

Abstract
Vitiligo is a depigmentation disorder of the skin. It is primarily caused by the destruction of melanocytes or obstruction of the melanin synthesis pathway. Melanin is a type of skin pigment that determines skin color. The seeds of Vernonia anthelmintica (L.) Willd (Kaliziri) are used for treating skin diseases including vitiligo in traditional Uyghur medicine. 1,5-Dicaffeoylquinic acid (1,5-diCQA) is a natural polyphenolic compound widely distributed in plants and extracted from Kaliziri seeds. Therefore, in the present study, the effect of 1,5-diCQA on melanin synthesis in B16 cell was evaluated, and its molecular mechanism was explored. The results indicated that 1,5-diCQA treatment of B16 cells stimulated an increase of intracellular melanin level and tyrosinase (TYR) activity without cytotoxicity. Reverse transcription quantitative polymerase chain reaction results also indicated that 1,5-diCQA may markedly improve the protein expression and RNA transcription of microphthalmia-associated transcription factor (MITF), melanogenic enzyme Tyr, tyrosinase-related protein 1 (TRP 1) and tyrosinase-related protein 2 (TRP 2). Additional results identified that 1,5-diCQA may promote the phosphorylation of p38 mitogen-activated protein kinase (p38 MAPK) and extracellular signal-regulated kinase (ERK) MAPK. Notably, the increased levels of intracellular melanin synthesis and tyrosinase expression induced by 1,5-diCQA treatment were significantly attenuated by the protein kinase A (PKA) inhibitor H-89. Intracellular cyclic adenosine monophosphate (cAMP) concentration and phosphorylation of cAMP-response element binding protein was increased following 1,5-diCQA treatment. These results indicated that 1,5-diCQA stimulated melanogenesis via the MAPK and cAMP/PKA signaling pathways in B16 cells, which has potential therapeutic implications for vitiligo.
Keywords: 1, 5-dicaffeoylquinic acid; vitiligo; melanogenesis; mitogen-activated protein kinase signal pathway
Introduction
Vitiligo is a depigmentation disorder of the skin clinically characterized by the appearance of disfiguring circumscribed skin macules. It is hypothesized that the disease primarily results from the destruction of melanocytes or obstruction of the melanin synthesis pathway (1,2). Melanin is an essential component in the pigmentary system of the skin, which serves a vital role in the prevention from damage by ultraviolet light (3). Melanocytes synthesize melanin via a process termed melanogenesis in the melanosome, a specialized organelle of melanocytes (4). Melanogenesis is a physiological process leading to the production of melanin pigment and a crucial step for the regulation of melanocyte functions, including photoprotection (5). Melanogenesis is regulated by melano-genic enzymes, including tyrosinase (TYR), tyrosinase-related protein 1 (TRP 1) and tyrosinase-related protein 2 (TRP 2) (6). TYR directly mediates the production of melanin through the oxidation of melanogenesis-associated substrates tyrosine (7). Microphthalmia-associated transcription factor (MITF) is a master regulator of the transcription of genes involved in melanin synthesis (8).
Previous studies have identified signaling pathways that mediate melanogenesis A summary obtained from Niu and Aisa (9) is presented in Fig. 1. Among them, the p38 mitogen-activated protein kinase (p38 MAPK) pathway may upregulate melanogenesis by increasing MITF expression (9,10). The extracellular signal-regulated kinase mitogen-activated protein kinase (ERK MAPK)-dependent MITF expression pathway is also involved in melanogenesis (11). In addition, the cyclic adenosine monophosphate (cAMP)/protein kinase A (PKA) signal pathway is one of the primary pathways that may increase the expression of MITF, which is also associated with cAMP responsive element binding (CREB) and CREB binding protein (CBP) (12). Previously, the Wnt/β-catenin signal pathway was identified to serve an important role in melanin synthesis through the nuclear mediator MITF (13).
Figure 1.

Melanogenesis. Obtained from Niu and Aisa (9). TYR, tyrosinase; TRP-1, tyrosinase-related protein 1; TRP-2, tyrosinase-related protein 2; MITF, microphthalmia-associated transcription factor; MAPK, mitogen-activated protein kinase; P38, p38 MAPK; ERK, extracellular signal-regulated kinase; JNK, c-Jun N-terminal kinase; cAMP, intracellular cyclic adenosine monophosphate; Dvl, Dishevelled; Go/Gq, G protein; GSK-3β, glycogen synthase kinase 3β; CRE, cAMP-response element; CREB, cAMP-response element binding protein; CBP, CREB binding protein; TCF-LEF, T cell factor-lymphoid enhancer factor; PKA, protein kinase A; α-MSH, α melanocyte stimulating hormone; PI3K, phosphoinositide 3-kinase; Akt, protein kinase B; NO, nitric oxide; MC1R, melanocortin 1 receptor; SCF, stem cell factor; AC, adenylyl cyclase; c-Kit, proto-oncogene c-Kit.
Vernonia anthelmintica (L.) Willd. (Kaliziri), a member of the Asteraceae family, is an erect forb or shrub that is broadly distributed in subtropical and tropical areas throughout Asia and Africa (14). It has been demonstrated that Kaliziri possesses a number of pharmacological properties, including anti-inflammatory, antibacterial, antioxidant, hypoglycemic and antithrombotic activities (15). The seeds are used in traditional therapy as treatment for skin diseases including vitiligo (16).
1,5-Dicaffeoylquinic acid (1,5-diCQA; Fig. 2) is a class of natural polyphenolic compounds widely distributed in plants (17,18). Previously, it was also identified that 1,5-diCQA exhibits antioxidant signal properties that upregulate glutathione synthesis by stimulating the nuclear factor-like 2 pathway in astrocytes and protects them from cell death (19). 1,5-diCQA also protects primary neurons from amyloid β 1-42-induced apoptosis via the phosphoinositide 3-kinase/protein kinase B signal pathway (20).
Figure 2.
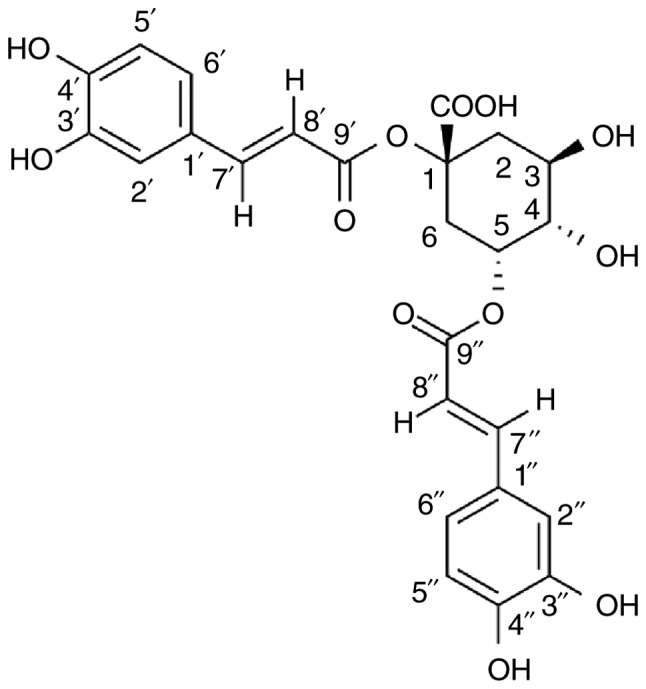
Chemical structure of 1,5-dicaffeoylquinic acid.
Therefore, the present study evaluated the effect of 1,5-diCQA, which was isolated from Kaliziri seeds, on melanin synthesis in murine B16 cells, and examined its molecular mechanism.
Materials and methods
1,5-DiCQA extraction
1,5-DiCQA was extracted from Vernonia anthelmintica (L.) Willd. Briefly, Vernonia anthelmintica seeds (1 kg) were pulverized. Ethanol (40%) was added and refluxed at 80°C three times, each time for 1 h. The extracts were combined and concentrated. This extract was subsequently purified by HPD-300 macroporous resin (diameter, 1:8; loading volume, 6 BV; Cangzhou Bon Adsorber Technology Co., Ltd., Cangzhou, China). Following drying, a 60 g powder was obtained. The powder was taken and dissolved in ethanol, and separated by medium pressure preparative chromatography. At last, the fraction was separated and purified by semi-preparative chromatography to obtain 1,5-diCQA, according to a previously described method (21).
Materials
The obtained 1,5-diCQA (white solid; purity, 98%) was dissolved in dimethyl sulfoxide (DMSO; 100 mM) and stored at −20 °C as a stock solution. ERK (cat. no. 4696), phosphorylated (p)-ERK (Thr202/Tyr204; cat. no. 9106), p38 (cat. no. 9212), p-P38 (Thr180/Tyr182; cat. no. 5140), CREB (cat. no. 9104), p-CREB (Ser133; cat. no. 9196), glycogen synthase kinase 3β (GSK-3β; cat. no. 9832), p-GSK-3β (cat. no. 9323), β-catenin (cat. no. 8480) and p-β-catenin (ser675; cat. no. 4176) antibodies were purchased from Cell Signaling Technology, Inc., (Danvers, MA, USA). p-MITF (cat. no. SAB4301514) antibody, phenyl methyl sulfonyl fluoride and the components of the whole cell lysis buffer for western blot analysis were obtained from Sigma-Aldrich, Merck KGaA (Darmstadt, Germany). β-actin antibody (cat. no. MA1115), rabbit anti-goat secondary antibody (cat. no. BA1060), goat anti-rabbit secondary antibody (cat. no. BA1054) and goat anti-mouse secondary antibody (cat. no. BA1050) were purchased from Wuhan Boster Biological Technology, Ltd., (Wuhan, China). TYR (cat. no. sc-7833), MITF (cat. no. sc-52938), TRP 1 (cat. no. sc-25543) and TRP 2 (cat. no. sc-25544) antibodies were purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). The bicinchoninic acid (BCA) protein assay kit was purchased from Beijing Biomed Gene Technology Co., Ltd. (Beijing, China). The ERK (PD98059), JNK (SP600125) and p38 MAPK inhibitors (SB203580) were purchased from Beyotime Institute of Biotechnology (Haimen, China). The JNK inhibitor (SP600125) was obtained from EMD Biosciences (EMD Millipore, Billerica, MA, USA) and dissolved in DMSO. Deoxynucleotide triphosphates (dNTPs, cat. no. BH7201B) were purchased from TAKARA BIO INC. (Yokohama, Japan). The Recombinant RNasin Ribonuclease Inhibitor (cat. no. N2511) and 100 U Moloney murine leukemia virus (M-MLV) reverse transcriptase (cat. no. M170A) were purchased from Promega Corporation (Madison, WI, USA). Power SYBR™-Green PCR Master mix (cat. no. 4367659) was purchased from Applied Biosystems (Thermo Fisher Scientific, Inc., Waltham, MA, USA). 8-Methoxypsoralen (8-MOP, cat. no. M3501) was purchased from Sigma-Aldrich (Merck KGaA) and dissolved in DMSO (50 mM) for storage at −20°C as a stock solution.
Cell culture
B16 murine melanoma cells were obtained from the Cell Bank of Type Culture Collection of the Chinese Academy of Sciences, (Beijing, China). Cells were grown in Dulbecco's modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10% heat-inactivated fetal bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100 µg/ml streptomycin and 100 U/ml penicillin (Gibco; Thermo Fisher Scientific, Inc.; cat. no. 15140-122) in a humidified atmosphere with 5% CO2 at 37°C.
Cell viability assay
To examine the effects of 1,5-diCQA on cell viability, a TransDetect Cell Counting kit (CCK) assay (cat. no. FC101; Beijing Transgen Biotech, Co., Ltd., Beijing, China) was performed according to the manufacturer's protocol. Briefly, B16 cells were seeded in 96-well plates (5x103 cells/well) and allowed to adhere at 37°C for 12 h. Cells were then treated with 0, 5, 25, 50, 100, 200 or 400 µM 1,5-diCQA for 48 h. Following treatment, 10 µl CCK solution was added into each well and cells were incubated at 37°C for an additional 2 h. Optical absorbance was determined at 450 nm with a Spectra Max M5 plate reader (Molecular Devices, LLC, Sunnydale, CA, USA). The absorbance of cells without treatment was considered as 100% cell survival. Each treatment was performed in quintuplicate, and each experiment was repeated three times. To observe the cell morphology, B16 cells were seeded in 6-well plates (1x104 cells/well) and allowed to adhere at 37°C for 12 h. Cells were subsequently treated with 0, 5, 50 or 100 µM 1,5-diCQA for 48 h. Cell morphology was observed and images were captured at room temperature using a LEICA DMI 8 fluorescence inversion microscope (magnification, x200; Leica Microsystems GmbH, Wetzlar, Germany).
Melanin content assay
The effect of 1,5-diCQA on melanogenesis in B16 cells was investigated according to a previously published protocol, with certain modifications (22). More specifically, intracellular melanin content was measured by using the NaOH dissolution method: B16 cells were seeded in a 6-well plate at a density of 2x105 cells/well, and then allowed to attach for 12 h. Then, cells were treated with 1,5-diCQA at 0, 5, 50, 100 µM or 8-MOP at 50 µM for 48 h, and SB203580 (10 µM); PD98059 (1 µM); SP600125 (10 µM) for 2 h prior to 1,5-diCQA application. Cells were washed with PBS and harvested with 100 µl radioimmunoprecipitation assay lysis buffer (Beyotime Institute of Biotechnology) in 1.5 Eppendorf (Ep) tubes. Samples were centrifuged at −4°C and 12,000 × g for 20 min, and the supernatant was obtained to determine the protein concentrations. Sediment was dissolved in 150 µl 1M NaOH (with 10% DMSO) for 1 h at 80°C, and solubilized melanin was measured at 405 nm. The amount of melanin was calculated by normalizing the total melanin values with the protein content.
Tyrosinase activity assay
Tyrosinase activity was estimated by measuring the rate of L-DOPA oxidation as previously described, with certain modifications (23). B16 cells were seeded in a 6-well plate at a density of 3×105 cells/well and allowed to attach for 12 h. Then, cells were treated with 1,5-diCQA at 0, 5, 50, 100 µM or 8-MOP at 50 µM for 24 h; SB203580 (10 µM); PD98059 (1 µM); or SP600125 (10 µM) for 2 h prior to 1,5-diCQA application. Cells were then washed with ice-cold PBS twice, 100 µl lysis buffer (1% sodium deoxycholate and 1% Triton X-100 in PBS) was added and cells were collected with a cell scraper into an Ep tube. All tubes were incubated at −20°C for 30 min, and then centrifuged at −4°C and 12,000 × g for 20 min. Following centrifugation, the supernatants were obtained for determining the protein concentrations and tyrosinase activity assay. Protein concentrations were determined using the BCA protein assay kit. A reaction mixture containing 90 µl cell lysate and 10 µl 10 mM L-DOPA was added to each well of a 96-well plate for each sample. Following a 20–60-min incubation (according to the content of dopachrome formation) at 37°C in the dark, the dopachrome product was detected at 490 nm by a multi-plate reader (Spectra Max M5/M5e), The tyrosinase activity of each sample was calculated and corrected for the concentrations of proteins, and normalized with protein content levels.
Reverse transcription quantitative polymerase chain reaction (RT-qPCR)
Total cellular RNA was prepared from B16 cells using TRIzol reagent® (Invitrogen; Thermo Fisher Scientific, Inc.) according to the manufacturer's protocol. The quality of RNA samples was assessed using 1.5% agarose gel electrophoresis, at 100 V for 20 min and was subsequently analyzed using Quantity One version 3 (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The concentration of total RNA was determined using a NanoDrop 2000 spectrophotometer (NanoDrop Technologies; Thermo Fisher Scientific, Inc., Wilmington DE, USA). cDNA was synthesized from 1.0 µg total RNA using the Reverse Transcriptase M-MLV according to the protocol of the manufacturer. The cDNA from each sample was used as a template in the RT-qPCR to detect the mRNA level of the 1,5-diCQA target genes. qPCR was conducted as previously described, with certain modifications (24). qPCR was performed in a 7300 Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific, Inc.) in a final volume of 25 µl [forward and reverse primers, 0.25 mol/l each; Power SYBR®-Green PCR Master mix (cat. no. 4367659); and a 1 µl cDNA sample]. The thermoycling conditions were as follows: 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. PCR primers were as follows: TYR forward, 5′GAG AAG CGA GTC TTG ATT AG3′ and reverse, 5′TGG TGC TTC ATG CGC AAA ATC3′; TRP 1 forward, 5′GGC CTC TGA GGT TCT TTA AT3′ and reverse, 5′AAT GAC AAA TTG AGG GTG AG3′; TRP 2 forward, 5′ATG AGA AAC TGC CAA CCT TA3′ and reverse, 5′AGG AGT GAG GCC AAG TTA TGA3′; MITF forward, 5′AGT ACA GGA GCT GGA GAT G3′ and reverse, 5′GTG AGA TCC AGA GTT GTC GT3′ (25); β-actin forward, 5′ATG AGA AGG AGA TCA CTG C3′ and reverse, 5′CTG CGC AAG TTA GGT TTT GT3′ (26). Relative expression was analyzed and determined by 2−ΔΔCq method, normalizing the data to β-actin mRNA levels (27,28). Experiments were repeated in triplicate.
Western blot analysis
Protein samples that were collected during the melanin content assay was used for western blot analysis. The lysates were denatured in SDS-PAGE protein loading buffer 5X (cat. no. AR1112; Wuhan Boster Biological Technology, Ltd.) separated on 10% SDS-PAGE at 80 V, and transferred onto polyvinylidene fluoride membranes for 2 h at 400 A. Membrane blocking was performed with 5% skim milk dissolved in TBS with 1% Tween-20 (TBST) at room temperature for 1 h and the membrane was incubated with primary antibodies at dilutions of 1:1,000 at 4°C overnight. Subsequent to washing in TBST, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies at a dilution of 1:2,000 for 1 h at room temperature. The membranes were then washed with TBST. Proteins were visualized by enhanced chemiluminescent western blotting detection reagents (GE Healthcare, Chicago, IL, USA). Densitometry analysis was performed using Quantity One version 3 (Bio-Rad Laboratories, Inc.).
Determination of intracellular cAMP levels
B16 melanoma cells were treated with 1,5-diCQA at 0, 5, 50 or 100 µM at 37°C for 12 h. Intracellular cAMP levels were measured using a cAMP ELISA kit (cat. no. STA-500; Cell Biolabs, Inc., San Diego, CA, USA) following the manufacturer's protocol.
Statistical analysis
All data are expressed as mean ± standard deviation. Statistical analysis was performed by one-way analysis of variance followed by Tukey's post-hoc test for multiple comparisons using SPSS 19.0 (IBM Corp., Armonk, NY, USA) and Graphpad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was considered to indicate a statistically significant difference.
Results
Effect of 1,5-diCQA on B16 cell viability
The total melanin content in the skin is determined by the number of melanocytes and the amount of melanin synthesized by single cells (29). In order to avoid the possibility that effect of melanin synthesis was due to the cell viability, the cytotoxicity of 1,5-diCQA to B16 cells was first determined. B16 cells treated at various concentrations of 1,5-diCQA were incubated for 48 h and compared with the 1,5-diCQA untreated cells. Cell viability was measured using a CCK assay. The results indicated that 1,5-diCQA exhibited no cytotoxicity at concentration ranges of 0–400 µM (Fig. 3A). Concurrently, 1,5-diCQA did not induce any change in cell morphology when compared with the control cells at the concentration of 0–100 µM (Fig. 3B). Accordingly, it was determined that 1,5-diCQA is not cytotoxic to melanoma cells.
Figure 3.

Effects of 1,5-diCQA on B16 cell viability and morphology. (A) B16 cells were treated with 0.1% dimethyl sulfoxide as a vehicle control, or with 1,5-diCQA at 5, 25, 50, 100, 200 and 400 µM for 48 h and cell viability was measured by CCK assay. (B) Cell morphology was observed under a microscope at magnification, ×200. (1) control; (2) 5; (3) 50; and (4) 100 µM 1,5-diCQA. Values are expressed as the mean ± standard deviation of three separate independent experiments. 1,5-diCQA, 1,5-dicaffeoylquinic acid.
Effect of 1,5-diCQA on melanin formation and Tyr activity
The effect of 1,5-diCQA on melanin production in B16 cells was determined by a melanin content assay. As indicated in Fig. 4A and B, intracellular melanin levels increased in response to 1,5-diCQA treatment in a dose- and time-dependent manner, which suggests that 1,5-diCQA may promote melanin synthesis in B16 cells. Fig. 4C denotes the cell precipitation following centrifugation and visual observation. It indicates that cells became darker following the addition of 1,5-diCQA. Studies investigating pigmentary disorders have primarily focused on tyrosinase activity, as it is the most important enzyme in melanin biosynthesis in the melanocytes (30). The results also suggested that 1,5-diCQA promotes intracellular Tyr activity in B16 cells in a dose-dependent manner after 24 h of treatment (Fig. 4D).
Figure 4.

Effects of 1,5-diCQA on the melanin contents in B16 cells. Melanin content was measured in dose- and time-dependent manners. (A) B16 cells were treated with 0.1% dimethyl sulfoxide as a blank control, 8-MOP (50 µM) as a positive control or 1,5-diCQA at 5, 50 and 100 µM for 48 h. (B) Melanin content was assayed in a time-dependent manner following treatment with 1,5-diCQA at 50 µM. (C) B16 cells were treated with 1,5-diCQA at 5, 50 and 100 µM for 48 h, and cell precipitation following centrifugation is indicated. (D) B16 cells were treated with 1,5-diCQA at 5, 50 and 100 µM for 24 h, and TYR activity was measured. (E) B16 cells were treated with 1,5-diCQA at 5, 50 and 100 µM for 48 h. TYR, TRP1, TRP2 and MITF protein expression levels were detected by western blot analysis, and β-actin was used as a loading control. (F) B16 cells were treated with 100 µM 1,5-diCQA for 0,12, 24, 36 and 48 h. TYR, TRP1, TRP2 and MITF protein expression levels were detected by western blot analysis, and β-actin was used as a loading control. Values are expressed as the mean ± standard deviation of three separate independent experiments. Each percentage value in the treated cells was calculated with respect to that in the control cells. *P<0.05, **P<0.01 and ***P<0.001 vs. control group. TYR, tyrosinase; TRP-1, tyrosinase-related protein 1; TRP-2, tyrosinase-related protein 2; MITF, microphthalmia-associated transcription factor; 1,5-diCQA, 1,5-dicaffeoylquinic acid.
Effects of 1,5-diCQA on the expression of melanogenesis-associated genes
The effect of 1,5-diCQA on the expression of melanogenic genes was additionally explored by western blot analysis and RT-qPCR. Western blot analysis indicated that 1,5-diCQA significantly increased the protein expression levels of melanogenic genes TYR, TRP 1, TRP 2 and transcription factor MITF in a dose- and time-dependent manner (Fig. 4E and F). In order to confirm whether the changes in these protein expressions were due to the changes in RNA levels, the effects of 1,5-diCQA on mRNA levels was also detected, and RNA levels of MITF and its downstream genes TYR, TRP 1, and TRP 2. The results demonstrated that the transcriptional levels of MITF, TYR, TRP 1, and TRP 2 were significantly increased in B16 cells in the presence of 1,5-diCQA in a dose-dependent manner (Fig. 5).
Figure 5.

Effects of 1,5-diCQA on the transcriptional TYR family and MITF. B16 cells were treated with 1,5-diCQA at 5, 50 and 100 µM for 24 h. The mRNA levels of TYR family and MITF were detected using reverse transcription quantitative polymerase chain reaction. **P<0.01 and ***P<0.001 vs. untreated control. TYR, tyrosinase; MITF, microphthalmia-associated transcription factor; 1,5-diCQA, 1,5-dicaffeoylquinic acid.
Effects of 1,5-diCQA on the MAPK and Wnt signaling pathways
It has been demonstrated that the phosphorylation of MAPK and signaling cascades of ERK, JNK and p38 regulate melanin production (31). As the results of the present study indicated that 1,5-diCQA increased melanin production via the induction of the melanogenic enzyme and pigmentation-associated transcription factor MITF, the melanin-associated signal pathways involved were additionally explored. To investigate the involvement of the MAPK signal pathway in 1,5-diCQA-promoted melanin synthesis, western blot analysis was performed on B16 cells following 1,5-diCQA treatment. The results demonstrated that the phosphorylation levels of p38 and ERK, but not JNK, were significantly increased following 30 min treatment with different concentrations of 1,5-diCQA (Fig. 6). These data indicated that 1,5-diCQA may increase melanin synthesis by increasing the levels of p38 and ERK phosphorylation. Therefore, inhibitors of p38 (SB203580), ERK (PD98059) and JNK (SP600125) were applied to verify our prior hypothesis. B16 cells were pre-treated with different inhibitors for 2 h prior to the addition of 1,5-diCQA (100 µM). Then, melanin content and Tyr activities were measured. The results indicated that SB203580 and PD98059 may reverse the 1,5-diCQA effects on melanin content and TYR activities (Fig. 7A–D). However, no effects from the JNK inhibitor (SP600125) were observed (Fig. 7E and F), which was consistent with the western blotting results. In addition, MITF expression was measured by western blot analysis following treatment with SB203580, PD98059, SP600125 and 1,5-diCQA (100 µM) for 48 h (Fig. 8). The results also indicate that 1,5-diCQA increased melanin content through the phosphorylation of p38 and ERK, but not JNK.
Figure 6.
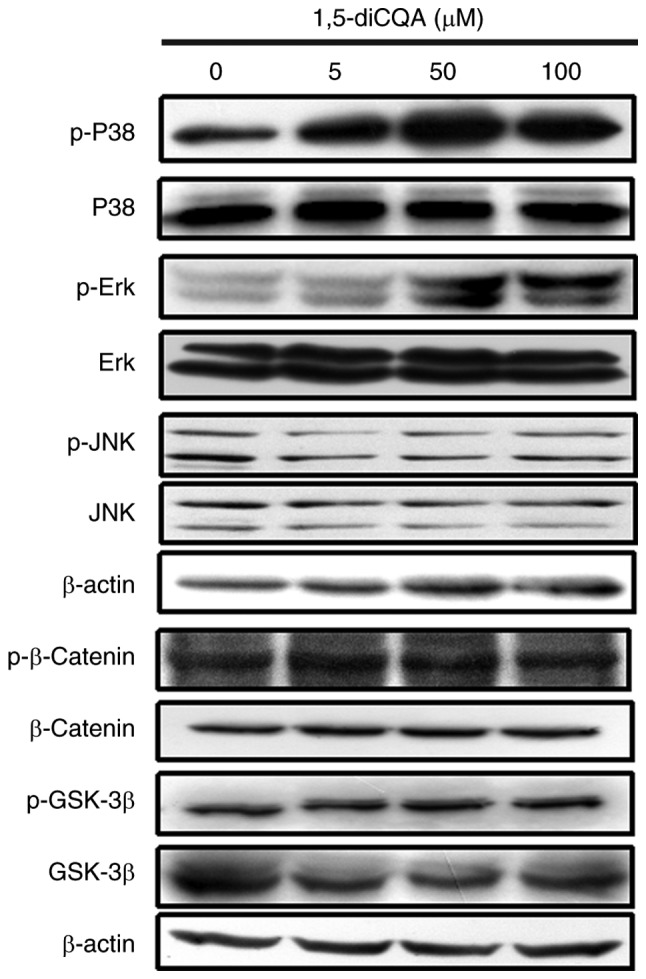
Effects of 1,5-diCQA on the MAPK and Wnt signal pathways. B16 cells were treated with 5, 50 and 100 µM 1,5-diCQA for 30 min, and total and phosphorylated forms of p38, ERK and JNK was measured by western blot analysis. Levels of total and phosphorylated forms of β-catenin and GSK-3β were measured following treatment with 1,5-diCQA for 48 h. p, phos-phorylated; MAPK, mitogen-activated protein kinase; P38, p38 MAPK; ERK, extracellular signal-regulated kinase; JNK, c-Jun N-terminal kinase; GSK-3β, glycogen synthase kinase 3β; 1,5-diCQA, 1,5-dicaffeoylquinic acid.
Figure 7.

Effects of p38, ERK and JNK inhibitors on 1,5-diCQA-induced melanin synthesis. B16 cells were pre-incubated with SB203580 (10 µM) for 2 h prior to the addition of 1,5-diCQA (100 µM), and then (A) incubated for 48 h for the measurement of melanin content or (B) incubated for 24 h for the measurement of TYR activity. (C) B16 cells were pre-incubated with PD98059 (10 µM) for 2 h prior to the addition of 1,5-diCQA (100 µM), and then (C) incubated for 48 h for the measurement of melanin content or (D) incubated for 24 h for the measurement of TYR activity. B16 cells were pre-incubated with SP600125 (10 µM) for 2 h prior to the addition of 1,5-diCQA (100 µM), and then (E) incubated for 48 h for the measurement of melanin content or (F) incubated for 24 h for the measurement of TYR activity. Each percentage value in the treated cells was calculated with respect to that in the control cells. Values are expressed as the mean ± standard deviation of three separate independent experiments. **P<0.01 and ***P<0.001 vs. untreated control group. ##P<0.01 and ###P<0.001 vs. single drug treatment group. P38, p38 mitogen-activated protein kinase; ERK, extracellular signal-regulated kinase; JNK, c-Jun N-terminal kinase; 1,5-diCQA, 1,5-dicaffeoylquinic acid.
Figure 8.
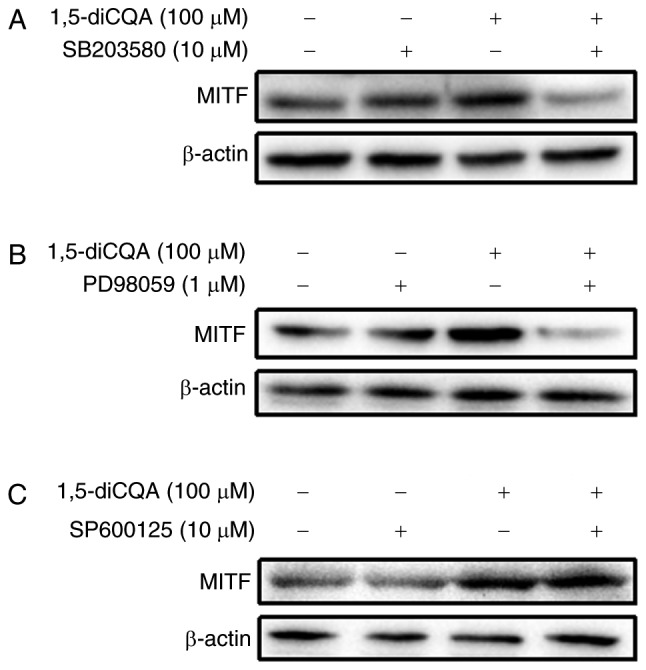
Effects of p38 mitogen-activated protein kinase, extracellular signal-regulated kinase and c-Jun N-terminal kinase inhibitors on 1,5-diCQA-induced MITF expression. B16 cells were pre-incubated with (A) SB203580 (10 µM), (B) PD98059 (1 µM) or (C) SP600125 (10 µM) for 2 h prior to the addition of 1,5-diCQA (100 µM), and then incubated for 48 h. MITF expression was measured by western blot analysis.
The Wnt signal pathway was demonstrated to be closely associated with melanin synthesis (32). In order to investigate whether the Wnt signal pathway was involved in the effects of 1,5-diCQA on melanogenesis, the changes in β-catenin and GSK-3β in B16 cells following treatment with 1,5-diCQA were measured by western blot analysis. B16 cells were treated with 5, 50 and 100 µM 1,5-diCQA for 48 h, and total and phosphorylated forms of β-catenin and GSK-3β were measured. The results indicated that neither total nor phosphorylated β-catenin and GSK-3β were altered following 1,5-diCQA treatment (Fig. 6).
Effects of 1,5-diCQA on the PKA signaling pathway
It is well-known that the PKA signaling pathway is involved in melanogenesis. Increased cellular cAMP levels may activate PKA. In turn, activated PKA may activate CREB, leading to the upregulation of MITF transcription (33). Therefore, to determine whether the effects of 1,5-diCQA on melanogenesis were also mediated by the PKA signal pathway, a range of experiments using the PKA inhibitor H89 were conducted to identify if PKA affected this process. B16 cells were pre-incubated with H89 (10 µM) for 2 h prior to the addition of 1,5-diCQA (100 µM), and then incubated for 48 h for the measurement of melanin content and expression of MITF, or incubated for 24 h for the measurement of TYR activity. The results indicated that the effects of 1,5-diCQA on melanogenesis were inhibited following pre-incubation with H89 (Fig. 9A and B). MITF expression was also decreased in the samples that were treated with H89 (10 µM) and 1,5-diCQA (100 µM) (Fig. 9F). In addition, cAMP content of the B16 cells following treatment with 1,5-diCQA was measured. The results confirmed that cAMP content was also significantly increased in response to 1,5-diCQA treatment in a dose-dependent manner (Fig. 9C). The levels of CREB induced by PKA, which may activate MITF transcription levels, were also investigated using western blot analysis. Phosphorylation of CREB was significantly increased following treatment with 1,5-diCQA in time- and dose-dependent manners. (Fig. 9D and E).
Figure 9.

Effects of 1,5-diCQA on the PKA signal pathway. B16 cells were pre-incubated with H89 (10 µM) for 2 h prior to the addition of 1, 5-diCQA (100 µM), and then (A) incubated for 48 h for the measurement of melanin content or (B) incubated for 24 h for the measurement of TYR activity. (C) B16 cells were treated with 0.1% dimethyl sulfoxide and 8-MOP as positive controls or 1,5-diCQA at 5, 50 and 100 µM for 12 h, and then cAMP content was measured by a cAMP-ELISA kit. (D) B16 cells were treated with 5, 50 and 100 µM 1,5-diCQA for 48 h, and levels of total and phosphorylated CREB were measured by western blot analysis. (E) B16 cells were treated with 1,5-diCQA (100 µM) for 0, 12, 24, 36 and 48 h, and levels of total and phosphorylated CREB were measured by western blot analysis. (F) B16 cells were pre-incubated with H89 (10 µM) for 2 h prior to the addition of 1,5-diCQA (100 µM), and then incubated for 48 h. MITF expression levels were the measured by western blot analysis. **P<0.01 and ***P<0.001 vs. untreated control group. ##P<0.01 and ###P<0.001 vs. single treatment group. PKA, protein kinase A; TYR, tyrosinase; MITF, microphthalmia-associated transcription factor; 1,5-diCQA, 1,5-dicaffeoylquinic acid; p-phosphorylated; CREB, cAMP-response element binding protein.
Discussion
Melanin serves a pivotal role in solar UV irradiation-induced skin injury. Lack of melanin is involved in the development of skin diseases, including vitiligo and albinism (34). At present, a number of studies have focused on the specific mechanisms, including the induction of melanin biosynthesis and functional melanocytes, with the aim of developing novel therapeutic agents for vitiligo (35–37). Kaliziri is a plant that only grows in high-altitude areas of southern Xinjiang (China) and small regions of Pakistan and India. The seeds of Kaliziri have historically been used for treating skin diseases including vitiligo in Traditional Chinese medicine (16). Our study group have focused on identifying novel compounds in Kaliziri, and have isolated 1,5-diCQA using high performance liquid chromatography (HPLC) and preparative HLPC, which may be the active ingredient (38,39). Previous studies suggested that methyl 3,5-dicaffeoylquinate and 3,5-dicaffeoylquinic acid, two different structural analogues of 1,5-diCQA, induce melanin synthesis (40,41). Therefore, we hypothesized that 1,5-diCQA may have effects on melanin synthesis. In the present study, the effects of 1,5-diCQA on skin pigmentation induction and its underlying mechanism were investigated. The results indicated that 1,5-diCQA increased melanin production by the induction of the pigmentation-associated transcription factor MITF and melanogenic enzymes TYR, TRP 1 and TRP 2. Then, the present study attempted to elucidate the molecular mechanism of melanogenesis induction by 1,5-diCQA.
There are certain melanin-associated signal pathways involved in this process, through affecting the pigmentation-associated transcription factor MITF (42). It was demonstrated that p38 MAPK activation contributes to melanin production by activating CREB, which in turn activates MITF expression (43). In addition, the ERK MAPK pathway is also involved in melanogenesis (44). Therefore, the present study examined the effect of 1,5-diCQA on the MAPK signal pathway. It was identified that 1,5-diCQA upregulated the phosphorylation levels of ERK MAPK and p38 MAPK, while levels of JNK MAPK phosphorylation remained the same. This suggested that the p38 MAPK and ERK MAPK pathways may contribute to the stimulation of melanin production.
1,5-DiCQA is a structural analogs of isochlorogenic acid A, which stimulates melanin synthesis via the Wnt signaling pathway (41). Therefore, the present study examined the effect of 1,5-diCQA on the Wnt signal pathway. However, the levels of total and phosphorylated GSK-3β and β-catenin, which are key proteins in the Wnt signal pathway (13), were demonstrated to be similar prior and subsequent to 1,5-diCQA treatment. This indicates that the Wnt signaling pathway is not a major factor of 1,5-diCQA-induced melanin synthesis.
It has been demonstrated that cAMP up-regulation activates MAPK in B16 cells and normal human melanocytes (45). The cAMP signal pathway is modulated via the MAPK and PKA pathways during melanin synthesis (46). Once intracellular cAMP is accumulated, it activates PKA that subsequently phosphorylates CREB. Then CREB binds to the CRE motif of the MITF promoter and activates MITF transcription (12). Therefore, the effects of 1,5-diCQA on the PKA signal pathway was additionally examined. The results indicated that 1,5-diCQA effects may activate the PKA signaling pathway through the accumulation of cAMP content. In turn, activated PKA activates CREB, sequentially leading to the activation of MITF transcription. Eventually, MITF upregulates melanogenic genes, including TYR, TRP 1 and TRP 2 (Fig. 10).
Figure 10.
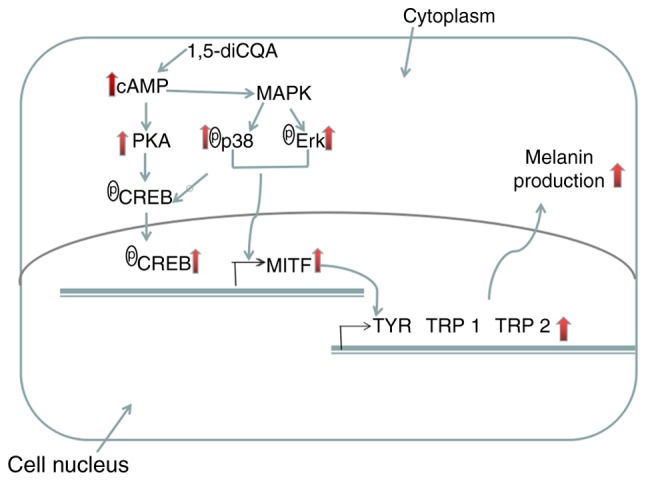
Potential mechanisms through which 1,5-diCQA functions in promoting melanin content in B16 cells. 1,5-diCQA increases the levels of p38 MAPK and ERK MAPK phosphorylation, and therefore increases the activation of MITF. 1,5-diCQA increase the cAMP content, which activates PKA that subsequently phosphorylates CREB. CREB binds to the CRE motif of the MITF promoter and activates MITF transcription TYR, tyrosinase; TRP-1, tyrosinase-related protein 1; TRP-2, tyrosinase-related protein 2; MITF, microphthalmia-associated transcription factor; 1,5-diCQA, 1,5-dicaffeoylquinic acid; MAPK, mitogen-activated protein kinase; P38, p38 MAPK; ERK, extracellular signal-regulated kinase; cAMP, intracellular cyclic adenosine monophosphate; CREB, cAMP-response element binding protein; PKA, protein kinase A.
In conclusion, the present study suggested that the 1,5-diCQA purified from Kaliziri promoted melanogenesis in B16 cells by activating the p38 MAPK, ERK MAPK and PKA signaling pathways. Taken together, the present study suggested that 1,5-diCQA may be a useful agent for treating hypopigmentation skin disorders.
Acknowledgments
Not applicable.
Funding
The present study was supported by grants from the Key Research and Development Project of Xinjiang Autonomous Region (grant no. 2016B03038-3) and Projects of International Science and Technology Cooperation of the Xinjiang Uyghur Autonomous Region (grant no. 20146020).
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Authors' contributions
HAA and NM conceived and designed the experiments and wrote the paper; NM and XYL performed the experiments; MK analyzed the data; HAA revised the paper.
Ethics approval and consent to participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
References
- 1.Boniface K, Seneschal J, Picardo M, Taïeb A. Vitiligo: Focus on clinical aspects, immunopathogenesis, and therapy. Clin Rev Allergy Immunol. 2018;54:52–67. doi: 10.1007/s12016-017-8622-7. [DOI] [PubMed] [Google Scholar]
- 2.Schallreuter KU, Bahadoran P, Picardo M, Slominski A, Elassiuty YE, Kemp EH, Giachino C, Liu JB, Luiten RM, Lambe T, et al. Vitiligo pathogenesis: Autoimmune disease, genetic defect, excessive reactive oxygen species, calcium imbalance, or what else? Exp Dermatol. 2008;17:139–140. doi: 10.1111/j.1600-0625.2007.00666.x. [DOI] [PubMed] [Google Scholar]
- 3.Gupta AK, Gover MD, Nouri K, Taylor S. The treatment of melasma: A review of clinical trials. J Am Acad Dermatol. 2006;55:1048–1065. doi: 10.1016/j.jaad.2006.02.009. [DOI] [PubMed] [Google Scholar]
- 4.Yamaguchi Y, Hearing VJ. Melanocytes and their diseases. Cold Spring Harb Perspect Med. 2014;4:a017046. doi: 10.1101/cshperspect.a017046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pillaiyar T, Manickam M, Jung SH. Recent development of signaling pathways inhibitors of melanogenesis. Cell Signal. 2017;40:99–115. doi: 10.1016/j.cellsig.2017.09.004. [DOI] [PubMed] [Google Scholar]
- 6.Rad HH, Yamashita T, Jin HY, Hirosaki K, Wakamatsu K, Ito S, Jimbow K. Tyrosinase-related proteins suppress tyrosinase-mediated cell death of melanocytes and melanoma cells. Exp Cell Res. 2004;298:317–328. doi: 10.1016/j.yexcr.2004.04.045. [DOI] [PubMed] [Google Scholar]
- 7.Kim YJ, Uyama H. Tyrosinase inhibitors from natural and synthetic sources: Structure, inhibition mechanism and perspective for the future. Cell Mol Life Sci. 2005;62:1707–1723. doi: 10.1007/s00018-005-5054-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Levy C, Khaled M, Fisher DE. MITF: Master regulator of melanocyte development and melanoma oncogene. Trends Mol Med. 2006;12:406–414. doi: 10.1016/j.molmed.2006.07.008. [DOI] [PubMed] [Google Scholar]
- 9.Niu C, Aisa HA. Upregulation of melanogenesis and tyrosinase activity: Potential agents for vitiligo. Molecules. 2017;22:E1303. doi: 10.3390/molecules22081303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yamaguchi Y, Brenner M, Hearing VJ. The regulation of skin pigmentation. J Biol Chem. 2007;282:27557–27561. doi: 10.1074/jbc.R700026200. [DOI] [PubMed] [Google Scholar]
- 11.Gu WJ, Ma HJ, Zhao G, Yuan XY, Zhang P, Liu W, Ma LJ, Lei XB. Additive effect of heat on the UVB-induced tyrosinase activation and melanogenesis via ERK/p38/MITF pathway in human epidermal melanocytes. Arch Dermatol Res. 2014;306:583–590. doi: 10.1007/s00403-014-1461-y. [DOI] [PubMed] [Google Scholar]
- 12.Buscà R, Ballotti R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment Cell Res. 2000;13:60–69. doi: 10.1034/j.1600-0749.2000.130203.x. [DOI] [PubMed] [Google Scholar]
- 13.Schepsky A, Bruser K, Gunnarsson GJ, Goodall J, Hallsson JH, Goding CR, Steingrimsson E, Hecht A. The microph-thalmia-associated transcription factor Mitf interacts with beta-catenin to determine target gene expression. Mol Cell Biol. 2006;26:8914–8927. doi: 10.1128/MCB.02299-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lin R, Chen YL. Flora of China. Vol. 4. Science Press; Beijing: 1985. Compositae; pp. 5–8. [Google Scholar]
- 15.Jamil S, Khan RA, Ahmed S, Fatima S. Evaluation of anti-inflammatory and anti-oxidant potential of seed extracts o Vernonia anthelmintica. Pak J Pharm Sci. 2017;30:755–760. [PubMed] [Google Scholar]
- 16.Tuerxuntayi A, Liu YQ, Tulake A, Kabas M, Eblimit A, Aisa HA. Kaliziri extract upregulates tyrosinase, TRP-1, TRP-2 and MITF expression in murine B16 melanoma cells. BMC Complement Altern Med. 2014;14:166. doi: 10.1186/1472-6882-14-166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Yang B, Meng Z, Dong J, Yan L, Zou L, Tang Z, Dou G. Metabolic profile of 1,5-dicaffeoylquinic acid in rats, an in vivo and in vitro study. Drug Metab Dispos. 2005;33:930–936. doi: 10.1124/dmd.104.002154. [DOI] [PubMed] [Google Scholar]
- 18.McDougall B, King PJ, Wu BW, Hostomsky Z, Reinecke MG, Robinson WE., Jr Dicaffeoylquinic and dicaffeoyltartaric acids are selective inhibitors of human immunodeficiency virus type 1 integrase. Antimicrob Agents Chemother. 1998;42:140–146. doi: 10.1128/AAC.42.1.140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Cao X, Xiao H, Zhang Y, Zou L, Chu Y, Chu X. 1-5-Dicaffeoylquinic acid-mediated glutathione synthesis through activation of Nrf2 protects against OGD/reper-fusion-induced oxidative stress in astrocytes. Brain Res. 2010;1347:142–148. doi: 10.1016/j.brainres.2010.05.072. [DOI] [PubMed] [Google Scholar]
- 20.Xiao HB, Cao X, Wang L, Run XQ, Su Y, Tian C, Sun SG, Liang ZH. 1-5-dicaffeoylquinic acid protects primary neurons from amyloid β 1-42-induced apoptosis via PI3K/Akt signaling pathway. Chin Med J (Engl) 2011;124:2628–2635. [PubMed] [Google Scholar]
- 21.Zheng Z, Wang X, Liu P, Li M, Dong H, Qiao X. Semi-preparative separation of 10 caffeoylquinic acid derivatives using high speed counter-current chromatogaphy combined with semi-preparative HPLC from the roots of burdock (Arctium lappa L.) Molecules. 2018;23:E429. doi: 10.3390/molecules23020429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Park SY, Jin ML, Kim YH, Kim Y, Lee SJ. Aromatic-turmerone inhibits α-MSH and IBMX-induced melanogenesis by inactivating CREB and MITF signaling pathways. Arch Dermatol Res. 2011;303:737–744. doi: 10.1007/s00403-011-1155-7. [DOI] [PubMed] [Google Scholar]
- 23.Zhou J, Ren T1, Li Y, Cheng A, Xie W, Xu L, Peng L, Lin J, Lian L, Diao Y, et al. Oleoylethanolamide inhibits α-melanocyte stimulating hormone-stimulated melanogenesis via ERK, Akt and CREB signaling pathways in B16 melanoma cells. Oncotarget. 2017;8:56868–56879. doi: 10.18632/oncotarget.18097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hout DR, Schweitzer BL, Lawrence K, Morris SW, Tucker T, Mazzola R, Skelton R, McMahon F, Handshoe J, Lesperance M, et al. Performance of a RT-PCR assay in comparison to fish and immunohistochemistry for the detection of ALK in non-small cell lung cancer. Cancers (Basel) 2017;9:E99. doi: 10.3390/cancers9080099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Zhu PY, Yin WH, Wang MR, Dang YY, Ye XY. Andrographolide suppresses melanin synthesis through Akt/GSK3β/β-catenin signal pathway. J Dermatol Sci. 2015;79:74–83. doi: 10.1016/j.jdermsci.2015.03.013. [DOI] [PubMed] [Google Scholar]
- 26.Zheng MF, Shen SY, Huang WD. DCA increases the antitumor effects of capecitabine in a mouse B16 melanoma allograft and a human non-small cell lung cancer A549 xenograft. Cancer Chemother Pharmacol. 2013;72:1031–1041. doi: 10.1007/s00280-013-2281-z. [DOI] [PubMed] [Google Scholar]
- 27.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–428. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 28.Lu XY, Li JQ, Liu XN, Li XB, Ma J. Characterization and expression analysis of six chitinase genes from the desert beetle microdera punctipennis in response to low temperature. Cryo Letters. 2014;35:438–448. [PubMed] [Google Scholar]
- 29.Smit NP, Kolb RM, Lentjes EG, Noz KC, van der Meulen H, Koerten HK, Vermeer BJ, Pavel S. Variations in melanin formation by cultured melanocytes from different skin types. Arch Dermatol Res. 1998;290:342–349. doi: 10.1007/s004030050315. [DOI] [PubMed] [Google Scholar]
- 30.Ramsden CA, Riley PA. Tyrosinase: The four oxidation states of the active site and their relevance to enzymatic activation, oxidation and inactivation. Bioorg Med Chem. 2014;22:2388–2395. doi: 10.1016/j.bmc.2014.02.048. [DOI] [PubMed] [Google Scholar]
- 31.Shen T, Heo SI, Wang MH. Involvement of the p38 MAPK and ERK signaling pathway in the anti-melanogenic effect of methyl 3,5-dicaffeoyl quinate in B16F10 mouse melanoma cells. Chem Biol Interact. 2012;199:106–111. doi: 10.1016/j.cbi.2012.06.004. [DOI] [PubMed] [Google Scholar]
- 32.Hart MJ, de los Santos R, Albert IN, Rubinfeld B, Polakis P. Downregulation of beta-catenin by human axin and its association with the APC tumor suppressor, beta-catenin and GSK3 beta. Curr Biol. 1998;8:573–581. doi: 10.1016/S0960-9822(98)70226-X. [DOI] [PubMed] [Google Scholar]
- 33.Kim DS, Cha SB, Park MC, Park SA, Kim HS, Woo WH, Mun YJ. Scopoletin stimulates melanogenesis via cAMP/PKA pathway and partially p38 activation. Biol Pharm Bull. 2017;14:2068–2074. doi: 10.1248/bpb.b16-00690. [DOI] [PubMed] [Google Scholar]
- 34.Kaidbey KH, Agin PP, Sayre RM, Kligman AM. Kligman, hotoprotection by melanin-a comparison of black and caucasian skin. J Am Acad Dermatol. 1979;1:249–260. doi: 10.1016/S0190-9622(79)70018-1. [DOI] [PubMed] [Google Scholar]
- 35.Sato K, Ando R, Kobayashi H, Nishio T. 2-Ethoxybenzamide stimulates melanin synthesis in B16F1 melanoma cells via the CREB signaling pathway. Mol Cell Biochem. 2016;423:39–52. doi: 10.1007/s11010-016-2823-x. [DOI] [PubMed] [Google Scholar]
- 36.Lee KM, Lee KY, Choi HW, Cho MY, Kwon TH, Kawabata S, Lee BL. Activated phenoloxidase from Tenebrio molitor larvae enhances the synthesis of melanin by using a vitellogenin-like protein in the presence of dopamine. Eur J Biochem. 2000;267:3695–3703. doi: 10.1046/j.1432-1327.2000.01402.x. [DOI] [PubMed] [Google Scholar]
- 37.Kumar R, Parsad D, Rani S, Bhardwaj S, Srivastav N. Glabrous lesional stem cells differentiated into functional mela-nocytes: New hope for repigmentation. J Eur Acad Dermatol Venereol. 2016;30:1555–1560. doi: 10.1111/jdv.13686. [DOI] [PubMed] [Google Scholar]
- 38.Maimaiti Z, Turak A, Aisa HA. Two new compounds from the seeds o Vernonia anthelmintica. J Asian Nat Prod Res. 2017;19:862–868. doi: 10.1080/10286020.2016.1269760. [DOI] [PubMed] [Google Scholar]
- 39.Turak A, Aisa HA. Three new elemanolides from the seeds o Vernonia anthelmintica. J Asian Nat Prod Res. 2018;20:313–320. doi: 10.1080/10286020.2017.1343304. [DOI] [PubMed] [Google Scholar]
- 40.Kim HJ, Kim JS, Woo JT, Lee IS, Cha BY. Hyperpigmentation mechanism of methyl 3,5-dicaffeoylquinate through activation of p38 andMITF induction of tyrosinase. Acta Biochim Biophys Sin (Shanghai) 2015;47:548–556. doi: 10.1093/abbs/gmv040. [DOI] [PubMed] [Google Scholar]
- 41.Mamat N, Dou J, Lu X, Eblimit A, Haji Akber A. Isochlorogenic acid A promotes melanin synthesis in B16 cell through the β-catenin signal pathway. Acta Biochim Biophys Sin (Shanghai) 2017;49:800–807. doi: 10.1093/abbs/gmx072. [DOI] [PubMed] [Google Scholar]
- 42.Pei T, Zheng C, Huang C, Chen X, Guo Z, Fu Y, Liu J, Wang Y. Systematic understanding the mechanisms of vitiligo pathogenesis and its treatment by Qubaibabuqi formula. J Ethnopharmacol. 2016;190:272–287. doi: 10.1016/j.jep.2016.06.001. [DOI] [PubMed] [Google Scholar]
- 43.Ahn JH, Jin SH, Kang HY. LPS induces melanogenesis through p38 MAPK activation in human melanocytes. Arch Dermatol Res. 2008;300:325–329. doi: 10.1007/s00403-008-0863-0. [DOI] [PubMed] [Google Scholar]
- 44.Yanase H, Ando H, Horikawa M, Watanabe M, Mori T, Matsuda N. Possible involvement of ERK 1/2 in UVA-induced melanogenesis in cultured normal human epidermal melanocytes. Pigment Cell Res. 2001;14:103–109. doi: 10.1034/j.1600-0749.2001.140205.x. [DOI] [PubMed] [Google Scholar]
- 45.Buscà R, Abbe P, Mantoux F, Aberdam E, Peyssonnaux C, Eychène A, Ortonne JP, Ballotti R. Ras mediates the cAMP-dependent activation of extracellular signal-regulated kinases (ERKs) in melanocytes. EMBO J. 2000;19:2900–2910. doi: 10.1093/emboj/19.12.2900. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Jung HG, Kim HH, Paul S, Jang JY, Cho YH, Kim HJ, Yu JM, Lee ES, An BJ, Kang SC, Bang BH. Quercetin -3-O-β-D-g lucopyranosyl-(1→6)-β-D-glucopyranoside suppresses melanin synthesis by augmenting p38 MAPK and CREB signaling pathways and subsequent cAMP down-regulation in murine melanoma cells. Saudi J Biol Sci. 2015;22:706–713. doi: 10.1016/j.sjbs.2015.03.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.