

Abstract
Post-conflict affiliation is a mechanism favored by natural selection to manage conflicts in animal groups thus avoiding group disruption. Triadic affiliation towards the victim can reduce the likelihood of redirection (benefits to third-parties) and protect and provide comfort to the victim by reducing its post-conflict anxiety (benefits to victims). Here, we test specific hypotheses on the potential functions of triadic affiliation in Theropithecus gelada, a primate species living in complex multi-level societies. Our results show that higher-ranking geladas provided more spontaneous triadic affiliation than lower-ranking subjects and that these contacts significantly reduced the likelihood of further aggression on the victim. Spontaneous triadic affiliation significantly reduced the victim’s anxiety (measured by scratching), although it was not biased towards kin or friends. In conclusion, triadic affiliation in geladas seems to be a strategy available to high-ranking subjects to reduce the social tension generated by a conflict. Although this interpretation is the most parsimonious one, it cannot be totally excluded that third parties could also be affected by the negative emotional state of the victim thus increasing a third party’s motivation to provide comfort. Therefore, the debate on the linkage between third-party affiliation and emotional contagion in monkeys remains to be resolved.
Introduction
Conflicts in social animals can have various immediate and long-term outcomes. Immediately following a conflict, opponents may show a wide range of responses, from tolerance and avoidance of open conflict, to aggression1. Due to the potential negative consequences of agonistic interactions, such as renewed hostility towards the victim and conflict spreading across the social group, conflict management strategies are thought to have evolved to prevent and repair damage following conflicts that could be disruptive to the social group2. From an ecological point of view, conflicts can also lead to negative consequences in foraging activities, with victims spending less time searching for food because they are either excluded from access to resource or spend more time engaged in social vigilance3.
Among post-conflict behaviors, reconciliation is the main form of conflict resolution. De Waal & van Roosmalen4 first defined reconciliation as the tendency of aggressor and victim to contact each other shortly after a conflict and engage in affiliative behavioral patterns. In the subsequent decades, many studies have been devoted to exploring the dynamics and functions of the conciliatory mechanism5–8. The main advantage of reconciliation process is to end hostilities and restore the relationship between the opponents. When the aggressor and the victim share a good relationship or are kin, the probability of a conciliatory contact increases and consequently, so do the benefits derived from conflict resolution3. However, reconciliation can also entail a certain level of risk especially for the victim because approaching a former aggressor immediately after the fight can lead to a further attack9. Reconciliation may thus be absent or infrequent when aggression occurs in highly competitive contexts (e.g., feeding), if the intensity of the conflict is severe7,10 or if the conflict is highly directional11,12.
Dyadic reconciliation, however, is not the only post-conflict affiliative interaction available to limit the damage caused by aggression. Triadic post-conflict affiliation has been described in several primate (reviewed in13–15) and non-primate species (ravens, Corvus corax16; dogs, Canis familiaris17; wolves, Canis lupus18; prairie voles, Microtus ochrogaster19). Triadic post-conflict affiliation refers to affiliation between the two opponents and an uninvolved bystander (i.e. a ‘third party’). Such contacts may involve either aggressors or victims, and the third parties may be kin to either one of the opponents or unrelated to them20–23. The function of such contacts presumably varies according to the identity of the third party and whether the affiliation is directed to the aggressor or the victim13. Third party affiliation can be provided spontaneously (i.e., non-solicited or spontaneous triadic affiliation) or following a request for such affiliation by either the aggressor or the victim (i.e., solicited triadic affiliation)15–17,24.
Third party spontaneous affiliation towards the victim has been observed in a variety of mammalian and non-mammalian species (chimpanzees, Pan troglodytes4,25–32; gorillas, Gorilla spp.33–35, bonobos, Pan paniscus24,36–38, humans, Homo sapiens39, wolves18,40, ravens16, rooks41 and horses42, prairie voles19). Similar to dyadic reconciliation, the occurrence and frequency of third party affiliation is affected by several factors, such as the presence/absence of reconciliation, the relationship quality between the victim and the third party, the history of previous agonistic interactions and the level of redirection of a group (redirection is when a victim redirects aggression against a bystander)3. All these factors can affect the outcome of triadic post-conflict affiliation and do so in different ways depending on the species and the relationships of the particular individuals being observed.
When third party affiliation is spontaneously offered to socially bonded subjects (i.e., kin/friends) and significantly reduces anxiety in the victim, it can be labeled as consolation. Anxiety can be defined as an emotional state deriving from motivational conflict that can be induced by conditions of uncertainty43. If not resolved, the heightened level of anxiety can lead to reduced maintenance activities and altered social interactions by the victim, which, in turn, can have a negative impact on other group members.
In primates, consolation has been demonstrated in humans44, chimpanzees31,32 (although one study did not find evidence in this species29), bonobos24,37,38 and Tonkean macaques (Macaca tonkeana)45,46. It is interesting to note that consolation was not found in several macaque species whose social groups are based on more despotic relationships (Macaca fascicularis, M. fuscata, M. sylvanus, and M. nemestrina25). In despotic macaques, the distribution of affiliative dyadic interactions (e.g., grooming) is determined by rigid hierarchical and nepotistic rules47. In these species, the investment in creating and maintaining social bonds among unrelated subjects is extremely low and mainly focused on with high-ranking subjects47. The presence of consolation in the more tolerant Macaca tonkeana is not surprising given the extremely high levels of social bonding that extends beyond kin and more flexible dominance relationships46.
Geladas (Theropithecus gelada) live in what are referred to as multi-level societies48. The basic element of these societies consists of a One-Male Unit (OMU), a unit normally containing one adult male, six to eight adult females and their offspring. Some units may contain more than one adult male, but only one male typically copulates with the females. Several OMUs may live closely together forming a higher level of social organization. Depending on the degree of association among the constituent OMUs, such units are called “teams” if they associate regularly or “bands” if they associate less regularly49–51. Typically, adult males dominate females and the males from the different OMUs avoid interacting with the females belonging to other OMUs52,53. Geladas are characterized by female philopatry and male dispersal. The strongest social relationships within an OMU are among the related females (high level of agonistic support, embracing and grooming), with the males engaging in friendly interactions with some the of those females54. Hamadryas baboons (Papio hamadryas) also form social groups based on OMUs but these are maintained by the males aggressively herding the females. In contrast, OMU integrity in geladas results from the strong bonds among group members55. The feeding ecology of geladas probably explains why this species shows a tolerant dominance hierarchy and high levels of affiliation. Geladas’ diet is mainly based on grass, an abundant resource, which is impossible to monopolize. This can lead to a low level of competition among group members and reduced differences in foraging afforded by differences in social rank48,56. Geladas are considered a socially tolerant species, a characterization reflected in the use of coalitionary support in favor of the victims of aggression57 and by the frequent occurrence of affiliative behaviors (e.g., grooming, embracing, alloparental care) among unit members, especially among the females57–59.
Given the importance of social relationships in geladas, group cohesion could easily be disrupted by unresolved conflict, making conflict resolution particularly important in maintaining group integrity. The first study demonstrating the presence of dyadic reconciliation in geladas showed that the behavior occurs in the first two minutes after the occurrence of aggression60, a finding later confirmed by Leone and Palagi61. Because of the presence of reconciliation and their high levels of cohesiveness and tolerance, geladas are good candidates to test specific hypotheses on the potential roles of third party affiliation towards the victim, including the consolatory function.
Many hypotheses have been formulated to explain the existence/evolution of triadic post-conflict affiliation. The Self-Protection Hypothesis predicts that triadic affiliation protects the third party from becoming the victim of redirection32,62–64. Redirection occurs when the victim immediately attacks another subject not involved in the previous conflict14,23. In this perspective, redirection should be common and affiliation should be received primarily from those individuals that are frequently the target of redirection and from the individuals ranking lower than the victim (presumably more at risk). If the Self-Protection Hypothesis applies to geladas, we expect that the probability of redirection should be reduced as a result of third party affiliation.
Reconciliation between former opponents restores the relationship jeopardized by the conflict and reduces distress both in the victim and in the aggressor3. The Substitute for Reconciliation Hypothesis5 predicts that third party affiliation towards the victim can also play a role in reducing its anxiety. According to this hypothesis, we expect that geladas show third party affiliation toward the victim more frequently in the absence of reconciliation.
According to the Victim Protection Hypothesis third party affiliation protects the victim against further attacks thus providing direct benefits for the victim. This could be extremely important not only for the victim itself, but also to limit the diffusion of aggression across the group. The latter would be important in maintaining cohesion. If so, we expect that third party affiliation would significantly reduce the probability of renewed attacks on the recipient of previous aggression.
The Consolation Hypothesis predicts that third party affiliation is primarily received from a victim’s closely bonded/related partner and that such affiliation reduces the victim’s anxiety. In primates, changes in anxiety level can be assessed by scoring the occurrence of self-directed behaviors, such as scratching65–70. According to the empathic gradient hypothesis71, subjects sharing strong affiliative bonds are more prone to engage in consolatory contacts. Moreover, the victim’s anxiety would decrease because closely bonded partners (kin and friends) are supposed to be more effective in relieving anxiety, due to the good relationship quality shared with the victim25,30,64,72. If consolation is the driving mechanism of third party affiliation in geladas, we expect that it should be biased towards kin/friends and effective in reducing victim’s anxiety.
The aim of the present study was to test these hypotheses in captive geladas involving long-term data collection focused on triadic post-conflict mechanisms. Although the hypotheses are tested and discussed separately, they are not mutually exclusive, with different combinations of benefits possible among individuals with different relationships.
Results
The colony of geladas, subject of the study, was made up of two One-Male Units (OMUs). Across the four years of observation the two OMUs were composed of 47 subjects including all age-class combinations. The exact composition of each group including the age of each subject in the years observed is provided in Supplementary Table 1.
Demonstration and quantification of the phenomenon
We gathered behavioral data through the Post-Conflict/Matched-Control method [PC-MC20] resulting in 671 PC-MC pairs (see the Methods for details). To demonstrate the presence of spontaneous triadic post-conflict affiliation, we considered only the subjects with at least eight PC-MCs during in which reconciliation did not take place. For each animal we determined the number of three types pairings. In ‘attracted’ pairs affiliative contacts occurred earlier in the PC than in the MC (or they did not occur at all in the MC). In ‘dispersed’ pairs the affiliative contacts occurred earlier in the MC than in the PC (or they did not occur at all in the PC). In ‘neutral’ pairs affiliative contacts did not occur in any of the two conditions (PC and MC) or both occurred in the same minute after the ending of the conflict.
Geladas engaged in spontaneous triadic affiliation towards the victim (for an operational description of spontaneous triadic affiliation see the Method section). The rates of third party post-conflict spontaneous affiliation were higher in PCs than in MCs for the first minute of the post-conflict period (attracted pairs >dispersed pairs; Wilcoxon’s T1min = 0.00; n = 17; ties = 0; p = 0.0001; with Bonferroni correction p < 0.01; TCT1min = 26.49% ± 3.98 SE; Fig. 1).
Figure 1.
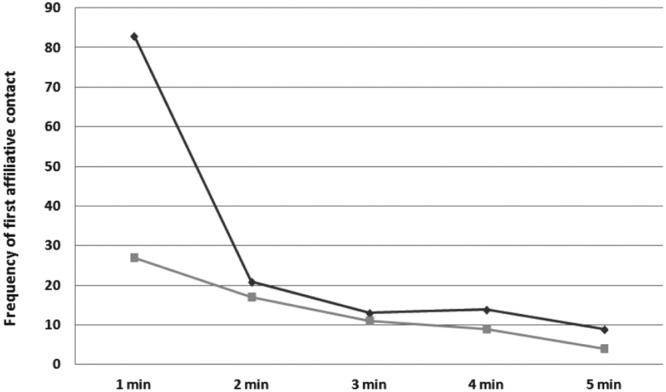
Temporal distribution of first affiliative contacts in PCs (dark grey) and MCs (light grey) for unsolicited post-conflict affiliation. Frequencies of first affiliative contacts are depicted on the Y axis. The phenomenon was present only in the first minute of observation (T1min = 0.00; n = 17; ties = 0; p = 0.0001; T2min = 5.00, n = 17, ties = 3; p = 0.502; T3min = 2.00; n = 17, ties = 11, p = 0.688; T4min = 1.00; n = 17; ties = 11, p = 0.188; T5min = 1.00, n = 17, ties = 10, p = 0.281).
To demonstrate solicited triadic post-conflict affiliation (see the Method section), we considered only the subjects with at least eight PC-MCs during which reconciliation did not take place. We did not find any evidence for the presence of solicited triadic affiliation in our study group. The Wilcoxon signed-rank test did not show any significant difference between attracted and dispersed pairs (Wilcoxon’s T1min = 6.00; n = 19; ties = 5 p = 0.187; Wilcoxon’s T2min = 6.00; n = 19; ties = 8; p = 0.464; Wilcoxon’s T3min = 4.00; n = 19; ties = 10; p = 0.516; Wilcoxon’s T4min = 7.00; n = 19; ties = 10; p = 0.148; Wilcoxon’s T5min = 5.00; n = 19; ties = 10; p = 0.973).
Due to the absence of solicited triadic affiliation all further analyses were focused on spontaneous triadic affiliation. We measured the ‘Triadic Contact Tendency’ (TCT) defined as the number of attracted minus the number of dispersed pairs divided by the total number of PC–MC pairs (see Methods for further details). The most frequent patterns used by animals when engaging in spontaneous triadic affiliation were playing, affiliative body interactions (touching/embracing) and affiliative facial expressions and vocalizations (lip-smacking/grunting + moan). Playing (mean TCTplay = 42.71 ± 17.61 SE), body interactions (TCTtouching/embracing = 23.53 ± 11.02 SE) and affiliative facial expressions and vocalizations (TCTlip-smacking/grunting+moan = 54.76 ± 16.98 SE) were significantly higher during the first minute of PCs than during the first minute of MCs (Wilcoxon’s Tplaying = 9.00; n = 16; ties = 4; p = 0.018; Wilcoxon’s Ttouching/embracing = 14.00; n = 19; ties = 6; p = 0.028; Wilcoxon’s Tlip-smacking/grunting+moan = 10.00; n = 14; ties = 1; p = 0.011, with Bonferroni correction p < 0.02). On the other hand, we did not find any difference between attracted and dispersed pairs for contact sitting (Wilcoxon’s Tcontact sitting = 31.50; n = 17; ties = 2 p = 0.115) and grooming (Wilcoxon’s Tgrooming = 20.00; n = 14; ties = 4; p = 0.590).
Self-Protection Hypothesis
Of the 29 different victims of aggression, only six individuals showed redirection. The mean level of redirection calculated on the six subjects indicates that the phenomenon was rare (1.31% ± 0.38 SE).
Victim Protection Hypothesis
The rate of renewed conflicts towards the victim in absence of reconciliation and third-party affiliation was significantly higher in PC compared to MC in the first four minutes after the aggression (Wilcoxon’s T1min = 14.50; ties = 3; N = 27; p = 0.0001; Wilcoxon’s T2min = 0; ties = 9; N = 27; p = 0.0001; Wilcoxon’s T3min = 12; ties = 11; N = 27; p = 0.004; Wilcoxon’s T4min = 18; ties = 12; N = 27; p = 0.017; Wilcoxon’s T5min = 17; ties = 16; N = 27; p = 0.154; with Bonferroni correction p < 0.01; Supplementary Fig. 1) To understand the effect of triadic affiliation in absence of reconciliation, we selected the 2–4 min time window for the analysis of renewed aggression. In the following analysis, the 1 min was excluded because it was characterized by the presence of the triadic affiliation (Fig. 1). The 2–4 min time window was selected because in this period the rate of PC renewed aggression remained significantly higher than the rate of aggression recorded in the same time window in the MC. The 5 min was excluded because the rates of aggression did not differ between the PC and the MC. The frequency of renewed aggression significantly differed across the three conditions: without spontaneous third-party affiliation (PC-no contact), following spontaneous third-party affiliation (PC-contact), and matched-control (MC) (Friedman’s Chi-square = 20.414, N = 18, df = 2, p = 0.0001). We found that renewed aggression in PC-no contact were significantly more frequent than those in PC-contact (post-hoc Dunnett’s test: q = 5.00, p < 0.001) and in the MC (PC-no contact vs MC contact: Dunnett’s test: q = 5.18, p < 0.001). Finally, we did not find any significant difference between renewed aggression in PC-contact and MC (Dunnett’s test: q = 0.47, ns; Fig. 2) (with Bonferroni correction p < 0.02).
Figure 2.
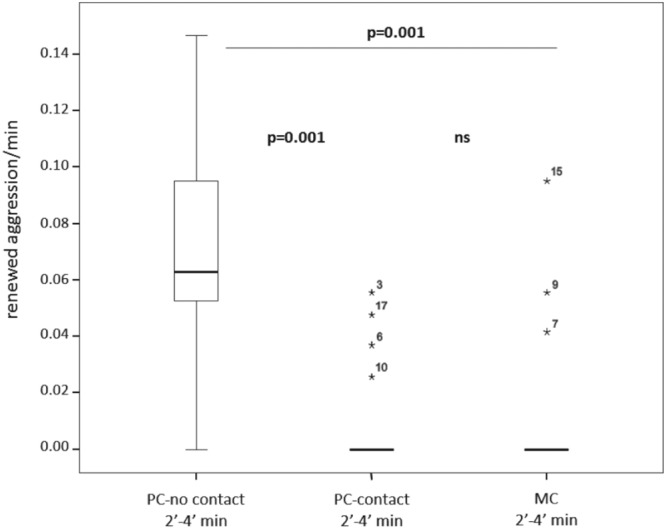
Boxplots showing the rate of renewed aggression per minute of observation (2–4 min) in absence of postconflict third-party affiliation (PC-no contact; in absence of reconciliation), in presence of post-conflict third-party affiliation (PC-contact; in absence of reconciliation) and in absence of conflict (MC). The box plots show the median and 25th and 75th percentiles; the numbered whiskers indicate the outlier values within 1.5 times the interquartile range, IQR. N = 18.
Substitute for Reconciliation Hypothesis
Third-party affiliation was more frequent in absence of reconciliation than in its presence (Wilcoxon’s T = 8.00; ties = 1; N = 29; p = 0.0001).
Consolation Hypothesis
Scratching rates during PCs-no contact were significantly higher than those during MCs across a 4-min time window (2–5 min), therefore the analysis of scratching in presence of spontaneous third-party affiliation was limited to that time-window (Supplementary Fig. 2). Scratching rates were significantly different across all the three conditions: without spontaneous third-party affiliation (PC-no contact), following spontaneous third-party affiliation (PC-contact), and matched-control (MC) (Friedman’s Chi-square = 12.52, N = 22, df = 2, p = 0.001). We found that scratching rates in PCs-contact were lower than those in MCs (post-hoc Dunnett’s test: q = 2.81, p < 0.01). Both scratching rates in PCs-contact and in MCs were significantly lower than those recorded in PCs-no contact (PC-contact vs PC-no contact: Dunnett’s test: q = 3.48, p < 0.001; MC vs PC-no contact: Dunnett’s test: q = 2.11, p < 0.05) (Fig. 3) (with Bonferroni correction p < 0.02).
Figure 3.
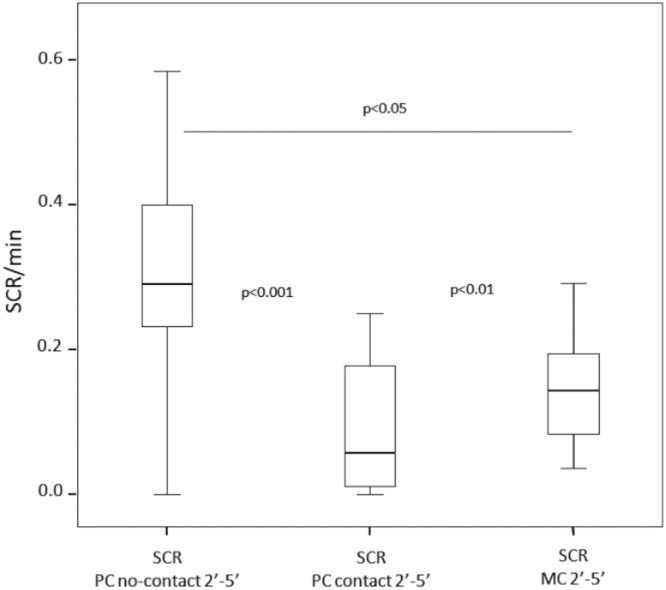
Boxplots showing the scratching per minute of observation (2–5 min) in absence of postconflict third-party affiliation (PC-no contact; in absence of reconciliation), in presence of post-conflict third-party affiliation (PC-contact; in absence of reconciliation) and in absence of conflict (MC).
To verify which factors (Table 1) affected the Triadic Contact Tendency (TCT) levels between victims and third party, we ran a Linear Mixed Model (LMM). We tested models for each combination involving the variables of interest, spanning from a single-variable model to a model including all the fixed factors (full model). All possible variable combinations were tested. The best model included the interaction of kinship with bonding (“kinship*bondinggr”), the sexes forming the dyad (sexcombination), the age of the subjects forming the dyad (agecombination), the rank of the victim (Normalized David’s Scorevictim) and the rank of the third-party (Normalized David’s Scorethird-party) (for the best model Akaike’s Corrected Information Criterion, AICc = 535.92; for the next-best model Akaike’s Corrected Information Criterion, AICc = 550.08; ΔAIC = 14.16) and explained about 94.18% of the distribution (see Table 2 for the AICc of the second and third model tested; AICc of the intercept = 612.318). Within this model only Normalized David’s Scorethird-party was a statistically significant factor with high-ranking individuals providing more spontaneous contacts than low ranking individuals (Table 3; Fig. 4).
Table 1.
Description of variables used in LMM analyses. NDS = Normalized David’s Scores.
| NAME | TYPE |
|---|---|
| Dependent Variable | |
| TCT | Scale (Triadic Contact Tendency) |
| Fixed Explanatory variables | |
| Individual characteristics | |
| Sex Combination | Nominal (00 = male-male; 01 = male-female; 10 = female-male; 11 = female-female) |
| Age Combination | Nominal (11 = adult-adult, 00 = immature-immature, 10 = adult-immature, 01 = immature-adult) |
| NDSVICTIM | Scale |
| NDSTHIRD-PARTY | Scale |
| Relational characteristics | |
| Kinship | Dichotomous (1 = kin, 0 = non-kin) |
| Affiliation levels | |
| Social bonding (grooming) | Ordinal (0 = week, 1 = medium, 2 = strong) |
| Random Variables | |
| VictimID*Third-PartyID | Nominal |
Table 2.
Table showing the fixed variables included in the three models showing the lowest values of the Akaike Corrected Information Criterion (AICC).
| Random effects | Fixed effects | AICc | Δ AICc | EXP(−0.5 * Δi) | Wi |
|---|---|---|---|---|---|
| VictimID*Third-PartyID | Kinship*Bondinggr;NDSvictim;NDSthird-party;SEXcombination;AGEcombination | 535.92 | 0.00 | 1.00 | 0.94 |
| Bondinggr; NDSvictim;NDSthird-party;SEXcombination;AGEcombination;Kinship | 550.08 | 14.16 | 0.03 | 0.03 | |
| Bondinggr;NDSthird-party;SEXcombination;AGEcombination;Kinship | 551.89 | 15.97 | 0.02 | 0.02 |
The difference between the AIC of the best model and the AIC of each other model (ΔAICc) and Akaike Weight (Wi) are also reported. NDS = Normalized David’s Scores.
Table 3.
Best LMM explaining the frequency of TCT (AICc = 535.920).
| TCT (AICc = 547.874) | F | df1 | df2 | Sig. |
|---|---|---|---|---|
| Fixed Explanatory Variables | ||||
| Intercept | 0.855 | 13 | 59 | 0.602 |
| SEX combination | 0.700 | 3 | 59 | 0.556 |
| AGE combination | 0.321 | 3 | 59 | 0.810 |
| KIN*BONDgr | 0.816 | 5 | 59 | 0.543 |
| NDSVICTIM | 0.132 | 1 | 59 | 0.718 |
| NDSTHIRD-PARTY | 5.981 | 1 | 59 | 0.017 |
| Random variables | Variance | |||
| Victim*Third-party ID | 1.013 | 0.311 | ||
Figure 4.
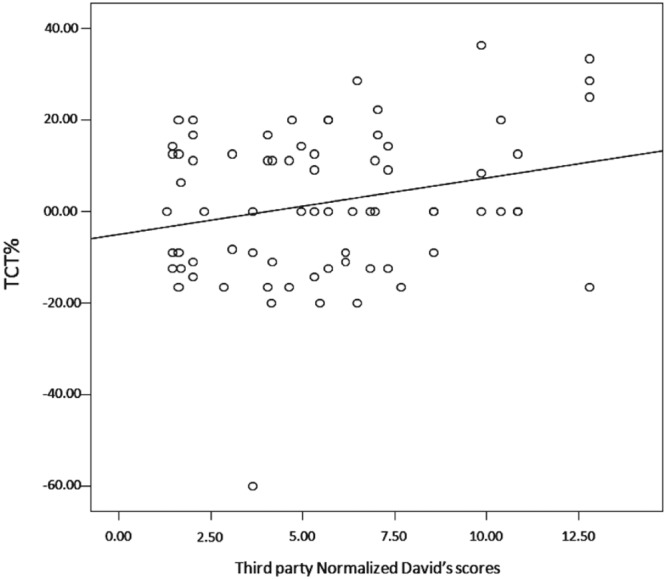
Scatter plot showing the positive correlation between TCT% values and Third-party Normalized David’s scores (R2 = 0.06).
Discussion
To understand why and how animals respond to agonistic interactions provides valuable insights into the evolution of socio-cognitive and emotional capacities, especially if comparative studies are conducted by applying the same methodological approaches across different species18,30,45,57,63. The presence of unsolicited triadic affiliation in geladas permitted the testing some hypotheses about the potential functions of such social behavior. The findings from this study could then be compared to those from other primate and non-primate species. Our data do not support the Self-Protection hypothesis, but do provide support for the Victim Protection and the Substitute for Reconciliation hypotheses. As only two of the three criteria defining consolation were met, our findings cannot draw any clear-cut conclusions about the Consolation hypothesis. In the following sections we discuss our findings with regard to these hypotheses and their implications for a better understanding of the functions of post-conflict affiliation in geladas.
Geladas engaged in unsolicited triadic affiliation towards the victim in the first minute after a conflict (Fig. 1) and this is consistent with the results from several other primate35,36,45,72 and non-primate species18,19,41. The most surprising result is the absence of the solicited version of triadic contacts. Interestingly, while both spontaneous and solicited triadic contacts are present in bonobos, only spontaneous triadic contacts lead to the relief from distress in the victim24. This result underlines the functional dichotomy of spontaneous and non-spontaneous post-conflict affiliation. It seems that also in geladas the spontaneity of the contact is a key factor in triadic post-conflict affiliation.
In geladas, reconciliation60,61 and third party affiliation are both present, at least in captive populations. Our results show that when reconciliation fails to occur, the frequency of third party affiliation increases (supporting the Substitute for Reconciliation hypothesis). Detecting general patterns of post-conflict affiliative contacts is relatively easy, but parsing out the proximate and ultimate causes of post-conflict mechanisms is more challenging given the strong dependence of affiliative interactions on a wide range of variables related both to the social context and to the variability in the types of affiliative contacts preferentially used by different individuals4,27,29,73,74. Leone and Palagi61 demonstrated that geladas living in the same colony did not show any preference in the behavioral pattern used to reconcile (e.g., grooming, contact sitting, lip-smacking/vocalizations). On the contrary, our findings suggest that bystanders selectively engaged in short play bouts, affiliative body interactions (touching/embracing) and facial expressions and vocalizations (lip-smacking/grunting + moan) to affiliate with the victim. Therefore, although third party affiliation can function as a substitute for reconciliation, these two conflict management phenomena are expressed differently. Play was generally preferred by juveniles/subadults, even though adults also exchanged some playful contacts. A similar result was also found in gorillas34. In this species the immature animals, who were the best “consolers”, engaged in short play bouts with the victims to affiliate with them34. In geladas, third parties preferred engaging in rapid behavioral patterns, such as play, facial expressions and vocalizations, possibly because of the high level of arousal of the performers in these post-conflict contexts. Social grooming and sitting closely together with bodily contact tend to occur under more relaxed conditions.
Many authors describe triadic, post-conflict affiliation as an investment from which both victims and bystanders should gain direct and/or indirect benefits3,36,75–77. Given that among group living primates, victims can redirect aggression towards others, it is even possible to hypothesize that bystanders should avoid interacting with the recipient of aggression78,79. This is not the case of geladas, where triadic post-conflict interactions do not seem to be so risky given the extremely low levels of redirected aggression. In this view, the Self Protection hypothesis cannot explain our results. Macaca tonkeana, a species sharing with Theropithecus gelada tolerant social relationships and third party affiliation, also lacks redirection45. In contrast, other species of the Macaca genus, which have more despotic social relationships, show high levels of redirection and the absence of third party affiliation45,80–82. Our results also contrast with the function of spontaneous third-party affiliation found in mandrills (Mandrillus sphinx), a more despotic species63,83. Schino and Marini63 found that victims received most affiliation from those bystanders that were frequently the target of redirection and that bystander affiliation reduced the probability of redirected aggression. Therefore, in mandrills triadic post-conflict affiliation functions as a self-protection strategy, as the third-party can gain an immediate benefit. It appears, therefore, clear that post-conflict triadic affiliation works differently depending on the social style of the species.
From the victim’s perspective, receiving spontaneous triadic affiliation can provide short-term benefits. We found that after the triadic contact, the scratching rates of the victim significantly decreased, thus demonstrating that this kind of affiliation can reduce anxiety in the victim (Fig. 3). Moreover, triadic post-conflict affiliation protects the victim against further attacks, as the frequency of renewed aggression decreased after third-party interactions had occurred (supporting the Victim Protection hypothesis) (Fig. 2). A previous study carried out on the same colony of geladas showed that the probability of renewed aggression on the victim was not significantly affected by the presence of reconciliation61. These differing findings suggest that spontaneous third party affiliation and dyadic reconciliation can have different functions, leading to different social outcomes.
Even though the Victim Protection hypothesis has been rarely tested, third party affiliation has now been shown to protect victims in four primate species (chimpanzees28, bonobos24, Tonkean macaques45, geladas in the present study). As far as we know, only two studies have explicitly tested this hypothesis in species of monkeys with differing social styles. Third party affiliation in the egalitarian Tonkean macaque protects the victim from further attacks45, whereas such effect was not found in mandrills63, a species with a more despotic social style83. The apparently contrasting results deriving from these studies are in line with the different social dynamics characterizing the two species and, more specifically, with their different levels of tolerance sensu84.
The distribution of the triadic consolatory tendency (TCT) in our study was significantly affected by the rank of bystanders, although other factors also converged in explaining the best model (see Tables 2 and 3). The highest-ranking bystanders provided the highest levels of triadic affiliation to the victim (Fig. 4). In the same study group, we found that dominance rank also played a role in agonistic support, which was mainly provided by high-ranking individuals to the victims of the ongoing aggression57. In geladas, supporting the victim during the conflict and providing protection and comfort when the conflict ceased, may be driven by the so-called ‘community concern’, a pacifying motivation to maintain the stability of the group which is beneficial for all group members57,85,86. Therefore, triadic affiliation and agonistic support could be viewed as two sides of the same coin, providing protection to the victim and, consequently, bringing benefits to the whole group.
The effects of triadic affiliation on reducing anxiety and lowering the probability to be the target of renewed aggression, are two direct benefits gained by the victim. But the finding that affiliation was not biased toward either kin or friends, supports that the main benefit for the third party involved is related to group stability and cohesion. Based on our data, there are two possible, but not mutually exclusive, explanations linked to the motivations driving the third-party towards the victim (Fig. 5). Firstly, the third-party could be affected by the behavioural arousal shown by the victim and be motivated to reduce it by providing comfort. Secondly, the motivation leading the third-party to affiliate with the victim is the reduction of renewed aggression. By protecting the victim against potential renewed aggression, the third party’s intervention can break the cycle of aggression57. This may be especially true, if as suggested by our data, that affiliation is provided by the highest-ranking members of the group. In geladas57, as in many other primate species14, highest-ranking individuals play an important role in conflict management because their interventions are more effective in maintaining peaceful relationships57. The affiliative contact offered by high-ranking third party animals is most effective in reducing the likelihood of renewed aggression towards the victim as well as their level of anxiety.
Figure 5.
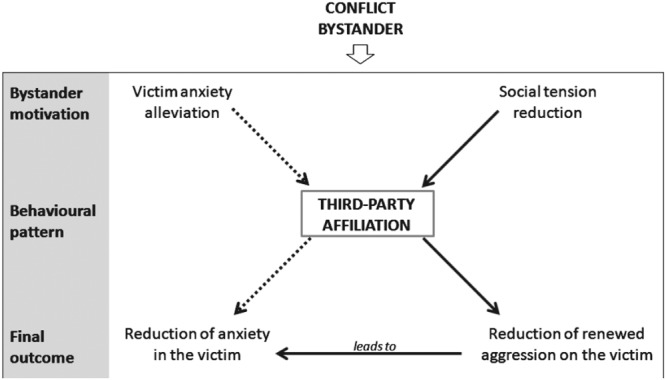
The model we proposed on the basis of our findings on geladas. For the explanation see the Discussion.
The reduction of victim’s anxiety together with a bias towards kin and friends are the two main prerequisites that must be satisfied to consider spontaneous third-party affiliation as consolation. Therefore, our data do not explicitly support the Consolation hypothesis, even though they support the Victim Protection and the Substitute for Reconciliation hypotheses. We want to emphasize that the reduction of victim’s anxiety could also be a by-product deriving from a conflict management strategy enacted by high-ranking individuals to reduce further aggression (Fig. 5).
To determine the motivation driving subjects to engage in triadic affiliation, we should focus on the third party. For example, we should consider if the third party shows higher levels of anxiety after perceiving the emotional state of the victim as it has been demonstrated in prairie voles under laboratory conditions19. Obviously, this kind of assessment is very difficult to perform under naturalistic conditions, especially in species living in large groups where numerous bystanders could become the third party affiliating with the victim.
The direct influence of social bonding and kinship on the distribution of triadic contacts could be less easily detectable in highly cohesive groups87 characterized by small differences between the levels of affiliation (as measured by grooming) of each dyad. Furthermore, the same argument can be applied to the quality of relationships measured by the level of relatedness, which is particularly high between gelada females belonging both to the same One-Male Unit (OMU) and even to different OMUs as demonstrated by Snyder-Mackler et al.88. For this reason, it could be extremely interesting to expand the study to wild populations in order to compare the level of reconciliation and third-party affiliation within each OMU and across different OMUs.
Methods
Subjects and housing
The colony of geladas, housed at the NaturZoo (Rheine, Germany), is held in two enclosures with both an indoor (a room about 36 m2) and outdoor facility (an island of 2,700 m2 surrounded by a boundary ditch). The size of the island allows the scattering of geladas and, consequently, the formation of small groups with variable composition. The enclosures are equipped with environmental enrichments allowing geladas to move freely in all three dimensions. Specifically, the outside enclosure is located in an open naturally hilly area equipped with trees, branches, ropes, and dens. The animals were fed with grass, vegetables, and pellets, which were scattered on the ground twice a day (9:30 a.m., 2:30 p.m.). Water was available ad libitum. No stereotypic or aberrant behaviors have been observed in this group.
Individual identification of the animals was based on sex, age, and distinctive external features (scars, size, missing fur patches, fur color, and facial traits). Thanks to the accurate birth recording made by the staff of the zoo, kinship was known. In 2007, the two OMUs were housed in the same enclosure and, in 2009, 2010 and 2011 they lived separately in two different enclosures.
This study was approved by The University of Pisa (Animal Care and Use Board). Since the study was purely observational the committee waived the need for a permit.
Data collection
We collected behavioral data during a 6-month period in 2007 (June–November), a 4-month period in 2009 (June–September), a 2-month period in 2010 (July-August) and a 2-month period in 2011 (July-August). Data were collected through voice- and video-recorders. We gathered 1,809 hours of observations, which took place daily over 6-hr periods spanning morning (from 6:00 a.m.) and evening (until 10:00 p.m.) and all hours were equally represented.
Six observers (two per period) collected the data, with all observers being trained by the same person (the first author). The presence of at least two observers was necessary for the concurrent use of different techniques. Training was over when the observations produced a Cohen’s kappa higher than 0.8089. We checked for observation reliability regarding the behaviors object of this study (aggression, touching, playing, grooming, contact sitting, lip-smacking, grunting/moan, scratching) at the beginning of each month and we never obtained values below 0.80.
We defined scratching (used to measure primate anxiety)65,66,68–70 as a repeated movement of the hand or foot during which the fingertips are drawn across the individual’s fur. A new scratching event was assigned when the scratched body part changed or when scratching was resumed after more than 5 seconds.
Agonistic events were collected using all-occurrences sampling90. Then, after the last aggressive action of any agonistic event, we followed the victim through the focal-animal sampling90 for a 5-min post-conflict observation (PC). Matched Control focal observations (MC), were conducted the day after the conflict at the same time as the original PC and in absence of any agonistic interaction in the preceding 5 minutes20. Moreover, we started observing the victim for the MC when the opponents had the opportunity to socially interact (they could see each other) but were not interacting20,91. The rare polyadic interactions (involving >2 aggressors but only one victim) were split into dyadic components92. For both PCs and MCs we recorded (1) starting/ending time (minute), (2) affiliative behaviors (grooming, contact-sitting, play, touching, lip-smacking, grunting, moan) between the victim and other group members, (3) victim’s scratching (see the definition in the Result section), (4) identity of individuals interacting with the victim (5) exact moment of each interaction within the five-minute time window.
The first individual affiliating with the victim in the MCs could be every individual of the group with the exclusion of the individual who had attacked the victim in the PC (i.e., the aggressor). The selection of one specific individual as third party in the MC would produce a strong statistical bias in the evaluation of the presence of the phenomenon. For this reason, in the MC we considered as first third party each possible group mate (the previous aggressor of the corresponding PC was clearly excluded).
An aggressive event was defined as redirection when a victim attacked a group member (different from the former aggressor) within five minutes after the previous attack (PC duration).
During each focal (PC), the first affiliative contact following an aggressive event could occur between the victim and the aggressor (reconciliation), be directed by a “third party” (an individual other then the aggressor) toward the victim (spontaneous triadic party affiliation), or by the victim toward a “third party” (solicited triadic party affiliation)28,29,36,62. Specifically, when the bystander approached the victim and initiated the first affiliative contact interaction toward the victim the triadic party affiliation was defined as ‘spontaneous’. Instead, when the victim approached a bystander and initiated the first affiliative interaction, the triadic party affiliation was considered as ‘solicited’. The cases in which the initiator was unclear were excluded from the analysis.
For each animal we determined the number of attracted, dispersed and neutral pairs over all PC-MC pairs. In attracted pairs, affiliative contacts occurred earlier in the PC than in the MC (or they did not occur at all in the MC), whereas in dispersed pairs the affiliative contacts occurred earlier in the MC than in the PC (or they did not occur at all in the PC). In neutral pairs, affiliative contacts occurred during the same minute in the PC and the MC, or no contact occurred in either the PC or the MC.
We extracted background information on social bonding among group members using the grooming interactions recorded during focal observations other than PCs and MCs. We followed each focal subject (a single bout focal = 30 min) at different times to obtain data covering the entire day (about 6 hours of observations) in balanced proportions. Every two days all the subjects were observed at least 30 minutes.
Definitions and data analyses
We measured the ‘Triadic Contact Tendency’ (TCT) defined as the number of attracted minus the number of dispersed pairs divided by the total number of PC–MC pairs93. To evaluate which behavior was preferentially used in third-party affiliation, via the same methodology we calculated TCTs for the five different behaviors involved in the post-conflict third-party affiliation: affiliative body interactions (touching/embracing), contact sitting, playing, grooming, facial expressions/vocalizations (lip-smacking/grunting + moan). The lip-smacking facial expression has an affiliative function in geladas and consists in a protrusion of the lips that are smacked together repeatedly. Grunting and moan in geladas, as it occurs for baboon species94,95, have an important role in the affiliation mechanisms96,97.
For each of the behaviours we scored attracted and dispersed pairs (see)62. We made use of non-parametric statistics when the data did not have a normal distribution. Each non-parametric test was performed at an individual level (each subject compared only once to avoid the problem linked to the pseudo replication of the data. To check for the presence of the post-conflict phenomena we applied the Wilcoxon signed-rank test. We used the non-parametric ANOVA Friedman’s test to compare scratching rates across three conditions: PC-no contact (absence of third-party affiliation), PC-contact (after third-party affiliation, excluding cases with reconciliation) and MC. Scratching rates during PCs-no contact were significantly higher than those during MCs in the 2–5 min time window, therefore only this period (4 minutes) was considered as relevant for the Friedman test (see Supplementary Fig. 1). Only the events not characterized by reconciliation were considered for this analysis. In case of significance across the three conditions, we ran the Dunnett’s multiple comparison test (post-hoc test) to determine which pairs of conditions differed significantly98. In case of multiple comparisons we adjusted the p-value according to the Bonferroni’s correction.
The analysis was performed minute-by-minute to ascertain that the decrease of the phenomena analyzed was actually due to the contact provided and not to their natural fluctuating over time after the agonistic contact30.
The dyadic TCT variable was normally distributed (Kruskal-Wallis, ns). Via Linear Mixed Models (used for normal distribution, LMM) we evaluated the influence of individual and relationship characteristics on TCT values. Individual characteristics included age class (adult or immature) and sex. Relationship characteristics included rank, kinship and affiliation levels. Rank was measured by using Normalized David’s Scores (Table 1). Normalized David’s scores (NDS) were calculated on the basis of a dyadic dominance index (Dij) in which the observed proportion of wins (Pij) is corrected for the chance occurrence of the observed outcome. The chance occurrence of the observed outcome is calculated on the basis of a binomial distribution with each animal having an equal chance of winning or losing in every dominance encounter99. The correction is necessary when, as in the case of our study groups, the interaction numbers greatly differ between dyads. We considered as kin-related those individuals belonging to grandmother/mother/offspring dyads and siblings (r ≥ 0.25), the others were considered non-related (non-kin). Affiliation levels were determined using grooming interactions within each dyad. Affiliation rates across dyads were arranged according to a decreasing order. We categorized the relationship quality of the dyads as strong if their grooming levels fell into the upper quartile and as medium if their grooming levels fell into the inter-quartile. All the weakly bonded dyads fell in the bottom quartile. Victim’s and third-party’s identities were entered as random factors (nominal variables) (Table 1).
We tested models for each combination involving the variables of interest, spanning from a single-variable model to a model including all the fixed factors (full model). All possible variable combinations were tested. To select the best model we used the Akaike’s Corrected Information Criterion (AICc), a measure for comparing mixed models based on the −2 (Restricted) log likelihood. The AICc corrects the Akaike’s Information Criterion (AIC) for small sample sizes. As the sample size increases, the AICc converges to AIC. The AICc allows a comparison of multiple competing models and an estimation of the best approximation of the ‘true’ process underlying the biological phenomenon under study. The model with a lower value of AICc was considered to be the best model. To quantitatively select the best model we used Akaike weights (wi). These weights provide a reliable measure of the strength of evidence for each model, and represents the ratio of delta AICc (Δi) values for each model relative to the whole set of R candidate models91. The value of the Akaike weight (wi) indicates the probability that the model is the best among the whole set of candidate models. As suggested by Symonds and Moussalli100, “… models with Δi values greater than 10 are sufficiently poorer than the best AIC model as to be considered implausible” (p. 17). All tests were performed via SPSS 20.0.
Electronic supplementary material
Acknowledgements
We thank Johann Achim, the Director of the RheineZoo, for permission to work at the zoo and to the gelada keepers for allowing and facilitating this work; Giada Mancini and Virginia Pallante for collaboration in data collection. Thanks are also due to W. Rebo for his important clarifying input in discussing results. We wish to thank Sergio Pellis for his accurate linguistic revision. This research was supported with private funding and complies with current laws of Italy and Germany.
Author Contributions
E.P. designed the study and trained all the observers. E.P., A.L., E.D., P.F.F. carried out the study on their own private expenses. A.L. collected data. E.P., A.L. and E.D. analysed data. E.P., A.L., E.D., P.F.F. wrote the manuscript. All authors have read and approved the manuscript for publication.
Availability of Data
Data have been submitted as Supplementary Materials.
Competing Interests
The authors declare no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Elisabetta Palagi, Email: elisabetta.palagi@unipi.it.
Pier Francesco Ferrari, Email: pierfrancesco.ferrari@isc.cnrs.fr.
Electronic supplementary material
Supplementary information accompanies this paper at 10.1038/s41598-018-33548-y.
References
- 1.de Waal FBM. Primates - a natural heritage of conflict resolution. Science. 2000;289:586–590. doi: 10.1126/science.289.5479.586. [DOI] [PubMed] [Google Scholar]
- 2.Preuschoft, S., van Schaik, C. P. Dominance and communication. In Natural Conflict Resolution (ed. Aureli, F., de Waal, F. B. M.) 5, 77–105 (Berkeley University of California Press, 2000).
- 3.Aureli F, Cords M, van Schaik CP. Conflict resolution following aggression in gregarious animals: A predictive framework. Anim. Behav. 2002;64:325–343. doi: 10.1006/anbe.2002.3071. [DOI] [Google Scholar]
- 4.de Waal FBM, van Roosmalen A. Reconciliation and consolation among chimpanzees. Behav. Ecol. Sociobiol. 1979;5:55–66. doi: 10.1007/BF00302695. [DOI] [Google Scholar]
- 5.Aureli, F., de Waal, F. B. M. In Natural Conflict Resolution (ed. Aureli, F., de Waal, F. B. M.) 1, 3–12 (Berkeley University of California Press, 2000).
- 6.Cordoni G, Palagi E. Reconciliation in wolves (Canis lupus): new evidence for a comparative perspective. Ethology. 2008;114:298–308. doi: 10.1111/j.1439-0310.2008.01474.x. [DOI] [Google Scholar]
- 7.Palagi E, Antonacci D, Norscia I. Peacemaking on treetops: first evidence of reconciliation from a wild prosimian. Anim. Behav. 2008;76:737–747. doi: 10.1016/j.anbehav.2008.04.016. [DOI] [Google Scholar]
- 8.Palagi E, Cordoni G, Demuru E, Bekoff M. Fair play and its connection with social tolerance, reciprocity and the ethology of peace. Behaviour. 2016;153:1195–1216. [Google Scholar]
- 9.Cordoni G, Palagi E. Being a victim or an aggressor: Different functions of triadic post-conflict interactions in wolves (Canis lupus lupus) Aggr. Behav. 2015;41(6):526–536. doi: 10.1002/ab.21590. [DOI] [PubMed] [Google Scholar]
- 10.Norscia I, Palagi E. Do wild brown lemurs reconcile? Not always. J. Ethol. 2011;29:181–185. doi: 10.1007/s10164-010-0228-y. [DOI] [Google Scholar]
- 11.Aureli F, van Schaik CP, van Hooff JARAM. Functional aspects of reconciliation among captive long-tailed macaques (Macaca fascicularis) Am. J. Primatol. 1989;19:39–51. doi: 10.1002/ajp.1350190105. [DOI] [PubMed] [Google Scholar]
- 12.Arnold, K., Aureli, F. Postconflict reconciliation. Primates in Perspective, 2 (2007).
- 13.Das, M. Conflict management via third parties: post-conflict affiliation of the aggressor. In Natural Conflict Resolution (ed. Aureli, F., de Waal, F. B. M.) 13, 263–266 (Berkeley University of California Press, 2000).
- 14.Watts, D. P., Colmenares, F., Arnold, K. Redirection, consolation and male policing: how target of aggression interact with bystanders. In Natural Conflict Resolution (ed. Aureli, F., de Waal, F. B. M.) 14, 281–288 (Berkeley University of California Press, 2000).
- 15.Palagi E, Chiarugi E, Cordoni G. Peaceful post-conflict interactions between aggressors and bystanders in captive lowland gorillas. Am. J. Primatol. 2008;70:949–955. doi: 10.1002/ajp.20587. [DOI] [PubMed] [Google Scholar]
- 16.Fraser ON, Bugnyar T. Do ravens show consolation? Responses to distressed others. PLoS ONE. 2010;5:e10605. doi: 10.1371/journal.pone.0010605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Cools. AKA, van Hout AJM, Nelissen MHJ. Canine reconciliation and third-party initiated postconflict affiliation: do peacemaking social mechanisms in dogs rival those of higher primates? Ethology. 2008;114:53–63. doi: 10.1111/j.1439-0310.2007.01443.x. [DOI] [Google Scholar]
- 18.Palagi E, Cordoni G. Postconflict third-party affiliation in Canis lupus: do wolves share similarities with the great apes? Anim. Behav. 2009;78:979–986. doi: 10.1016/j.anbehav.2009.07.017. [DOI] [Google Scholar]
- 19.Burkett JP, et al. Oxytocin-dependent consolation behavior in rodents. Science. 2016;351:375–378. doi: 10.1126/science.aac4785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.de Waal FBM, Yoshihara D. Reconciliation and redirected affection in rhesus monkeys. Behaviour. 1983;85:224–241. doi: 10.1163/156853983X00237. [DOI] [Google Scholar]
- 21.Judge PG. Dyadic and triadic reconciliation in pigtail macaques (Macaca nemestrina) Am. J. Primatol. 1991;23:225–237. doi: 10.1002/ajp.1350230403. [DOI] [PubMed] [Google Scholar]
- 22.Das M, Penke Z, van Hooff JARAM. Affiliation between aggressors and third parties following conflicts in long-tailed macaques (Macaca fascicularis) Int. J. Primatol. 1997;18:159–181. doi: 10.1023/A:1026316501593. [DOI] [Google Scholar]
- 23.Castles DL, Whiten A. Post-conflict behaviour of wild olive baboons. I. Reconciliation, redirection and consolation. Ethology. 1998;104:126–147. doi: 10.1111/j.1439-0310.1998.tb00057.x. [DOI] [Google Scholar]
- 24.Palagi E, Norscia I. Bonobos protect and console friends and kin. PloS One. 2013;8(11):e79290. doi: 10.1371/journal.pone.0079290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.de Waal, F. B. M., Aureli, F. Consolation, reconciliation and a possible difference between macaques and chimpanzees. In Reaching into thought: The Minds of Great Apes. (ed. Russon, A. E., Bard, K. A., Parker, S. T.) 80–110 (Cambridge Cambridge University Press 1996).
- 26.Wittig RM, Boesch C. ‘Decision-making’ in conflicts of wild chimpanzees (Pan troglodytes): an extension of the relational model. Behav. Ecol. Sociobiol. 2003;54:491–504. doi: 10.1007/s00265-003-0654-8. [DOI] [Google Scholar]
- 27.Kutsukake N, Castles DL. Reconciliation and post-conflict third-part affiliation among wild chimpanzees in the Mahale Mountains, Tanzania. Primates. 2004;45:157–165. doi: 10.1007/s10329-004-0082-z. [DOI] [PubMed] [Google Scholar]
- 28.Palagi E, Cordoni G, Borgognini SB. Possible roles of consolation in captive chimpanzees (Pan troglodytes) Am. J. Phys. Anthropol. 2006;129:105–111. doi: 10.1002/ajpa.20242. [DOI] [PubMed] [Google Scholar]
- 29.Koski SE, Sterck E. Triadic postconflict affiliation in captive chimpanzees: does consolation console? Anim. Behav. 2007;73:133–142. doi: 10.1016/j.anbehav.2006.04.009. [DOI] [Google Scholar]
- 30.Fraser ON, Stahl D, Aureli F. Stress reduction through consolation in chimpanzees. Proc. Natl. Acad. Sci. USA. 2008;105:8557–8562. doi: 10.1073/pnas.0804141105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fraser ON, Koski SE, Wittig RM, Aureli F. Why are bystanders friendly to recipients of aggression? Commun. Integr. Biol. 2009;2:285–291. doi: 10.4161/cib.2.3.8718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Romero T, Castellanos MA, de Waal FBM. Consolation as possible expression of sympathetic concern among chimpanzees. Proc. Nat. Acad. Sci. USA. 2010;107:12110–12115. doi: 10.1073/pnas.1006991107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Watts DP. Post-conflict social events in wild mountain gorillas (Mammalia, Hominoidea). I. Social interactions between opponents. Ethology. 1995;100:139–157. doi: 10.1111/j.1439-0310.1995.tb00321.x. [DOI] [Google Scholar]
- 34.Cordoni G, Palagi E, Borgognini Tarli S. Reconciliation and consolation in captive western gorillas. Int. J. Primatol. 2006;27:1365–1382. doi: 10.1007/s10764-006-9078-4. [DOI] [Google Scholar]
- 35.Mallavarapu S, Stoinski TS, Bloomsmith MA, Maple TL. Postconflict behaviour in captive western lowland gorillas (Gorilla gorilla gorilla) Am. J. Primatol. 2006;68:789–810. doi: 10.1002/ajp.20279. [DOI] [PubMed] [Google Scholar]
- 36.Palagi E, Paoli T, Borgognini Tarli S. Reconciliation and consolation in captive bonobos (Pan paniscus) Am. J. Primatol. 2004;62:15–30. doi: 10.1002/ajp.20000. [DOI] [PubMed] [Google Scholar]
- 37.Clay Z, de Waal FBM. Bonobos respond to distress in others: consolation across the age spectrum. PLoS ONE. 2013;8:e55206. doi: 10.1371/journal.pone.0055206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Clay Z, de Waal FBM. Development of socio-emotional competence in bonobos. Proc. Nat. Acad. Sci. USA. 2013;110:18121–18126. doi: 10.1073/pnas.1316449110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Fujisawa KK, Kutsukake N, Hasegawa T. Peacemaking and consolation in Japanese preschoolers witnessing peer aggression. J. Comp. Physiol. 2006;120:48–57. doi: 10.1037/0735-7036.120.1.48. [DOI] [PubMed] [Google Scholar]
- 40.Baan C, Bergmüller R, Smith DW, Molnar B. Conflict management in free-ranging wolves. Canis lupus. Anim. Behav. 2014;90:327–334. doi: 10.1016/j.anbehav.2014.01.033. [DOI] [Google Scholar]
- 41.Seed AM, Clayton NS, Emery NJ. Postconflict third-party affiliation in rooks. Corvus frugilegus. Curr. Biol. 2007;17:152–158. doi: 10.1016/j.cub.2006.11.025. [DOI] [PubMed] [Google Scholar]
- 42.Cozzi A, Sighieri C, Gazzano A, Nicol CJ, Baragli P. Post-conflict friendly reunion in a permanent group of horses (Equus caballus) Behav. Process. 2010;85:185–190. doi: 10.1016/j.beproc.2010.07.007. [DOI] [PubMed] [Google Scholar]
- 43.Aureli F. Post‐conflict anxiety in nonhuman primates: The mediating role of emotion in conflict. Aggr. Behav. 1997;23(5):315–328. doi: 10.1002/(SICI)1098-2337(1997)23:5<315::AID-AB2>3.0.CO;2-H. [DOI] [Google Scholar]
- 44.Singer T. The neuronal basis and ontogeny of empathy and mind reading: review of literature and implications for future research. Neurosci. Biobehav. Rev. 2006;30:855–863. doi: 10.1016/j.neubiorev.2006.06.011. [DOI] [PubMed] [Google Scholar]
- 45.Palagi E, Dall’Olio S, Demuru E, Stanyon R. Exploring the evolutionary foundations of empathy: consolation in monkeys. Evol. Human Behav. 2014;35(4):341–349. doi: 10.1016/j.evolhumbehav.2014.04.002. [DOI] [Google Scholar]
- 46.Thierry, B., Singh, M. & Kaumanns, W. (eds). Macaque Societies. A Model for the Study of Social Organization. (Edimburgh, UK: Cambridge University Press 2004)
- 47.Thierry, B. Covariation of conflict management patterns across macaque species. In Natural conflict Resolution (eds de Waal, F. B. M. & Aureli, F), pp. 106–128. (Berkeley, USA: University of California Press 2000).
- 48.Kawai M, Ohsawa H, Mori U, Dunbar R. Social organization of gelada baboons: social units and definitions. Primates. 1983;24:13–24. doi: 10.1007/BF02381450. [DOI] [Google Scholar]
- 49.Kawai, M. In Ecological and Sociological Studies of Gelada Baboons. Contributions to Primatology (ed Kawai, M.) 16 (Kodansha Tokyo & Karger Basel, 1979). [PubMed]
- 50.Mori, U. Social structure of gelada baboons. In Ecological and Sociological Studies of Gelada Baboons (ed Kawai, M.) 243–247 (Kodansha Tokyo & Karger Basel, 1979).
- 51.Dunbar RIM. Infant-use by male gelada in agonistic contexts: agonistic buffering, progeny protection or soliciting support? Primates. 1984;25:28–35. doi: 10.1007/BF02382293. [DOI] [Google Scholar]
- 52.Smuts, B. B. Gender, aggression, and influence. Primate Societies 400–412 (1987).
- 53.Dunbar RIM, Dunbar P. Maternal time budgets of gelada baboons. Anim. Behav. 1988;36:970–980. doi: 10.1016/S0003-3472(88)80055-1. [DOI] [Google Scholar]
- 54.Dunbar RIM. Structure of gelada baboon reproductive units. III. The male’s relationship with his females. Anim. Behav. 1983;31:565–575. [Google Scholar]
- 55.Bramblett CA. Coalitions among gelada baboons. Primates. 1970;11:327–333. doi: 10.1007/BF01730637. [DOI] [Google Scholar]
- 56.le Roux A, Beehner JC, Bergman TJ. Female philopatry and dominance patterns in wild geladas. Am. J. Primatol. 2011;73:422–430. doi: 10.1002/ajp.20916. [DOI] [PubMed] [Google Scholar]
- 57.Pallante V, Stanyon R, Palagi E. Agonistic support towards victims buffers aggression in geladas (Theropithecus gelada) Behaviour. 2016;153:1217–1243. doi: 10.1163/1568539X-00003369. [DOI] [Google Scholar]
- 58.Mori A, Belay G, Iwamoto T. Changes in unit structures and infanticide observed in Arsi geladas. Primates. 2003;44:217–223. doi: 10.1007/s10329-002-0013-9. [DOI] [PubMed] [Google Scholar]
- 59.Palagi E, Leone A, Mancini G, Ferrari PF. Contagious yawning in gelada baboons as a possible expression of empathy. Proc. Nat. Acad. Sci. USA. 2009;106:19262–19267. doi: 10.1073/pnas.0910891106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Swedell L. Patterns of reconciliation among captive gelada baboons (Theropithecus gelada): A brief report. Primates. 1997;38(3):325–330. doi: 10.1007/BF02381619. [DOI] [Google Scholar]
- 61.Leone A, Palagi E. Reconciling conflicts in a one-male society: the case of geladas. Primates. 2010;51:203–212. doi: 10.1007/s10329-010-0188-4. [DOI] [PubMed] [Google Scholar]
- 62.Call J, Aureli F, de Waal FBM. Postconflict third party affiliation in stumptailed macaques. Anim. Behav. 2002;63:209–216. doi: 10.1006/anbe.2001.1908. [DOI] [Google Scholar]
- 63.Schino G, Marini C. Self-protective function of post-conflict bystander affiliation in mandrills. PLoS One. 2012;7:e38936. doi: 10.1371/journal.pone.0038936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wittig RM, Boesch C. Receiving post-conflict affiliation from the enemy’s friend reconciles former opponents. PLoS ONE. 2010;5:e13995. doi: 10.1371/journal.pone.0013995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Maestripieri D, Schino G, Aureli F, Troisi A. A modest proposal: displacement activities as an indicator of emotions in primates. Anim. Behav. 1992;44:967–979. doi: 10.1016/S0003-3472(05)80592-5. [DOI] [Google Scholar]
- 66.Aureli F, de Waal FBM. Inhibition in social behaviour in chimpanzees under high density conditions. Am. J. Phys. Anthropol. 1997;41:213–228. doi: 10.1002/(SICI)1098-2345(1997)41:3<213::AID-AJP4>3.0.CO;2-#. [DOI] [PubMed] [Google Scholar]
- 67.Baker KC, Aureli F. Behavioural indicators of anxiety: an empirical test in chimpanzees. Behaviour. 1997;134:1031–1050. doi: 10.1163/156853997X00386. [DOI] [Google Scholar]
- 68.Troisi A. Displacement activities as a behavioural measure of stress in nonhuman primates and human subjects. Stress. 2002;5:47–54. doi: 10.1080/102538902900012378. [DOI] [PubMed] [Google Scholar]
- 69.Palagi E, Norscia I. Scratching around stress: Hierarchy and reconciliation make the difference in wild brown lemurs (Eulemur fulvus) Stress. 2011;14:93–97. doi: 10.3109/10253890.2010.505272. [DOI] [PubMed] [Google Scholar]
- 70.Sclafani V, Norscia I, Antonacci D, Palagi E. Scratching around mating: factors affecting anxiety in wild Lemur catta. Primates. 2012;53:247–254. doi: 10.1007/s10329-012-0294-6. [DOI] [PubMed] [Google Scholar]
- 71.de Waal FBM, Preston SD. Mammalian empathy: behavioural manifestations and neural basis. Nat. Rev. Neurosci. 2017;18:498–509. doi: 10.1038/nrn.2017.72. [DOI] [PubMed] [Google Scholar]
- 72.Fraser ON, Aureli F. Reconciliation, consolation and post-conflict behavioural specificity in chimpanzees. Am. J. Primatol. 2008;70:1114–1123. doi: 10.1002/ajp.20608. [DOI] [PubMed] [Google Scholar]
- 73.de Waal FBM. The integration of dominance and social bonding in primates. Q. Rev. Biol. 1986;61:459–479. doi: 10.1086/415144. [DOI] [PubMed] [Google Scholar]
- 74.Arnold K, Whiten A. Post-conflict behaviour of wild chimpanzees (Pan troglodytes schweinfurthii) in the Budongo Forest, Uganda. Behaviour. 2001;138:649–690. doi: 10.1163/156853901316924520. [DOI] [Google Scholar]
- 75.Cords, M., Aureli, F. Reconciliation and relationship qualities. In Natural Conflict Resolution (ed. Aureli, F., de Waal, F. B. M.) 177–178 (Berkeley University of California Press, 2000).
- 76.Preuschoft S, Wang X, Aureli F, de Waal FBM. Reconciliation in captive chimpanzees: a re-evaluation with controlled methods. Int. J. Primatol. 2002;23:29–50. doi: 10.1023/A:1013293623158. [DOI] [Google Scholar]
- 77.Watts DP. Intracommunity coalitionary killing of an adult male chimpanzee at Ngogo, Kibale National Park, Uganda. Int. J. Primatol. 2004;25:507–521. doi: 10.1023/B:IJOP.0000023573.56625.59. [DOI] [Google Scholar]
- 78.Aureli F, Das M, Verleur D, van Hooff JARAM. Post-conflict social interactions among barbary macaques (Macaca sylvanus) Int. J. Primatol. 1994;15:471–484. doi: 10.1007/BF02696104. [DOI] [Google Scholar]
- 79.Schino G, Geminiani S, Rosati L, Aureli F. Behavioural and emotional response of Japanese macaque (Macaca fuscata) mothers after their offspring receive an aggression. J. Comp. Psychol. 2004;118:340–346. doi: 10.1037/0735-7036.118.3.340. [DOI] [PubMed] [Google Scholar]
- 80.Cooper MA, Bernstein IS. Counter aggression and reconciliation in Assamese macaques (Macaca assamensis) Am. J. Primatol. 2002;56(4):215–30. doi: 10.1002/ajp.1076. [DOI] [PubMed] [Google Scholar]
- 81.Aureli F, Schaik CP. Post-conflict behaviour in long-tailed macaques (Macaca fascicularis) Ethology. 1991;89:89–100. doi: 10.1111/j.1439-0310.1991.tb00296.x. [DOI] [Google Scholar]
- 82.Aureli F, Cozzolino R, Cordischi C, Scucchi S. Kin-oriented redirection among Japanese macaques: an expression of a revenge system? Anim. Behav. 1992;44:283–291. doi: 10.1016/0003-3472(92)90034-7. [DOI] [Google Scholar]
- 83.Otovic P, Partan SR, Bryant JB, Hutchinson E. Let’s call a truce… for now: the silent bared-teeth face expression in mandrills (Mandrillus sphinx) during baseline and post-conflict conditions. Ethology. 2014;120:1118–1127. doi: 10.1111/eth.12285. [DOI] [Google Scholar]
- 84.Thierry, B. Management patterns across macaque species. In Natural Conflict Resolution (ed. Aureli, F., de Waal, F. B. M.) 106–128 (Berkeley University of California Press, 2000).
- 85.de Waal, F. B. M. Good Natured (Harvard University Press, 1996).
- 86.Von Rohr CR, et al. Impartial third-party interventions in captive chimpanzees: a reflection of community concern. PLoS One. 2012;7:e32494. doi: 10.1371/journal.pone.0032494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Matsuda I, et al. Comparisons of intraunit relationships in nonhuman primates living in multilevel social systems. Int. J. Primatol. 2012;33(5):1038–1053. doi: 10.1007/s10764-012-9616-1. [DOI] [Google Scholar]
- 88.Snyder-Mackler N, Alberts SC, Bergman TJ. The socio‐genetics of a complex society: female gelada relatedness patterns mirror association patterns in a multilevel society. Mol. Ecol. 2014;23(24):6179–6191. doi: 10.1111/mec.12987. [DOI] [PubMed] [Google Scholar]
- 89.Kaufman AB, Rosenthal R. Can you believe my eyes? The importance of inter-observer reliability statistics in observations of animal behaviour. Anim. Behav. 2009;78:1487–1491. doi: 10.1016/j.anbehav.2009.09.014. [DOI] [Google Scholar]
- 90.Altmann J. Observational study of behaviour sampling methods. Behaviour. 1974;49:227–265. doi: 10.1163/156853974X00534. [DOI] [PubMed] [Google Scholar]
- 91.Kappeler PM, van Schaik CP. Methodological and evolutionary aspects of reconciliation among primates. Ethology. 1992;92:51–69. doi: 10.1111/j.1439-0310.1992.tb00948.x. [DOI] [Google Scholar]
- 92.de Waal FBM, van Hooff JARAM. Side-directed communication and agonistic interactions in chimpanzees. Behaviour. 1981;77:164–198. doi: 10.1163/156853981X00211. [DOI] [Google Scholar]
- 93.Veenema HC, Das M, Aureli F. Methodological improvements for the study of reconciliation. Behav. Proc. 1994;31:29–37. doi: 10.1016/0376-6357(94)90035-3. [DOI] [PubMed] [Google Scholar]
- 94.Cheney DL, Seyfarth RM, Silk JB. The role of grunts in reconciling opponents and facilitating interactions among adult female baboons. Anim. Behav. 1995;50:249–257. doi: 10.1006/anbe.1995.0237. [DOI] [Google Scholar]
- 95.Palombit RA, Cheney DL, Seyfarth RM. Male grunts as mediators of social interaction with females in wild chacma baboons (Papio cynocephalus ursinus) Behaviour. 1999;136:221–242. doi: 10.1163/156853999501298. [DOI] [Google Scholar]
- 96.Gustison ML, le Roux A, Bergman TJ. Derived vocalisations of geladas (Theropithecus gelada) and the evolution of vocal complexity in primates. Phil. Trans. R. Soc. B. 2012;367:1847–1859. doi: 10.1098/rstb.2011.0218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Bergman TJ. Speech-like vocalized lip-smacking in geladas. Curr. Biol. 2013;23:268–269. doi: 10.1016/j.cub.2013.02.038. [DOI] [PubMed] [Google Scholar]
- 98.Siegel, S. & Castellan, N. J. J. Nonparametric statistics: for the behavioural. Sciences. (New York McGraw-Hill, 1988).
- 99.De Vries H, Stevens JM, Vervaecke H. Measuring and testing the steepness of dominance hierarchies. Anim. Behav. 2006;71:585–592. doi: 10.1016/j.anbehav.2005.05.015. [DOI] [Google Scholar]
- 100.Symonds MR, Moussalli A. A brief guide to model selection, multimodel inference and model averaging in behavioural ecology using Akaike’s information criterion. Behav. Ecol. Sociobiol. 2011;65:13–21. doi: 10.1007/s00265-010-1037-6. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data have been submitted as Supplementary Materials.