

Abstract
Selective androgen receptor modulators (SARMs) now under development can protect against muscle and bone loss without causing prostate growth or polycythemia. 17β-Hydroxyestra-4,9,11-trien-3-one (trenbolone), a potent testosterone analog, may have SARM-like actions because, unlike testosterone, trenbolone does not undergo tissue-specific 5α-reduction to form more potent androgens. We tested the hypothesis that trenbolone-enanthate (TREN) might prevent orchiectomy-induced losses in muscle and bone and visceral fat accumulation without increasing prostate mass or resulting in adverse hemoglobin elevations. Male F344 rats aged 3 mo underwent orchiectomy or remained intact and were administered graded doses of TREN, supraphysiological testosterone-enanthate, or vehicle for 29 days. In both intact and orchiectomized animals, all TREN doses and supraphysiological testosterone-enanthate augmented androgen-sensitive levator ani/bulbocavernosus muscle mass by 35–40% above shams (P ≤ 0.001) and produced a dose-dependent partial protection against orchiectomy-induced total and trabecular bone mineral density losses (P < 0.05) and visceral fat accumulation (P < 0.05). The lowest doses of TREN successfully maintained prostate mass and hemoglobin concentrations at sham levels in both intact and orchiectomized animals, whereas supraphysiological testosterone-enanthate and high-dose TREN elevated prostate mass by 84 and 68%, respectively (P < 0.01). In summary, low-dose administration of the non-5α-reducible androgen TREN maintains prostate mass and hemoglobin concentrations near the level of shams while producing potent myotrophic actions in skeletal muscle and partial protection against orchiectomy-induced bone loss and visceral fat accumulation. Our findings indicate that TREN has advantages over supraphysiological testosterone and supports the need for future preclinical studies examining the viability of TREN as an option for androgen replacement therapy.
Keywords: androgen, 5α-reductase, steroid, testosterone, selective androgen receptor modulator
in men, hypogonadism (i.e., reduced circulating testosterone) results in a variety of detrimental effects, including the loss of skeletal muscle mass and bone mineral density (BMD) and increased visceral adiposity (52). Testosterone replacement therapy has been proposed as a means of reducing these unfavorable changes (6). However, replacement doses of testosterone result in only minor improvements in muscle strength (39) and BMD (61) and only small reductions in adiposity (27). In contrast, supraphysiological testosterone effectively augments skeletal muscle mass (7, 8, 57), reduces adiposity (65), and increases BMD (2) in men and in orchiectomized animal models (10, 66) but also increases the risk for a variety of side effects, of which polycythemia, prostate enlargement, and increased incidence of prostate biopsy occur most frequently (13). Selective androgen receptor (AR) modulators (SARMs) are currently under development with the aim of producing anabolic effects in skeletal muscle and bone without prostate enlargement, polycythemia, or other androgenic side effects (33).
17β-Hydroxyestra-4,9,11-trien-3-one (trenbolone) is a potent synthetic testosterone analog (67) that does not undergo 5α-reduction to more potent metabolites (46). As such, trenbolone may induce less growth in prostate and other androgenic tissues that highly express 5α-reductase. This is in contrast to testosterone, which has approximately threefold greater potency in androgenic tissues that highly express 5α-reductase (63) due to its conversion to dihydrotestosterone (DHT).
The primary purpose of this study was to determine the effects of trenbolone-enanthate (TREN; a slowly released trenbolone ester) on a variety of androgen-sensitive tissues, including skeletal muscle, bone, visceral adiposity, hemoglobin (Hb), and the prostate of rodents. Because trenbolone is selectively metabolized to weaker androgens in vivo, we hypothesized that TREN will produce dose-dependent anabolic effects in skeletal muscle, bone, and fat that are at least equal to those of supraphysiological testosterone while producing a smaller increase in Hb and less growth of the prostate.
METHODS
Animal Care
Barrier-raised and viral pathogen-free Fischer F344 male rats aged 3 mo were obtained from Charles River Laboratories (Wilmington, MA). Animals were housed individually in a temperature- and light-controlled room on a 12:12-h light-dark cycle. Rats were fed an ad libitum diet of Purina rodent chow containing 3.3 kcal/g distributed as 58.9% carbohydrate, 12.4% fat, and 28.7% protein (no. 5001; Purina Mills, St. Louis, MO) and tap water. All experimental procedures conformed to the Institute for Laboratory Animal Research Guide to the Care and Use of Experimental Animals and were approved by the Institutional Animal Care and Use Committee at the Gainesville Veterans Affairs Medical Center.
Pilot Study Experiment Design: Verify Hormone Delivery and Suppression of Endogenous Sex Hormones
A pilot study was performed to verify delivery of TREN (Steraloids, Newport, RI) following intramuscular administration and to evaluate the suppressive effects of TREN administration on circulating testosterone and DHT concentrations. For this experiment, intact 3-mo-old male Fisher F344 rats received intramuscular injections of either TREN (7.0 mg·wk−1·animal−1) or vehicle (sesame oil) at baseline and 7 days later (n = 5/group). Blood was sampled via tail tip amputation at baseline and every 2 days thereafter. Ten days following the initial injection, the rats were euthanized by an intraperitoneal injection of 120 mg/kg pentobarbital sodium, and blood was collected via cardiac puncture for the measurement of serum androgen concentrations.
Experimental Design Study 1: Muscle, Bone, Adipose, and Prostate Tissue Responses to Trenbolone Administration in Gonadectomized Male Rats
Male Fisher 344 rats, aged 3 mo, were divided into six groups (n = 10/group), including sham surgery plus vehicle (SHAM), orchiectomy (ORX) plus vehicle, ORX plus low-dose TREN (ORX + low TREN; 1.0 mg/wk), ORX plus moderate-dose TREN (ORX + mod TREN; 3.5 mg/wk), ORX plus high-dose TREN (ORX + high TREN; 7.0 mg/wk), or ORX plus supraphysiological testosterone-enanthate (ORX + TE; 7.0 mg/wk). We have reported previously that ORX induces catabolic effects in muscle, bone, and kidney and increases visceral fat accumulation (9), whereas supraphysiological TE administration prevents those effects but results in significant prostate enlargement (10, 11, 66). Blood was sampled via tail tip amputation prior to surgery and weekly thereafter. Food consumption was measured weekly. On day 29, rats were euthanized via intraperitoneal pentobarbital sodium (120 mg/kg) injection, and blood was collected via cardiac puncture for Hb analysis and hormone analyses. The levator ani/bulbocavernosus (LABC) muscle complex (an androgen-responsive skeletal muscle complex in rodents), soleus, plantaris, semimembranosus, retroperitoneal fat pads, left and right femurs and tibiae, kidney, and prostate were excised and weighed. With the exception of the femora, all tissues were snap-frozen in liquid nitrogen and stored at −80°C until further analysis. After weighing, the femora were measured using a digital caliper (Mitutoyo, Aurora, IL), wrapped in saline-soaked gauze, and stored at −20°C to maintain the mechanical properties of the bone (44).
Surgery.
Animals received sham surgery or bilateral closed ORX, which involved removal of testes, epididymis, and epididymal fat. All surgeries were performed aseptically under isoflurane anesthesia. After surgery, rats received a nutritional supplement (Jell-O plus protein and fat) daily for 2 days to promote weight maintenance.
Hormone delivery.
TREN and TE (Savient Pharmaceutical, East Brunswick, NJ) were dissolved in sesame oil prior to intramuscular injection. Once weekly injections were alternated between the right and left quadriceps musculature and administered under brief isoflurane anesthesia. Sham surgery animals received vehicle sesame oil injections, where orchiectomized rats received either low (1.0 mg/wk), moderate (3.5 mg/wk), or high doses (7.0 mg/wk) of TREN, TE (7.0 mg/wk), or vehicle immediately following surgery and once every 7 days thereafter.
Hb and serum hormone analyses.
Whole blood Hb was analyzed in duplicate immediately upon euthanization using the Hb Pro Professional Hemoglobin Testing System (ITC, Edison, NJ), which has a sensitivity of 4.0 g/dl and an intra-assay coefficient of variation (CV) of <3.3%. Serum was subsequently separated and stored at −80°C for further analysis. All hormones were assayed in duplicate using commercially available kits. Serum trenbolone was evaluated using qualitative enzyme-linked immunosorbent assay, which has a sensitivity of 0.1 ng/ml and an intra-assay CV of 3.76% (Neogen, Lexington, KY), and a standard curve was developed using trenbolone (Sigma-Aldrich, St. Louis, MO). Testosterone was assayed using a quantitative EIA kit (Diagnostic Systems Laboratories, Webster, TX) with a sensitivity of 0.04 ng/ml and an intra-assay covariance of 5.7%. DHT was assayed using a quantitative EIA kit (Alpco Diagnostics, Salem, NH) with a sensitivity of 6 pg/ml and an intra-assay covariance of 5.4%. Osteocalcin (a marker of bone formation) was determined with a Rat-Mid EIA, which has a sensitivity of 50 ng/ml and an intra-assay CV of <8% (Immunodiagnostic Systems, Fountain Hills, AZ). Bone resorption was measured by evaluating tartrate-resistant acid phosphatase form 5b (TRAP5b; an early marker of bone resorption) (1) with a RatTrap EIA (Immunodiagnostic systems), which has a sensitivity of 0.1 U/l and an intra-assay CV <5.8%, and by evaluating COOH-terminal telopeptide a1 chain of type 1 collagen (a late marker of bone resorption) with a RatLaps EIA (Immunodiagnostic Systems), which has a sensitivity of 2.0 ng/ml and an intra-assay CV <9.2%.
Bone morphometry and mechanical strength.
Prior to peripheral quantitative computerized tomography (pQCT), the femurs were thawed to room temperature and were kept in saline-soaked gauze except during measurement. The left femoral diaphysis and metaphysis were scanned by pQCT with a Stratec XCT Research M Instrument (Norland Medical Systems, Fort Atkinson, WI). Scans were performed at a distance of 5 (metaphysis) and 15 mm (diaphysis) proximal to the distal end of the femur for measurements of cancellous and cortical bone structure, respectively. The structural variables that were measured include total, trabecular, and cortical bone area (mm2), content (mg/mm), and density (mg/cm3).
Subsequent to pQCT, the midshaft of the left femora was subjected to a medial/lateral three-point bending test, using an MTS material testing machine (MTS Systems, Erden Prairie, MN) with methods we have reported previously (66). Before mechanical testing, a preload (10 N) was applied on the medial surface of the femur using a steel cross-bar fixture. The bending load was applied at 1.0 mm/s until failure. From the load-deformation curve, the following parameters were determined for the femoral shaft: breaking load, yield load, stiffness, and displacement. Bone mechanical strength is expressed as both force (measured in N) and stress (measured in Mpa).
Experimental Design Study 2: Prostate and LABC Responses to Trenbolone Administration in Intact Male Rats
Twenty male Fisher 344 rats, aged 3 mo, remained intact and were divided into the following four groups (n = 5/group): vehicle injection (SHAM), supraphysiological testosterone-enanthate (TE; 7.0 mg/wk), low-dose TREN (low TREN; 1.0 mg/wk), and one-half the low-dose TREN (half-Low TREN; 0.5 mg/wk). Animals remained intact to determine the effects of TREN on prostate and LABC masses and Hb concentrations when administered in the presence of endogenous sex hormones, a situation that simulates humans undergoing androgen replacement therapy. Once weekly intramuscular injections were delivered in a manner identical to that described in experiment 1. On day 29, rats were euthanized via intraperitoneal pentobarbital (120 mg/kg) injection, and blood was collected via cardiac puncture. The LABC muscle complex and prostate were excised and weighed. Whole blood Hb and serum testosterone, DHT, and trenbolone were analyzed using the methods and commercially available kits discussed previously (see experiment 1). For this experiment, the low TREN was a dose identical to that administered in experiment 1, a dose that produced robust muscle growth and preservation of BMD in ORX rats while maintaining prostate mass at the level of SHAMs. Additionally, TREN was administered in a dose lower than what was administered in experiment 1 (i.e., half-low TREN) to further evaluate the dose-response effects of TREN within skeletal muscle and prostate.
Statistical Analysis
Results are reported as means ± SE, and P < 0.05 was defined as the threshold of significance. One-Way ANOVAs (for normally distributed data) were used to separately analyze dependent variables, and Tukey's post hoc test was performed for multiple comparisons among groups when appropriate. The Kruskal-Wallis and Mann-Whitney tests were performed when data were not normally distributed. Hormone values that were below the lowest detectable standard are reported as such and were assigned a value equal to the sensitivity of each individual assay for the purposes of statistical analysis. Data were analyzed with the SPSS version 15.0.0 statistical software package.
RESULTS
Pilot Experiment: Verification of Hormone Delivery and Suppression of Endogenous Sex Hormones
Intramuscular TREN administration elevated serum trenbolone concentrations throughout the 7 days following injection (Fig. 1). On day 7, rats received an additional TREN injection, which resulted in peak trenbolone concentrations (46.0 ± 5.6 ng/ml) occurring on day 9. Thus, once weekly TREN injections resulted in a sustained elevation of serum trenbolone, with peak concentrations occurring within 48 h. Within 10 days of administration, TREN also suppressed serum testosterone of intact animals by 80% (2.55 ± 0.46 vs. 0.49 ± 0.07, P ≤ 0.05) and serum DHT by 70% (644 ± 99 vs. 187 ± 7, P ≤ 0.05) compared with intact vehicle-treated animals.
Fig. 1.

Weekly intramuscular injections of trenbolone-enanthate (TREN) elevate serum trenbolone in a sustainable manner. Intact male F344 rats were injected with 7.0 mg of TREN or vehicle under isoflurane anesthesia at baseline and 7 days later (as indicated by arrow). Blood samples were obtained by tail tip prior to injection and every 2 days thereafter for 10 days. Values are means ± SE; n = 5. Some error bars are within the width of the symbol.
Experiment 1: Study on Muscle, Bone, Adipose, and Prostate Tissue Responses to Trenbolone Administration in Gonadectomized Male Rats
Serum androgen concentrations.
Increasing doses of TREN resulted in a progressive increase in serum trenbolone (P ≤ 0.001; Fig. 2A). Serum testosterone was 1.9 ± 0.1 ng/ml in SHAMs (Fig. 2B) and was elevated 18-fold following TE administration (34.7 ± 2.5, P ≤ 0.01). Testosterone was low to undetectable following ORX (0.04 ng/ml) and in all TREN treatment groups (ORX + low TREN: 0.04 ng/ml; ORX + mod TREN: 0.07 ± 0.03 ng/ml; ORX + high TREN: 0.06 ± 0.02 ng/ml). Serum DHT was 284 ± 33 pg/ml in SHAMs (Fig. 2C) and was elevated 34-fold by TE administration (9,774 ± 522 pg/ml, P ≤ 0.001). DHT was reduced 70% with ORX compared with SHAMs (86 ± 7 pg/ml, P ≤ 0.001) and remained reduced following all TREN treatments (ORX + low TREN: 93 ± 5 pg/ml; ORX + mod TREN: 150 ± 5 pg/ml; ORX + high TREN: 182 ± 6 pg/ml).
Fig. 2.
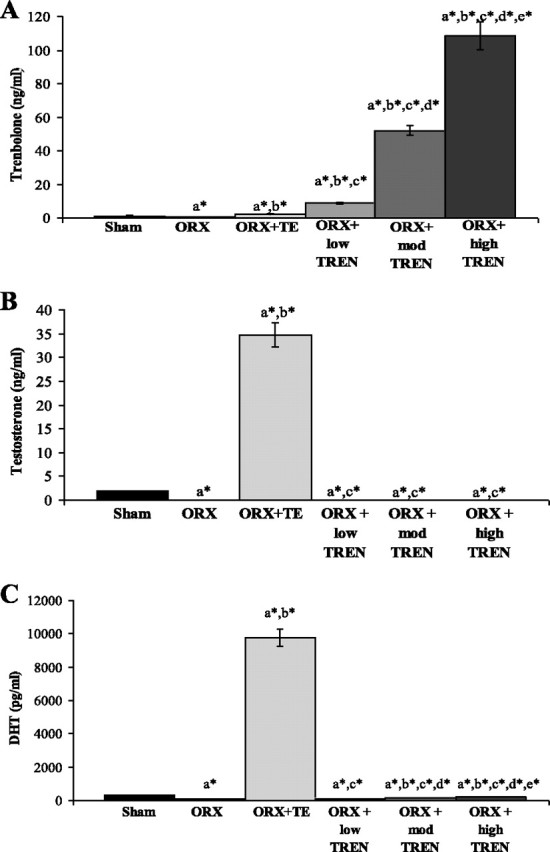
Effects of orchiectomy (ORX), testosterone-enanthate (TE), or graded doses of TREN on serum trenbolone (A), testosterone (B), and dihydrotestosterone (DHT; C) at euthanization. Values are means ± SE; n = 9–10/group for trenbolone and DHT and 6/group for testosterone. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 [avs. vehicle (SHAM); bvs. ORX; cvs. ORX + TE; dvs. ORX + low-dose TREN (low TREN); evs. ORX + moderate-dose TREN (mod TREN)]. High TREN, high-dose TREN.
Weight gain, food consumption, and adiposity.
During the first 3 days following surgery, ORX animals experienced a 75% reduction in weight gain compared with SHAMs, and weight gain remained lower than SHAMs throughout the duration of the study (P ≤ 0.01; Fig. 3A). Both TE and TREN (at all doses) restored weight gain during the first 3 days following ORX. However, after the initial 3-day post-surgery period, total weight gain was gradually reduced in the TE- and TREN-treated groups such that at euthanization the weight gain in all androgen-treated groups was 30–50% lower than in SHAMs (P ≤ 0.001) and not different from ORX (SHAM: 41.6 ± 2.5 g; ORX: 27.6 ± 1.9 g; ORX + TE: 25.7 ± 2.8 g; ORX + low TREN: 29.7 ± 1.5 g; ORX + mod TREN: 25.1 ± 1.1 g; ORX + high TREN: 20.7 ± 1.8 g). No differences in total weight gain were present between the TE and low and mod TREN treatments, although weight gain was 30% lower following high TREN treatment compared with low TREN (P ≤ 0.05).
Fig. 3.

Effects of ORX, TE, or graded doses of TREN on weekly body weight (BW) change (A) and total BW change (B) corrected for food consumption. Rats received ORX vs. sham surgery and were treated with intramuscular injections of vehicle, TE (7.0 mg/wk), or TREN (1.0, 3.5, or 7.0 mg/wk) for a period of 29 days. BW gain was reduced by ORX on day 3, and this reduction was initially prevented by all drug treatment groups. At euthanization, the BW change corrected for food intake was similarly reduced in all ORX groups compared with SHAM. Values are means ± SE; n = 9–10/group. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. ORX; cvs.ORX + TE; dvs. ORX + low TREN; evs. ORX + mod TREN).
ORX also reduced total food consumption throughout the experiment by 14% when compared with SHAMs (P ≤ 0.01), and this reduction was not prevented by any androgen treatment (data not shown). Change in body mass was subsequently corrected for total food consumption to determine whether food consumption influenced the observed differences in weight gain. Following this correction, the change in body mass gain corrected for food intake for the ORX and for the androgen treatments remained 20–40% below that of SHAMs (Fig. 3B). Thus, it appeared that reduced food intake only partially accounted for the reductions in body mass gain following ORX.
ORX induced a nonsignificant 18% increase in retroperitoneal (retro) fat mass compared with SHAMs (Fig. 4A). Conversely, TE administration reduced retro fat mass by 27% compared with ORX (P ≤ 0.01). TREN treatment also resulted in a dose-dependent reduction in retro fat mass. Specifically, low TREN restored retro fat mass to the level of SHAMs, moderate TREN reduced retro fat mass by 35% compared with ORX (P ≤ 0.001) and by 23% compared with low TREN (P ≤ 0.05), and high TREN reduced retro fat mass by 43–51% compared with SHAMs, low TREN, and ORX animals (P ≤ 0.001) and also by 34% compared with TE treatment (P ≤ 0.05).
Fig. 4.
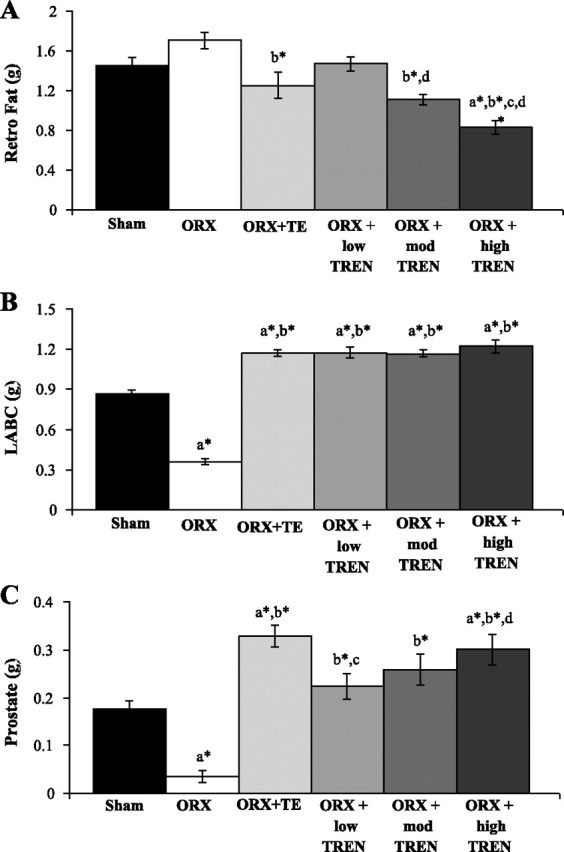
Effects of ORX, TE, or graded doses of TREN on retroperitoneal (retro) fat (A), levator ani/bulbocavernosus (LABC) muscle complex (B), and prostate masses (C). Rats received ORX vs. sham surgery and were treated with intramuscular injections of vehicle, TE (7.0 mg/wk), or TREN (1.0, 3.5, or 7.0 mg/wk) for a period of 29 days. Values are means ± SE; n = 9–10/group. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. ORX; cvs. ORX + TE; dvs. ORX + low TREN; evs. ORX + mod TREN).
Muscle.
ORX resulted in a 59% reduction in the mass of the LABC muscle complex compared with SHAMs (Fig. 4B). All androgen treatments completely prevented this reduction and resulted in an additional 35–40% augmentation of the LABC above SHAMs (P ≤ 0.001). Ultimately, the LABC mass was ∼3.3-fold greater than the ORX group in all androgen treatment groups (P ≤ 0.001). The masses of the plantaris, soleus, and semimembranosus were unaltered by any treatment (data not shown).
Prostate.
ORX resulted in an 80% reduction in prostate mass compared with SHAMs (P ≤ 0.001; Fig. 4C). TE and high TREN administrations increased prostate mass by 84 (P ≤ 0.001) and 68% (P ≤ 0.01), respectively, compared with SHAMs, whereas, neither low nor mod TREN elevated prostate mass above SHAMs. Ultimately, the low TREN resulted in a prostate that was 34% smaller than TE treatment (P ≤ 0.05). No differences in prostate mass were present between the high TREN and TE treatments.
Kidney and Hb.
ORX reduced kidney mass by 10% compared with SHAMs (P ≤ 0.001; Table 1). TE administration prevented this reduction and further increased kidney mass by 14% compared with SHAMs (P ≤ 0.001). Similarly, low TREN completely prevented the ORX-induced loss of kidney mass (P ≤ 0.001), whereas both mod and high TREN increased kidney mass by ∼19% compared with SHAMs (P ≤ 0.001).
Table 1.
Effects of ORX, TE, or graded doses of TREN on kidney mass and Hb concentrations
| SHAM | ORX | ORX + TE | ORX + Low TREN | ORX + Mod TREN | ORX + High TREN | |
|---|---|---|---|---|---|---|
| Kidney, g | 0.89 ± 0.01 | 0.79 ± 0.01a* | 1.00 ± 0.01a*,b* | 0.95 ± 0.01b* | 1.05 ± 0.02a*,b*,d* | 1.06 ± 0.02a*,b*,d* |
| Hb, mg/dl | 15.6 ± 0.2 | 15.7 ± 0.2 | 16.6 ± 0.2 | 16.7 ± 0.3 | 17.3 ± 0.4a*,b* | 17.0 ± 0.3a,b |
Values are means ± SE; n = 9–10/group.
ORX, orchiectomy; TE, testosterone-enanthate; TREN, trenbolone-enanthate; SHAM, vehicle group; low, mod, and high TREN, low-, moderate- and high-dose TREN groups, respectively.
Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. ORX; cvs. ORX + TE; dvs. ORX + low TREN).
No differences in whole blood Hb concentrations were observed between the SHAM, ORX, ORX + TE, or ORX + low TREN animals (Table 1). However, both mod and high TREN administration elevated Hb by ∼8–10% compared with SHAMs and ORX (P ≤ 0.05).
BMD and bone mechanical strength.
ORX resulted in a 12–14% reduction in total bone mineral content (BMC) and total BMD and also reduced trabecular (t)BMC and tBMD by ∼50% at the distal femoral metaphysis compared with shams (P ≤ 0.001; Table 2). Conversely, TE administration prevented the ORX-induced reductions at this skeletal site, ultimately resulting in an 11–12% increase in total BMC (P < 0.05) and total BMD (P ≤ 0.001) and a 62% increase in tBMC and tBMD (P ≤ 0.001). Additionally, all TREN treatments ameliorated the ORX-induced reductions at this skeletal site in a dose-dependent manner, with low TREN treatment increasing total BMD by 6% and tBMD by 37% (P < 0.05), mod TREN treatment increasing total BMD by 9% and tBMC and tBMD by 45% (P ≤ 0.01), and high TREN treatment increasing total BMD by 9% and tBMC and tBMD by 55% (P ≤ 0.001) compared with ORX. No differences were present between TE and any TREN treatments for the bone mineral measurements at this skeletal site, although TREN treatment did not completely restore tBMC and tBMD to SHAM values (P < 0.05). The total and trabecular tissue areas remained unaltered at the distal femoral metaphysis, and no differences were present at the femoral diaphysis for any measurement (data not shown). Additionally, the femoral mass and length remained unaltered by all treatments, and no statistically significant differences were present between any bone mechanical measurements at the femoral midshaft (Table 2).
Table 2.
Effects of ORX, TE, or graded doses of TREN on femoral anthropometric measurements, metaphyseal BMC and BMD, and midshaft bone mechanical characteristics
| SHAM | ORX | ORX + TE | ORX + Low TREN | ORX + Mod TREN | ORX + High TREN | |
|---|---|---|---|---|---|---|
| Femoral mass, g | 0.78 ± 0.01 | 0.75 ± 0.01 | 0.78 ± 0.01 | 0.77 ± 0.01 | 0.77 ± 0.01 | 0.77 ± 0.01 |
| Femoral length, mm | 37.1 ± 0.1 | 36.4 ± 0.2 | 36.8 ± 0.2 | 36.9 ± 0.1 | 36.8 ± 0.1 | 36.7 ± 0.2 |
| Total BMC, mg/mm | 12.7 ± 0.3 | 10.9 ± 0.3a* | 12.1 ± 0.4b | 11.5 ± 0.3a | 11.9 ± 0.3 | 11.9 ± 0.2 |
| Total BMD, mg/cm3 | 668 ± 6 | 587 ± 8a* | 657 ± 10b* | 624 ± 7a*,b | 632 ± 8a,b* | 641 ± 9b* |
| Trabecular BMC, mg/mm | 2.0 ± 0.1 | 1.0 ± 0.1a* | 1.6 ± 0.1b* | 1.4 ± 0.1a* | 1.5 ± 0.1a*,b | 1.5 ± 0.1a,b* |
| Trabecular BMD, mg/cm3 | 344 ± 16 | 177 ± 13a* | 288 ± 17b* | 243 ± 15a*,b | 257 ± 14a*,b* | 274 ± 16a,b* |
| Breaking load, N | 136 ± 12 | 134 ± 11 | 138 ± 7 | 139 ± 11 | 139 ± 10 | 137 ± 10 |
| Yield load, N | 93 ± 17 | 90 ± 20 | 93 ± 12 | 90 ± 17 | 116 ± 16 | 98 ± 20 |
| Stiffness, N/mm | 245 ± 32 | 227 ± 30 | 270 ± 25 | 257 ± 30 | 253 ± 26 | 256 ± 34 |
Values are means ± SE; n = 9–10/group.
BMC, bone mineral content; BMD, bone mineral density.
Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. ORX; cvs. ORX + TE; dvs. ORX + low TREN).
Systemic markers of bone remodeling.
One week following surgery, serum Trap5b [an early marker of bone resorption (1)] was 9.4 ± 0.6 (SHAM), 11.7 ± 0.9 (ORX), 10.2 ± 0.4 (ORX + low TREN), 8.2 ± 0.4 (ORX + mod TREN), 7.6 ± 0.4 (ORX + high TREN), and 9.4 ± 0.8 (ORX + TE) (Fig. 5A). ORX resulted in a nonsignificant 25% increase in Trap5b, which was completely prevented by TE. Low TREN also resulted in a nonsignificant 15% reduction in Trap5b compared with ORX, whereas both mod TREN and high TREN reduced Trap5b by 30% compared with ORX (P < 0.01). At euthanization, serum C-telopeptide (a late marker of bone resorption) was 33.3 ± 2.6 (SHAM), 36.3 ± 2.3 (ORX), 30.0 ± 2.2 (ORX + low TREN), 32.0 ± 2.6 (ORX + mod TREN), 26.5 ± 2.7 (ORX + high TREN), and 33.7 ± 2.2 (ORX + TE). High TREN reduced serum C-telopeptide by 27% compared with ORX (P ≤ 0.05), with no other significant differences present between groups (Fig. 5B). At euthanization, serum osteocalcin (a marker of bone formation) was 679 ± 56 (SHAM), 892 ± 95 (ORX), 435 ± 42 (ORX + low TREN), 320 ± 30 (ORX + mod TREN), 282 ± 26 (ORX + high TREN), and 380 ± 48 (ORX + TE) (Fig. 5C). ORX resulted in a nonsignificant 31% increase in serum osteocalcin compared with SHAMs. All androgen treatment groups reduced serum osteocalcin by 36–58% compared with SHAMs (P ≤ 0.01) and by 51–68% compared with ORX (P ≤ 0.001). No differences were present between TREN- and TE-treated groups. In addition, serum osteocalcin was 26% lower in mod TREN (P = 0.063, trend) and 35% lower in high TREN (P ≤ 0.05) groups compared with the low TREN.
Fig. 5.
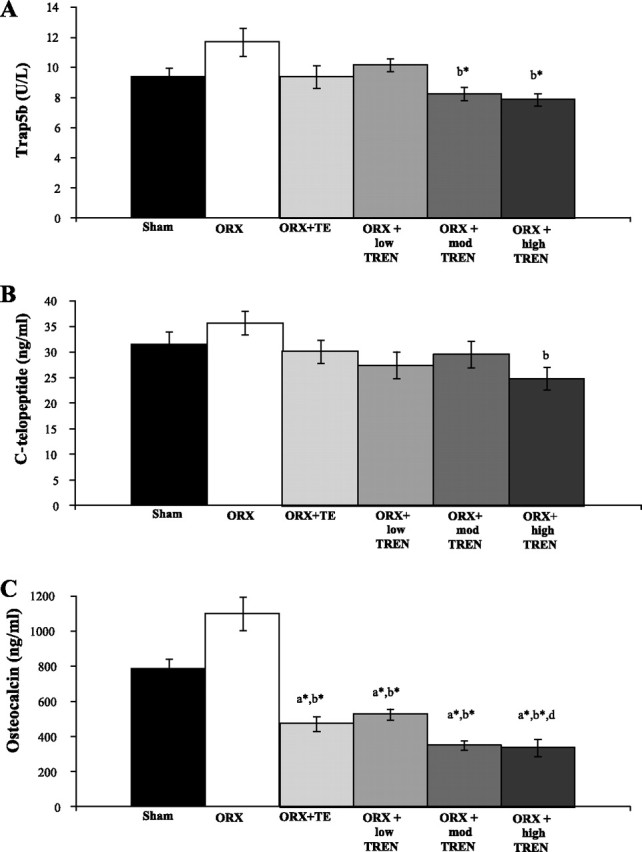
Effects of ORX, TE, or graded doses of TREN on serum tartrate-resistant acid phosphatase form 5b (Trap5b, an early marker of bone resorption; A), C-telopeptide (a later marker of bone resorption; B), and osteocalcin (C) at euthanization. Rats received ORX vs. sham surgery and were treated with intramuscular injections of vehicle, TE (7.0 mg/wk), or TREN (1.0, 3.5, or 7.0 mg/wk) for a period of 29 days. Values are means ± SE; n = 9–10/group. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. ORX; cvs.ORX + TE; dvs. ORX + low TREN; evs. ORX + mod TREN).
Experiment 2: Study on Prostate and LABC Responses to Trenbolone Administration in Intact Male Rats
Hormone responses.
At euthanization, serum trenbolone was 13.7 ± 0.6 ng/ml in intact low TREN-treated animals and 4.8 ± 0.3 ng/ml in half-low TREN-treated animals (P < 0.01; Fig. 6A). Serum testosterone was 5.7 ± 1.5 ng/ml in SHAM animals and was increased ninefold by TE administration (53.0 ± 4.5 ng/ml, P < 0.01; Fig. 6B). Serum testosterone was reduced to nearly undetectable concentrations by both low TREN (0.4 ± 0.1 ng/ml) and half-low TREN (0.3 ± 0.1 ng/ml) treatments (P < 0.01). Serum DHT was 962 ± 198 pg/ml in SHAM animals and was increased ∼13-fold in TE-treated animals (12,288 ± 1,037 pg/ml, P < 0.01), whereas DHT was reduced below SHAM values by low TREN treatment (126 ± 4 pg/ml, P < 0.01) and below both sham and low TREN treatment values by the half-Low TREN treatment (88 ± 2 pg/ml, P < 0.01) (Fig. 6C).
Fig. 6.
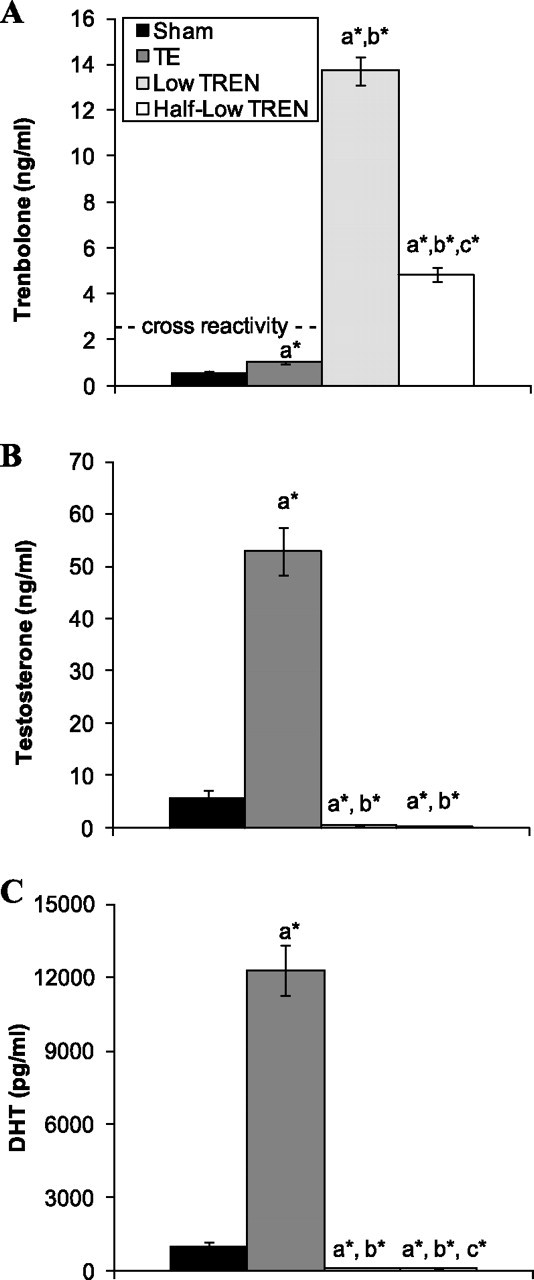
Effects of supraphysiological TE and graded doses of TREN on serum trenbolone (A), testosterone (B), and DHT (C) at euthanization. Intact male rats received intramuscular injections of vehicle, TE (7.0 mg/wk), or TREN (0.5 or 1.0 mg/wk) for a period of 29 days. Values are means ± SE; n = 5/group. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. TE; cvs. low TREN).
Weight gain.
At baseline, body mass was similar among all groups (SHAM: 270.7 ± 2.6 g; TE: 270.7 ± 3.7 g; low TREN: 271.6 ± 2.2 g; half-Low TREN: 271.4 ± 4.9 g). Body mass gain was reduced to a similar degree by all androgen treatments, which became significant on day 21 of the experiment (P < 0.05; Fig. 7A). Combined retroperitoneal and perirenal (visceral) fat mass was also reduced 25% by TE treatment (P < 0.05) and was nonsignificantly reduced 13–16% by both the low and half-low TREN treatments (Fig. 7B).
Fig. 7.
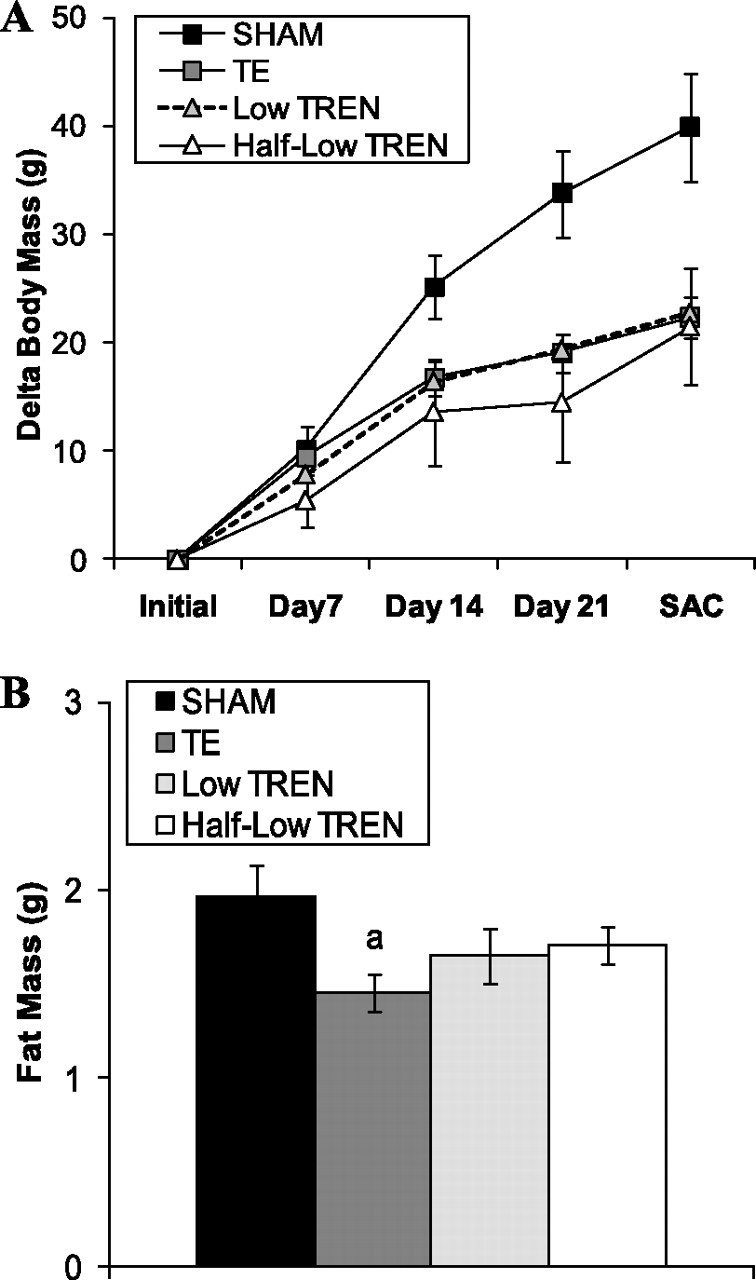
Effects of supraphysiological TE and graded doses of TREN on body mass change (A) and visceral fat pad mass (B). Intact male rats received intramuscular injections of vehicle, TE (7.0 mg/wk), or TREN (0.5 or 1.0 mg/wk) for a period of 29 days. Values are means ± SE; n = 5/group. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. TE; cvs. low TREN).
Prostate and LABC.
In intact animals, TE treatment increased prostate mass by 59% compared with SHAMs (P < 0.001; Fig. 8A). Low TREN maintained prostate mass at the level of SHAMs, ultimately resulting in a prostate that was 31% smaller than following TE treatment (P < 0.001). Half-low TREN treatment resulted in a nonsignificant 22% reduction in prostate mass compared with SHAMs, a 29% reduction in prostate mass compared with low TREN treatment (P < 0.01), and a 51% reduction compared with TE treatment (P < 0.001).
Fig. 8.
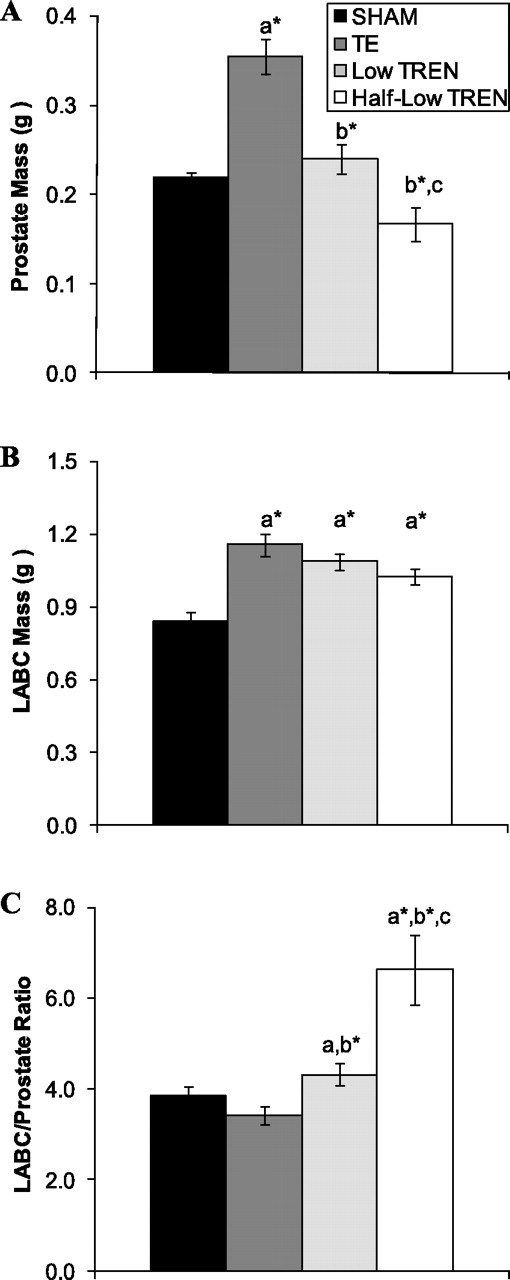
Effects of supraphysiological TE and graded doses of TREN on prostate mass (A), LABC muscle complex mass (B), and LABC/prostate ratio (C). Intact male rats received intramuscular injections of vehicle, TE (7.0 mg/wk), or TREN (0.5 or 1.0 mg/wk) for a period of 29 days. Values are means ± SE; n = 5/group. Letters indicate differences from respectively labeled groups at P < 0.05 or *P < 0.01 (avs. SHAM; bvs. TE; cvs. low TREN).
Mass of the LABC muscle complex was increased by 21–38% in TE- and TREN-treated animals (P < 0.01; Fig. 8B), with no differences present between any androgen treatment. The LABC/prostate ratio, which illustrates the anabolic/androgenic ratio of androgens, was unaltered by TE treatment (Fig. 8C). Following Low TREN treatment, the LABC/prostate ratio was increased by 12% compared with SHAMs (P < 0.05) and by 25% compared with TE treatment (P < 0.01). Ultimately, half-low TREN treatment resulted in the highest LABC/prostate ratio, a value that was 53% higher than low TREN treatment (P < 0.05), 72% higher than SHAMs (P < 0.01), and 92% higher than TE treatment (P < 0.01).
Kidney and Hb.
No differences in kidney mass (sham: 0.93 ± 0.02 g; TE: 1.01 ± 0.01 g; low TREN: 0.98 ± 0.02 g; half-low TREN: 0.94 ± 0.04 g) or Hb concentrations (sham: 14.6 ± 0.2 mg/dl; TE: 15.7 ± 0.2 mg/dl; low TREN: 15.5 ± 0.4 mg/dl; half-low TREN: 15.4 ± 0.4 mg/dl) were observed between groups.
DISCUSSION
Our results demonstrate that TREN prevents the deleterious alterations in body composition associated with ORX to the same extent as supraphysiological TE. Specifically, we observed that 1) regardless of dose, TREN and supraphysiological TE produced equally myotrophic responses in the androgen-sensitive LABC muscle complex in both intact and ORX animals, 2) TREN partially prevented ORX-induced bone loss to roughly the same extent as supraphysiological TE, and 3) at equal doses, TREN was somewhat more lipolytic than TE in visceral fat. In contrast, the lowest doses of TREN maintained prostate mass and Hb concentrations at the level of shams in both intact and orchiectomized animals, whereas supraphysiological TE and high-dose TREN produced prostate enlargement and/or elevations in Hb. Thus, at the lowest doses administered, TREN appears to have a higher and more clinically favorable anabolic/androgenic ratio than supraphysiological TE.
Skeletal muscle expresses ARs to varying degrees among specific muscle groups and among species. For example, humans respond robustly to androgenic stimuli due to the high percentages of AR-positive myonuclei (54). Conversely, certain rodent peripheral muscles respond poorly to androgenic stimuli due to the their low concentrations of AR-positive myonuclei (e.g., extensor digitorum longus with 7% AR-positive myonuclei) (34), whereas the rodent LABC muscle complex contains ∼74% AR-positive myonuclei and experiences a substantial atrophic response to androgen ablation and a robust myotrophic response to androgen administration (34). In our study, the lower limb muscles were insensitive to androgen status; however, ORX reduced LABC muscle mass, whereas TREN and TE robustly augmented LABC muscle mass. These results demonstrate clearly that TREN is at least as myotrophic as supraphysiological TE within androgen-sensitive skeletal muscle because even the lowest TREN doses produced equal growth of the LABC muscle in intact and ORX animals compared with supraphysiological TE. The mechanism(s) underlying the TREN-induced augmentation of skeletal muscle mass requires further clarification (67), although it is suspected that TREN acts via 1) AR activation, since trenbolone possesses three times the affinity of testosterone for the AR (5), 2) upregulation of endogenous growth factors (e.g., IGF-I or fibroblast growth factor) and/or increased responsiveness of skeletal muscle to such growth factors (29, 60), and/or 3) anticatabolic mechanisms associated with reductions in endogenous glucocorticoid activity (48, 49).
In our study, androgen treatment [either TE or TREN (at all doses)] prevented the initial weight reduction associated with surgical ORX to a similar extent. However, at euthanization, body weight gain was similarly reduced among all ORX groups compared with SHAMs, regardless of androgen treatment. Similarly, all intact androgen-treated animals experienced a reduced body weight gain compared with SHAMs. It remains unclear why TE did not maintain body weight gain throughout these studies, as we have reported previously (66). The reduced weight accrual may be at least partially explained by the lower food consumption within all ORX groups throughout the duration of experiment 1. However, the reduced food consumption did not cause a measurable loss of lower limb skeletal muscle mass. Conversely, we observed that ORX increased retroperitoneal fat mass, albeit nonsignificantly, and that visceral fat mass was reduced by TREN in a dose-dependent manner. Furthermore, at equal doses, TREN induced a greater magnitude of fat loss than TE. These results are somewhat surprising considering that all androgen treatments produced similar myotrophic growth of the androgen-sensitive LABC muscle complex and induced a substantial and roughly equivalent prevention of the ORX-induced BMD loss. Thus, it appears that TREN is capable of preventing the initial catabolic responses associated with ORX while inducing more potent lipolytic effects within visceral adiposity than endogenous androgens, at least on a dose-to-dose equivalent. Future research examining the mechanisms underlying the lipolytic effects of TREN is appealing considering the preponderance of health decrements associated with central adiposity (15).
Clearly, androgens influence the development of male secondary sexual characteristics, skeletal muscle, bone, and fat, among other tissues (59). Testosterone is the primary circulating endogenous androgen that produces biological responses following nuclear interactions with ARs or through nongenomic signaling pathways (19). However, in tissues that highly express 5α-reductase (e.g., prostate), testosterone is transformed to its more potent metabolite DHT, which increases its androgenic potency within these tissues (63). As such, prostate enlargement is a concern associated with testosterone replacement therapy in hypogonadal men (13). In our studies, the lowest doses of TREN did not induce prostate enlargement in ORX or intact animals, whereas supraphysiological TE and the highest TREN dose increased prostate mass to a similar magnitude. These results are interesting considering that TREN binds to ARs with approximately three times the affinity of TE and an affinity roughly equal to that of DHT, the most potent endogenous androgen. In contrast to testosterone, certain anabolic steroids (e.g., 19-nortestosterone) are transformed to less potent metabolites by 5α-reductase or are not substrates for 5α-reductase (e.g., 7α-methyl-19-nortestosterone) due to slight variations in the structures of these compounds (58). Similarly, the reduced prostate enlargement associated with TREN appears to result from the presence of a 3-oxotriene structure that prevents A-ring reduction of this steroid (46), ultimately limiting its ability to undergo 5α-reduction or aromatization. As a result, trenbolone is primarily metabolized to its less potent 17α-isoform (epitrenbolone) or trendione in humans (55) and in other mammalian species (17, 31, 45, 46), underlying its equal potency in tissues that highly express 5α-reductase (e.g., prostate) vs. those that do not (e.g., skeletal muscle) (67).
Another concern with androgen replacement therapy is the potential to initiate or accelerate the growth of undiagnosed prostate cancer, although several recent meta-analyses have failed to detect an increased risk of prostate cancer following testosterone replacement in hypogonadal men (13, 18). In our studies, TREN administration resulted in a nearly complete ablation of circulating testosterone and DHT, the primary endogenous androgens implicated in prostate growth, through mechanisms apparently associated with feedback inhibition; however, we did not directly evaluate the possibility that TREN may initiate or accelerate prostate cancer growth. Several previous reports indicate that trenbolone appears to produce little genotoxic activity and is not an initiator of cancer in various in vitro models (47, 51), although trenbolone has been shown capable of inducing morphological changes in the Syrian hamster embryo cell transformation assay (an in vitro mutagenicity model) (50, 51). As such, future research examining the effects of TREN and other androgens using in vitro and/or in vivo xenograft models of androgen-responsive prostate cancer is warranted and should be conducted prior to this agent being advanced to clinical testing.
Certain androgens also undergo in vivo aromatization (e.g., the conversion of testosterone to estradiol) and subsequently exert systemic or tissue-specific estrogen receptor-mediated effects (28). The aromatization of testosterone appears to be essential for bone development in men (62), as demonstrated by the identification of several men who suffer from congenital aromatase deficiency resulting in osteopenia, which is treatable with estradiol but not testosterone (28). Similarly, male aromatase knockout mice develop severe osteopenia and adverse skeletal development, which is reversible with estradiol administration (37, 41, 43). Conversely, a number of side effects commonly associated with supraphysiological androgen administration are influenced by excessive systemic or tissue-specific aromatization of androgens, including gyncecomastia (12), prostate cancer (14), fluid retention (56), and premature epiphyseal closure (42). We were unable to measure circulating estrogens in our study, since the currently available methods lack the sensitivity required for this analysis in orchiectomized male rodents. However, in vitro evidence indicates that trenbolone and its primary metabolites are relatively nonestrogenic (30), and in vivo evidence demonstrates that trenbolone induces antiestrogenic effects in oviparous species (3, 16, 23, 32, 38, 53). Additionally, it has been reported that trenbolone does not undergo aromatization due to its 3-oxotriene structure (46), although conflicting reports exist regarding the influence of trenbolone on circulating estrogens in mammalian species (24, 26). It is plausible that the antiestrogenic effects and apparently nonaromatizable aspects of trenbolone partially underlie the slightly diminished bone-protective effects and the apparent adynamic bone state (i.e., reduced markers of bone resorption and bone formation) that we observed following TREN treatment of ORX animals. These results may provide further evidence indicating that the bone-protective effects of androgens are at least partially mediated by the systemic and/or tissue-specific aromatization to estrogens. Clearly, future research examining bone histomorphometry would provide further insight into the effects of TREN on bone turnover. Regardless, our findings indicate that androgens are capable of exerting direct bone protection, as evidenced by the TREN-induced partial prevention of BMD loss and the reversal of ORX-induced increases in serum markers of bone resorption and formation, results that ultimately assisted in the maintenance of bone strength. As such, future research evaluating the mechanism(s) underlying the bone-protective effects of TREN and other nonaromatizable androgens are warranted, especially considering the influence of estrogens on bone health and on the side effects associated with excessive androgen administration.
Recently, SARMs that produce anabolic responses within muscle and bone without inducing prostate enlargement or other androgenic side effects have been developed (33). The mechanism(s) through which steroidal and nonsteroidal SARMs produce selective tissue-specific anabolic responses has not been fully elucidated, although it has been postulated that their reduced activity within the prostate and other androgenic tissues may result from their inability to undergo tissue-specific 5α-reduction to more potent metabolites (22). Our results demonstrate that TREN, a non-5α-reducible testosterone analog, exhibits SARM-like properties since it prevented the ORX-induced reductions in muscle and bone and also reduced fat mass while maintaining prostate mass and Hb at the level of intact animals. In fact, the half-low TREN treatment reduced prostate mass in intact animals, albeit nonsignificantly, whereas it increased LABC muscle mass, which corroborates previous work indicating that TREN remains myotrophic when administered at doses lower than in our current study (4, 20, 21, 35, 36, 40), and may also be capable of reducing prostate mass in developing animals (25, 64). The likely mechanisms underlying these findings are that 1) TREN ablates circulating testosterone and DHT in intact animals and 2) TREN undergoes tissue-specific biotransformation to less potent androgens in vivo. The long-term SARM-like potential of TREN remains to be determined, as does its anabolic/androgenic effects within older hypogonadal rodents, a model that may more appropriately mimic the effects of aging and/or hypogonadism in men.
In conclusion, administration of TREN, a potent non-5α-reducible and nonestrogenic synthetic testosterone analog, produces robust myotrophic effects, partial inhibition of bone loss, prevention of visceral fat accumulation, and maintenance of Hb and prostate mass at the level of intact animals, at least at the lowest dose administered. These results suggest that lower-dose TREN induces favorable SARM-like effects on musculoskeletal tissue and adiposity and within specific accessory sex organs. Future research examining the safety and efficacy of this androgen in preclinical settings appears to be appropriate because the anabolic/androgenic ratio appears to be higher and because the risk/benefit ratio appears to be less than that of supraphysiological TE, at least in regard to prostate enlargement, although evaluating TREN and other SARMs in models of androgen-responsive prostate cancer is necessary prior to these agents being recommended for clinical testing.
GRANTS
This work was supported by a Veterans Department Merit Award to S. E. Borst.
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
REFERENCES
- 1.Alatalo SL, Peng Z, Janckila AJ, Kaija H, Vihko P, Vaananen HK, Halleen JM. A novel immunoassay for the determination of tartrate-resistant acid phosphatase 5b from rat serum. J Bone Miner Res : 134–139, 2003. [DOI] [PubMed] [Google Scholar]
- 2.Amory JK, Watts NB, Easley KA, Sutton PR, Anawalt BD, Matsumoto AM, Bremner WJ, Tenover JL. Exogenous testosterone or testosterone with finasteride increases bone mineral density in older men with low serum testosterone. J Clin Endocrinol Metab : 503–510, 2004. [DOI] [PubMed] [Google Scholar]
- 3.Ankley GT, Defoe DL, Kahl MD, Jensen KM, Makynen EA, Miracle A, Hartig P, Gray LE, Cardon M, Wilson V. Evaluation of the model anti-androgen flutamide for assessing the mechanistic basis of responses to an androgen in the fathead minnow (Pimephales promelas). Environ Sci Technol : 6322–6327, 2004. [DOI] [PubMed] [Google Scholar]
- 4.Ashby J, Lefevre PA, Tinwell H, Odum J, Owens W. Testosterone-stimulated weanlings as an alternative to castrated male rats in the Hershberger anti-androgen assay. Regul Toxicol Pharmacol : 229–238, 2004. [DOI] [PubMed] [Google Scholar]
- 5.Bauer ER, Daxenberger A, Petri T, Sauerwein H, Meyer HH. Characterisation of the affinity of different anabolics and synthetic hormones to the human androgen receptor, human sex hormone binding globulin and to the bovine progestin receptor. Apmis : 838–846, 2000. [DOI] [PubMed] [Google Scholar]
- 6.Bhasin S, Calof OM, Storer TW, Lee ML, Mazer NA, Jasuja R, Montori VM, Gao W, Dalton JT. Drug insight: Testosterone and selective androgen receptor modulators as anabolic therapies for chronic illness and aging. Nat Clin Pract Endocrinol Metab : 146–159, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bhasin S, Woodhouse L, Casaburi R, Singh AB, Bhasin D, Berman N, Chen X, Yarasheski KE, Magliano L, Dzekov C, Dzekov J, Bross R, Phillips J, Sinha-Hikim I, Shen R, Storer TW. Testosterone dose-response relationships in healthy young men. Am J Physiol Endocrinol Metab : E1172–E1181, 2001. [DOI] [PubMed] [Google Scholar]
- 8.Bhasin S, Woodhouse L, Casaburi R, Singh AB, Mac RP, Lee M, Yarasheski KE, Sinha-Hikim I, Dzekov C, Dzekov J, Magliano L, Storer TW. Older men are as responsive as young men to the anabolic effects of graded doses of testosterone on the skeletal muscle. J Clin Endocrinol Metab : 678–688, 2005. [DOI] [PubMed] [Google Scholar]
- 9.Borst SE, Conover CF. Orchiectomized Fischer 344 male rat models body composition in hypogonadal state. Life Sci : 411–415, 2006. [DOI] [PubMed] [Google Scholar]
- 10.Borst SE, Conover CF, Carter CS, Gregory CM, Marzetti E, Leeuwenburgh C, Vandenborne K, Wronski TJ. Anabolic effects of testosterone are preserved during inhibition of 5α-reductase. Am J Physiol Endocrinol Metab : E507–E514, 2007. [DOI] [PubMed] [Google Scholar]
- 11.Borst SE, Lee JH, Conover CF. Inhibition of 5α-reductase blocks prostate effects of testosterone without blocking anabolic effects. Am J Physiol Endocrinol Metab : E222–E227, 2005. [DOI] [PubMed] [Google Scholar]
- 12.Braunstein GD. Aromatase and gynecomastia. Endocr Relat Cancer : 315–324, 1999. [DOI] [PubMed] [Google Scholar]
- 13.Calof OM, Singh AB, Lee ML, Kenny AM, Urban RJ, Tenover JL, Bhasin S. Adverse events associated with testosterone replacement in middle-aged and older men: a meta-analysis of randomized, placebo-controlled trials. J Gerontol A Biol Sci Med Sci : 1451–1457, 2005. [DOI] [PubMed] [Google Scholar]
- 14.Carruba G. Estrogen and prostate cancer: an eclipsed truth in an androgen-dominated scenario. J Cell Biochem : 899–911, 2007. [DOI] [PubMed] [Google Scholar]
- 15.Cornier MA, Dabelea D, Hernandez TL, Lindstrom RC, Steig AJ, Stob NR, Van Pelt RE, Wang H, Eckel RH. The metabolic syndrome. Endocr Rev : 777–822, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dorts J, Richter CA, Wright-Osment MK, Ellersieck MR, Carter BJ, Tillitt DE. The genomic transcriptional response of female fathead minnows (Pimephales promelas) to an acute exposure to the androgen, 17beta-trenbolone. Aquat Toxicol : 44–53, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Evrard P, Maghuin-Rogister G, Rico AG. Fate and residues of trenbolone acetate in edible tissues from sheep amd calves implanted with tritium-labeled trenbolone acetate. J Anim Sci : 1489–1496, 1989. [DOI] [PubMed] [Google Scholar]
- 18.Fernández-Balsells MM, Murad MH, Lane M, Lampropulos JF, Albuquerque F, Mullan RJ, Agrwal N, Elamin MB, Gallegos-Orozco JF, Wang AT, Erwin PJ, Bhasin S, Montori VM. Clinical review 1: Adverse effects of testosterone therapy in adult men: a systematic review and meta-analysis. J Clin Endocrinol Metab : 2560–2575, 2010. [DOI] [PubMed] [Google Scholar]
- 19.Foradori CD, Weiser MJ, Handa RJ. Non-genomic actions of androgens. Front Neuroendocrinol : 169–181, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Freyberger A, Ellinger-Ziegelbauer H, Krotlinger F. Evaluation of the rodent Hershberger bioassay: testing of coded chemicals and supplementary molecular-biological and biochemical investigations. Toxicology : 77–88, 2007. [DOI] [PubMed] [Google Scholar]
- 21.Freyberger A, Hartmann E, Krötlinger F. Evaluation of the rodent Hershberger bioassay using three reference (anti)androgens. Arh Hig Rada Toksikol : 131–139, 2005. [PubMed] [Google Scholar]
- 22.Gao W, Dalton JT. Ockham's razor and selective androgen receptor modulators (SARMs): are we overlooking the role of 5alpha-reductase? Mol Interv : 10–13, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hemmer MJ, Cripe GM, Hemmer BL, Goodman LR, Salinas KA, Fournie JW, Walker CC. Comparison of estrogen-responsive plasma protein biomarkers and reproductive endpoints in sheepshead minnows exposed to 17beta-trenbolone. Aquat Toxicol : 128–136, 2008. [DOI] [PubMed] [Google Scholar]
- 24.Henricks DM, Edwards RL, Champe KA, Gettys TW, Skelley GC, Gimenez T. Trenbolone, estradiol-17 beta and estrone levels in plasma and tissues and live weight gains of heifers implanted with trenbolone acetate. J Anim Sci : 1048–1056, 1982. [DOI] [PubMed] [Google Scholar]
- 25.Hotchkiss AK, Nelson RJ. An environmental androgen, 17beta-trenbolone, affects delayed-type hypersensitivity and reproductive tissues in male mice. J Toxicol Environ Health A : 138–140, 2007. [DOI] [PubMed] [Google Scholar]
- 26.Hunt DW, Henricks DM, Skelley GC, Grimes LW. Use of trenbolone acetate and estradiol in intact and castrate male cattle: effects on growth, serum hormones, and carcass characteristics. J Anim Sci : 2452–2462, 1991. [DOI] [PubMed] [Google Scholar]
- 27.Isidori AM, Giannetta E, Greco EA, Gianfrilli D, Bonifacio V, Isidori A, Lenzi A, Fabbri A. Effects of testosterone on body composition, bone metabolism and serum lipid profile in middle-aged men: a meta-analysis. Clin Endocrinol : 280–293, 2005. [DOI] [PubMed] [Google Scholar]
- 28.Jones ME, Boon WC, Proietto J, Simpson ER. Of mice and men: the evolving phenotype of aromatase deficiency. Trends Endocrinol Metab : 55–64, 2006. [DOI] [PubMed] [Google Scholar]
- 29.Kamanga-Sollo E, White ME, Hathaway MR, Chung KY, Johnson BJ, Dayton WR. Roles of IGF-I and the estrogen, androgen and IGF-I receptors in estradiol-17beta- and trenbolone acetate-stimulated proliferation of cultured bovine satellite cells. Domest Anim Endocrinol : 88–97, 2008. [DOI] [PubMed] [Google Scholar]
- 30.Le Guevel R, Pakdel F. Assessment of oestrogenic potency of chemicals used as growth promoter by in-vitro methods. Hum Reprod : 1030–1036, 2001. [DOI] [PubMed] [Google Scholar]
- 31.Metzler M. Metabolism of some anabolic agents: toxicological and analytical aspects. J Chromatograph : 11–21, 1989. [DOI] [PubMed] [Google Scholar]
- 32.Miracle A, Ankley G, Lattier D. Expression of two vitellogenin genes (vg1 and vg3) in fathead minnow (Pimephales promelas) liver in response to exposure to steroidal estrogens and androgens. Ecotoxicol Environ Saf : 337–342, 2006. [DOI] [PubMed] [Google Scholar]
- 33.Mohler ML, Bohl CE, Jones A, Coss CC, Narayanan R, He Y, Hwang DJ, Dalton JT, Miller DD. Nonsteroidal selective androgen receptor modulators (SARMs): dissociating the anabolic and androgenic activities of the androgen receptor for therapeutic benefit. J Med Chem : 3597–3617, 2009. [DOI] [PubMed] [Google Scholar]
- 34.Monks DA, O'Bryant EL, Jordan CL. Androgen receptor immunoreactivity in skeletal muscle: enrichment at the neuromuscular junction. J Comp Neurol : 59–72, 2004. [DOI] [PubMed] [Google Scholar]
- 35.Moon HJ, Kang TS, Kim TS, Kang IH, Ki HY, Kim SH, Han SY. OECD validation of phase 3 Hershberger assay in Korea using surgically castrated male rats with coded chemicals. J Appl Toxicol : 350–355, 2009. [DOI] [PubMed] [Google Scholar]
- 36.Moon HJ, Kang TS, Kim TS, Kang IH, Kim SH, Han SY. OECD validation of phase-3 Hershberger assay using the stimulated weanling male rat in Korea. J Appl Toxicol 2010. [DOI] [PubMed] [Google Scholar]
- 37.Murata Y, Robertson KM, Jones ME, Simpson ER. Effect of estrogen deficiency in the male: the ArKO mouse model. Mol Cell Endocrinol : 7–12, 2002. [DOI] [PubMed] [Google Scholar]
- 38.Orn S, Yamani S, Norrgren L. Comparison of vitellogenin induction, sex ratio, and gonad morphology between zebrafish and Japanese medaka after exposure to 17alpha-ethinylestradiol and 17beta-trenbolone. Arch Environ Contam Toxicol : 237–243, 2006. [DOI] [PubMed] [Google Scholar]
- 39.Ottenbacher KJ, Ottenbacher ME, Ottenbacher AJ, Acha AA, Ostir GV. Androgen treatment and muscle strength in elderly men: A meta-analysis. J Am Geriatr Soc : 1666–1673, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Owens W, Gray LE, Zeiger E, Walker M, Yamasaki K, Ashby J, Jacob E. The OECD program to validate the rat Hershberger bioassay to screen compounds for in vivo androgen and antiandrogen responses: phase 2 dose-response studies. Environ Health Perspect : 671–678, 2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Oz OK, Hirasawa G, Lawson J, Nanu L, Constantinescu A, Antich PP, Mason RP, Tsyganov E, Parkey RW, Zerwekh JE, Simpson ER. Bone phenotype of the aromatase deficient mouse. J Steroid Biochem Mol Biol : 49–59, 2001. [DOI] [PubMed] [Google Scholar]
- 42.Oz OK, Millsaps R, Welch R, Birch J, Zerwekh JE. Expression of aromatase in the human growth plate. J Mol Endocrinol : 249–253, 2001. [DOI] [PubMed] [Google Scholar]
- 43.Oz OK, Zerwekh JE, Fisher C, Graves K, Nanu L, Millsaps R, Simpson ER. Bone has a sexually dimorphic response to aromatase deficiency. J Bone Miner Res : 507–514, 2000. [DOI] [PubMed] [Google Scholar]
- 44.Pelker RR, Friedlaender GE, Markham TC, Panjabi MM, Moen CJ. Effects of freezing and freeze-drying on the biomechanical properties of rat bone. J Orthop Res : 405–411, 1984. [DOI] [PubMed] [Google Scholar]
- 45.Pottier J, Busigny M, Grandadam JA. Plasma kinetics, excretion in mild and tissue levels in the cow following implantation of trenbolone acetate. J Anim Sci : 962–968, 1975. [DOI] [PubMed] [Google Scholar]
- 46.Pottier J, Cousty C, Heitzman RJ, Reynolds IP. Differences in the biotransformation of a 17 beta-hydroxylated steroid, trenbolone acetate, in rat and cow. Xenobiotica : 489–500, 1981. [DOI] [PubMed] [Google Scholar]
- 47.Richold M. The genotoxicity of trenbolone, a synthetic steroid. Arch Toxicol : 249–258, 1988. [DOI] [PubMed] [Google Scholar]
- 48.Santidrian S, Thompson JR. Effect of sex, testosterone propionate and trienbolone acetate on the rate of growth and myofibrillar protein degradation in growing young rats. Reproduccion : 33–41, 1982. [PubMed] [Google Scholar]
- 49.Santidrián S, Thompson JR, Young VR. Effect of trienbolone acetate on the rate of myofibrillar protein breakdown in young adrenalectomized male rate treated with corticosterone. Arch Farmacol Toxicol : 333–340, 1981. [PubMed] [Google Scholar]
- 50.Schiffmann D, Hieber L, Schmuck G, Pechan R, Metzler M, Henschler D. Trenbolone induces micronucleus formation and neoplastic transformation in Syrian hamster embryo fibroblasts but not in mouse C3H10T1/2 cells. Arch Toxicol : 49–53, 1988. [DOI] [PubMed] [Google Scholar]
- 51.Schiffmann D, Metzler M, Neudecker T, Henschler D. Morphological transformation of Syrian hamster embryo fibroblasts by the anabolic agent trenbolone. Arch Toxicol : 59–63, 1985. [DOI] [PubMed] [Google Scholar]
- 52.Seftel A. Male hypogonadism. Part II: etiology, pathophysiology, and diagnosis. Int J Impot Res : 223–228, 2006. [DOI] [PubMed] [Google Scholar]
- 53.Seki M, Fujishima S, Nozaka T, Maeda M, Kobayashi K. Comparison of response to 17 beta-estradiol and 17 beta-trenbolone among three small fish species. Environ Toxicol Chem : 2742–2752, 2006. [DOI] [PubMed] [Google Scholar]
- 54.Sinha-Hikim I, Taylor WE, Gonzalez-Cadavid NF, Zheng W, Bhasin S. Androgen receptor in human skeletal muscle and cultured muscle satellite cells: up-regulation by androgen treatment. J Clin Endocrinol Metab : 5245–5255, 2004. [DOI] [PubMed] [Google Scholar]
- 55.Spranger B, Metzler M. Disposition of 17 beta-trenbolone in humans. J Chromatograph : 485–492, 1991. [PubMed] [Google Scholar]
- 56.Stachenfeld NS, Taylor HS. Effects of estrogen and progesterone administration on extracellular fluid. J Appl Physiol : 1011–1018, 2004. [DOI] [PubMed] [Google Scholar]
- 57.Storer TW, Woodhouse L, Magliano L, Singh AB, Dzekov C, Dzekov J, Bhasin S. Changes in muscle mass, muscle strength, and power but not physical function are related to testosterone dose in healthy older men. J Am Geriatr Soc : 1991–1999, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Sundaram K, Kumar N, Monder C, Bardin CW. Different patterns of metabolism determine the relative anabolic activity of 19-norandrogens. J Steroid Biochem Mol Biol : 253–257, 1995. [DOI] [PubMed] [Google Scholar]
- 59.Swerdloff RS, Wang C, Hines M, Gorski R. Effect of androgens on the brain and other organs during development and aging. Psychoneuroendocrinology : 375–383, 1992. [DOI] [PubMed] [Google Scholar]
- 60.Thompson SH, Boxhorn LK, Kong WY, Allen RE. Trenbolone alters the responsiveness of skeletal muscle satellite cells to fibroblast growth factor and insulin-like growth factor I. Endocrinology : 2110–2117, 1989. [DOI] [PubMed] [Google Scholar]
- 61.Tracz MJ, Sideras K, Boloña ER, Haddad RM, Kennedy CC, Uraga MV, Caples SM, Erwin PJ, Montori VM. Testosterone use in men and its effects on bone health. A systematic review and meta-analysis of randomized placebo-controlled trials. J Clin Endocrinol Metab : 2011–2016, 2006. [DOI] [PubMed] [Google Scholar]
- 62.Vandenput L, Ohlsson C. Estrogens as regulators of bone health in men. Nat Rev Endocrinol : 437–443, 2009. [DOI] [PubMed] [Google Scholar]
- 63.Wilson JD. The role of 5alpha-reduction in steroid hormone physiology. Reprod Fertil Dev : 673–678, 2001. [DOI] [PubMed] [Google Scholar]
- 64.Wilson VS, Lambright C, Ostby J, Gray LE. In vitro and in vivo effects of 17beta-trenbolone: a feedlot effluent contaminant. Toxicol Sci : 202–211, 2002. [DOI] [PubMed] [Google Scholar]
- 65.Woodhouse LJ, Gupta N, Bhasin M, Singh AB, Ross R, Phillips J, Bhasin S. Dose-dependent effects of testosterone on regional adipose tissue distribution in healthy young men. J Clin Endocrinol Metab : 718–726, 2004. [DOI] [PubMed] [Google Scholar]
- 66.Yarrow JF, Conover CF, Purandare AV, Bhakta AM, Zheng N, Conrad B, Altman MK, Franz SE, Wronski TJ, Borst SE. Supraphysiological testosterone enanthate administration prevents bone loss and augments bone strength in gonadectomized male and female rats. Am J Physiol Endocrinol Metab : E1213–E1222, 2008. [DOI] [PubMed] [Google Scholar]
- 67.Yarrow JF, McCoy SC, Borst SE. Tissue selectivity and potential clinical applications of trenbolone (17beta-hydroxyestra-4,9,11-trien-3-one): A potent anabolic steroid with reduced androgenic and estrogenic activity. Steroids : 377–389, 2010. [DOI] [PubMed] [Google Scholar]