

Abstract Abstract
An extended incubation strategy to culture slow growing members of anaerobic fungi resulted in the isolation of a novel anaerobic fungus from the rumen of a goat after 15 days. The novel genus, represented by type strain G1SC, showed filamentous monocentric thallus development and produced uniflagellate zoospores, hence, showing morphological similarity to the genera Piromyces, Buwchfawromyces, Oontomyces and Pecoramyces. However, strain G1SC showed genetic similarity to the genus Anaeromyces, which, though produces uniflagellate zoospore, also exhibits polycentric thallus development. Moreover, unlike Anaeromyces, strain G1SC did not show hyphal constrictions, instead produced a branched, determinate and anucleate rhizoidal system. This fungus also displayed extensive sporangial variations, both exogenous and endogenous type of development, short and long sporangiophores and produced septate sporangia. G1SC utilised various complex and simple substrates, including rice straw and wheat straw and produced H2, CO2, formate, acetate, lactate, succinate and ethanol. Phylogenetic analysis, using internal transcribed spacer 1 (ITS1) and D1/D2 domain of large-subunit (LSU) rRNA locus, clearly showed a separate lineage for this strain, near Anaeromyces. The ITS1 based geographical distribution studies indicated detection of environmental sequences similar (93–96%) to this strain from cattle faeces. Based on morphological and molecular characterisation results of strain G1SC, we propose a novel anaerobic fungus Liebetanzomycespolymorphusgen. et sp. nov., in the phylum Neocallimastigomycota.
Keywords: Anaerobe, fungal diversity, novel genus, phylogeny, rumen fungi, taxonomy
Introduction
Anaerobic fungi inhabit the gut of various herbivorous animals where they play a pivotal role in the degradation of lignocellulosic feed (Gruninger et al. 2014). Some recent reports even suggest the presence of anaerobic fungi outside the gut of terrestrial animals and within the gut of sea animals and sediments (Ivarsson et al. 2016; Edwards et al. 2017; Picard 2017). Taxonomically, anaerobic fungi are the only member of phylum Neocallimastigomycota, class Neocallimastigomycetes, order Neocallimastigales and family Neocallimastigaceae (Hibbett et al. 2007). So far, ten genera of these fungi have been reported, namely Anaeromyces, Caecomyces, Cyllamyces, Neocallimastix, Orpinomyces, Piromyces, Buwchfawromyces, Feramyces, Oontomyces and Pecoramyces, the last four of which have been described very recently (Callaghan et al. 2015; Dagar et al. 2015; Hanafy et al. 2017; Hanafy et al. 2018).
The total number of anaerobic fungal species is reported to be 29, but this number is not validated due to several taxonomy related issues, including incorrect or repetitive naming (Paul et al. 2018). Several culture-independent studies, moreover, suggest the presence of at least 25 genus- and 119 species-equivalent taxa of anaerobic fungi, highlighting the need for continuous attempts for the cultivation of these additional taxa into axenic cultures (Kittelmann et al. 2012; Koetschan et al. 2014; Paul et al. 2018). This suggested number of total genera and species may even be higher as some of the primers used during routine ITS-based studies are also known to neglect certain cultures (Callaghan et al. 2015). These observations, therefore, reaffirm the need to look into vastly unexplored and unreported diversity of anaerobic fungi.
Many of the anaerobic fungal genera like Orpinomyces, Caecomyces, Neocallimastix, Piromyces etc. have a ubiquitous occurrence and can be found in diverse animal species, while some are reportedly host-specific (Paul et al. 2018). The genus Oontomyces has been reported to be a camel-specific genus (Dagar et al. 2015) and some uncultured genus designates have been reported to be specific to hosts like Somali ass, wallaby or American bison (Paul et al. 2018). Similarly, the presence of recently described genera Pecoramyces and Feramyces has been found limited to foregut fermenters and wild undomesticated animals, respectively (Hanafy et al. 2018). However, it is interesting to note that many genera remain uncultured even from the routinely sampled domesticated herbivores, a fact which needs further attention. In this paper, we report the isolation of a novel anaerobic fungus Liebetanzomycespolymorphus, isolated from goat rumen after prolonged incubation of 15 days.
Materials and methods
Sampling, cultivation and preservation
The rumen digesta samples were collected from goats (n=3) slaughtered at Kondhwa slaughterhouse, Pune (India). The samples were immediately brought to the laboratory (within 1 h), pooled and homogenised for 10 minutes under the gas phase of CO2 before making dilutions (up to 10-4) in the anaerobic diluent (McSweeney et al. 2005). The serum roll bottle method (Miller and Wolin 1974) was used to isolate pure cultures of anaerobic fungi from different dilutions. Briefly, the inoculum (0.5 ml) was added to 125 ml glass serum bottles containing 10 ml fungal culture medium (pH 6.8 ± 0.1), which comprised (per litre) of 3 g yeast extract, 5 g tryptone, 150 ml each of solution 1 (0.3% K2HPO4), solution 2 [0.3% KH2PO4, 0.6% (NH4)2SO4, 0.6% NaCl, 0.06% MgSO4.7H2O and 0.06% CaCl2.2H2O] and clarified rumen fluid, 1 ml resazurin (0.1%), 1 ml hemin (0.05%), 6 g NaHCO3, 1 g L-cysteine-HCl, 5 g cellobiose and 20 g agar. The antibiotics, benzylpenicillin and streptomycin sulphate (final concentration 200 µg/ml), were also added to inhibit bacterial growth.
All the roll bottles were incubated at 39±1 °C for 3 weeks and inspected regularly for the development of fungal colonies. The morphologically distinct colonies were picked under anaerobic conditions and inoculated into the fresh liquid culture medium. The serum roll bottle method was repeated two more times to get the axenic fungal cultures. All the cultures were cryopreserved at -80 °C and -196 °C for short-term and long-term storage, respectively using ethylene glycol (final concentration 0.64 M) as the cryoprotectant (Callaghan et al. 2015).
Morphological characterisation
The colony morphology of the cultures was measured after 3 d growth on cellobiose roll bottles, using a stereomicroscope (Leica M205 FA) equipped with a digital camera (Leica DFC450 C). For documenting the microscopic features, the cultures were grown on different carbon sources like rice straw, wheat straw, cellulose, xylan, starch, cellobiose, lactose, maltose, sucrose, glucose, xylose and fructose for 3 d. The microscopic features like thallus morphology, shape and size of sporangia, zoospore shape and flagellation etc. were documented using a differential interference contrast (DIC) microscope (Olympus BX53) equipped with a digital camera (Olympus DP 73) and scanning electron microscope (Carl Zeiss EVO MA15), respectively. The samples for scanning electron microscopy were prepared as described by Ho et al. (1988). To determine the monocentric/polycentric growth patterns, nuclei positions were visualised following straining with bisbenzimide (Fliegerova et al. 2002) or DAPI (Callaghan et al. 2015) using a fluorescence microscope (Nikon Eclipse 80i) equipped with a monochrome digital camera (Media Cybernetics) or confocal microscope (Leica TCS SP8), respectively. All images were processed in GIMP (version 2.8.14) and then compiled using Inkscape (version 0.91) software.
Molecular characterisation and phylogenetic analysis
For molecular characterisation, the genomic DNA was extracted using the CTAB DNA extraction protocol (Sirohi et al. 2013). The complete internal transcribed spacer (ITS1-5.8S-ITS2; ITS) and D1/D2 domain of large-subunit (LSU) ribosomal DNA were amplified using ITS1/ITS4 and NL1/NL4 primer pairs (Edwards et al. 2017). The sequencing was outsourced to 1st BASE (Singapore) and the obtained sequences were compiled and edited manually using BioEdit software (Hall 1999). Since most of the culture-independent studies have generated only ITS1 sequences, the ITS sequence was trimmed to obtain only the partial ITS1 region. Sequence similarity searches were performed using GenBank BLASTn. For phylogenetic analyses, the ITS1 and LSU sequences representing different anaerobic fungal genera and uncultured representatives, were downloaded from the NCBI GenBank database. All the sequences were aligned using the ClustalW programme (Thompson et al. 1994) with default settings in MEGA7 (Kumar et al. 2016). The aligned sequences were used to construct a phylogenetic tree in MEGA7 using the maximum-likelihood method based on the Tamura-Nei model (Tamura and Nei 1993) and tested by 500 bootstrap replications. The genus Gromochytriummamkaevae was used as the outgroup for both ITS1 (accession number: KF586842) as well as LSU (accession number: NR_132054) based trees. The ITS1 and LSU alignments have been submitted to TreeBASE under submission ID 22988.
Substrate utilisation, enzyme activities and fermentation product analyses
The obtained strains were grown in a fungal culture medium without yeast extract or tryptone and cellobiose was replaced by different substrates (Table 1) as a carbon source (Hanafy et al. 2017; Hanafy et al. 2018). Following initial growth for 2 d on monosaccharides and disaccharides and 5 d on polysaccharides, the cultures were subcultured three times at 10% inoculum size to evaluate their substrate utilisation abilities. The growth was measured in terms of visible biomass accumulation and total gas production. The cultures were scored based on their ability to grow luxuriantly (++) or slowly (+) within the stipulated incubation periods or following extended incubation up to 10 days. The avicelase, CMCase, xylanase and β-glucosidase activities of type strain were also determined following its growth on cellulose, xylan, wheat straw and rice straw for 5 d as per the method described previously (Dagar et al. 2018). The fermentation gases (H2, CO2), volatile fatty acids (VFAs; acetate, propionate, butyrate etc.) and alcohols (ethanol and butanol) of growth positive substrates were determined using gas chromatographs equipped with flame ionisation or thermal conductivity detectors, similar to previous reports (Dighe et al. 1998; Kamalaskar et al. 2010; Singh et al. 2016). The organic acids (formate, lactate, succinate, malate etc.) were analysed using HPLC LC20A (Shimadzu, Japan) equipped with a refractive index detector (Thakker et al. 2006). An ordination of the obtained fermentation products was also generated to see if there is any pattern of substrate utilisation using Non-metric Multidimensional Scaling (NMDS) analysis based on the distance matrix in PAST 3.20 software (Hammer et al. 2001).
Table 1.
Substrate utilisation pattern of Liebetanzomycespolymorphus strains G1SC and G6SC compared with other genera of monocentric and uniflagellate filamentous anaerobic fungi.
| Substrate | Lp | Am | Pc | Pr | |
| Polysaccharide | Rice straw | ++ | ND | ND | ND |
| Wheat straw | ++ | ND | ND | ND | |
| Cellulose | ++ | ++ | ++ | ++ | |
| Xylan | ++ | ++ | ++ | ++ | |
| Starch | ++ | ++ | ++ | ++ | |
| Inulin | – | – | – | ++ | |
| Raffinose | – | – | ++ | ++ | |
| Chitin | – | ND | ND | – | |
| Alginate | – | ND | ND | – | |
| Pectin | ++ | – | – | – | |
| Disaccharide | Cellobiose | ++ | ++ | ++ | ++ |
| Sucrose | + | ++ | ++ | ++ | |
| Maltose | ++ | ++ | ++ | ++ | |
| Trehalose | – | ND | ND | + | |
| Lactose | + | + | ++ | – | |
| Monosaccharide | Glucose | ++ | ++ | ++ | ++ |
| Xylose | ++ | ++ | ++ | ++ | |
| Mannose | – | – | – | ++ | |
| Fructose | ++ | ++ | ++ | ++ | |
| Arabinose | – | – | – | – | |
| Ribose | – | ND | – | – | |
| Glucuronic acid | – | ND | ND | – | |
| Galactose | – | – | – | – | |
| Peptide | Peptone | – | ND | ND | – |
| Tryptone | – | ND | ND | – | |
Strains: LpLiebetanzomycespolymorphus strain G1SC and G6SC AmAnaeromycesmucronatus strain BF2 (Breton et al. 1990) PcPiromycescommunis strain SM5 (Trinci et al. 1994) PrPecoramycesruminantium strain C1A and S4B (Hanafy et al. 2017). – (negative), ++ (positive), + (weak growth/ slow growth/ growth visible after 4–10 d of incubation), ND (not documented)
Results and discussion
Growth and morphological characterisation
For the routine anaerobic fungal isolation, the inoculated roll tubes are usually incubated for 2–4 d after which the developed colonies are picked anaerobically and pure cultures are obtained. However, we chose to incubate the roll tubes for an extended period of time i.e. up to 3 weeks. The decision for extended incubation was taken in the wake of the fact that several genera of anaerobic fungi remain to be uncultured and incubation time may be one of the limiting factors. As shorter incubation times favours fast-growing cultures, some slow growing cultures might be omitted and which may grow after prolonged incubation. The prolonged incubation might also help some cultures to cope better in stressed conditions of different growth environment, the presence of antibiotics or some oxygen exposure during sample collection or processing. In a previous study, prolonged incubation was recommended for the isolation of anaerobic bacteria from clinical specimens and correct bacteriological diagnosis (Wren 1980). Janssen et al. (2002) also speculated that the extended incubation period may help isolation of previously uncultured groups of soil bacteria. In this case, the extended incubation of 15 d resulted in the development of two fungal colonies, but no further growth even after 21 d of incubation. Following isolation and establishment of pure cultures, the morphological, molecular and substrate utilisation characteristics of both strains, namely G1SC and G6SC were found to be identical. Therefore, one of them, i.e. strain G1SC, was denoted as the type strain and used for detailed characterisation. The type strain has been deposited to the MACS collection of microorganisms (MCM), Agharkar Research Institute, Pune, India under the accession number MCMB-1469.
The colony morphology of strain G1SC is shown in Fig. 1A–C, showing 1–2 mm sized colony, attached to rice straw (Fig. 1A) and at different stages of growth. A large number of newly formed sporangia at the periphery of the colony were observed, as shown by numerous dot-like structures of varying sizes (Fig. 1B–C). A similar observation was also made in broth culture, where numerous fungal thalli, attached to the bottom of glass bottles, were seen on the initial period of growth (Suppl. material 1: Figure S1), which later developed into a thin mat or biofilm-like structure (Fig. 1D) similar to Pecoramyces (Hanafy et al. 2017). Zoospores were abundantly produced, mostly uniflagellate (Fig. 1E–F) and rarely biflagellate (Fig. 1G), spherical to ovoid in size (5–6 µm in diameter) and the flagellum 15–20 µm in length. The zoospores were found to germinate either endogenously or exogenously (Fig. 1H–I) into different shapes of sporangia (Fig. 1J) like globose or ellipsoidal at the very early stages of development. Different forms of rhizoidal development, like long single rhizoid (Fig. 1K), two rhizoids (Fig. 1L) and even multiple rhizoids originating from sporangia (Fig. 1M), were also seen.
Figure 1.

Macroscopic and microscopic features of Liebetanzomycespolymorphus. Colony morphology on agar roll tubes (A–C), showing the development of a colony attached (A) to a straw particle (arrowed), dense growth in the centre surrounded by numerous sporangia and zoospores (B–C) causing expansion of colony size. Growth in liquid medium showing a biofilm-like growth (D). Zoospores are spherical and Uniflagellate (E–F) or biflagellate (G). Germinating zoospore (H) showing a zoospore cyst (arrowed), presence and absence of sporangiophore indicating the endogenous and exogenous type of sporangial development (I) and different shapes of sporangia (I, J). Early stages of thallus development showing a single (K), bifurcated (L) and multifurcated (M) rhizoidal system. Scale bar: 1 mm (A–C); 10 µM (E–M).
We also noticed pleomorphism in sporangial and rhizoidal structures of strain G1SC on different substrates like rice straw (Fig. 2A), wheat straw (Fig. 2B), cellulose (Fig. 2C), xylan (Fig. 2D), starch (Fig. 2E), cellobiose (Fig. 2F), lactose (Fig. 2G), maltose (Fig. 2H), sucrose (Fig. 2I), glucose (Fig. 2J), xylose (Fig. 2K) and fructose (Fig. 2L). In the case of complex substrates like rice straw, wheat straw, cellulose and xylan, the rhizoidal growth was observed to be more extensively branched. Conversely, thicker and less branched rhizoidal growth was noticed on dimeric and monomeric substrates, probably due to the ready availability of fermentable sugars. It was also observed that, on rice and wheat straw, an exogenous type of sporangia development with short or long sporangiophores was more prominent in comparison to endogenous sporangia on all other substrates. The presence of sporangiophores on complex straw particles might be helpful for sporangia to come out of straw and release zoospores to further areas for faster colonisation. The pleomorphism, in morphological features with a change in culture conditions, particularly with carbon source, is also well described earlier (Brookman et al. 2000; Gruninger et al. 2014). However, the sporangial shapes varied, not only on different carbon sources, but also on the same carbon source (Fig. 3). Different sporangial shapes like globose, ellipsoid, clavate, ovoid, with or without a papilla were noticed on the same substrate i.e. rice straw as shown in Fig. 3A–F. Similar polymorphism was also observed on several other substrates, e.g. xylose (Fig. 3G–I) and cellobiose (Fig. 3J–L), where conspicuous irregular sporangial structures were seen. These findings, therefore, highlight the pleomorphic nature of strain G1SC, irrespective of the carbon sources used in the growth medium.
Figure 2.

Microscopic features of Liebetanzomycespolymorphus showing pleomorphism in sporangial and rhizoidal structures, when grown on different carbon sources like rice straw (A), wheat straw (B), cellulose (C), xylan (D), starch (E), cellobiose (F), lactose (G), maltose (H), sucrose (I), glucose (J), xylose (K) and fructose (L). The exogenous sporangia appearing on a sporangiophore (A–B) and endogenous sporangia of different shapes (C–L) are shown. Scale bar: 20 µM.
Figure 3.

Microscopic images of Liebetanzomycespolymorphus. On rice straw (A–F) sporangia of varying sizes and shapes, like elongate (A), ellipsoid (B), ovoid (C), clavate (E) and globose (F). The zoospore cyst (A–C arrowed) is also visible, highlighting bipolar germination. Sporangiophore of varying length and shape, from short to long (A), sometimes eggcup shaped (C) and constricted (E arrowed) is shown. The sporangia with a papilla (D arrowed) and septum (B, C, D, E starred) are also indicated. Sporangia and thalli of irregular morphology on xylose (G–I) and cellobiose (J–L), including some pseudo-intercalary sporangia (J–L) can be observed. Scale bar: 20 µM.
The sporangial size also varied in diameter (10–90 µm wide, 10–75 µm long), always borne at the terminal end of a variable length sporangiophore (15–80 µm), in the case of exogenous development. Several cyst-like structures were also visible on the sporangiophore (Fig. 3A–C) highlighting bipolar germination of zoospores. In some cases, an eggcup shaped sporangiophore at the bottom of sporangium (Fig. 3C) and a constricted sporangiophore were also noted (Fig. 3E). In the case of soluble substrates, few pseudo-intercalary sporangia, similar to Oontomyces and Feramyces (Dagar et al. 2015; Hanafy et al. 2018), were also noted (Fig. 3J–L). The thallus development was clearly monocentric as evident by the mature sporangia full of zoospores (Fig. 4A–D, I), anucleated rhizoids (Fig. 4E–I) and a single thallus having a single sporangium (Fig. 4J–K). All these morphological features were similar to various Piromyces spp., Buwchfawromyces, Oontomyces and Pecoramyces, which also have the monocentric thallus and extensive variations in sporangial shape and size and produce uniflagellate zoospores (Ho and Barr 1995). Similar to P.rhizinflatus, the sporangia of strain G1SC showed a constricted, isthmus-like neck (Fig. 2C, J), sporangiophore appearing like an eggcup (Fig. 3C) like in P.communis and sporangium with a papilla (Fig. 3D) similar to P.mae (Ho and Barr 1995; Li et al. 1990).
Figure 4.

Phase contrast (A, C, E, G), fluorescence (B, D, F, H) and confocal (I) microscopic images of Liebetanzomycespolymorphus showing elongate (A–B) and triangular (C–D) sporangium filled with zoospores. Nuclei were seen in sporangium but not in sporangiophore (B) or rhizoidal system (F, H, I). No constrictions were observed in rhizoids. Scanning electron microscopy images of elongate (J) and globose (K) sporangium showing monocentric thallus. Scale bar: 20 µM (A–I); 10 µM (J–K).
The similarities in morphological features of monocentric and uniflagellate genera make it very difficult to identify and differentiate this group of anaerobic fungi. It is also interesting to note that, so far, the maximum number of species has been described in the Piromyces genus and most of the newly described genera of anaerobic fungi share morphological similarities with these different species, including strain G1SC. These observations point towards the possibility that some of these newly described genera might have been isolated previously as well but were identified as different species of Piromyces. It is also worth noting that all newly described genera have primarily been described using molecular tools, thus emphasising the need to use these modern tools.
Molecular characterisation and phylogenetic analysis
The successful amplification of ITS and LSU regions yielded product sizes of ca. 700 bp and 750 bp, respectively. The obtained sequences were submitted to NCBI GenBank to obtain the accession numbers for strain G1SC (MH468765 and MH468763; ITS and LSU) and G6SC (MH468766 and MH468764; ITS and LSU). The ITS region based sequence similarity search results showed that strain G1SC was 88.14% and 87.03% similar to A.robustus (accession number: NR 148182) and A.contortus (accession number: MG605706), respectively. Likewise, the LSU region of strain G1SC was most closely similar (97.71%) to A.contortus (accession number: MG605690). Although the sequence similarity of ca. 97% usually relates to a novel culture at the species level with its nearest match, the stark morphological dissimilarities of strain G1SC with A.contortus led to its classification as a novel genus.
To better understand the ecological distribution of strain G1SC, only ITS1 based searches were performed, which indicated <96% sequence similarities with several uncultured representatives of phylum Neocallimastigomycota (Suppl. material 2: Figure S2) reported from the USA (Liggenstoffer et al. 2010). The nearest matches were with different clones obtained from cattle (93–96%) and other clones of cattle, sheep and llama (88%-92%), indicating the presence of members of the G1SC clade in cattle and close relatives in sheep and llama. The nearest matches of G1SC clade were also found in the SP4 clade designated by Paul et al. (2018), represented by accession number GQ767184, which consisted of 120 sequences of uncultured fungi, all from cattle faeces. The results, hence, indicate the presence of related fungi in both large as well as small ruminants. Surprisingly, no similar sequences were obtained from goat, which might be due to a lesser number of culture-independent studies conducted on these animals. The phylogenetic analysis of LSU and ITS1 regions showed distinct lineage for the G1SC clade, supported by high bootstrap values (Fig. 5 and Fig. 6), nearest to the Anaeromyces cluster. The results, therefore, underscore the importance of morphology-based identifications along with molecular tools, as none on its own can be relied on for accurate identification of anaerobic fungi.
Figure 5.

The LSU based maximum-likelihood tree is showing the phylogenetic position of Liebetanzomycespolymorphus with other members of phylum Neocallimastigomycota. Bootstrap values (>50%) based on 500 replicates are indicated at branching points. The GenBank accession number of each strain is listed in parentheses. Scale bar: 0.1 substitutions per site.
Figure 6.

The ITS1 based maximum-likelihood tree is showing the phylogenetic position of Liebetanzomycespolymorphus with nearest uncultured clones and other members of phylum Neocallimastigomycota. Bootstrap values (>50%) based on 500 replicates are indicated at branching points. The GenBank accession number of each strain is listed in parentheses. Scale bar: 0.5 substitutions per site.
Substrate utilisation, enzyme activities and fermentation product analyses
The substrate utilisation profiles of strains G1SC and G6SC exhibited exactly identical patterns (Table 1). Both these strains could utilise various polymeric and monomeric substrates. Our results were also compared to substrate utilisation profiles of other genera of monocentric and uniflagellate filamentous anaerobic fungi. All these genera were found to utilise cellulose, xylan, starch, cellobiose, sucrose, maltose, glucose and fructose, while none utilised chitin, ribose, peptone and tryptone as the sole carbon and energy source. For inulin, raffinose, alginate, pectin, trehalose, mannose, arabinose, glucuronic acid and galactose, varying results were obtained. Interestingly, only strains G1SC and G6SC were found positive for pectin utilisation amongst compared genera.
The enzyme activities (µmol/ml/h) of type strain G1SC showed maximum enzyme activities on crude substrates like rice straw and wheat straw, instead of pure cellulose and xylan (Table 2). The highest avicelase (1.05 ± 0.19) and xylanase (88.33 ± 1.00) activities were obtained on rice straw, while highest CMCase (7.73 ± 0.31) and β-glucosidase (1.64 ± 0.08) activities were measured in wheat straw. When compared to the enzyme activities of Neocallimastix, Piromyces, Orpinomyces and Anaeromyces, grown on wheat straw (Dagar et al. 2018), the strain G1SC displayed poor β-glucosidase activities. The avicelase activities of strain G1SC were comparable to Orpinomyces and Anaeromyces, but lower than most strains of Neocallimastix and Piromyces. The CMCase and xylanase activities of strain G1SC were comparable to Orpinomyces, Neocallimastix and most strains of Piromyces, but higher than Anaeromyces. These results suggest that strain G1SC is a moderate lignocellulose degrader and can be of use in applications involving lignocellulose degradation.
Table 2.
Enzyme activities (µmol/mL/h) of Liebetanzomycespolymorphus strain G1SC on different substrates at 5 d of incubation.
| Enzyme activity | Substrate used | |||
| Rice straw | Wheat straw | Cellulose | Xylan | |
| Avicelase | 1.05 ± 0.19 | 0.68 ± 0.10 | 0.73 ± 0.06 | – |
| CMCase | 4.55 ± 0.14 | 7.73 ± 0.31 | 5.56 ± 0.98 | – |
| Xylanase | 88.33 ± 1.00 | 79.44 ± 0.11 | – | 54.69 ± 2.31 |
| Β-glucosidase | 1.53 ± 0.07 | 1.64 ± 0.08 | 0.69 ± 0.02 | |
– ±: standard deviation of three replicates.
The substrate utilisation results were also confirmed by measuring the fermentation products on each substrate (Table 3). The strain G1SC produced hydrogen, carbon dioxide, formate, acetate, lactate, succinate and ethanol on all substrates, but did not produce propionate, butyrate, butanol or malate which is in line with the previous reports (Trinci et al. 1994; Edwards et al. 2017). The absence of propionate and butyrate also implies that there was no bacterial contamination and all products are solely of fungal origin. The fermentation product quantities also indicated the efficiency of substrate utilisation. The substrates like sucrose and lactose, which recorded poor growth after 2 d of incubation, also showed lesser production of formate, acetate and lactate and the absence of succinate. Likewise, the polymeric substrate like rice straw, wheat straw, cellulose and xylan showed lesser production of hydrogen, formate, acetate, lactate, succinate and ethanol at 2 d of incubation (Table 3), which increased substantially after 5 d (Suppl. material 4: Table S1). The NMDS plot showed clear clustering of various carbon sources based on fermentation products, an observation duly supported by a low stress value (Suppl. material 3: Figure S3). These results, therefore, signify the usefulness of metabolite profiling for measuring the substrate utilisation abilities of anaerobic fungi.
Table 3.
Fermentation products of Liebetanzomycespolymorphus strain G1SC on different substrates after 2 d of incubation.
| Substrate | Fermentation product (in mM) | ||||||
| Hydrogen | Carbon dioxide | Formate | Acetate | Lactate | Succinate | Ethanol | |
| Rice straw | 58.10 ± 3.57 | 4.82 ± 0.16 | 16.43 ± 0.04 | 26.07 ± 0.24 | 1.79 ± 0.04 | 0.29 ± 0.00 | 10.35 ± 0.05 |
| Wheat straw | 45.42 ± 2.98 | 5.46 ± 0.13 | 11.44 ± 0.07 | 19.77 ± 0.44 | 1.72 ± 0.04 | 0.24 ± 0.00 | 8.27 ± 0.13 |
| Cellulose | 26.86 ± 3.11 | 6.38 ± 0.05 | 4.61 ± 0.05 | 13.26 ± 0.52 | 0.73 ± 0.02 | ND | 7.27 ± 0.01 |
| Xylan | 27.71 ± 2.48 | 7.12 ± 0.11 | 5.01 ± 0.06 | 8.30 ± 0.12 | 4.01 ± 0.06 | 0.12 ± 0.00 | 6.96 ± 0.03 |
| Starch | 44.09 ± 0.54 | 5.99 ± 0.03 | 9.45 ± 0.10 | 14.58 ± 0.28 | 15.11 ± 0.38 | 0.34 ± 0.00 | 7.69 ± 0.13 |
| Pectin | 42.59 ± 1.18 | 6.38 ± 0.09 | 7.64 ± 0.08 | 11.32 ± 0.22 | 3.56 ± 0.04 | 0.15 ± 0.01 | 8.01 ± 0.12 |
| Cellobiose | 58.05 ± 0.63 | 5.64 ± 0.03 | 9.75 ± 0.02 | 15.69 ± 0.29 | 9.35 ± 0.04 | 0.30 ± 0.01 | 8.46 ± 0.04 |
| Sucrose | 29.21 ± 1.02 | 6.99 ± 0.05 | 3.77 ± 0.10 | 7.61 ± 0.44 | 0.66 ± 0.00 | ND | 9.11 ± 0.06 |
| Maltose | 53.26 ± 3.01 | 5.59 ± 0.12 | 8.26 ± 0.13 | 17.45 ± 0.40 | 10.16 ± 0.06 | 0.22 ± 0.00 | 7.85 ± 0.05 |
| Lactose | 39.08 ± 1.83 | 6.51 ± 0.08 | 4.26 ± 0.01 | 8.98 ± 0.13 | 0.47 ± 0.01 | ND | 8.52 ± 0.04 |
| Glucose | 49.79 ± 0.86 | 5.84 ± 0.04 | 10.11 ± 0.18 | 16.75 ± 0.19 | 7.18 ± 0.02 | 0.31 ± 0.00 | 8.65 ± 0.09 |
| Xylose | 53.80 ± 1.41 | 5.62 ± 0.09 | 8.38 ± 0.19 | 12.58 ± 0.47 | 7.90 ± 0.06 | 0.16 ± 0.00 | 8.39 ± 0.03 |
| Fructose | 61.05 ± 1.59 | 5.28 ± 0.06 | 11.79 ± 0.05 | 17.05 ± 0.71 | 15.97 ± 0.03 | 0.30 ± 0.00 | 9.10 ± 0.05 |
– ±: standard deviation of three replicates.
Liebetanzomyces polymorphus
Joshi, G.W. Griff. & Dagar, gen. et sp. nov.
Diagnosis.
Strictly anaerobic fungus with determinate, monocentric thallus with single terminal sporangium of varying shape and size and uniflagellate zoospores. The clade is defined by the sequences accession numbers MH468765 (ITS1, 5.8S, ITS2 complete) and MH468763 (LSU, partial sequence). The most genetically similar genus is Anaeromyces, which is defined as forming a polycentric thallus (Breton et al. 1990.FEMS Microbiol. Lett. 58, p.177), in contrast to the monocentric Liebetanzomyces.
A strict anaerobic fungus, isolated from the rumen of a goat. In roll tubes of cellobiose agar medium, the fungus forms medium to large circular colonies (0.5–2 mm diam.). In liquid cellobiose medium, the fungal thalli attach to the sides and bottom of glass bottles. Monocentric thallus development, producing a single terminal sporangium per thallus. Also, the fungus produces few pseudo-intercalary sporangia but only on soluble substrates. An extensive anucleate rhizoidal system without constrictions is formed. The sporangia vary in size (10–90 µm wide, 10–75 µm long), shape (globose, ellipsoid, clavate, ovoid, elongate or irregular) and sometimes bear papillae. The sporangium is borne on variable length sporangiophore (15–80 µm), sometimes forming an eggcup-like structure below the sporangium or showing cyst-like structure. Zoospores are produced abundantly, mostly uniflagellate, rarely biflagellate, spherical (5–6 µm in diameter) in size and flagellum of 15–20 µm in length. The zoospores may germinate either endogenously or exogenously. The clade is defined by the sequences accession numbers MH468765 (ITS1, 5.8S, ITS2 complete) and MH468763 (LSU, partial sequence). The ex-type culture (strain G1SC) is stored cryogenically in liquid nitrogen at bioenergy group, Agharkar Research Institute, Pune, India. The holotype is a 3 d old culture of G1SC preserved in 5% glutaraldehyde and deposited at the MACS-collection of microorganisms (MCM) of Agharkar Research Institute, Pune, India and isotype material at the Aberystwyth University biorepository (code ABS).
Etymology.
‘Liebetanz’ is assigned to honour Erwin Liebetanz (Liebetanz 1910) by taking all nine letters of his surname (i.e. LIEBETANZ) and who was the first to observe anaerobic gut fungus Piromyces as a flagellated organism in 1910; myces = the Greek name for fungus.
The specific epithet polymorphus is for different polymorphic sporangial shapes displayed by this fungus.
Conclusions
The morphological and molecular characterisation results clearly indicate that the strain G1SC represents a novel genus Liebetanzomycespolymorphus in the phylum Neocallimastigomycota. Liebetanzomycespolymorphus displays a monocentric thallus and produces uniflagellate zoospores, thus, it is morphologically similar to Piromyces, Buwchfawromyces, Oontomyces and Pecoramyces. Liebetanzomycespolymorphus exhibits extensive sporangial variations and is genetically near but dissimilar to Anaeromyces. The ITS1 and LSU based phylogenetic analysis also confirmed the distinct lineage of Liebetanzomycespolymorphus. The results suggest that both morphological and molecular tools should be used in tandem to determine the uniqueness of any anaerobic fungal culture, as the use of either one independently can result in a wrong identification.
Supplementary Material
Acknowledgements
The authors acknowledge the support of Department of Biotechnology (DBT) for the financial support (project no. BT/PR15694/PBD/26/506/2015). We are grateful to Dr Karthick Balasubramanian for his valuable help in the suitable presentation of results and Dr Rajesh Kumar K.C. for helpful discussions. We also thank the Director, Agharkar Research Institute for providing the required infrastructure support.
Citation
Joshi A, Lanjekar VB, Dhakephalkar PK, Callaghan TM, Griffith GW, Dagar SS (2018) Liebetanzomyces polymorphus gen. et sp. nov., a new anaerobic fungus (Neocallimastigomycota) isolated from the rumen of a goat. MycoKeys 40: 89–110. https://doi.org/10.3897/mycokeys.40.28337
Supplementary materials
Figure S1
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: multimedia
Explanation note: Photographs showing fungal thalli (marked by arrows) of Liebetanzomyces polymorphus attached to the sides (A) and bottom (B) of glass bottles when grown in liquid medium.
{kind=link}
Figure S2
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: multimedia
Explanation note: The ITS1 based maximum-likelihood tree based on the Tamura 3-parameter model (Tamura 1992) is showing the phylogenetic position of Liebetanzomyces polymorphus with nearest uncultured clones. The genus Anaeromyces mucronatus (accession number: NR_111156) was used as the outgroup. Bootstrap values (>50%) based on 500 replicates are indicated at branching points. The GenBank accession number of each strain is listed before the name. Bar, 0.02 substitutions per site.
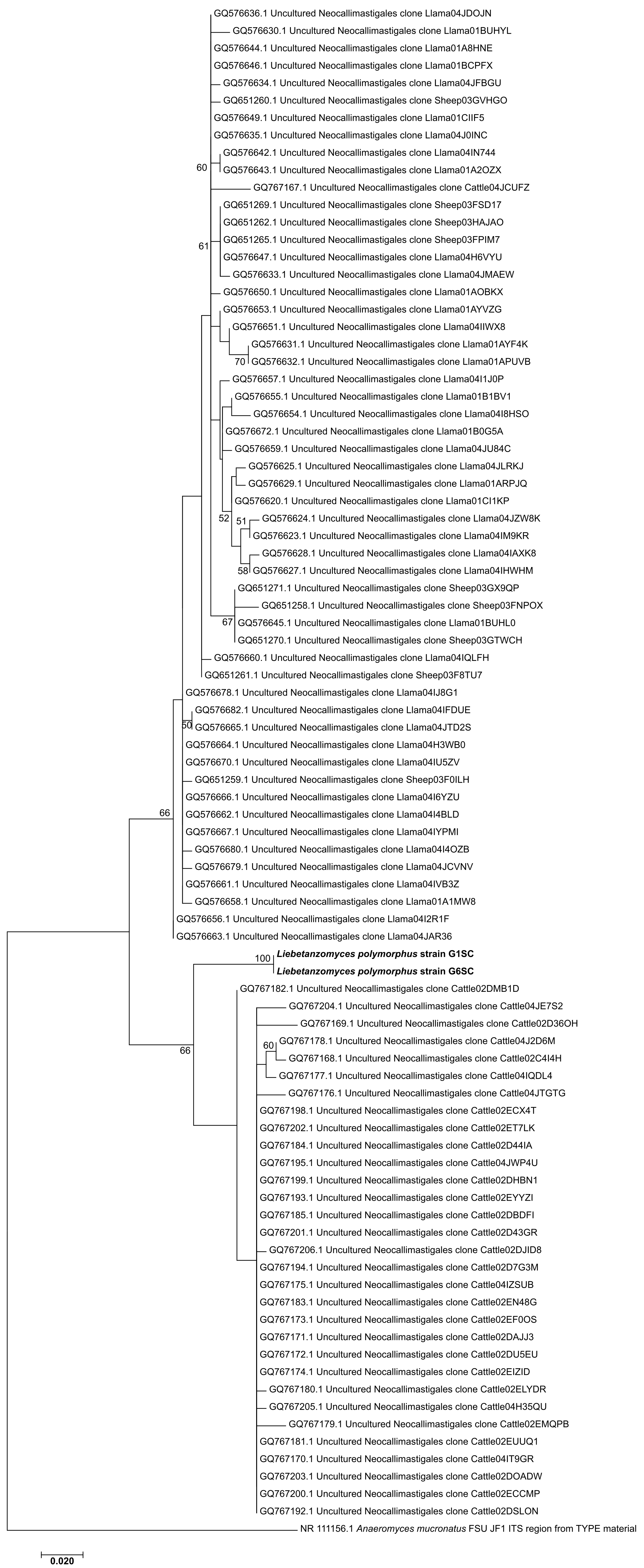{kind=link}
Figure S3
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: multimedia
Explanation note: The Non-metric multidimensional scaling (NMDS) plot showing clustering of various carbon sources based on fermentation products. Stress value, 0.012.
{kind=link}
Table S1
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: species data
Explanation note: Fermentation products of Liebetanzomyces polymorphus strain G1SC on different substrates after 5 d of incubation.
Reference
- Breton A, Bernalier A, Dusser M, Fonty G, Gaillard-Martinie B, Guillot J. (1990) Anaeromycesmucronatus nov. gen., nov. sp. A new strictly anaerobic rumen fungus with polycentric thallus. FEMS microbiology letters 70: 177–182. 10.1111/j.1574-6968.1990.tb13974.x [DOI] [PubMed] [Google Scholar]
- Brookman J, Mennim G, Trinci A, Theodorou M, Tuckwell D. (2000) Identification and characterization of anaerobic gut fungi using molecular methodologies based on ribosomal ITS1 and 18S rRNA. Microbiology 146: 393–403. 10.1099/00221287-146-2-393 [DOI] [PubMed] [Google Scholar]
- Callaghan TM, Podmirseg SM, Hohlweck D, Edwards JE, Puniya AK, Dagar SS, Griffith GW. (2015) Buwchfawromyceseastonii gen. nov., sp. nov.: a new anaerobic fungus (Neocallimastigomycota) isolated from buffalo faeces. MycoKeys 9. 10.3897/mycokeys.9.9032 [DOI]
- Dagar SS, Kumar S, Griffith GW, Edwards JE, Callaghan TM, Singh R, Nagpal AK, Puniya AK. (2015) A new anaerobic fungus (Oontomycesanksri gen. nov., sp. nov.) from the digestive tract of the Indian camel (Camelusdromedarius). Fungal Biology 119: 731–737. 10.1016/j.funbio.2015.04.005 [DOI] [PubMed] [Google Scholar]
- Dagar SS, Kumar S, Mudgil P, Puniya AK. (2018) Comparative evaluation of lignocellulolytic activities of filamentous cultures of monocentric and polycentric anaerobic fungi. Anaerobe 50: 76–79. 10.1016/j.anaerobe.2018.02.004 [DOI] [PubMed] [Google Scholar]
- Dighe AS, Shouche YS, Ranade DR. (1998) Selenomonaslipolytica sp. nov., an obligately anaerobic bacterium possessing lipolytic activity. International Journal of Systematic and Evolutionary Microbiology 48: 783–791. 10.1099/00207713-48-3-783 [DOI] [PubMed] [Google Scholar]
- Edwards JE, Forster RJ, Callaghan TM, Dollhofer V, Dagar SS, Cheng Y, Chang J, Kittelmann S, Fliegerova K, Puniya AK, Henske JK, Gilmore SP, O’Malley MA, Griffith GW, Smidt H. (2017) PCR and omics based techniques to study the diversity, ecology and biology of anaerobic fungi: insights, challenges and opportunities. Frontiers in Microbiology 8: 1657. 10.3389/fmicb.2017.01657 [DOI] [PMC free article] [PubMed]
- Fliegerova K, Pažoutová S, Mrazek J, Kopečný J. (2002) Special properties of polycentric anaerobic fungus Anaeromycesmucronatus. Acta Veterinaria Brunensis 71: 441–444. 10.2754/avb200271040441 [DOI] [Google Scholar]
- Gruninger RJ, Puniya AK, Callaghan TM, Edwards JE, Youssef N, Dagar SS, Fliegerova K, Griffith GW, Forster R, Tsang A, McAllister T, Elshahed MS. (2014) Anaerobic fungi (phylum Neocallimastigomycota): advances in understanding their taxonomy, life cycle, ecology, role and biotechnological potential. FEMS Microbiology Ecology 90: 1–17. 10.1111/1574-6941.12383 [DOI] [PubMed] [Google Scholar]
- Hall TA. (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic acids symposium series. [London]: Information Retrieval Ltd., c1979–c2000., 95–98.
- Hammer Ø, Harper DAT, Ryan PD. (2001) PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4: 1–9. [Google Scholar]
- Hanafy RA, Elshahed MS, Liggenstoffer AS, Griffith GW, Youssef NH. (2017) Pecoramycesruminantium, gen. nov., sp. nov., an anaerobic gut fungus from the feces of cattle and sheep. Mycologia 109: 231–243. 10.1080/00275514.2017.1317190 [DOI] [PubMed] [Google Scholar]
- Hanafy RA, Elshahed MS, Youssef NH. (2018) Feramycesaustinii, gen. nov, sp. nov., an anaerobic gut fungus from rumen and fecal samples of wild Barbary sheep and fallow deer. Mycologia. 10.1080/00275514.2018.1466610 [DOI] [PubMed]
- Hibbett DS, Binder M, Bischoff JF, Blackwell M, Cannon PF, Eriksson OE, Huhndorf S, James T, Kirk PM, Lucking R, Thorsten Lumbsch H, Lutzoni F, Matheny PB, McLaughlin DJ, Powell MJ, Redhead S, Schoch CL, Spatafora JW, Stalpers JA, Vilgalys R, Aime MC, Aptroot A, Bauer R, Begerow D, Benny GL, Castlebury LA, Crous PW, Dai YC, Gams W, Geiser DM, Griffith GW, Gueidan C, Hawksworth DL, Hestmark G, Hosaka K, Humber RA, Hyde KD, Ironside JE, Koljalg U, Kurtzman CP, Larsson KH, Lichtwardt R, Longcore J, Miadlikowska J, Miller A, Moncalvo JM, Mozley-Standridge S, Oberwinkler F, Parmasto E, Reeb V, Rogers JD, Roux C, Ryvarden L, Sampaio JP, Schussler A, Sugiyama J, Thorn RG, Tibell L, Untereiner WA, Walker C, Wang Z, Weir A, Weiss M, White MM, Winka K, Yao YJ, Zhang N. (2007) A higher-level phylogenetic classification of the Fungi. Mycological Research 111: 509–547. 10.1016/j.mycres.2007.03.004 [DOI] [PubMed] [Google Scholar]
- Ho Y, Abdullah N, Jalaludin S. (1988) Penetrating structures of anaerobic rumen fungi in cattle and swamp buffalo. Microbiology 134: 177–181. 10.1099/00221287-134-1-177 [DOI] [Google Scholar]
- Ho Y, Barr D. (1995) Classification of anaerobic gut fungi from herbivores with emphasis on rumen fungi from Malaysia. Mycologia: 655–677. 10.1080/00275514.1995.12026582 [DOI]
- Ivarsson M, Schnürer A, Bengtson S, Neubeck A. (2016) Anaerobic Fungi: A Potential Source of Biological H(2) in the Oceanic Crust. Frontiers in Microbiology 7: 674. 10.3389/fmicb.2016.00674 [DOI] [PMC free article] [PubMed]
- Janssen PH, Yates PS, Grinton BE, Taylor PM, Sait M. (2002) Improved culturability of soil bacteria and isolation in pure culture of novel members of the divisions Acidobacteria, Actinobacteria, Proteobacteria, and Verrucomicrobia. Applied and Environmental Microbiology 68: 2391–2396. 10.1128/aem.68.5.2391-2396.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kamalaskar LB, Dhakephalkar P, Meher K, Ranade D. (2010) High biohydrogen yielding Clostridium sp. DMHC-10 isolated from sludge of distillery waste treatment plant. International Journal of Hydrogen Energy 35: 10639–10644. 10.1016/j.ijhydene.2010.05.020 [DOI] [Google Scholar]
- Kittelmann S, Naylor GE, Koolaard JP, Janssen PH. (2012) A proposed taxonomy of anaerobic fungi (Class Neocallimastigomycetes) suitable for large-scale sequence-based community structure analysis. PLoS One 7: e36866. 10.1371/journal.pone.0036866 [DOI] [PMC free article] [PubMed]
- Koetschan C, Kittelmann S, Lu J, Al-Halbouni D, Jarvis GN, Müller T, Wolf M, Janssen PH. (2014) Internal transcribed spacer 1 secondary structure analysis reveals a common core throughout the anaerobic fungi (Neocallimastigomycota). PLoS One 9: e91928. 10.1371/journal.pone.0091928 [DOI] [PMC free article] [PubMed]
- Kumar S, Stecher G, Tamura K. (2016) MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution 33: 1870–1874. 10.1093/molbev/msw054 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Heath IB, Bauchop T. (1990) Piromycesmae and Piromycesdumbonica, two new species of uniflagellate anaerobic chytridiomycete fungi from the hindgut of the horse and elephant. Canadian Journal of Botany 68: 1021–1033. 10.1139/b90-129 [DOI] [Google Scholar]
- Liebetanz E. (1910) Die parasitischen Protozoen des Wiederkauermagens. Archiv für Protistenkunde 19: 19–80. [Google Scholar]
- Liggenstoffer AS, Youssef NH, Couger M, Elshahed MS. (2010) Phylogenetic diversity and community structure of anaerobic gut fungi (phylum Neocallimastigomycota) in ruminant and non-ruminant herbivores. The ISME Journal 4: 1225. 10.1038/ismej.2010.49 [DOI] [PubMed]
- McSweeney CS, Denman SE, Mackie RI. (2005) Rumen bacteria. In: Makkar HPS, McSweeney CS. (Eds) Methods in Gut Microbial Ecology for Ruminants.Springer Netherlands, Dordrecht, 23–37. 10.1007/1-4020-3791-0_2 [DOI]
- Miller TL, Wolin MJ. (1974) A Serum bottle modification of the hungate technique for cultivating obligate anaerobes. Applied Microbiology 27: 985–987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paul SS, Bu D, Xu J, Hyde KD, Yu Z. (2018) A phylogenetic census of global diversity of gut anaerobic fungi and a new taxonomic framework. Fungal Diversity 89: 253–266. 10.1007/s13225-018-0396-6 [DOI] [Google Scholar]
- Picard KT. (2017) Coastal marine habitats harbor novel early-diverging fungal diversity. Fungal Ecology 25: 1–13. 10.1016/j.funeco.2016.10.006 [DOI] [Google Scholar]
- Singh KG, Lapsiya KL, Gophane RR, Ranade DR. (2016) Optimization for butanol production using Plackett-Burman Design coupled with Central Composite Design by Clostridiumbeijerenckii strain CHTa isolated from distillery waste manure. International Research Journal of Biochemical Technology 7: 1063–1068. [Google Scholar]
- Sirohi SK, Choudhury PK, Dagar SS, Puniya AK, Singh D. (2013) Isolation, characterization and fibre degradation potential of anaerobic rumen fungi from cattle. Annals of Microbiology 63: 1187–1194. 10.1007/s13213-012-0577-6 [DOI] [Google Scholar]
- Tamura K. (1992) Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G+ C-content biases. Molecular Biology and Evolution 9: 678–687. 10.1093/oxfordjournals.molbev.a040752 [DOI] [PubMed] [Google Scholar]
- Tamura K, Nei M. (1993) Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution 10: 512–526. 10.1093/oxfordjournals.molbev.a040023 [DOI] [PubMed] [Google Scholar]
- Thakker C, Bhosale S, Ranade D. (2006) Formation of succinic acid by Klebsiellapneumoniae MCM B-325 under aerobic and anaerobic conditions. Journal of Microbiology and Biotechnology 16: 870–879. [Google Scholar]
- Thompson JD, Higgins DG, Gibson TJ. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research 22: 4673–4680. 10.1093/nar/22.22.4673 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trinci APJ, Davies DR, Gull K, Lawrence MI, Bonde Nielsen B, Rickers A, Theodorou MK. (1994) Anaerobic fungi in herbivorous animals. Mycological Research 98: 129–152. 10.1016/S0953-7562(09)80178-0 [DOI] [Google Scholar]
- Wren M. (1980) Prolonged primary incubation in the isolation of anaerobic bacteria from clinical specimens. Journal of Medical Microbiology 13: 257–264. 10.1099/00222615-13-2-257 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: multimedia
Explanation note: Photographs showing fungal thalli (marked by arrows) of Liebetanzomyces polymorphus attached to the sides (A) and bottom (B) of glass bottles when grown in liquid medium.
Figure S2
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: multimedia
Explanation note: The ITS1 based maximum-likelihood tree based on the Tamura 3-parameter model (Tamura 1992) is showing the phylogenetic position of Liebetanzomyces polymorphus with nearest uncultured clones. The genus Anaeromyces mucronatus (accession number: NR_111156) was used as the outgroup. Bootstrap values (>50%) based on 500 replicates are indicated at branching points. The GenBank accession number of each strain is listed before the name. Bar, 0.02 substitutions per site.
Figure S3
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: multimedia
Explanation note: The Non-metric multidimensional scaling (NMDS) plot showing clustering of various carbon sources based on fermentation products. Stress value, 0.012.
Table S1
This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Akshay Joshi, Vikram B. Lanjekar, Prashant K. Dhakephalkar, Tony M. Callaghan, Gareth W. Griffith, Sumit Singh Dagar
Data type: species data
Explanation note: Fermentation products of Liebetanzomyces polymorphus strain G1SC on different substrates after 5 d of incubation.