

Summary
High prevalence numbers of extended‐spectrum beta‐lactamase‐ (ESBL‐)/plasmid‐mediated AmpC beta‐lactamase‐ (pAmpC‐) producing Escherichia coli in broiler chicken and their distribution along the broiler production chain is an ongoing problem in food production. We, therefore, investigated resistant isolates along the broiler production chain to determine whether there is a constantly occurring direct vertical transmission of the ESBL‐/pAmpC‐producing E. coli from the parent flocks to their offspring or not. We, furthermore, analysed the isolates concerning the occurrence of virulence factors and their ability to form biofilms to estimate their potential to effectively colonize broiler chickens and/or persist and survive in the environment of the broiler production facilities. Using whole genome sequencing, we could show that ESBL‐/pAmpC‐producing E. coli were likely transferred in a step‐wise process along the broiler production chain but not directly from the parent flock to the fattening flock with every single batch of offspring chickens. Additionally, resistant E. coli strains showing an extraintestinal pathogenic genotype as well as high numbers of virulence‐associated genes including the production of curli fibres and cellulose have high capabilities to persist and spread in the broiler production chain.
Introduction
Extended‐spectrum beta‐lactamase‐ (ESBL‐) and plasmid‐mediated AmpC beta‐lactamase‐ (pAmpC‐) producing Enterobacteriaceae are an increasing problem in public health and veterinary medicine (Pitout and Laupland, 2008; Ewers et al., 2012; Kaesbohrer et al., 2012). A major concern is the high prevalence of these resistant bacteria in the broiler production chain. ESBL‐/pAmpC‐producing Enterobacteriaceae were frequently isolated from broiler fattening farms worldwide (Bortolaia et al., 2010; Randall et al., 2011; Kameyama et al., 2013; Maciuca et al., 2015; Trongjit et al., 2016) but were also found in broiler (grand) parent flocks (Dierikx et al., 2013; Agersø et al., 2014; Mo et al., 2014; Zurfluh et al., 2014). Therefore, different transmission scenarios of the resistant bacteria in the broiler production pyramid are discussed: A transmission from prior stages into the fattening farms (Giovanardi et al., 2005; Dierikx et al., 2013; Nilsson et al., 2014; Huijbers et al., 2016) as well as an insufficient cleaning and disinfection procedure in the chicken barns (Hiroi et al., 2012; Luyckx et al., 2015b). We recently showed a pseudo‐vertical transfer of resistant bacteria from the parent flocks into the hatchery via contaminated eggshells (Projahn et al., 2017). In addition, it was proved by whole genome analyses that there is in fact a horizontal transfer of ESBL producers between consecutively fattened flocks regardless of cleaning and disinfection procedures (Daehre et al., 2017). However, it still remained unclear whether the resistant bacteria are permanently and repeatedly transferred along the whole broiler production chain with each production cycle, or if this transfer only occurs occasionally and certain strains, once introduced on the farm, are circulating constantly.
Escherichia coli naturally inhabits the gastrointestinal tract of mammals and birds and can be classified into commensal and pathogenic strains. Intestinal pathogenic E. coli (IPEC) are causing diarrhoea syndromes and seldom colonize healthy humans whereas extraintestinal pathogenic E. coli (ExPEC) often innocuously colonize the gut but have the ability to cause severe infections such as meningitis or blood stream infections (Vila et al., 2016). Recent studies compared the occurrence of certain virulence‐associated genes (VAGs) in avian pathogenic E. coli (APEC), uropathogenic E. coli (UPEC) and commensal E. coli from humans and broiler chickens (Kemmett et al., 2013; de Carli et al., 2015; van Hoek et al., 2016; Paixao et al., 2016; Silveira et al., 2016). It turned out that certain VAGs (e.g. fimC, iha, tsh, ireA, neuC, astA, irp2, vat, iucD, chuA, iss) could not be statistically significant linked to a particular type of pathogenicity, but were also detected in strains from healthy hosts. Therefore, some of these factors are increasingly termed as ‘fitness factors’ because they contribute to a successful colonization and enhanced survival in the gut and the environment but not necessarily cause diseases (Smith et al., 2007; Frommel et al., 2013; Wigley, 2015; Vila et al., 2016). ESBL‐/pAmpC‐producing E. coli from healthy hosts were usually classified as commensal strains but recent investigations found out that these resistant strains also show characteristics of ExPEC or ExPEC‐like strains and, therefore, have the ability for an enhanced colonization of the gut.
The presented study aimed three questions. First, is there a direct top‐down transmission of ESBL‐/pAmpC‐producing E. coli from the parent flock via the hatchery to the respective fattening flock in the absence of an antibiotic treatment? Second, is the prevalence of ESBL‐/pAmpC‐producing E. coli in healthy broiler flocks influenced by their status as an ExPEC? Third, have the isolates found in the broiler production chain a higher ability to persist and survive in the environment?
To investigate the top‐down transmissions across all stages of the broiler production chain (parent flock, hatchery, fattening flock/barn), we analysed 44 ESBL‐/pAmpC‐producing E. coli from two different broiler production chains (chain C and F) which were not treated with antibiotics by whole genome analyses (Daehre et al., 2017; Projahn et al., 2017). Ten ESBL‐/pAmpC‐positive strains from parent screening samples (S) and other parent or fattening flocks (A/11, B/41, D, E/73) showing the same phylogroup and resistance genes that were found in chain C or F, respectively, were also included to show a potential circulation of certain strains within the broiler production chain (please see Fig. 4 and Table S1 for assignment/glossary of samples and strains). Secondly, to address the question whether there is a difference in the VAGs related to ExPEC between high and low prevalent strains, 89 selected ESBL‐/pAmpC‐positive strains from three healthy broiler flocks (C/41, E/74, F/74) with different prevalence were analysed for their VAGs. In addition, to determine the potential to persist and survive in the environment, the expression of two major biofilm‐associated extracellular matrix components (cellulose, curli fimbriae) was investigated for 13 strains from these three flocks.
Figure 4.
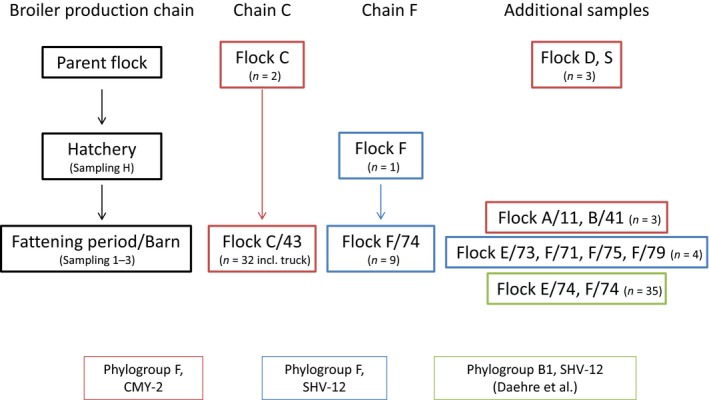
Origin and number of E. coli isolates used in the study for transmission investigations using WGS. Isolates of the same genetic characteristics (phylogroup + ESBL/pAmpC gene) are coloured in red, blue and green, respectively. Additional samples from the fattening period of flock F (F/71, F/75, F/79) were collected at the same time from other barns within the same farm as F/74. Isolates from flock E/74 and F/74 were only investigated concerning VAGs and Biofilm production.
Results
Phylogenetic analyses
In silico multilocus sequence typing (MLST) using whole genome sequences revealed three different sequence types (STs). Isolates, previously determined as phylogroup F harbouring a bla CMY‐2 gene and sampled from the broiler chain C were assigned as ST‐354. Samples from parent flocks D and screening parent flock S were also of ST‐354 whereas isolates of the fattening flocks A/11 and B/41 turned out to be of ST‐38. All isolates of phylogroup F harbouring a bla SHV‐12 gene of broiler chain F as well as from the additional samples belonged to the ST‐117.
Comparative analyses of single nucleotide polymorphisms (SNPs) were carried out for isolates of the MLST types ST‐354 (n = 37) and ST‐117 (n = 14) respectively (Figs 1 and 2). The calculation of the number of SNP differences between the ST‐354 strains (bla CMY‐2) resulted in one large cluster (fattening cluster: 0‐32 SNPs difference, max. 6.4 SNPs Mbp−1), which included all the isolates from the fattening flock C/41 (n = 31) as well as the isolate 5146 from the transportation truck of the same flock (Fig. 1). The SNP differences between this fattening cluster and the two strains (5008, 4989) from the respective parent flock C were calculated as 53 and 78 SNPs (10.5 and 15.6 SNPs Mbp−1) respectively. The SNP differences between the fattening cluster and the samples from parent flock D and screening parent flock S were calculated as up to 118 SNPs (23.6 SNPs Mbp−1).
Figure 1.
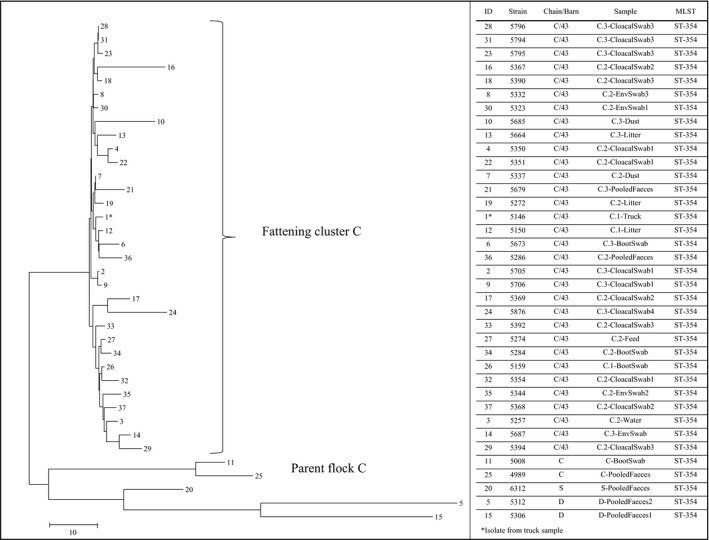
Phylogenetic tree of ST‐354 strains. A neighbour‐joining tree was calculated (with 1000 bootstraps) using a SNP distance matrix calculated by MEGA7 and the Harvest suite. Isolates are indicated by the respective strain number and the sample from which they originate as well as the chain and barn number of the respective sampling. The sample name presents further information like the sampling time points (C.1 – arrival of the chicks at the farm, C.2 – middle of the fattening period, C.3 – end of the fattening period) and the sample matrix.
Figure 2.
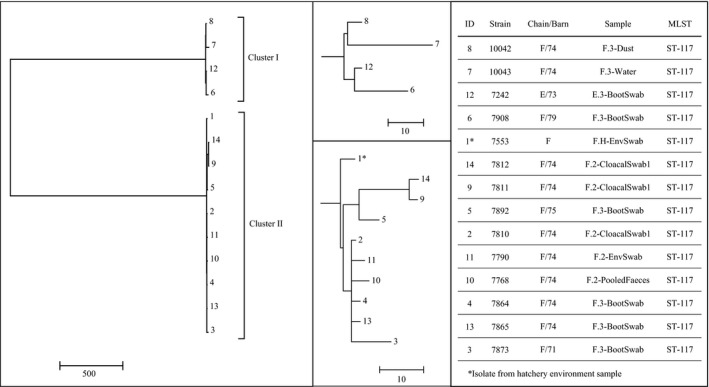
Phylogenetic tree of ST‐117 strains. A neighbour‐joining tree was calculated (with 1000 bootstraps) using a SNP distance matrix calculated by MEGA7 and the Harvest suite. Isolates are indicated by the respective strain number and the sample from which they originate as well as the chain and barn number of the respective sampling (E/73, F/71, F/74, F/75, F/79). The sample name presents further information like the sampling time points (F.H – Hatchery, F.2 – middle of the fattening period of chain F, F.3 – end of the fattening period of chain F, E.3 – end of the fattening period of chain E which was fattened at the same farm previously to the chickens of chain F) and the sample matrix.
The SNP difference calculation of the ST‐117 strains revealed two clusters (Fig. 2). The first one includes only four isolates of which two were collected from samples of fattening flock F/74 (chain F) as well as two screening isolates of fattening flocks E/73 and F/79. The second cluster comprises the isolate 7553 from the hatchery environment (chain F), seven isolates of the fattening period of flock F/74 (chain F) as well as two isolates from other flocks fattened at the same farm at the same time as fattening flock F (barn 74) but in different barns (F/71, F/75). SNP differences among the strains of the clusters varied between 23 and 68 SNPs (max. 13.6 SNPs Mbp−1) and 4 to 51 (max. 10.2 SNPs Mbp−1) SNPs, respectively, whereas the differences between both clusters were determined as 3114 to 3147 SNPs (max. 629.4 SNPs Mbp−1).
Virulence‐associated genes
Whole genome contigs of the 89 strains were also investigated for their virulence profiles analysing 89 VAGs (Table S3) of which 19 genes were typically associated with IPEC strains (InVAGs) and 70 with ExPEC strains (ExVAGs). All strains were negative concerning the occurrence of InVAGs. In contrast, of the 70 ExVAGs, 37 genes were detected in ST‐354 strains (53%; chain C, screening samples D+S), up to 25 genes in ST‐117 strains (36%; chain F, screening samples E/73, F/71, F/75, F/79) and in the isolates of ST‐38 (36%; samples A/11, B/41), respectively, and 19 ExPEC‐related VAGs in the ST‐2307 isolates (27%). All isolates of the different MLST types share 14 ExVAGs which were as follows: crl (curli fibre gene), csgA (curli fibre‐encoding gene), feoA/B (major bacterial ferrous iron transporter, iron (II) transport system), fimC and fimH (type 1 fimbriae), matA (meningitis‐associated and temperature‐regulated fimbriae), iroN (catecholate salmochelin receptor), iss (increased serum survival), sitA and sitC (Salmonella iron transport system gene), cvi (structural genes of colicin V operon), ompA (outer membrane protein), traT (transfer protein), astA (heat stable cytotoxin associated with enteroaggregative E. coli) and malX (pathogenicity‐associated island marker CFT073). Most variations between the isolates of the different MLST types were observed in the numbers of genes which contribute to adhesion and iron uptake (Table 1). Moreover, certain traits were exclusively found in isolates of a respective ST. bfpM (bundle‐forming pilus), ibeA (invasion of brain endothelium), the pap operon (Pap pili adhesin), tia (toxigenic invasion locus in ETEC isolates) and neuC (K1 capsular polysaccharide) were detected in isolates of the ST‐354 whereas fyuA (yersiniabactin receptor), ireA (iron‐responsive element), irp2 (yersiniabactin synthesis) and pic (serine protease autotransporter) occurred only in ST‐117 strains. hek/hra (heat‐resistant agglutinin) could be detected in the three ST‐38 isolates from the parent flocks A and B, only. Even though all analysed isolates showed very high numbers of detectable VAGs, according to the definition of ExPEC strains by Johnson et al. (2003)(≥2 of papA/C, afa/dra, sfa/foc, iutA and kpsMTII), only the ST‐354 strains and two of three ST‐38 strains (3565, 10026) could be assigned as an ExPEC. Interestingly, ExPEC‐related invasion genes also occurred in ST‐354 and ST‐38 strains, only.
Table 1.
Mean values of detected virulence‐associated genes (VAGs). Highly variable categories are highlighted in bold
| MLST | ST‐354 | ST‐117 (cluster 1) | ST‐117 (cluster 2) | ST‐2307 | ST‐38 |
|---|---|---|---|---|---|
| Total no. of ExVAGs (70;14a) | 36.7 | 21 | 24.3 | 19 | 23.7 |
| Adhesion (34;5a) | 15.9 | 4 | 5 | 5 | 7 |
| Invasion (4;0a) | 2 | 0 | 0 | 0 | 1 |
| Iron uptake (12;4a) | 8.8 | 8 | 11 | 6 | 8 |
| Protection (6;4a) | 6 | 3 | 4 | 4 | 4.7 |
| Toxins (12;1a) | 3 | 3 | 3.3 | 3 | 2 |
| Miscellaneous (2;1a) | 1 | 2 | 2 | 1 | 1 |
Number of genes which were detected in all isolates.
Macrocolony assay
Two sets of isolates were analysed by the macrocolony assay. The first one comprised thirteen isolates from the whole genome approach, at least three isolates of each MLST type (chain C: ST‐354, chain F: ST‐117, screening samples from A/11+B/41: ST‐38, Daehre et al. chain E: ST‐2307). Isolates of fattening flocks C/41, F/74 and E/74 occurred with different prevalence in the respective fattening flocks (flock C/41: 73.1%, flock E/74: 26.4%, flock F/74: 5.7%). The second strain set included 42 isolates from investigations of seven different broiler production chains covering all detected genetic profiles (phylogroup + ESBL‐/pAmpC gene) to get an overall impression on cellulose and curli production in different resistant isolates from the whole broiler production chain (Daehre et al., 2017; Projahn et al., 2017) (Tables S1 and S2). Isolates of the first set showed both, production of cellulose and curli, regardless of the flock prevalence, whereas in the second one, various phenotypes were determined (Fig. 3). In total, 28.6% (12/42) of the second strain set showed neither a production of curli nor cellulose, 7.1% (3/42) showed only a cellulose‐positive phenotype and 9.5% (4/42) of the isolates were positive for a curli production, only. Most of the strains (52.4%, 22/42) were phenotypically positive for both, curli and cellulose production. Only 16.7% (7/42) could not be assigned to a distinct phenotype in the macrocolony assay.
Figure 3.
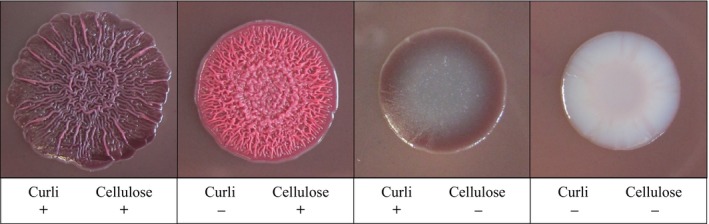
Macrocolony assay. Categories of detected phenotypes in the investigated isolates. The figure shows exemplary macrocolonies of isolates 6818 (+/+), 6922 (−/+), 3233 (+/−), 6985 (−/−).
Discussion
We performed our study addressing the questions of a direct top‐down transmission of ESBL‐/pAmpC‐producing E. coli from the parent flock via the hatchery to the respective fattening flock in the absence of an antibiotic treatment as well as the role of the ExPEC status and the ability to form biofilms on the prevalence of ESBL‐/pAmpC‐producing E. coli in healthy broiler flocks. Therefore, we investigated isolates (ST‐354) from a parent flock and the corresponding fattening flock (chain C) using whole genome analyses. It turned out that the isolates of the parent flock and the fattening flock are closely related but the grouping into different clusters and the analyses of the SNP differences showed that there was no direct vertical spread of these resistant bacteria. ESBL strains from the fattening period, collected during a time frame of five weeks, differed in not more than 6.4 SNPs Mbp−1. In contrast, isolates of the parent flock were collected only 17 days earlier but the SNP differences between isolates of the parent flock and the fattening flock were about 10.5 SNPs Mbp−1. The number of SNP differences between the strains of the fattening period is higher than the numbers published for clonal outbreaks of enterohemorrhagic E. coli (EHEC) (de Been et al., 2014; Berenger et al., 2015; Rusconi et al., 2016) but overall lower than those described for interspecies transmissions of clonal ST‐410 E. coli strains (Schaufler et al., 2016b). These findings suggest that ESBL‐/pAmpC‐resistant E. coli were more likely transferred from the parent flock into the production chain via an earlier event but not directly along the investigated batch of broiler chicken.
In addition, we isolated an ESBL‐E. coli strain from the transportation truck and from the litter inside of the barn from the first day of the fattening period which clustered together with all the other strains from the whole fattening period of chain C. This indicates that a transmission of the resistant bacteria can also occur via the transportation process and that the resistant bacteria must have been already present in the hatchery than directly transmitted from the parent flock. This is in concordance with our findings that the resistant bacteria were introduced into the hatchery via contaminated eggshells and the hatchery acts as a bottleneck for the spread of these strains (Projahn et al., 2017).
Investigation of the ST‐117 strains of chain F underlines the fact that there were ESBL E. coli already present in the hatchery which lead to the colonization of the recently hatched chicks. Whole genome analyses revealed two clusters. The first one comprised strains from the environment of the hatchery and the fattening period confirming the hatchery as a contamination source. The second ST‐117 cluster comprised strains of the fattening flock F as well as strains of two other barns from the same farm. Due to the small number of SNP differences between the isolates, a transmission or exchange of these ESBL E. coli between the barns of the same farm is very likely. It was previously shown that (ESBL‐) E. coli can survive the cleaning and disinfection procedures in the chicken barns which finally leads to a spread and the colonization of broiler chickens on the same farms (Luyckx et al., 2015a,b; Daehre et al., 2017). Interestingly, the two clusters differ in up to 629.4 SNPs Mbp−1 indicating that two different clonal lines of ST‐117 strains were introduced into the same flock.
We previously investigated up to 150 samples from the fattening period of seven flocks each showing that there is an overall highly diverse E. coli population on these fattening farms (Daehre et al., 2017). Together with the results from this study, we could show that not only one prominent vertical or horizontal transmission route of ESBL producers exists. More precisely, certain ESBL‐/pAmpC‐producing E. coli seemed to be more likely introduced into the broiler production chain decades ago and now circulate constantly in different broiler production‐related environments and evolve even without antibiotic pressure (Nilsson et al., 2014; Huijbers et al., 2016; Mo et al., 2016).
Investigations on the phenotypic characteristics and virulence patterns of ESBL‐/pAmpC‐producing E. coli indicate that certain traits usually associated with virulence factors correlate with the ability to adhere, persist and adapt to certain environmental circumstances (Guenther et al., 2012; Rodiger et al., 2015; Mo et al., 2016; Paixao et al., 2016; Schaufler et al., 2016a). In our study, high prevalence strains of ST‐354 could be assigned as ExPEC and overall showed the highest number of ExVAGs, especially genes referring to adhesion factors, compared to low prevalent strains of ST‐117 and ST‐2307. ST‐354 isolates harboured the pap operon which was shown to be responsible for the in vitro adhesion to not only human but also animal cell lines (Stromberg et al., 1990; Frommel et al., 2013). But intestinal colonization is also depending on the iron uptake ability of E. coli (Nowrouzian et al., 2006; Tenaillon et al., 2010). On the one hand, ST‐117 strains of our study had the highest number of genes referring to iron uptake, on the other hand, these isolates had low numbers of genes which contribute to adherence factors. Furthermore, these strains were found with low prevalence in the investigated fattening flock F (5.7%). Interestingly, ST‐2307 isolates had overall the lowest numbers of ExVAGs including adherence traits but occurred with higher prevalence in the fattening flock (23.5%) than ST‐117 strains assuming that not only high numbers of VAGs but a certain combination or set of virulence traits can promote the colonization of the avian gut. However, specific factors for the binding to avian host cells are poorly understood and should be therefore further investigated.
The ability to form bacterial biofilms is a benefit in the survival against host defence factors, antibiotics, physical and chemical stress as well as disinfectants (Hall‐Stoodley et al., 2004; Flemming and Wingender, 2010). Using a macrocolony assay, we could show that more than half of our tested strains from both investigated sets were able to produce curli fibres and the exopolysaccharide cellulose which are necessary to form stable and strong biofilm matrices (Olsen et al., 1989; Barnhart and Chapman, 2006; Uhlich et al., 2006; Serra et al., 2013, 2015). These positive strains were isolated from all the seven investigated flocks by Daehre et al. and included also the 14 selected isolates from the whole genome analyses of ST‐354, ST‐117, ST‐38 and ST‐2307 from our study. Here, the production of curli fibres and cellulose was not linked to a certain phylogroup or ESBL/pAmpC resistance gene and was also independent from the prevalence of these strains in the respective flocks (flock C: 73.1%, flock E: 26.4%, flock F: 5.7%) indicating that resistant E. coli occurring on broiler farms in general show good abilities to survive on the farms and in the environment. Cellulose also has high water holding capacities which protects the bacteria against dehydration (O'Sullivan, 1997). This could be a big advantage in surviving, for example, the cleaning and disinfection procedure in the hatchery and the broiler fattening farms. Daehre et al. already showed that the ST‐2307 strains were transmitted from one broiler flock to the consecutively fattened flock in the same barn. The results of the present study suggest that biofilm production as well as high numbers of gene conferring to adherence might support the persistence of resistant bacteria and the stable colonization of broiler chickens.
Addressing our first question, we demonstrated that the transmission of ESBL‐/pAmpC‐producing Enterobacteriaceae in the broiler production chain is more or less a summary of different transmission routes including persistence and circulation of different clonal lineages independently from antibiotic usage instead of a direct vertical top‐down transmission along the broiler production chain. Second, the occurrence of high numbers of ExPEC VAGs seemed to contribute the colonization process. However, regarding the avian host, not only high numbers of VAGs but also a certain set of virulence traits seemed to contribute this colonization. This needs to be further investigated. Finally, high numbers of isolates producing curli fibres and cellulose were detected which support the survival of resistant strains in the environment of the broiler production chain.
Experimental procedures
Bacterial isolates
ESBL‐/pAmpC‐producing E. coli originate from individual animal samples (cloacal swabs) and environmental samples (swabs and housing environment) of different stages of the broiler production chain (parent flock, hatchery, fattening farm) collected in Germany during the years 2014 and 2015 (Daehre et al., 2017; Projahn et al., 2017).
In total, 54 isolates were selected on the basis of their phylogroup and the bla resistance gene for detailed genomic analyses using WGS to determine possible transmission events along the broiler production chain (Fig. 4; Table S1).
Escherichia coli of phylogroup F encoding a CMY‐2 cephamycinase were collected from samples of the parent flock C (n = 2) as well as from the respective fattening flock C (barn 43; n = 31) and the transportation truck (n = 1) of fattening flock C from the hatchery to the fattening farm. Additionally, strains showing the same characteristics were isolated from screening samples (faeces or boot swab), collected at different time points within the study, of parent flock D (n = 2) and parent screening flock S (n = 1) and the fattening flocks A (barn 11; n = 1) and B (barn 41; n = 2).
Isolates of phylogroup F coding for a SHV‐12 beta‐lactamase were determined from the investigation of the broiler chain F. Here, an environmental gauze swab from the hatchery (hatching of the flock F chicks; n = 1) as well as different samples from the fattening period of flock F (barn 74; cloacal swabs, gauze swabs, environmental swabs, housing environment; n = 9) were analysed. Additionally, isolates of phylogroup F encoding a SHV‐12 beta‐lactamase from boot swabs of fattening barns E/71, F/73, F/75 and F/79 were included in the comparative genome analyses. These samples originate from other flocks fattened at the same time at the same farm as flock F or E, respectively, but in different barns.
Escherichia coli isolates of phylogroup B1 harbouring a bla SHV‐12 gene (n = 35) were included in the study for the investigation of VAGs. These isolates were derived from two broiler flocks subsequently fattened in the same barn (flock E/74 and F/74) and from an additional gauze swab from the transportation truck of flock F (Daehre et al., 2017).
Whole genome sequencing
Genomic DNA preparation was performed using the MasterPure™ DNA purification kit (Epicentre, Illumina) according to the manufacturer's instructions. Whole genome sequencing was performed using the Nextera XT Kit for library preparation and the Illumina MiSeq Reagent Kit v3 (300‐bp paired‐end sequencing with 50–100× coverage). The NGS tool kit (Patel and Jain, 2012) was used for quality control of the sequence read data. High‐quality reads were de novo assembled into contiguous sequences using CLC Genomics Workbench 8.0 (CLC bio, Aarhus, Denmark), and annotation of draft genomes was performed with RAST server (Aziz et al., 2008).
Phylogenetic analyses
Whole genome data were used for in silico determination of MLST types using the CGE Bacterial Analysis Pipeline (Thomsen et al., 2016).
The bacterial core genome and the number of SNP differences were calculated between isolates of the same MLST type using Harvest suite 1.0 (Treangen et al., 2014) and mega 7.0 (Kumar et al., 2016). Phylogenetic trees were constructed using a pairwise distance matrix and the neighbour‐joining algorithm (mega 7.0, 1000 bootstraps) based on the number of calculated SNP differences between the isolates.
Virulence‐associated genes
The presence of 89 virulence‐associated genes (VAGs) in the genomes of all 54 investigated isolates (Fig. 4) was checked using an automated in‐house blast search. VAG analyses were also performed for additional 35 isolates of the ST‐2307 (fattening flock E), which originate from samples collected from the same barn as fattening flock F but from the previous fattening round (Table S3), to overall investigate samples from fattening flocks with differing ESBL/pAmpC prevalence (flock C: 73.1%, flock E: 26.4%, flock F: 5.7%). Strains were previously investigated concerning a possible horizontal transmission scenario (Daehre et al., 2017).
Phenotypic characterization
To determine the expression of biofilm‐associated extracellular matrix components (cellulose, curli fimbriae), a macrocolony assay (Schaufler et al., 2016a) was applied to thirteen selected isolates from the whole genome approach (at least three isolates of each determined MLST type) as well as to 38 E. coli and four E. fergusonii isolates of the studies of Projahn et al. and Daehre et al. representing all the different detected isolate profiles (phylogroup + bla resistance gene) determined during the investigation of seven broiler production chains (Tables S1 and S2).
Conflict of Interest
The authors declare no conflict of interest.
Supporting information
Table S1. Isolates used in the study for investigation of vertical transmissions, occurence of virulence associated genes and production onf curli.
Table S2. Isolates of the second set investigated by a macrocolony assay. Isolates comprise all detected genotypes (phylogroup+bla resistance).
Table S3. Numbers and occurrence of the 80 investigated virulence associated genes.
Acknowledgements
The presented study was funded by the Federal Ministry of Education and Research (BMBF) RESETII consortium (grant no.: 01KI1313C, http://www.reset-verbund.de). The authors gratefully thank Maja Thieck for excellent technical assistance as well as Inga Eichhorn and Petra Kriencke (Institute for Microbiology, Freie Universität Berlin, Berlin, Germany) for performing the whole genome sequencing.
Microbial Biotechnology (2018) 11(6), 1017–1026
Funding Information
The presented study was funded by the Federal Ministry of Education and Research (BMBF) RESETII consortium (grant no.: 01KI1313C, http://www.reset-verbund.de).
References
- Agersø, Y. , Jensen, J.D. , Hasman, H. , and Pedersen, K. (2014) Spread of extended spectrum cephalosporinase‐producing Escherichia coli clones and plasmids from parent animals to broilers and to broiler meat in a production without use of cephalosporins. Foodborne Pathog Dis 11: 740–746. 10.1089/fpd.2014.1742. [DOI] [PubMed] [Google Scholar]
- Aziz, R.K. , Bartels, D. , Best, A.A. , DeJongh, M. , Disz, T. , Edwards, R.A. , et al (2008) The RAST Server: rapid annotations using subsystems technology. BMC Genom 9: 75 10.1186/1471-2164-9-75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnhart, M.M. , and Chapman, M.R. (2006) Curli biogenesis and function. Annu Rev Microbiol 60: 131–147. 10.1146/annurev.micro.60.080805.142106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Been, M. , Lanza, V.F. , de Toro, M. , Scharringa, J. , Dohmen, W. , Du, Y. , et al (2014) Dissemination of cephalosporin resistance genes between Escherichia coli strains from farm animals and humans by specific plasmid lineages. PLoS Genet 10: e1004776 10.1371/journal.pgen.1004776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berenger, B.M. , Berry, C. , Peterson, T. , Fach, P. , Delannoy, S. , Li, V. , et al (2015) The utility of multiple molecular methods including whole genome sequencing as tools to differentiate Escherichia coli O157:H7 outbreaks. Euro Surveill 20: pii=30073. 10.2807/1560-7917.ES.2015.20.47.30073. [DOI] [PubMed] [Google Scholar]
- Bortolaia, V. , Guardabassi, L. , Trevisani, M. , Bisgaard, M. , Venturi, L. , and Bojesen, A.M. (2010) High diversity of extended‐spectrum beta‐lactamases in Escherichia coli isolates from Italian broiler flocks. Antimicrob Agents Chemother 54: 1623–1626. 10.1128/AAC.01361-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Carli, S. , Ikuta, N. , Lehmann, F.K.M. , da Silveira, V.P. , de Melo Predebon, G. , Fonseca, A.S.K. , and Lunge, V.R. (2015) Virulence gene content in Escherichia coli isolates from poultry flocks with clinical signs of colibacillosis in Brazil. Poult Sci 94: 2635–2640. 10.3382/ps/pev256. [DOI] [PubMed] [Google Scholar]
- Daehre, K. , Projahn, M. , Semmler, T. , Roesler, U. , and Friese, A. (2017) ESBL‐/AmpC‐producing Enterobacteriaceae in broiler farms – transmission dynamics at farm level. Microbial Drug Resistance [Epub ahead of print] 10.1089/mdr.2017.0150 [DOI] [PubMed] [Google Scholar]
- Dierikx, C.M. , van der Goot Jeanet, A. , Smith, H.E. , Kant, A. , and Mevius, D.J. (2013) Presence of ESBL/AmpC‐producing Escherichia coli in the broiler production pyramid: a descriptive study. PLoS ONE 8: e79005 10.1371/journal.pone.0079005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ewers, C. , Bethe, A. , Semmler, T. , Guenther, S. , and Wieler, L.H. (2012) Extended‐spectrum beta‐lactamase‐producing and AmpC‐producing Escherichia coli from livestock and companion animals, and their putative impact on public health: a global perspective. Clin Microbiol Infect 18: 646–655. 10.1111/j.1469-0691.2012.03850.x. [DOI] [PubMed] [Google Scholar]
- Flemming, H.‐C. , and Wingender, J. (2010) The biofilm matrix. Nat Rev Microbiol 8: 623–633. 10.1038/nrmicro2415. [DOI] [PubMed] [Google Scholar]
- Frommel, U. , Lehmann, W. , Rodiger, S. , Bohm, A. , Nitschke, J. , Weinreich, J. , et al (2013) Adhesion of human and animal Escherichia coli strains in association with their virulence‐associated genes and phylogenetic origins. Appl Environ Microbiol 79: 5817–5829. 10.1128/AEM.01384-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giovanardi, D. , Campagnari, E. , Ruffoni, L.S. , Pesente, P. , Ortali, G. , and Furlattini, V. (2005) Avian pathogenic Escherichia coli transmission from broiler breeders to their progeny in an integrated poultry production chain. Avian Pathol 34: 313–318. 10.1080/03079450500179046. [DOI] [PubMed] [Google Scholar]
- Guenther, S. , Bethe, A. , Fruth, A. , Semmler, T. , Ulrich, R.G. , Wieler, L.H. , and Ewers, C. (2012) Frequent combination of antimicrobial multiresistance and extraintestinal pathogenicity in Escherichia coli isolates from urban rats (Rattus norvegicus) in Berlin, Germany. PLoS ONE 7: e50331 10.1371/journal.pone.0050331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall‐Stoodley, L. , Costerton, J.W. , and Stoodley, P. (2004) Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2: 95–108. 10.1038/nrmicro821. [DOI] [PubMed] [Google Scholar]
- Hiroi, M. , Matsui, S. , Kubo, R. , Iida, N. , Noda, Y. , Kanda, T. , et al (2012) Factors for occurrence of extended‐spectrum β‐lactamase‐producing Escherichia coli in broilers. J Vet Med Sci 74: 1635–1637. [DOI] [PubMed] [Google Scholar]
- van Hoek, A.H.A.M. , Stalenhoef, J.E. , van Duijkeren, E. , and Franz, E. (2016) Comparative virulotyping of extended‐spectrum cephalosporin‐resistant E. coli isolated from broilers, humans on broiler farms and in the general population and UTI patients. Vet Microbiol 194: 55–61. 10.1016/j.vetmic.2016.04.008. [DOI] [PubMed] [Google Scholar]
- Huijbers, P.M.C. , Graat, E.A.M. , van Hoek Angela, H.A.M. , Veenman, C. , de Jong Mart, C.M. , and van Duijkeren, E. (2016) Transmission dynamics of extended‐spectrum β‐lactamase and AmpC β‐lactamase‐producing Escherichia coli in a broiler flock without antibiotic use. Preventive veterinary medicine 131: 12–19. 10.1016/j.prevetmed.2016.07.001. [DOI] [PubMed] [Google Scholar]
- Johnson, J.R. , Murray, A.C. , Gajewski, A. , Sullivan, M. , Snippes, P. , Kuskowski, M.A. , and Smith, K.E. (2003) Isolation and molecular characterization of nalidixic acid‐resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob Agents Chemother 47: 2161–2168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaesbohrer, A. , Schroeter, A. , Tenhagen, B.‐A. , Alt, K. , Guerra, B. , and Appel, B. (2012) Emerging antimicrobial resistance in commensal Escherichia coli with public health relevance. Zoonoses Public Health 59: 158–165. 10.1111/j.1863-2378.2011.01451.x. [DOI] [PubMed] [Google Scholar]
- Kameyama, M. , Chuma, T. , Yabata, J. , Tominaga, K. , Iwata, H. , and Okamoto, K. (2013) Prevalence and epidemiological relationship of CMY‐2 AmpC β‐lactamase and CTX‐M extended‐spectrum β‐lactamase‐producing Escherichia coli isolates from broiler farms in Japan. J Vet Med Sci 75: 1009–1015. [DOI] [PubMed] [Google Scholar]
- Kemmett, K. , Humphrey, T. , Rushton, S. , Close, A. , Wigley, P. , and Williams, N.J. (2013) A longitudinal study simultaneously exploring the carriage of APEC virulence associated genes and the molecular epidemiology of faecal and systemic E. coli in commercial broiler chickens. PLoS ONE 8: E67749 10.1371/journal.pone.0067749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumar, S. , Stecher, G. , and Tamura, K. (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol 33: 1870–1874. 10.1093/molbev/msw054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luyckx, K. , Dewulf, J. , van Weyenberg, S. , Herman, L. , Zoons, J. , Vervaet, E. , et al (2015a) Comparison of sampling procedures and microbiological and non‐microbiological parameters to evaluate cleaning and disinfection in broiler houses. Poult Sci 94: 740–749. 10.3382/ps/pev019. [DOI] [PubMed] [Google Scholar]
- Luyckx, K.Y. , van Weyenberg, S. , Dewulf, J. , Herman, L. , Zoons, J. , Vervaet, E. , et al (2015b) On‐farm comparisons of different cleaning protocols in broiler houses. Poult Sci 94: 1986–1993. 10.3382/ps/pev143. [DOI] [PubMed] [Google Scholar]
- Maciuca, I.E. , Williams, N.J. , Tuchilus, C. , Dorneanu, O. , Guguianu, E. , Carp‐Carare, C. , et al (2015) High prevalence of Escherichia coli‐producing CTX‐M‐15 extended‐spectrum beta‐lactamases in poultry and human clinical isolates in Romania. Microbial Drug Resist 21: 651–662. 10.1089/mdr.2014.0248. [DOI] [PubMed] [Google Scholar]
- Mo, S.S. , Norstrom, M. , Slettemeas, J.S. , Lovland, A. , Urdahl, A.M. , and Sunde, M. (2014) Emergence of AmpC‐producing Escherichia coli in the broiler production chain in a country with a low antimicrobial usage profile. Vet Microbiol 171: 315–320. 10.1016/j.vetmic.2014.02.002. [DOI] [PubMed] [Google Scholar]
- Mo, S.S. , Slettemeas, J.S. , Berg, E.S. , Norstrom, M. , and Sunde, M. (2016) Plasmid and host strain characteristics of Escherichia coli resistant to extended‐spectrum cephalosporins in the Norwegian broiler production. PLoS ONE 11: e0154019 10.1371/journal.pone.0154019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nilsson, O. , Borjesson, S. , Landen, A. , and Bengtsson, B. (2014) Vertical transmission of Escherichia coli carrying plasmid‐mediated AmpC (pAmpC) through the broiler production pyramid. J Antimicrob Chemother 69: 1497–1500. 10.1093/jac/dku030. [DOI] [PubMed] [Google Scholar]
- Nowrouzian, F.L. , Adlerberth, I. , and Wold, A.E. (2006) Enhanced persistence in the colonic microbiota of Escherichia coli strains belonging to phylogenetic group B2: role of virulence factors and adherence to colonic cells. Microbes Infect 8: 834–840. 10.1016/j.micinf.2005.10.011. [DOI] [PubMed] [Google Scholar]
- Olsen, A. , Jonsson, A. , and Normark, S. (1989) Fibronectin binding mediated by a novel class of surface organelles on Escherichia coli . Nature 338: 652–655. 10.1038/338652a0. [DOI] [PubMed] [Google Scholar]
- O'Sullivan, A.C. (1997) Cellulose: the structure slowly unravels. Cellulose 4: 173–207. 10.1023/A:1018431705579. [DOI] [Google Scholar]
- Paixao, A.C. , Ferreira, A.C. , Fontes, M. , Themudo, P. , Albuquerque, T. , Soares, M.C. , et al (2016) Detection of virulence‐associated genes in pathogenic and commensal avian Escherichia coli isolates. Poult Sci 95: 1646–1652. 10.3382/ps/pew087. [DOI] [PubMed] [Google Scholar]
- Patel, R.K. , and Jain, M. (2012) NGS QC Toolkit: a toolkit for quality control of next generation sequencing data. PLoS ONE 7: e30619 10.1371/journal.pone.0030619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitout, J.D.D. , and Laupland, K.B. (2008) Extended‐spectrum β‐lactamase‐producing Enterobacteriaceae. An emerging public‐health concern. Lancet Infect Dis 8: 159–166. 10.1016/S1473-3099(08)70041-0. [DOI] [PubMed] [Google Scholar]
- Projahn, M. , Daehre, K. , Roesler, U. , and Friese, A. (2017) Extended‐spectrum‐beta‐lactamase‐ and plasmid‐encoded cephamycinase‐producing Enterobacteria in the broiler hatchery as a potential mode of pseudo‐vertical transmission. Appl Environ Microbiol 83: e02364 10.1128/AEM.02364-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Randall, L.P. , Clouting, C. , Horton, R.A. , Coldham, N.G. , Wu, G. , Clifton‐Hadley, F.A. , et al (2011) Prevalence of Escherichia coli carrying extended‐spectrum β‐lactamases (CTX‐M and TEM‐52) from broiler chickens and turkeys in Great Britain between 2006 and 2009. The Journal of antimicrobial chemotherapy 66: 86–95. 10.1093/jac/dkq396. [DOI] [PubMed] [Google Scholar]
- Rodiger, S. , Kramer, T. , Frommel, U. , Weinreich, J. , Roggenbuck, D. , Guenther, S. , et al (2015) Intestinal Escherichia coli colonization in a mallard duck population over four consecutive winter seasons. Environ Microbiol 17: 3352–3361. 10.1111/1462-2920.12807. [DOI] [PubMed] [Google Scholar]
- Rusconi, B. , Sanjar, F. , Koenig, S.S.K. , Mammel, M.K. , Tarr, P.I. , and Eppinger, M. (2016) Whole genome sequencing for genomics‐guided investigations of Escherichia coli O157:H7 outbreaks. Front Microbiol 7: 985 10.3389/fmicb.2016.00985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaufler, K. , Semmler, T. , Pickard, D.J. , de Toro, M. , de La Cruz, F. , Wieler, L.H. , et al (2016a) Carriage of extended‐spectrum beta‐lactamase‐plasmids does not reduce fitness but enhances virulence in some strains of pandemic E. coli lineages. Front Microbiol 7: 336 10.3389/fmicb.2016.00336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaufler, K. , Semmler, T. , Wieler, L.H. , Wöhrmann, M. , Baddam, R. , Ahmed, N. , et al (2016b) Clonal spread and interspecies transmission of clinically relevant ESBL‐producing Escherichia coli of ST410‐another successful pandemic clone? FEMS Microbiol Ecol 92: fiv155 10.1093/femsec/fiv155. [DOI] [PubMed] [Google Scholar]
- Serra, D.O. , Richter, A.M. , and Hengge, R. (2013) Cellulose as an architectural element in spatially structured Escherichia coli biofilms. J Bacteriol 195: 5540–5554. 10.1128/JB.00946-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Serra, D.O. , Klauck, G. , and Hengge, R. (2015) Vertical stratification of matrix production is essential for physical integrity and architecture of macrocolony biofilms of Escherichia coli . Environ Microbiol 17: 5073–5088. 10.1111/1462-2920.12991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silveira, F. , Maluta, R.P. , Tiba, M.R. , de Paiva, J.B. , Guastalli, E.A.L. , and da Silveira, W.D. (2016) Comparison between avian pathogenic (APEC) and avian faecal (AFEC) Escherichia coli isolated from different regions in Brazil. Vet J 217: 65–67. 10.1016/j.tvjl.2016.06.007. [DOI] [PubMed] [Google Scholar]
- Smith, J.L. , Fratamico, P.M. , and Gunther, N.W. (2007) Extraintestinal pathogenic Escherichia coli . Foodborne Pathog Dis 4: 134–163. 10.1089/fpd.2007.0087. [DOI] [PubMed] [Google Scholar]
- Stromberg, N. , Marklund, B.I. , Lund, B. , Ilver, D. , Hamers, A. , Gaastra, W. , et al (1990) Host‐specificity of uropathogenic Escherichia coli depends on differences in binding specificity to Gal alpha 1‐4Gal‐containing isoreceptors. EMBO J 9: 2001–2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tenaillon, O. , Skurnik, D. , Picard, B. , and Denamur, E. (2010) The population genetics of commensal Escherichia coli . Nat Rev Microbiol 8: 207–217. 10.1038/nrmicro2298. [DOI] [PubMed] [Google Scholar]
- Thomsen, M.C.F. , Ahrenfeldt, J. , Cisneros, J.L.B. , Jurtz, V. , Larsen, M.V. , Hasman, H. , et al (2016) A bacterial analysis platform: an integrated system for analysing bacterial whole genome sequencing data for clinical diagnostics and surveillance. PLoS ONE 11: e0157718 10.1371/journal.pone.0157718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Treangen, T.J. , Ondov, B.D. , Koren, S. , and Phillippy, A.M. (2014) The harvest suite for rapid core‐genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol 15: 524 10.1186/PREACCEPT-2573980311437212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trongjit, S. , Angkittitrakul, S. , and Chuanchuen, R. (2016) Occurrence and molecular characteristics of antimicrobial resistance of Escherichia coli from broilers, pigs and meat products in Thailand and Cambodia provinces. Microbiol Immunol 60: 575–585. 10.1111/1348-0421.12407. [DOI] [PubMed] [Google Scholar]
- Uhlich, G.A. , Cooke, P.H. , and Solomon, E.B. (2006) Analyses of the red‐dry‐rough phenotype of an Escherichia coli O157:H7 strain and its role in biofilm formation and resistance to antibacterial agents. Appl Environ Microbiol 72: 2564–2572. 10.1128/AEM.72.4.2564-2572.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vila, J. , Saez‐Lopez, E. , Johnson, J.R. , Romling, U. , Dobrindt, U. , Canton, R. , et al (2016) Escherichia coli. An old friend with new tidings. FEMS Microbiol Rev 40: 437–463. 10.1093/femsre/fuw005. [DOI] [PubMed] [Google Scholar]
- Wigley, P. (2015) Blurred lines. Pathogens, commensals, and the healthy gut. Front Vet Sci 2: 40 10.3389/fvets.2015.00040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zurfluh, K. , Wang, J. , Klumpp, J. , Nüesch‐Inderbinen, M. , Fanning, S. , and Stephan, R. (2014) Vertical transmission of highly similar blaCTX‐M‐1‐harbouring IncI1 plasmids in Escherichia coli with different MLST types in the poultry production pyramid. Front Microbiol 5: 519 10.3389/fmicb.2014.00519. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Isolates used in the study for investigation of vertical transmissions, occurence of virulence associated genes and production onf curli.
Table S2. Isolates of the second set investigated by a macrocolony assay. Isolates comprise all detected genotypes (phylogroup+bla resistance).
Table S3. Numbers and occurrence of the 80 investigated virulence associated genes.