increased compared to metformin-treated diabetic control (p<0.05).
3.2. GPAE Treatment Improved Testicular Impairment and Number of Implantation Sites
Testicular tissue sections are shown in Figure 1. On the basis of the testes histology, the normal group showed normal spermatogenesis (Figure 1(a)), whereas the diabetic control demonstrated disrupted spermatogenesis as the lumen of seminiferous tubule was almost empty (Figure 1(b)). Similarly, the same histology could be seen in the metformin-treated diabetic control group. Alternatively, the GPAE-treated groups (150, 300, and 450 mg/kg) showed significant changes in the testes histology with the appearance of normal Sertoli and Leydig cells and undisrupted spermatogenesis (Figures 1(d)–1(f)).
Figure 1.

Cross-section of testes for each group at magnification 400×. (a) Normal (nontreated) control, (b) diabetic control (nontreated diabetic), (c) metformin-treated diabetic as well as Gynura procumbens aqueous extract (GPAE) treatment; (d) 150 mg/kg, (e) 300 mg/kg, and (f) 450 mg/kg on diabetic rats.
Furthermore, the number of implantation sites was observed in the uterine horn after 16 days of gestation (Table 2, Figure 2). The number of implantation sites in the normal control was 10 compared with none in the diabetic control rats. For the metformin-treated diabetic group, the number of implantation sites was still low (three sites). Conversely, the implantation sites in the females copulated with GPAE-treated groups increased significantly in the range of 7 to 11 sites, and the highest number was observed in the 450 mg/kg dosage group (Table 2).
Table 2.
The effect of G. procumbens aqueous extract treatment on the number of female implantation sites upon copulation with the treated group male rats. Different letters beside the numbers suggest significant difference according to one-way ANOVA (p < 0.05).
| Group | Number of implantation sites |
|---|---|
| Normal | 10 ± 0.62 |
| Diabetes | 00 ± 0.29 |
| Metformin | 03 ± 1.06a |
| 150 mg/kg | 07 ± 0.49a,b |
| 300 mg/kg | 06 ± 0.36a,b |
| 450 mg/kg | 11 ± 0.57a,b |
aSignificantly increased compared to diabetic control (p<0.05); bsignificantly increased compared to metformin-treated diabetic control (p<0.05).
Figure 2.
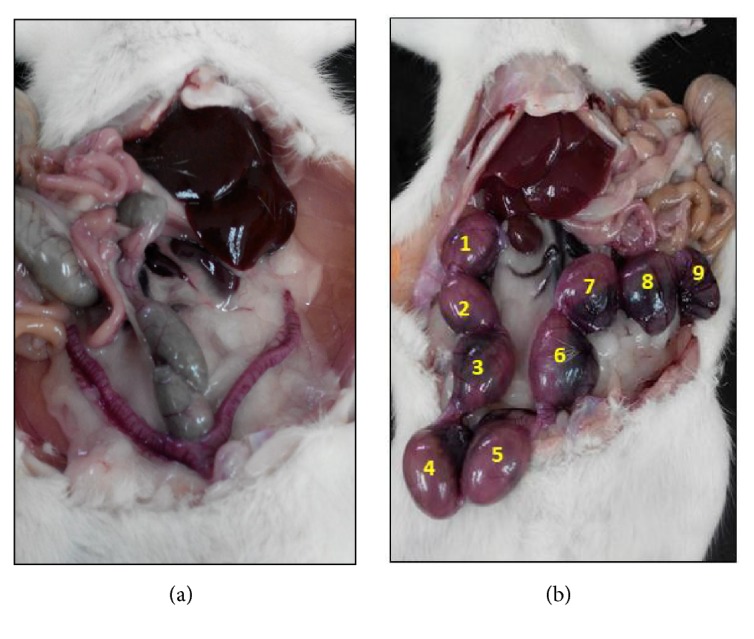
Representative figures of the number of implantation sites after mated with male rats from the (a) diabetic group (nontreated) and (b) Gynura procumbens aqueous extract (GPAE) treated group (450 mg/kg).
3.3. Shotgun Proteomics Identified Proteins Related to Various Biological Processes
To study the effect of GPAE on the sperm proteins, a shotgun proteomics approach was performed. The 450 mg/kg dose group was used, because it was the most effective treatment compared with the other three control groups. A total of 666 proteins were identified in the first initial run (data not shown) and through a stringent search using MaxQuant software, a total of 88 proteins were found in all groups (Supplementary Table S1). For better insight of the identified proteins, they were annotated using Blast2GO software and classified into cellular component, biological process, and molecular function (Figure 3).
Figure 3.

Classification of 88 sperm proteins by gene ontology: cellular component, molecular function, and biological process.
Most of the identified proteins in the biological processes played roles in the metabolic (79.5%), developmental (61.4%), biological regulation (65.9%), and reproduction (26.1%) (Figure 3). Furthermore, the most common molecular activities of the identified proteins were binding (80.7%), catalytic (64.8%), and transporter activities (15.9%) (Figure 3). On the basis of the results, the identified proteins manifested diverse biological roles and molecular activities, specifically in the metabolic pathway.
The 88 identified proteins were then classified according to their biological functions. All proteins were categorised into four general groupings: reproduction, sperm development, locomotion, and sperm metabolic process (Table 3). Fifteen proteins were found to play some roles in reproduction, including cysteine-rich secretory protein-1, zona pellucida binding protein, and outer dense fibre protein. Meanwhile, 10 proteins were involved in the sperm developmental process including carboxylic ester hydrolase, enolase, and lipocalin-5 (Table 3). Eight proteins including tubulin, l-lactate dehydrogenase, and phosphoglycerate kinase were categorised in sperm locomotion (Table 3). Many of the proteins (55) were involved in the metabolic process (Table 3), which is an important event in the living body, particularly in sperm. A large amount of energy is needed for sperm motility and fertility. The metabolic process includes TCA cycle, glycolysis, purine and reductase metabolism. Several proteins involved in these processes were found in this study including glyceraldehyde-3-phosphate, pyruvate dehydrogenase, and glucose-6-phosphate (Table 3).
Table 3.
Classification of 88 identified sperm proteins according to their biological processes. All proteins have Q value of 0 (Supplementary Table 1).
| No | ID | Protein names | Peptides | Sequence Coverage | Molecular Weight | Score | MS/MS Count |
|---|---|---|---|---|---|---|---|
| Proteins involved in reproduction | |||||||
|
| |||||||
| 1 | P12020 | Cysteine-rich secretory protein 1 (32 kDa epididymal protein) (Acidic epididymal glycoprotein) (Protein D) (Protein E) (Protein IV) (Sialoprotein) (Sperm-coating glycoprotein) (SCP) | 8 | 32.1 | 27.847 | 89.627 | 78 |
| 2 | P31044 | Phosphatidylethanolamine-binding protein 1 (PEBP-1) (23 kDa morphine-binding protein) (HCNPpp) (P23K) [Cleaved into: Hippocampal cholinergic neurostimulating peptide (HCNP)] | 8 | 73.8 | 20.801 | 314.36 | 111 |
| 3 | Q6AXU2 | Zona pellucida binding protein, isoform CRA_b (Zona pellucida-binding protein) | 17 | 40.3 | 45.129 | 323.31 | 188 |
| 4 | A0A0H2UHA4 | Mitochondria-eating protein | 12 | 17.4 | 67.794 | 145.83 | 37 |
| 5 | M0R4L7 | Histone H2B | 6 | 48.4 | 13.91 | 85.052 | 41 |
| 6 | A0A0H2UHP1 | Retinal dehydrogenase 1 | 17 | 34 | 56.589 | 323.31 | 146 |
| 7 | D3ZE94 | Outer dense fiber of sperm tails 3 (Outer dense fiber of sperm tails 3 (Predicted)) | 13 | 53.1 | 27.879 | 135.33 | 72 |
| 8 | D4ACV3 | Histone H2A | 6 | 43.7 | 13.645 | 323.31 | 52 |
| 9 | M0R660 | Glyceraldehyde-3-phosphate dehydrogenase (EC 1.2.1.12) | 13 | 38.1 | 35.783 | 323.31 | 83 |
| 10 | P18163 | Long-chain-fatty-acid--CoA ligase 1 (EC 6.2.1.3) (Long-chain acyl-CoA synthetase 1) (LACS 1) (Long-chain-fatty-acid--CoA ligase, liver isozyme) | 19 | 37.1 | 78.178 | 323.31 | 109 |
| 11 | P55063 | Heat shock 70 kDa protein 1-like (Heat shock 70 kDa protein 1L) (Heat shock 70 kDa protein 3) (HSP70.3) | 27 | 43.7 | 70.548 | 323.31 | 202 |
| 12 | G3V7X0 | Outer dense fiber of sperm tails 2, isoform CRA_e (Outer dense fiber protein 2) | 60 | 65 | 81.346 | 323.31 | 360 |
| 13 | Q4KLL5 | Ropporin-1 (Rhophilin-associated protein 1) | 10 | 58.5 | 23.96 | 322.26 | 67 |
| 14 | Q6AXR4 | Beta-hexosaminidase subunit beta (EC 3.2.1.52) (Beta-N-acetylhexosaminidase subunit beta) (Hexosaminidase subunit B) (N-acetyl-beta-glucosaminidase subunit beta) | 18 | 26.6 | 61.527 | 198.66 | 66 |
| 15 | Q6P502 | T-complex protein 1 subunit gamma (TCP-1-gamma) (CCT-gamma) | 11 | 30.1 | 60.646 | 186.2 | 52 |
|
| |||||||
| Proteins involved in sperm development process | |||||||
|
| |||||||
| 1 | F1M786 | Carboxylic ester hydrolase (EC 3.1.1.-) | 14 | 22.5 | 61.398 | 323.31 | 107 |
| 2 | P06911 | Epididymal-specific lipocalin-5 (Androgen-dependent epididymal 18.5 kDa protein) (Epididymal retinoic acid-binding protein) (E-RABP) (Epididymal secretory protein I) (ESP-I) [Cleaved into: Epididymal-specific lipocalin-5, B form; Epididymal-specific lipocalin-5, C form] | 9 | 58 | 20.67 | 235.53 | 89 |
| 3 | A0A0G2K3K2 | Actin, cytoplasmic 1 | 22 | 62.7 | 41.792 | 323.31 | 457 |
| 4 | G3V7J0 | Aldehyde dehydrogenase family 6, subfamily A1, isoform CRA_b (Methylmalonate-semialdehyde dehydrogenase [acylating], mitochondrial) | 12 | 35.1 | 57.747 | 205 | 70 |
| 5 | G3V9D8 | Carboxylic ester hydrolase (EC 3.1.1.-) | 11 | 24.8 | 62.17 | 323.31 | 104 |
| 6 | P04905 | Glutathione S-transferase Mu 1 (EC 2.5.1.18) (GST 3-3) (GSTM1-1) (Glutathione S-transferase Yb-1) (GST Yb1) | 12 | 55.5 | 25.914 | 195.33 | 56 |
| 7 | P11951 | Cytochrome c oxidase subunit 6C-2 (Cytochrome c oxidase polypeptide VIc-2) | 3 | 36.8 | 8.4548 | 38.138 | 31 |
| 8 | Q6YFQ1 | Cytochrome c oxidase subunit 6B2 (Cytochrome c oxidase subunit VIb isoform 2) (COX VIb-2) (Cytochrome c oxidase subunit VIb, testis-specific isoform) | 4 | 46.6 | 10.472 | 28.904 | 28 |
| 9 | P10715 | Cytochrome c, testis-specific | 13 | 77.1 | 11.742 | 323.31 | 114 |
| 10 | Q5BJ93 | Enolase 1, (Alpha) (RCG31027, isoform CRA_a) | 35 | 77.2 | 47.127 | 323.31 | 472 |
|
| |||||||
| Proteins involved in locomotion | |||||||
|
| |||||||
| 1 | G3V7C6 | Tubulin beta chain | 44 | 81.6 | 61.186 | 323.31 | 1073 |
| 2 | Q6AYX2 | L-lactate dehydrogenase (EC 1.1.1.27) | 22 | 72.3 | 35.712 | 323.31 | 195 |
| 3 | B5DEN4 | L-lactate dehydrogenase (EC 1.1.1.27) | 8 | 28 | 36.45 | 323.31 | 47 |
| 4 | P68370 | Tubulin alpha-1A chain (Alpha-tubulin 1) (Tubulin alpha-1 chain) [Cleaved into: Detyrosinated tubulin alpha-1A chain] | 21 | 47 | 50.135 | 20.5 | 23 |
| 5 | Q5XIV1 | Phosphoglycerate kinase (EC 2.7.2.3) | 36 | 75.8 | 45.01 | 323.31 | 545 |
| 6 | Q6AYM2 | Tektin-2 (Tektin-t) (Testicular tektin) | 6 | 17 | 50.296 | 54.918 | 23 |
| 7 | P63269 | Actin, gamma-enteric smooth muscle (Alpha-actin-3) (Gamma-2-actin) (Smooth muscle gamma-actin) [Cleaved into: Actin, gamma-enteric smooth muscle, intermediate form] | 20 | 60.6 | 41.876 | 173.1 | 38 |
| 8 | Q68FR8 | Tubulin alpha-3 chain (Alpha-tubulin 3) [Cleaved into: Detyrosinated tubulin alpha-3 chain] | 24 | 54.7 | 49.959 | 323.31 | 363 |
|
| |||||||
| Proteins involved in metabolism | |||||||
|
| |||||||
| 1 | G3V8X9 | Serine (or cysteine) peptidase inhibitor, clade A (alpha-1 antiproteinase, antitrypsin), member 16 (Serine proteinase inhibitor HongrES1) | 24 | 48.2 | 47.181 | 323.31 | 192 |
| 2 | M0R8P3 | Calcium-binding tyrosine phosphorylation-regulated (RCG24939, isoform CRA_c) | 7 | 30.8 | 43.044 | 131.18 | 57 |
| 3 | O88767 | Protein/nucleic acid deglycase DJ-1 (EC 3.1.2.-) (EC 3.5.1.-) (EC 3.5.1.124) (Contraception-associated protein 1) (Protein CAP1) (Fertility protein SP22) (Maillard deglycase) (Parkinson disease protein 7 homolog) (Parkinsonism-associated deglycase) (Protein DJ-1) (DJ-1) | 8 | 33.9 | 19.974 | 173.44 | 77 |
| 4 | P00507 | Aspartate aminotransferase, mitochondrial (mAspAT) (EC 2.6.1.1) (EC 2.6.1.7) (Fatty acid-binding protein) (FABP-1) (Glutamate oxaloacetate transaminase 2) (Kynurenine aminotransferase 4) (Kynurenine aminotransferase IV) (Kynurenine--oxoglutarate transaminase 4) (Kynurenine--oxoglutarate transaminase IV) (Plasma membrane-associated fatty acid-binding protein) (FABPpm) (Transaminase A) | 14 | 44.7 | 47.314 | 218.74 | 98 |
| 5 | P02091 | Hemoglobin subunit beta-1 (Beta-1-globin) (Hemoglobin beta chain, major-form) (Hemoglobin beta-1 chain) | 21 | 70.7 | 15.979 | 323.31 | 481 |
| 6 | P02770 | Serum albumin | 52 | 74.5 | 68.73 | 323.31 | 688 |
| 7 | P04636 | Malate dehydrogenase, mitochondrial (EC 1.1.1.37) | 11 | 53.8 | 35.683 | 130.31 | 33 |
| 8 | P05065 | Fructose-bisphosphate aldolase A (EC 4.1.2.13) (Muscle-type aldolase) | 19 | 59.3 | 39.351 | 323.31 | 127 |
| 9 | P06761 | Endoplasmic reticulum chaperone BiP (EC 3.6.4.10) (78 kDa glucose-regulated protein) (GRP-78) (Binding-immunoglobulin protein) (BiP) (Heat shock protein 70 family protein 5) (HSP70 family protein 5) (Heat shock protein family A member 5) (Immunoglobulin heavy chain-binding protein) (Steroidogenesis-activator polypeptide) | 21 | 33 | 72.346 | 323.31 | 184 |
| 10 | P09606 | Glutamine synthetase (GS) (EC 6.3.1.2) (Glutamate decarboxylase) (EC 4.1.1.15) (Glutamate--ammonia ligase) | 10 | 30 | 42.267 | 323.31 | 156 |
| 11 | P10760 | Adenosylhomocysteinase (AdoHcyase) (EC 3.3.1.1) (S-adenosyl-L-homocysteine hydrolase) | 3 | 6.9 | 47.538 | 122.35 | 23 |
| 12 | P10818 | Cytochrome c oxidase subunit 6A1, mitochondrial (Cytochrome c oxidase polypeptide VIa-liver) | 5 | 63.1 | 12.301 | 63.576 | 48 |
| 13 | P10860 | Glutamate dehydrogenase 1, mitochondrial (GDH 1) (EC 1.4.1.3) (Memory-related gene 2 protein) (MRG-2) | 9 | 17 | 61.415 | 177.81 | 83 |
| 14 | P11517 | Hemoglobin subunit beta-2 (Beta-2-globin) (Hemoglobin beta chain, minor-form) (Hemoglobin beta-2 chain) | 18 | 64.6 | 15.982 | 323.31 | 158 |
| 15 | P11980 | Pyruvate kinase PKM (EC 2.7.1.40) (Pyruvate kinase muscle isozyme) | 15 | 40.1 | 57.817 | 323.31 | 84 |
| 16 | P12346 | Serotransferrin (Transferrin) (Beta-1 metal-binding globulin) (Liver regeneration-related protein LRRG03) (Siderophilin) | 17 | 28.4 | 76.394 | 317.85 | 139 |
| 17 | P16290 | Phosphoglycerate mutase 2 (EC 5.4.2.11) (EC 5.4.2.4) (BPG-dependent PGAM 2) (Muscle-specific phosphoglycerate mutase) (Phosphoglycerate mutase isozyme M) (PGAM-M) | 16 | 71.1 | 28.755 | 323.31 | 155 |
| 18 | P19804 | Nucleoside diphosphate kinase B (NDK B) (NDP kinase B) (EC 2.7.4.6) (Histidine protein kinase NDKB) (EC 2.7.13.3) (P18) | 3 | 38.8 | 17.283 | 55.113 | 20 |
| 19 | P20760 | Ig gamma-2A chain C region | 10 | 32.6 | 35.185 | 172.46 | 82 |
| 20 | P32551 | Cytochrome b-c1 complex subunit 2, mitochondrial (Complex III subunit 2) (Core protein II) (Ubiquinol-cytochrome-c reductase complex core protein 2) | 4 | 20.1 | 48.396 | 323.31 | 29 |
| 21 | P46462 | Transitional endoplasmic reticulum ATPase (TER ATPase) (EC 3.6.4.6) (15S Mg(2+)-ATPase p97 subunit) (Valosin-containing protein) (VCP) | 23 | 41.8 | 89.348 | 323.31 | 136 |
| 22 | Q4KLZ6 | Triokinase/FMN cyclase (Bifunctional ATP-dependent dihydroxyacetone kinase/FAD-AMP lyase (cyclizing)) [Includes: ATP-dependent dihydroxyacetone kinase (DHA kinase) (EC 2.7.1.28) (EC 2.7.1.29) (Glycerone kinase) (Triokinase) (Triose kinase); FAD-AMP lyase (cyclizing) (EC 4.6.1.15) (FAD-AMP lyase (cyclic FMN forming)) (FMN cyclase)] | 8 | 27.7 | 59.443 | 202.32 | 59 |
| 23 | Q4QR77 | Protein FAM166A | 4 | 18.4 | 36.993 | 102.93 | 31 |
| 24 | Q4V8H5 | Aspartyl aminopeptidase (Aspartyl aminopeptidase, isoform CRA_c) | 9 | 24.2 | 52.555 | 323.31 | 60 |
| 25 | Q4V8P4 | Rsb-66 protein (Rsb-66 protein, isoform CRA_b) (Sperm acrosome-associated 9) | 3 | 26.2 | 19.465 | 323.31 | 56 |
| 26 | Q5RK28 | Normal mucosa of esophagus-specific gene 1 protein | 9 | 83.1 | 9.5981 | 275.93 | 78 |
| 27 | Q5XI62 | Protein MENT (Methylated in normal thymocytes protein) | 10 | 45.3 | 37.64 | 246.1 | 65 |
| 28 | Q6AXN7 | 5'-nucleotidase, cytosolic IB | 21 | 39.5 | 64.922 | 323.31 | 134 |
| 29 | Q6AXX6 | Redox-regulatory protein FAM213A (Peroxiredoxin-like 2 activated in M-CSF stimulated monocytes) (Protein PAMM) (Sperm head protein 1) | 14 | 50.7 | 25.763 | 240.02 | 79 |
| 30 | Q6AY07 | Fructose-bisphosphate aldolase (EC 4.1.2.13) | 20 | 56.6 | 39.491 | 323.31 | 220 |
| 31 | Q6AY30 | Saccharopine dehydrogenase-like oxidoreductase (EC 1.-.-.-) | 12 | 38.5 | 47.088 | 204.86 | 50 |
| 32 | Q6P6V0 | Glucose-6-phosphate isomerase (GPI) (EC 5.3.1.9) (Autocrine motility factor) (AMF) (Neuroleukin) (NLK) (Phosphoglucose isomerase) (PGI) (Phosphohexose isomerase) (PHI) | 14 | 38.7 | 62.826 | 323.31 | 73 |
| 33 | Q6P762 | Alpha-mannosidase (EC 3.2.1.-) | 23 | 29.7 | 114.33 | 323.31 | 113 |
| 34 | Q8VI04 | Isoaspartyl peptidase/L-asparaginase (EC 3.4.19.5) (EC 3.5.1.1) (Asparaginase-like protein 1) (Asparaginase-like sperm autoantigen) (Beta-aspartyl-peptidase) (Glial asparaginase) (Isoaspartyl dipeptidase) (L-asparagine amidohydrolase) [Cleaved into: Isoaspartyl peptidase/L-asparaginase alpha chain; Isoaspartyl peptidase/L-asparaginase beta chain] | 14 | 49.8 | 34.41 | 323.31 | 45 |
| 35 | A0A0G2KAM3 | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial | 4 | 11.1 | 46.192 | 272.59 | 17 |
| 36 | A0A0G2JSJ3 | Solute carrier family 2 (Facilitated glucose transporter), member 3 (Solute carrier family 2, facilitated glucose transporter member 3-like) | 5 | 14 | 53.562 | 151.67 | 31 |
| 37 | A0A0G2JSJ8 | Fucosidase, alpha-L-1, tissue, isoform CRA_a (Tissue alpha-L-fucosidase) | 5 | 15.4 | 53.472 | 286.49 | 63 |
| 38 | A0A0G2JSV6 | Globin c2 (Hemoglobin alpha, adult chain 2) (RCG34342, isoform CRA_a) | 17 | 69.7 | 15.284 | 323.31 | 587 |
| 39 | A0A0G2JSZ5 | Protein disulfide-isomerase A6 (RCG62282, isoform CRA_a) | 6 | 17.3 | 48.76 | 96.801 | 26 |
| 40 | A0A0G2JTW9 | Hemoglobin, beta adult major chain | 10 | 70.7 | 15.988 | 138.77 | 32 |
| 41 | G3V8Q6 | cAMP-dependent protein kinase type II-alpha regulatory subunit | 6 | 29.9 | 45.48 | 173.45 | 23 |
| 42 | D4AA52 | Alpha-1-inhibitor III | 8 | 6.3 | 163.6 | 53.696 | 34 |
| 43 | A0A0H2UHE1 | Succinate--CoA ligase [ADP/GDP-forming] subunit alpha, mitochondrial (EC 6.2.1.4) (EC 6.2.1.5) (Succinyl-CoA synthetase subunit alpha) (SCS-alpha) | 6 | 21.1 | 37.559 | 218 | 38 |
| 44 | A0A0H2UHM5 | Protein disulfide-isomerase (EC 5.3.4.1) | 22 | 49 | 57.078 | 272.63 | 83 |
| 45 | B0K020 | CDGSH iron-sulfur domain-containing protein 1 (MitoNEET) | 6 | 57.4 | 12.097 | 95.47 | 82 |
| 46 | B1H216 | Globin c3 (Hemoglobin alpha, adult chain 2) (RCG34636, isoform CRA_a) | 16 | 69.7 | 15.328 | 318.53 | 41 |
| 47 | B6DYP8 | Glutathione S-transferase (EC 2.5.1.18) | 9 | 55.2 | 25.319 | 150.96 | 69 |
| 48 | D3Z9F9 | Similar to RIKEN cDNA 4930540L03 (Predicted) (Sperm acrosome-associated 1) | 5 | 25.4 | 37.335 | 302.34 | 54 |
| 49 | D3ZLJ6 | Amine oxidase (EC 1.4.3.-) | 5 | 11.7 | 71.083 | 31.503 | 33 |
| 50 | D3ZUM4 | Beta-galactosidase (EC 3.2.1.23) | 21 | 33.5 | 73.227 | 224.16 | 110 |
| 51 | D4A4R7 | RCG21015, isoform CRA_a (Serine (or cysteine) peptidase inhibitor, clade A, member 1F) | 4 | 16.5 | 46.985 | 33.071 | 36 |
| 52 | F1LML2 | Polyubiquitin-C | 8 | 90.2 | 91.072 | 323.31 | 98 |
| 53 | F1LN88 | Aldehyde dehydrogenase, mitochondrial (RCG21519, isoform CRA_a) | 16 | 44.7 | 56.516 | 323.31 | 103 |
| 54 | F1LP05 | ATP synthase subunit alpha | 17 | 25.9 | 59.812 | 303.31 | 85 |
| 55 | G3V6D3 | ATP synthase subunit beta (EC 3.6.3.14) | 21 | 57.8 | 56.344 | 323.31 | 153 |
3.4. Differentially Expressed Proteins Highlight GPAE Effect on Spermatogenic Proteins
Quantification analysis was performed using MaxQuant software to compare protein levels between the experimental groups, yielding four statistically significant (p < 0.05) protein changes between nontreated diabetic rats and GPAE-treated diabetic rats (Figure 4). These proteins were carboxylesterase (CES5A), cysteine-rich secretory protein 1 (CRISP1), phosphatidylethanolamine-binding protein 1 (PEBP1), and zona pellucida binding protein (ZPBP) (Figures 4(a)–4(d)).
Figure 4.

Four significantly expressed proteins in Gynura procumbens aqueous extract (GPAE) treatment compared to the diabetic nontreated group. These proteins including carboxylesterase (CES5A) (a), cysteine-rich secretory protein 1 (CRISP1) (b), phosphatidylethanolamine-binding protein1 (PEBP1) (c), and zona pellucida binding protein (ZPBP) (d) are related to sperm development and sperm-egg interaction.
CES5A protein level significantly increased in GPAE-treated groups (23.21 ± 0.48) compared with both diabetic and metformin-treated diabetic control, which was at 20.42 ± 0.49 and 22.60 ± 0.77, respectively. The protein level was matched with the normal control at 23.62 ± 0.54. Metformin-treated diabetic control showed a slight increase of protein level compared with diabetic control; however, the level was not significant. For CRISP1, GPAE-treated groups showed a significant increase of protein level (23.24 ± 0.58) compared with both diabetic (20.32 ± 0.22) and metformin-treated diabetic control (21.50 ± 0.81). The third protein found to be significantly expressed after treatment with GPAE was PEBP1. This protein was at a higher level in GPAE-treated groups (23.48 ± 0.84) compared with diabetic and metformin-treated diabetic control. The protein level matched with normal control (23.84 ± 0.66). Both diabetic and metformin-treated diabetic control showed lower protein level at 20.75 ± 0.48 and 21.57 ± 0.89, respectively. In addition, protein level of ZPBP was the highest in GPAE-treated groups at 24.19 ± 0.21, compared with diabetic (21.00 ± 0.59) and metformin-treated diabetic control (22.67 ± 0.21). All these proteins were highly expressed in both normal control as well as 450 mg/kg dose GPAE-treated group, in comparison with the nontreated diabetic rats. Although the average protein levels could be higher for metformin-treated diabetic rats compared with nontreated diabetic rats, most levels were not significantly different and not as high as either or both normal control and GPAE-treated rats (Figure 4).
4. Discussion
Diabetes mellitus has always been linked to reproductive dysfunction, mainly in the male reproductive system. Many studies have revealed that diabetes mellitus can affect spermatogenesis and steroidogenesis [20, 27]. In this study, the impact of GPAE herbal treatment on diabetic rats was assessed in regard to the androgen hormone levels, testes histology, and fertilization (implantation sites) as well as sperm protein level.
Diabetes mellitus has been shown to reduce androgen hormones such as LH, FSH, and testosterone [28]. LH and FSH are the main regulatory hormones used for the stimulation of steroid hormones including testosterone and gametogenesis in both men and women [4]. In the current study, LH, FSH, and testosterone levels were significantly elevated in the GPAE-treated groups compared with the diabetic control (nontreated diabetic) group (Table 1). It appears that the GPAE treatment significantly increases the androgen hormone levels to match the normal control group level. The role of FSH is to stimulate Sertoli cells in testes to provide nutrients for sperm throughout the spermatogenesis process [4], while LH is responsible for functional Leydig cells and testosterone production [28]. The increase of both LH and FSH levels (Table 1) as well as reduced hyperglycaemia in GPAE-treated rats may repair the Leydig cells, thus, increasing the testosterone level. As a consequence, a functional male reproductive system can be regenerated, assisting spermatogenesis and the testicular structure regeneration [29].
Evidently, the testes histology of the GPAE-treated group has been improved with Sertoli and Leydig cells regenerated and the sperm in the lumen restored (Figures 1(d)–1(f)) compared with nontreated diabetic rats (Figure 1(b)). This is consistent with our proteomics finding which has identified the increase of CES5A protein level in diabetic rats after GPAE treatment (Figure 4(a)). CES5A is known to be expressed in the corpus and caudal epididymis, as well as being secreted in the lumen [30]. It was reported that the regulation of the lipid environment by CES5A in the epididymal lumen is crucial for sperm maturation and storage [30]. Furthermore, another protein known as PEBP1 was significantly increased in the GPAE-treated group compared with the nontreated diabetic rats (Figure 4(c)). PEBP1 is involved in sperm development and capacitation [31]. It also plays a role as a decapacitation factor (DF) receptor [31] which is vital in the regulation of sperm function. Hence, the higher CES5A and PEBP1 levels in GPAE-treated rats may suggest the profertility impact of the herbal treatment on diabetic rats, particularly with regard to the regeneration of testes structure and spermatogenesis.
GPAE treatment was also shown to significantly increase sperm quality including sperm count, motility, viability, and morphology compared with the diabetic control group [14]. As such, this has also improved the fertility of the male rats as evidence by the significant number of implantation sites (7 to 11 sites) in fertilized female rats compared with the diabetic control group (Table 2). Congruently, a few differentially expressed proteins identified in this study were involved in fertilization process. For example, ZPBP was found to be significantly expressed in GPAE-treated group compared with diabetic and metformin-treated diabetic control groups (Figure 4(d)). ZPBP is localised to the acrosome region and is important for the binding of sperm to the zona pellucida of the oocyte [32, 33]. The binding of sperm to zona pellucida membrane is one of the most crucial parts in sperm-egg interaction which leads to a successful penetration. Another important protein for fertilization is CRISP1 which was also significantly expressed upon GPAE treatment (Figure 4(b)). CRISP1 was reported to participate in sperm-egg fusion through the interaction with complementary sites on the surface of the egg [34, 35]. The increase of both ZPBP and CRISP1 in GPAE-treated diabetic rats implies that the herbal treatment was effective in increasing fertility in diabetic rats of which the changes can be implicated at the molecular protein level.
In contrast, metformin treatment demonstrates no improvement in most of our fertility tests (Figure 1, Tables 1 and 2). Previous studies showed that the drug was able to reduce the diabetic rat's blood sugar [12, 14], signifying that metformin was only effective against diabetes. This is further corroborated by the observed levels of androgen hormones (LH, FSH, and testosterone) which were not significantly higher than the nontreated diabetic rats (Table 2). Without improvement in these hormones, the repair and regeneration processes may not be able to occur, leading to degenerated Leydig cells and defective testes histology (Figure 1(c)). Consequently, the fertility of the male rats was not effectively recovered, as observed by the low number of implantation sites (only three sites) in female rats (Table 2).
5. Conclusion
In conclusion, the results of this study provide evidence that GPAE may improve androgen hormone levels, testicular regeneration, and fertility (number of implantation sites). Proteomic analysis further implies that GPAE improved the fertility of diabetic male rats via increasing protein expression related to sperm maturation and sperm-egg interaction. Thus, GP may have potential as an antihyperglycaemia and profertility agent for diabetic patients. Further studies are needed to develop the herbal extract as an alternative medication to treat diabetes and fertility problems.
Acknowledgments
The authors would like to thank Malaysian Genome Institute for providing LCMS/MS service for protein identification. This work was supported by the Economic Transformation Programme Research Fund (ETP) Grant (ETP-2013-080), Research University Grant (GUP) (GUP-2016-056), and the Faculty of Science and Technology, Universiti Kebangsaan Malaysia.
Data Availability
All data used to support the findings of this study are included within the supplementary information file.
Conflicts of Interest
The authors declare that there are no conflicts of interest that could be perceived as prejudicing the impartiality of this study.
Supplementary Materials
Supplementary Table 1. A total of 88 sperm proteins identified using MaxQuant and Perseus analysis.
References
- 1.National Health & Morbidity Survey (NHMS) 2015 Institute for Public Health. Fact Sheet. 2015:5–6. [Google Scholar]
- 2.Alves M. G., Rato L., Carvalho R. A., Moreira P. I., Socorro S., Oliveira P. F. Hormonal control of Sertoli cell metabolism regulates spermatogenesis. Cellular and Molecular Life Sciences. 2013;70(5):777–793. doi: 10.1007/s00018-012-1079-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Soudamani S., Yuvaraj S., Malini T., Balasubramanian K. Experimental diabetes has adverse effects on the differentiation of ventral prostate during sexual maturation of rats. The Anatomical Record Part A: Discoveries in Molecular, Cellular, and Evolutionary Biology. 2005;287(2):1281–1289. doi: 10.1002/ar.a.20250. [DOI] [PubMed] [Google Scholar]
- 4.Arikawe A. P., Oyerinde A., Olatunji-Bello I. I., Obika L. F. O. Streptozotocin diabetes and insulin resistance impairment of spermatogenesis in adult rat testis: Central vs local mechanism. Nigerian Journal of Physiological Sciences . 2012;27(2):171–179. [PubMed] [Google Scholar]
- 5.FogliaVG Rosner JM., Ramos M., Lema BE. Sexual disturbances in the male diabetic rat. Hormone and Metabolic Research. 1996;1:72–77. doi: 10.1055/s-0028-1095155. [DOI] [PubMed] [Google Scholar]
- 6.Aitken R. J., Baker M. A. Oxidative stress, sperm survival and fertility control. Molecular and Cellular Endocrinology. 2006;250:p. 669. doi: 10.1016/j.mce.2005.12.026. [DOI] [PubMed] [Google Scholar]
- 7.Hakim P., Sani H. A., Noor M. M. Effects of Gynura procumbens on sperm quality and testosterone level in streptozotocin-induced type 1 diabetic rats. International Journal of Pharmacognosy and Phytochemical Research. 2016;8:22–30. [Google Scholar]
- 8.Hoe S.-Z., Lee C.-N., Mok S.-L., Kamaruddin M. Y., Lam S.-K. Gynura procumbens Merr. decreases blood pressure in rats by vasodilatation via inhibition of calcium channels. Clinics. 2011;66(1):143–150. doi: 10.1590/s1807-59322011000100025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ng H.-K., Poh T.-F., Lam S.-K., Hoe S.-Z. Potassium channel openers and prostacyclin play a crucial role in mediating the vasorelaxant activity of Gynura procumbens. BMC Complementary and Alternative Medicine. 2013;13 doi: 10.1186/1472-6882-13-188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kaur N., Kumar R., Yam M. F., Sadikun A., Sattar A. M. Z., Asmawi M. Z. Antihypertensive effect of gynura procumbens water extract in spontaneously hypertensive rats. International Journal of Applied Research in Natural Products. 2013;6(3):20–27. [Google Scholar]
- 11.Nisa F., Hermawan A., Murwanti R., Meiyanto E. Antiproliferative effect of gynura procumbens (lour.) Merr. Leaves etanolic extract on 7,12-dimethylbenz(a)antracene induced male rat liver. Advanced Pharmaceutical Bulletin (APB) 2012;2(1):99–106. doi: 10.5681/apb.2012.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Hassan Z., Yam M. F., Ahmad M., Yusof A. P. M. Antidiabetic properties and mechanism of action of gynura procumbens water extract in streptozotocin-induced diabetic rats. Molecules. 2010;15(12):9008–9023. doi: 10.3390/molecules15129008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Noor M. M., Radzuan N. R. M. Anti-hyperglycemic effect of gynura procumbens methanolic extract on fertility and libido of induced diabetic male rats. SAINS MALAYSIANA. 2012;41(12):1549–1556. [Google Scholar]
- 14.Kamaruzaman K. A., MatNoor M. Gynura procumbens leaf improves blood glucose level, restores fertility and libido of diabetic-induced male rats. SAINS MALAYSIANA. 2017;46(9):1471–1477. doi: 10.17576/jsm-2017-4609-16. [DOI] [Google Scholar]
- 15.Yunianto I., Bashah N., Noor M. Antifertility properties of Centella asiatica ethanolic extract as a contraceptive agent: Preliminary study of sperm proteomic. Asian Pacific Journal of Reproduction. 2017;6(5):212–216. doi: 10.4103/2305-0500.215931. [DOI] [Google Scholar]
- 16.Luthfi M. J., Kamalrudin A., Noor M. M. Effects of Lunasia amara blanco (Sanrego) on male fertility: A preliminary study on sperm proteomic analysis. Journal of Applied Pharmaceutical Science. 2017;7(8):085–091. [Google Scholar]
- 17.Tyanova S., Temu T., Sinitcyn P., et al. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nature Methods. 2016;13(9):731–740. doi: 10.1038/nmeth.3901. [DOI] [PubMed] [Google Scholar]
- 18.Dorus S., Wasbrough E. R., Busby J., Wilkin E. C., Karr T. L. Sperm proteomics reveals intensified selection on mouse sperm membrane and acrosome genes. Molecular Biology and Evolution. 2010;27(6):1235–1246. doi: 10.1093/molbev/msq007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Baker M. A., Hetherington L., Reeves G., Müller J., Aitken R. J. The rat sperm proteome characterized via IPG strip prefractionation and LC-MS/MS identification. Proteomics. 2008;8(11):2312–2321. doi: 10.1002/pmic.200700876. [DOI] [PubMed] [Google Scholar]
- 20.Alvez M. G., Martins A. D., Socorro S. C. J. E., Oliveira P. F. Molecular mechanisms beyond glucose transport in diabetes-related male infertility. Biochimica et Biophysica Acta. 2013:626–635. doi: 10.1016/j.bbadis.2013.01.011. [DOI] [PubMed] [Google Scholar]
- 21.Punithavathi V. R., Prince P. S. M., Kumar R., Selvakumari J. Antihyperglycaemic, antilipid peroxidative and antioxidant effects of gallic acid on streptozotocin induced diabetic Wistar rats. European Journal of Pharmacology. 2011;650(1):465–471. doi: 10.1016/j.ejphar.2010.08.059. [DOI] [PubMed] [Google Scholar]
- 22.Agmo A. Protocol male rat sexual behaviour. Brain Research Protocols. 1997;1:203–209. doi: 10.1016/s1385-299x(96)00036-0. [DOI] [PubMed] [Google Scholar]
- 23.Chauhan A., Agarwal M. Assessment of the contraceptive efficacy of the aqueous extract of Aegel marmelos Corr. leaves in the male albino rats. Human Fertility12. 2009:10–118. doi: 10.1080/14647270802657402. [DOI] [PubMed] [Google Scholar]
- 24.Biggers J. D., Whitten W. K., Whittingham D. The culture of mouse embryos in vitro. In: Daniel J. C., editor. Methods in Mammalian Embryology. San Francisco, CA, USA: Freeman; 1971. pp. 86–116. [Google Scholar]
- 25.Ramm S. A., McDonald L., Hurst J. L., Beynon R. J., Stockley P. Comparative proteomics reveals evidence for evolutionary diversification of rodent seminal fluid and its functional significance in sperm competition. Molecular Biology and Evolution. 2009;26(1):189–198. doi: 10.1093/molbev/msn237. [DOI] [PubMed] [Google Scholar]
- 26.Iovinella I., Caputo B., Michelucci E., Dani F. R., della Torre A. Candidate biomarkers for mosquito age-grading identified by label-free quantitative analysis of protein expression in Aedes albopictus females. Journal of Proteomics. 2015;128:272–279. doi: 10.1016/j.jprot.2015.08.002. [DOI] [PubMed] [Google Scholar]
- 27.Bhattacharya SM., Ghosh M., Nani N. Diabetes mellitus and abnormalities in semen analysis. Journal of Obstetrics Gynaecology Research40. 2014:167–171. doi: 10.1111/jog.12149. [DOI] [PubMed] [Google Scholar]
- 28.La Vignera S., Condorelli R., Vicari E., D'Agata R., Calogero A. E. Diabetes mellitus and sperm parameters. Journal of Andrology. 2012;33(2):145–153. doi: 10.2164/jandrol.111.013193. [DOI] [PubMed] [Google Scholar]
- 29.Khaki A., Khaki A. A., Hajhosseini L., Golzar F. S., Ainehchi N. The anti-oxidant effects of ginger and cinnamon on spermatogenesis dys-function of diabetes rats. African Journal of Traditional, Complementary and Alternative Medicines. 2014;11(4):1–8. doi: 10.4314/ajtcam.v11i4.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Ru Y.-F., Xue H.-M., Ni Z.-M., Xia D., Zhou Y.-C., Zhang Y.-L. An epididymis-specific carboxyl esterase CES5A is required for sperm capacitation and male fertility in the rat. Asian Journal of Andrology. 2015;17(2):292–297. doi: 10.4103/1008-682X.143314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gibbons R., Adeoya-Osiguwa S. A., Fraser L. R. A mouse sperm decapacitation factor receptor is phosphatidylethanolamine-binding protein 1. Reproduction. 2005;130(4):497–508. doi: 10.1530/rep.1.00792. [DOI] [PubMed] [Google Scholar]
- 32.Lin Y. N., Matzuk M. M. High-throughput discovery of germcell-specific genes. Seminars of Reproductive Medicine. 2005;23:201–212. doi: 10.1055/s-2005-872448. [DOI] [PubMed] [Google Scholar]
- 33.Yu Y., Xu W., Yi Y.-J., Sutovsky P., Oko R. The extracellular protein coat of the inner acrosomal membrane is involved in zona pellucida binding and penetration during fertilization: Characterization of its most prominent polypeptide (IAM38) Developmental Biology. 2006;290(1):32–43. doi: 10.1016/j.ydbio.2005.11.003. [DOI] [PubMed] [Google Scholar]
- 34.Cohen D. J., Ellerman D. A., Cuasnicu P. S. Mammalian sperm-egg fusion: Evidence that epididymal protein DE plays a role mouse gamete fusion. Biology of Reproduction. 2000;63(2):462–468. doi: 10.1095/biolreprod63.2.462. [DOI] [PubMed] [Google Scholar]
- 35.Cohen D. J., Rochwerger L., Ellerman D. A., Morgenfeld M. M., Busso D., Cuasnicú P. S. Relationship between the association of rat epididymal protein 'DE' with spermatozoa and the behavior and function of the protein. Molecular Reproduction and Development. 2000;56(2):180–188. doi: 10.1002/(SICI)1098-2795(200006)56:2<180::AID-MRD9>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Table 1. A total of 88 sperm proteins identified using MaxQuant and Perseus analysis.
Data Availability Statement
All data used to support the findings of this study are included within the supplementary information file.